

Abstract
Projection neurons throughout the mature mammalian neocortex extend efferent axons either through the ventrolaterally positioned internal capsule to subcortical targets or through the dorsally located midline corpus callosum to the contralateral cortex. In rats, the internal capsule is pioneered on E14, but the corpus callosum is not pioneered until E17, even though these two types of projection neurons are generated at the same time. Here we use axonal markers to demonstrate that early cortical axon growth is directed toward the nascent internal capsule, which could account for the timing difference in the development of the two efferent pathways. This directed axon growth may be due to a chemoattractant and/or a chemorepellent secreted by intermediate targets of cortical efferent axons, the nascent internal capsule, or the medial wall of the dorsal telencephalon (MDT), respectively. To test for these soluble activities, explants of E15 rat neocortex and intermediate targets were cocultured in collagen gels. Cortical axon outgrowth was directed toward the internal capsule, but outgrowth was nondirected and suppressed when cocultured with MDT, suggesting that the internal capsule releases a chemoattractant for cortical axons, whereas the MDT releases a chemosuppressant. Because the chemoattractant Netrin-1 is expressed in the internal capsule, we cocultured cortical explants with E13 rat floor plate, which expresses Netrin-1, or with Netrin-1-transfected or control-transfected 293T cells. Cortical axon growth was directed toward both floor plate and Netrin-1-transfected 293T cells, as it had been toward the internal capsule, but not toward control-transfected 293T cells. These findings suggest that early events in cortical axon pathfinding may be controlled by a soluble activity which attracts initial axon growth toward the internal capsule and that this activity may be due to Netrin-1.
- axon guidance
- axon pathfinding
- chemorepellent
- chemosuppression
- cortical development
- cortical efferents
- corpus callosum
- subplate
- internal capsule
- Netrin
The directed growth of axons along their appropriate pathways is a crucial early step in the establishment of appropriate neural connectivity. Mechanisms that may influence directed axon extension have been examined in a variety of neuronal populations with well characterized trajectories, including retinal ganglion cells (Brittis et al., 1992) and circumferentially projecting neuronal populations in the hindbrain and spinal cord (Goodman, 1996). However, relatively little is known about guidance mechanisms that influence the early behavior of axons in forebrain structures, such as the neocortex. The two major efferent pathways of the neocortex are the internal capsule through the ventral forebrain and the corpus callosum across the dorsal midline. Neurons in layers 5 and 6 of cortex project through the internal capsule to the thalamus, midbrain, hindbrain, and spinal cord. A separate population of neurons in these and other layers extend axons to the contralateral cortex via the corpus callosum (O’Leary and Koester, 1993).
Even though the generation of deep layer neurons that project either subcortically or callosally is coincident (Koester and O’Leary, 1993), there is a substantial temporal disparity in the development of these two pathways. Axons from the neocortical subplate pioneer the internal capsule (McConnell et al., 1989), first extending into it on E14 in rats (De Carlos and O’Leary, 1992), whereas the corpus callosum is pioneered on E17 by neurons in cingulate cortex (Koester and O’Leary, 1994). This timing difference suggests that intracortical axon extension directed medially toward the corpus callosum is delayed relative to that directed ventrolaterally toward the internal capsule. In the first part of the study presented here, we address this issue by using axonal markers to study the early trajectories of cortical axons. We find that early axon extension is directed toward the internal capsule.
Recent studies have begun to address mechanisms that may guide cortical axons along their efferent pathways. For example, it has been proposed that an early transient population of cells in the lateral ganglionic eminence, which extends processes into the cortex at E11.5 in hamsters and E13 in mice (developmentally similar to an E15 rat), serves as a scaffold to guide cortical axons into the nascent internal capsule in a contact-mediated manner (Metin and Godement, 1996). In E17 rats, a transient projection through the internal capsule into the cortical plate, formed by cells in the perireticular nucleus, has been suggested to arrange and stabilize corticothalamic projections (Clemence and Mitrofanis, 1992; Adams and Baker, 1995).
Aside from a ventral midline-derived chemorepellent activity suggested to steer olfactory tract axons laterally (Pini, 1993), there is surprisingly little evidence of a role for soluble molecules in the establishment of axonal pathways within the forebrain, especially given the substantial body of evidence that they act to direct axon growth in the midbrain, hindbrain, and spinal cord by both attraction and repulsion (Tessier-Lavigne et al., 1988; Placzek et al., 1990;Colamarino and Tessier-Lavigne, 1995; Guthrie and Pini, 1995; Shirasaki et al., 1995, 1996; Tamada et al., 1995; Serafini et al., 1996). The early directed growth of cortical axons could be influenced by chemoattractants secreted by the nascent internal capsule or by chemorepellents secreted by a midline dorsal telencephalic structure. In the second part of the study presented here, we provide evidence from collagen gel coculture experiments that the nascent internal capsule secretes a chemoattractant activity that promotes the directed growth of cortical axons, and that the chemoattractant Netrin-1 (Kennedy et al., 1994; Serafini et al., 1994) can mimic this activity. These findings suggest a role for an internal capsule-derived chemoattractant, possibly Netrin-1, in establishing the early directed growth of cortical axons along their subcortical pathway.
MATERIALS AND METHODS
Animals. The fetuses of timed pregnant Harlan Sprague Dawley rats were used for this study. The first 24 hr period after insemination is designated embryonic day (E) 0.
Immunostaining. Immunohistochemistry using the neuron-specific antibody TuJ1 directed against acetylated β-tubulin (Moody et al., 1989), was performed on embryos ages E12–E18. E12 embryos were immersion-fixed for 4 hr in 2% paraformaldehyde and stained as whole mounts following a protocol modified from Easter et al. (1993). The embryos were permeabilized by immersion in distilled water, followed by graded ethanols, then xylenes. They were rehydrated through graded ethanols to distilled water, immersed in −20°C acetone for 10 min, then rinsed in distilled water. Aside from the acetone step, all of the steps above were 5 min each. Embryos were then incubated overnight in PBS containing 0.2% fish gelatin, 0.25% Triton X-100, 0.1 m lysine, 0.001% H2O2, and 10% normal goat serum at 4°C. After rinsing, the embryos were incubated for 48–72 hr at 4°C in TuJ1 primary antibody diluted 1:500 in PBS containing 0.25% Triton X-100 and 10% normal goat serum, washed in PBS (3 × 1 hr), and incubated for 24 hr at 4°C in horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG diluted 1:100 in PBS containing 5% rat serum. After three 1 hr washes in PBS and two half hour washes in Tris buffer (pH 8.2), the HRP was reacted in the presence of 0.05% diaminobenzidine and 0.01% H2O2 in Tris buffer. After this reaction, the cortices were removed and flat-mounted in 90% glycerol for examination and photography.
Embryos ages E14–E18 were fixed by immersion (E14) or by perfusion (E15–E18) with 4% paraformaldehyde in 0.1 m sodium phosphate buffer. After overnight post-fixation and cryoprotection in 20% sucrose, the brains were removed, embedded in OCT, and sectioned at 14 μm on a cryostat. Sections were mounted on gelatin-coated slides and stored at −20°C. For immunostaining, the slides were rinsed in distilled water and immersed in a blocking solution of 4% nonfat dry milk and 0.25% Triton X-100 in PBS (0.1 m) for 30 min. Sections were incubated overnight at 4°C in TuJ1 primary antibody diluted 1:500 in the blocking solution, rinsed in PBS, and incubated with fluorescein isothiocyanate-conjugated goat anti-mouse IgG (Fisher Biotech) diluted 1:50 in PBS containing 5% rat serum. After rinsing in PBS, slides were coverslipped in 90% glycerol in PBS containing 5% n-propyl gallate. Sections were examined and photographed using a fluorescence microscope and fluorescein optics. Control sections were incubated in the blocking solution without primary antibody and showed no staining.
DiI labeling. Embryos ages E14–E17 were fixed by immersion or by perfusion with 10% buffered formalin followed by overnight post-fixation. Small injections of 0.2% 1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine perchlorate (DiI; Molecular Probes) (Honig and Hume, 1989) in dimethylformamide (Sigma) were made using a fine-tipped glass micropipette attached to a Picospritzer (General Valve, Fairfield, NJ). A single injection was made in each cortical hemisphere; the injection site was varied across cases. Injections were made to extend through the full thickness of the cortical wall. The number of cortical hemispheres successfully injected and analyzed at each age is given in Results. Brains were stored in the dark at room temperature for at least 3 weeks to allow DiI diffusion. The brains were then embedded in agar and coronally sectioned at 100 μm on a vibratome. Every section through the cortical hemisphere was examined with a fluorescence microscope under rhodamine illumination for the presence and location of DiI-labeled axons and cells. The sections were counterstained with bisbenzimide (0.002% in sodium phosphate buffer; Sigma) and photographed under ultraviolet (bisbenzimide) and rhodamine (DiI) illumination.
DiI labeling of cortical explants was done by making small injections (as described for the labeling of cortical hemispheres described above) into the collagen gel at a distance of ∼200–300 μm. Injections were placed to the right of the cortical explant on the “toward” side close to the “test” explant, and at a similar distance on the corresponding side in the cortex-alone cultures. Injected cultures were placed in darkness for 7 d to allow retrograde diffusion to occur and were then viewed and photographed under a fluorescence microscope using a rhodamine filter.
Explant preparation. Timed pregnant rats were anesthetized using sodium pentobarbital (60 mg/kg), and then their abdomens were swabbed with alcohol and opened using sterile techniques to expose the pups. Pups were dissected from the uterine horn one at a time, washed in cold L-15 supplemented with 0.6% d-glucose (Sigma; L-15/d-glucose), then transferred to a clean dish containing L-15/d-glucose in a sterile tissue-culture hood.
Cortical explants were taken from E15 embryos. The brain was excised from the skull, and the pial membranes were removed. For preparing the cortical explants, an oval-shaped section of dorsolateral neocortex was dissected from each hemisphere and cut into approximately 500 × 500 μm pieces that spanned the full thickness of the cortical wall. A tiny slit was then made in each piece in the rostromedial corner to allow for the identification of mediolateral orientation of the piece during plating. The explants of medial dorsal telencephalon (MDT) were taken from the medial wall of the dorsal telencephalon (presumptive cingulate cortex) just dorsal to the site where the corpus callosum will later form. MDT explants were dissected from a thick coronal section and then cut into 500 × 500 μm pieces that spanned from the pial to ventricular surfaces.
For preparation of the explants of nascent internal capsule, E15 brains were embedded in 3% low melting point agar (Seaplaque, FMC Bioproducts, Rockland, ME) in L-15/d-glucose and coronally sectioned at 200 μm on a vibratome. Sections were collected in sterile L-15/d-glucose and sorted for those containing the striatum. Explants of the nascent internal capsule were dissected from the center of the presumptive striatum from sections at the level where both the lateral and medial ganglionic eminence were protruding into the lateral ventricles. One internal capsule explant was derived from each hemisphere per appropriate section (approximately three sections per brain).
Floor plate explants were derived from E13 embryos. Embryos placed in L-15/d-glucose were decapitated and eviscerated, and the skin overlying the vertebral column was removed. The vertebral column was incubated in a solution of minimal essential medium (MEM, Life Technologies) containing 0.1% trypsin (Calbiochem, La Jolla, CA) and 0.001% DNase (Boehringer-Mannheim) on ice for 30 min, then transferred to ice-cold MEM containing 1.0% fetal bovine serum (FBS; Intergen, Purchase, NY) and 0.001% DNase (Drago et al., 1991). The overlying mesoderm and ectoderm were removed, and special care was taken to remove the notochord. A cut was made along the dorsal midline of the spinal cord, and the cord was flattened, exposing the floor plate in the center. Cuts were made along the lateral edges of the floor plate to remove the lateral spinal cord. The strip of floor plate was then cut into small pieces approximately 200 μm in diameter. All explants were kept on ice in fresh L-15/d-glucose until plating.
Netrin expression construct and 293T cell transfections.293T cells (DuBridge et al., 1987) were transfected with Netrin-1 using the calcium phosphate method (Ausebel et al., 1987). The Netrin-1 expression construct consisted of a pBluescript vector (Stratagene) containing from 5′ to 3′ the CAG promoter (Niwa et al., 1991), chicken Netrin-1 cDNA (Serafini et al., 1994), an internal ribosomal entry site (Ghattas et al., 1991), green fluorescent protein (GFP) fused to the membrane-localizing signal of growth associated protein-43 (Moriyoshi et al., 1996), and a polyA tail. The control construct was the same, except the Netrin-1 cDNA was placed in the reverse (antisense) direction. The day before transfection, confluent plates were split 1:10. Eighteen to twenty-six hours later, the subconfluent cells were transfected with a solution consisting of 20–27 μg of DNA and 31.25 μl of 2 m CaCl in 500 μl of HEPES-buffered saline added to the medium of a 10 cm dish of 293T cells. Twelve hours later, the transfection solution was removed and replaced with fresh DMEM/10% FBS. Twenty-four hours later, the cells were harvested, centrifuged to a pellet, and resuspended in 100–300 μl of growth medium (see below). Effectiveness of the transfection was assessed by the presence of GFP fluorescence in the cells using fluorescein illumination. To make the agar block of cells, 2% low melting point agar in L-15/d-glucose was added to a 35 mm dish and allowed to harden. A 1 cm square was cut out and removed, and the dish was placed on a warming block. To this cavity was added 10 μl of cells and 60 μl of molten 2% agar in L-15/d-glucose, which was mixed and allowed to harden by placing the dish on ice. Approximately 300 μm square cubes were then cut from the agar and placed in L-15/d-glucose on ice until plating.
Collagen matrix assays. Collagen was prepared from young rat tails (not more than 10 cm in length). At the time of plating, 900 μl of collagen solution was mixed with 100 μl of 10× MEM (Sigma) and 11–18 μl of 7.5% bicarbonate solution (Life Technologies) and placed immediately on ice. Cultures were plated into 4-well dishes (Nunc). Initially, 25 μl of collagen was placed in the bottom of the well and allowed to gel. Explants were then placed on top of the collagen base, and an additional 75 μl of collagen was added. Before the top layer of collagen gelled, the explants were positioned 150–400 μm apart; either the medial or the lateral edge of the cortical explants faced the test explant in equal numbers. The collagen was allowed to gel at 37°C for 15–20 min, and then 500 μl of growth medium consisting of DMEM/F12 (Life Technologies), 0.1% penicillin-streptomycin, 3.6% d-glucose, 2 mmglutamine, 5% rat serum, and 10% FBS (Gemini, Bio-Products, Calabasas, CA) was added to each well. Explants were cultured for 24 hr in a humidified incubator.
Analysis of axon growth in vivo and in vitro.To quantify the direction of extension of cortical axons in TuJ1-stained whole mounts of E12 cortex, we photographed the whole mounts and then made magnified images from which we traced individual neurons and their processes. The analysis only included axons that could be traced back to the cell body to be certain of the direction of extension. To determine the direction of axon extension, a line parallel to the medial–lateral axis of the cortex was passed through the center of the cell body. Axons were assigned to either medially projecting or laterally projecting groups and, in addition, scored as falling within 45° or 90° of the line.
Cocultures were analyzed and photographed using an inverted-phase microscope (Nikon). Growth preferences exhibited by the cortical explants were scored independently by three investigators blind to the source or transfection construct of the test explants (i.e., the nascent internal capsule, MDT, floor plate, or 293T cell cubes) and the orientation of the cortical explants. In the great majority of the cultures, the scores were unanimous. In the few nonunanimous cases, two of the three investigators gave the same score, which was the one used. Growth on the sides of the cortical explant facing toward or away from the test explant (i.e., the lateral or medial side) was compared and judged to be equivalent or greater on one or the other side. Cortical explants cultured alone were plated in the same orientation as cortex cocultured with test explants so that scoring could be performed on sides equivalent to the toward (to the right) and away (to the left) sides. Given the nonparametric nature of this analysis, the Mann–Whitney test was used to analyze statistical differences between the various explant pairings. To quantitate axon outgrowth, we counted the number of cortical axon bundles in either the quadrant toward or away from the test explant that had grown more than 150 μm away from the cortical explant. This distance was selected because it was difficult to distinguish individual bundles close to the cortical explants. For each coculture type, the mean and SEM were calculated and analyzed using Student’s t test. For figure preparation, black-and-white film negatives of the cocultures were digitized on a 35 mm scanner (Nikon), and the images were composed into figures on a Macintosh computer using Photoshop (Adobe).
RESULTS
In the mature rat neocortex, separate but spatially overlapping populations of cortical neurons project axons medially or laterally. Some of the medially projecting axons pass through the corpus callosum to the contralateral cortex, and some of the laterally projecting axons extend through the internal capsule to subcortical targets; the remaining axons project intracortically (O’Leary and Koester, 1993). To analyze the development of these cortical axon trajectories, we used the neuron-specific antibody TuJ1 (Moody et al., 1989) as a general axonal marker to reveal the overall distribution of cortical axons and the fluorescent lipophilic dye DiI as an anterograde and retrograde tracer to label select groups of axons and their cells of origin.
Cortical axons extend medially or laterally at later stages of embryonic development
Injections of DiI into the neocortex on E17 (n = 9) retrogradely label, as expected, cells both medial and lateral to the injection site (Fig. 1). This labeling pattern indicates that cortical neurons positioned lateral or medial to the injection have extended axons far enough medially or laterally, respectively, to be labeled. The retrogradely labeled cells located medial to the injection site are found in both the subplate and the cortical plate, whereas those lateral to the injection site are predominantly in the cortical plate. Labeled axons deep in the intermediate zone medial to the injection site are likely to be anterogradely labeled medially projecting cortical axons, although they have yet to reach the midline, whereas those more superficially located are a mixture of retrogradely labeled cortical axons and anterogradely labeled thalamocortical axons.

Cortical axons extend laterally and medially at E17. Pattern of labeling from a neocortical DiI injection in a rat brain fixed at E17. A–C are serial coronal sections progressing from rostral (R) to caudal (C); dorsal is to the top, lateral to theleft. One section is not shown between Cand D. Unlike at earlier ages, at E17, large numbers of retrogradely labeled cells are found both lateral and medial to the injection site (asterisk). Laterally placed cells extend axons medially along the callosal trajectory. CP, Cortical plate; SP, subplate; IZ, intermediate zone; NE, neuroepithelium;LV, lateral ventricle. Scale bar, 250 μm.
The appearance of the TuJ1-immunostained sections of E17 (n = 4) and E18 (n = 4) cortex reflects the overall disposition of these axonal projection systems (Fig.2). The intermediate zone is broad and contains thick fascicles of labeled axons, including a deep collection of fascicles that appears to be oriented obliquely to those in a broader collection of fascicles directly above it. This pattern is more obvious at E18.

Developmental series of TuJ1-immunostained coronal sections from brains of rat embryos ages E14–E18. Scale bar, 250 μm. In each photomontage, medial is to the left, dorsal to the top. Stars mark the expected crossing point of the corpus callosum. PP, Preplate. Other abbreviations as in Figure 1.
Early cortical axon growth in vivo is directed toward the internal capsule
TuJ1 immunostaining of sections of E14 and E15 cortex reveals a simple labeling pattern. The preplate overlying the neuroepithelium is heavily stained, indicative of the dense accumulation of early generated cortical neurons. The staining pattern is much thicker ventrolaterally than at more dorsal and medial positions (Fig. 2), largely because of an accumulation of axons. Dorsally and medially, axon staining is difficult to distinguish from that of the preplate layer. At E16, the preplate has split into a marginal zone and a subplate by the emerging cortical plate. The medial-to-lateral gradient in the increasing thickness of axonal accumulation is very prominent. In addition, an intermediate zone containing stained axons is evident throughout the neocortex (see Fig. 6). Because the first afferent axons do not reach the cortical intermediate zone until E16 (Catalano et al., 1991; De Carlos and O’Leary, 1992; De Carlos et al., 1995), most, if not all, of the labeled axons must arise from cortical neurons. The medial-to-lateral thickening of the intracortical axon pathway at E16 and earlier revealed by TuJ1 immunostaining suggests that cortical axogenesis proceeds in a lateral-to-medial gradient. This may be attributable in part to axogenesis paralleling the neurogenetic gradient in cortex (Rickmann et al., 1977; Smart and Smart, 1982;Uylings et al., 1990; Bayer and Altman, 1991), but in addition may reflect that most early cortical axon growth is directed laterally, leading to progressively greater axonal accumulation at more lateral positions.

The earliest cortical axon growth is predominantly directed laterally. A, Whole mount of an E12 rat cortical hemisphere immunostained with the neuron-specific marker TuJ1. Rostral is to the left, lateral is down. The asterisk marks the location from whichB and C are taken. B, C, Higher magnification images of lateral regions of the brain shown inA. Some immunostained cells whose axons are visible are indicated with arrows. Most axons extend laterally toward the internal capsule. Scale bars: A, 250 μm;B, C (shown in C), 50 μm.
To examine the growth trajectories of cortical axons at these early stages, we made discrete DiI injections into the cortical wall of aldehyde-fixed brains at sites varied in position along the medial–lateral and rostral–caudal cortical axes. The pattern of DiI labeling, both retrograde and anterograde, in relation to each injection site, reveals that at E14, E15, and E16, cortical axon growth is predominantly directed toward the ventrolaterally positioned internal capsule. Figure 3 presents a summary of the axonal trajectories at these ages. This basic pattern of cortical axon convergence on the internal capsule is similar to that recently described for corticofugal axons in embryonic hamsters (Metin and Godement, 1996), although at these ages in the rat (E14–E16), the great majority of cortical axons exhibit this behavior. Examples of the actual labeling patterns are presented in Figures 4 and5, as described below.

A dorsal see-through view of a cortical hemisphere summarizing the predominant pattern of cortical axon extension at E14, E15, and E16. Medial is to the left, rostral is to thetop. The approximate locations of the internal capsule and lateral ventricle underlying the cortical mantle are indicated. Although the most medial axons must take somewhat indirect trajectories, most axon extension at these ages is directed toward the ventrolaterally located internal capsule.

DiI injections into neocortex at E14 through E16 reveal that axon extension is predominantly directed laterally toward the internal capsule. Coronal sections: dorsal to thetop, lateral to the left. A, B, Adjacent sections through an E14 fixed brain showing the pattern of labeling from a single DiI injection (asterisks). Virtually all retrogradely labeled cells are found medial to the injection site (arrow), and anterogradely labeled axons are found lateral to it (arrowheads). This labeling pattern indicates that most cells extend axons laterally at this stage. The efferent axons extend within the intermediate zone (IZ), deep to the preplate (PP). The superficial-most labeling both medial and lateral to the DiI injection site is nonspecific labeling of the pia attributable to DiI diffusion. B′, Higher power view of the cortical wall medial to the injection site in B. Thearrowhead marks nonspecific DiI labeling in the pia. A row of retrogradely labeled cortical cells is marked by thearrow. C, Low magnification image showing the pattern of labeling from an injection (asterisk) at E16. Most retrogradely labeled cells were medial to the injection (arrow). D, E, Higher magnification images of cells (arrows) retrogradely labeled by such an injection. F, Retrogradely labeled cells from a more ventrolaterally placed injection. At this level, both subplate (SP) and cortical plate (CP) cells are clearly distinguishable. Both populations extend axons laterally; very few cells of either population extend axons medially at this age. Abbreviations as in Figure 1. Scale bars: A, B (shown inA), 250 μm; B′, 100 μm;C, 250 μm; D–F (shown inD), 50 μm.

DiI injections in frontal and frontomedial cortex in E16 brains. Coronal sections: dorsal to the top, lateral to the left. A, A′, An injection (asterisk) into frontal cortex retrogradely labeled cells at more medial positions as far as 600 μm caudal in presumptive retrosplenial cortex. B, An injection (asterisk) of DiI into medial cortex retrogradely labeled cells (B′) several hundred micrometers caudal in presumptive cingulate cortex. The midline is marked by anarrowhead. LV, Lateral ventricle. Scale bar, 150 μm.
After injections that span the thickness of the neocortical wall in E14 (n = 8) and E15 (n = 8) rats, virtually all retrogradely labeled cell bodies are located medial to the injection site, and anterogradely labeled axons are directed laterally from it (E14, Fig. 4A,B,B′; E15 not shown). At E14, most of the labeling of cells and axons must be attributable to the labeling of preplate neurons because cortical plate neurons are not generated in significant numbers until E14 (Bruckner et al., 1976; Lund and Mustari, 1977; Valverde et al., 1989). After similar injections on E16 (n = 8), we also find that virtually all retrogradely labeled cells are located medial to the injection site, whereas anterogradely labeled axons are lateral to it (Fig.4C). Far medial to the injection site, the cortical plate is not easily definable, and the identity of the retrogradely labeled cells is ambiguous (Fig. 4D,E). Nearer to the injection site, cells retrogradely labeled from the more laterally placed injections are located in both the cortical plate and subplate (Fig. 4F). Only rarely are retrogradely labeled cells present in the intermediate zone lateral or medial to the injection, indicating that migrating neurons do not extend axons for any significant distance. A similar pattern of labeling was observed after injections in frontal or frontomedial cortex on E16 (n= 6). Virtually all retrogradely labeled cells are located medial and caudomedial to the injection site, and anterogradely labeled axons extend lateral and rostrolateral from it (Fig. 5). Cells in medial cortex extend their axons rostrolaterally, some around the front of the lateral ventricle to reach the internal capsule. In conclusion, at E14, E15, and E16, cortical axon extension is predominantly directed toward the internal capsule, the pathway from cortex to subcortical targets.
Initial extension of the first cortical axons is predominantly directed laterally
At the earliest ages of cortical axon growth, the axons are too short to be revealed effectively with DiI injections. However, because the density of neurons and axons is low at this stage, unlike at later ages, TuJ1 can be used to examine the trajectories of individual axons. TuJ1 was used to immunolabel whole mounts of cortical hemispheres from E12 rat fetuses (n = 4), the age at which the first cortical neurons become postmitotic (Valverde et al., 1989). TuJ1 staining reveals a low density of preplate neurons, which in the most lateral region of the cortex have begun to extend axons. Most of these axons are directed laterally (Fig.6A), toward the future internal capsule. Many of the axons are directed laterally from their initiation point on the neuronal soma, whereas others initially extend obliquely, then turn to assume a laterally directed trajectory (Fig.6B,C). Quantification of the direction of cortical axon extension shows that 92% of axons with a definable trajectory (n = 97) are directed laterally, and 91% of these axons have a trajectory within 45° of a line that extends directly lateral (for details, see Materials and Methods). Thus, from the onset of cortical axogenesis, cortical axon growth is predominantly directed laterally toward the site of the future internal capsule, and remains so over the first 5 d of cortical axon extension (E12–E16).
A soluble signal released by the internal capsule attracts cortical axons
A priori, the directed growth of cortical axons toward the ventrolaterally located internal capsule could be promoted either by molecules expressed within cortex or soluble molecules that diffuse into it. To assess the latter possibility, we carried out coculture experiments in three-dimensional collagen gels using intermediate targets of cortical axons (Fig. 7). Specifically, we tested whether the MDT releases a soluble activity that causes cortical axons to grow away from the midline and therefore toward the internal capsule, or whether the nascent internal capsule may release a soluble activity that attracts cortical axons toward it (Fig. 7). Explants were taken from E15 rats, an age when cortical axons show a strong directional growth toward the internal capsule in vivo. For each culture, cortical axon growth was scored in two ways: by a blind scoring of axon growth preferences and by counting axon fascicles (for details, see Materials and Methods).
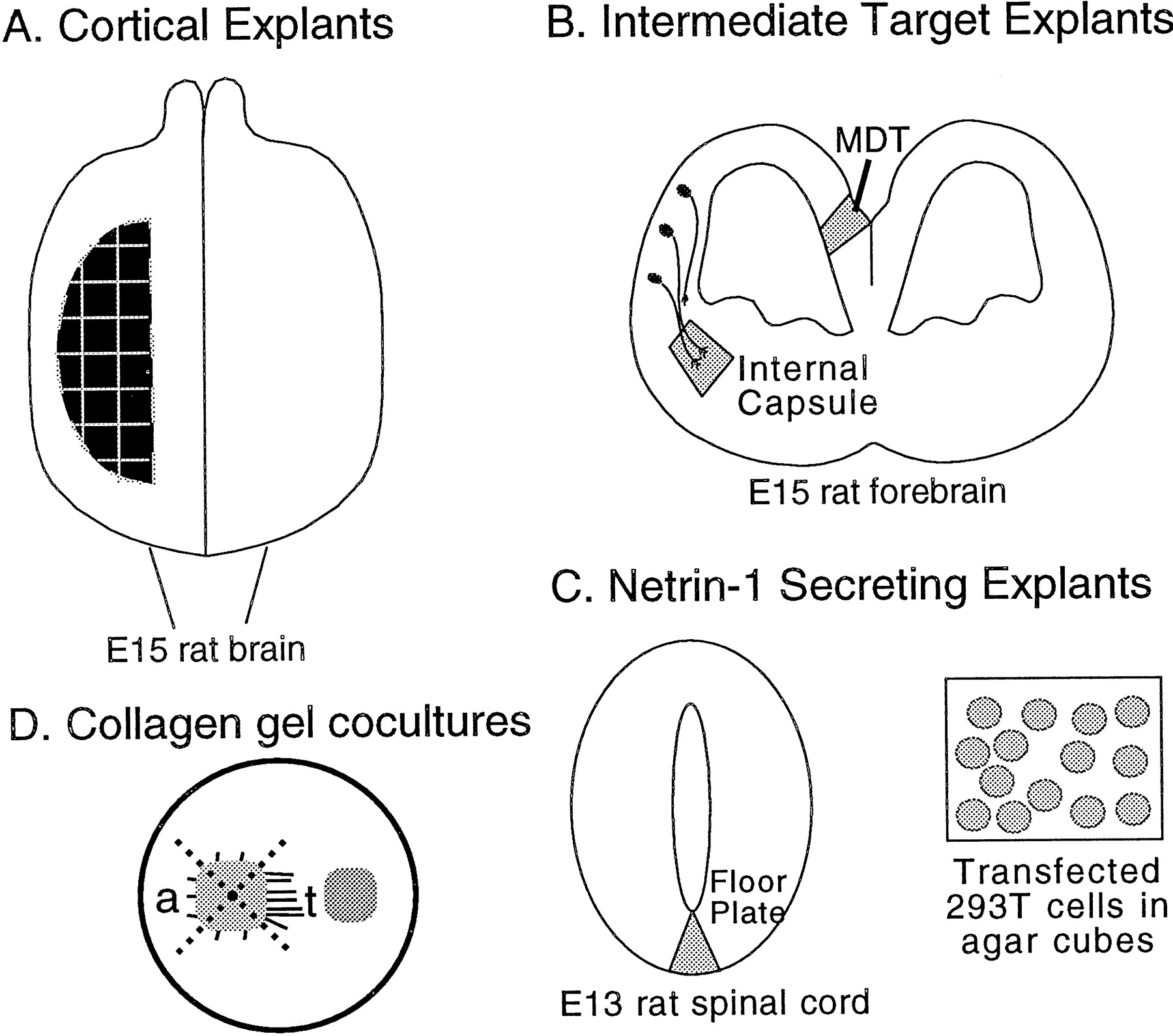
Schematic representation of coculture experiments. A, Dorsal view of an E15 rat brain showing where cortical explants were derived from within the dorsal telencephalon (i.e., neocortex). B, Coronal section through an E15 rat forebrain showing the locations from which explants of the intermediate target explants of cortical axons, the nascent internal capsule and the MDT, were dissected.C, Netrin-1-secreting explants were the floor plate, taken from an E13 rat spinal cord shown in transverse section, and agar-embedded HEK 293T cells transfected with Netrin-1 cDNA in the sense orientation. Agar-embedded HEK 293T cells transfected with Netrin-1 cDNA in the antisense orientation were used as a control.D, Cortical explants were cultured in collagen gels either alone or with test explants of either internal capsule, MDT, floor plate, or small agar cubes containing transfected 293T cells. For analysis of axon outgrowth, cortical explants were divided into four quadrants as shown by the dotted lines. The axon outgrowth from the sides of the cortical explant toward (t) and away (a) from the test explants was analyzed.
When cortical explants were cultured alone in collagen gels (n = 12), axon outgrowth was either symmetrical or more profuse on the caudal or lateral side of the explant (Table1; Fig. 8A). When cortex was cocultured with explants of MDT (n = 21), cortical axons maintained the characteristic pattern of axon outgrowth observed when they were cultured alone (Table 1; Fig.8B). Thus, this experiment provides no evidence that MDT releases a chemorepellent activity for early cortical axons. However, cortical explants cocultured with nascent internal capsule (n = 22) exhibited greater axon outgrowth on the side of the explant facing the internal capsule (Table 1; Fig.8C). A statistical analysis using the Mann–Whitney test indicates that the growth response of cortical axons toward explants of internal capsule was significantly different from the growth response of cortex cultured alone (p ≤ 0.05) or cocultured with explants of MDT (p ≤ 0.01) (Table 1). Counts of axon fascicles support the blind qualitative assessments of cortical explant growth preferences (Fig.9A). The number of axon fascicles extending from the side of cortical explants facing the internal capsule was more than threefold greater than the number extending from the opposite side; this difference is statistically significant (p ≤ 0.05, Student’s paired ttest). No significant difference was found in the number of axon bundles extending from the two sides of cortical explants in cultures of cortex alone or in cocultures with MDT. These findings demonstrate that, in vitro, the nascent internal capsule releases a soluble activity that promotes the growth of cortical axons toward it.
Summary of collagen gel coculture experiments

The internal capsule releases a chemoattractant activity for early cortical axons that can be mimicked by a soluble activity released by floor plate and by Netrin-1-transfected 293T cells. Cortical explants (ctx) were cultured in collagen gels either alone (A) or with test explants of MDT (B; MDT), nascent internal capsule (C; ic), floor plate (D;fp), or 293T cells transfected with Netrin-1 cDNA (E; Netrin-1 293T), or 293T cells transfected with a control construct (F; cont. 293T). The cortical explants were placed with their pial surface up, and either their medial or lateral side facing the test explant. A, E15 rat cortical explants cultured alone extended axons in a nondirected manner. B, Similarly, cortical explants cocultured with explants of E15 rat MDT extended axons in a nondirected manner. C, In contrast, cortical explants cocultured with explants of E15 rat internal capsule (ic) showed robust axon outgrowth directed toward the internal capsule explant. D–F, The effect of the internal capsule could be mimicked by coculturing the cortical explants with either floor plate explants (D) or agar cubes containing 293T cells transfected with Netrin-1 (E). In these cocultures, growth toward the floor plate or Netrin-1-transfected 293T cells was robust. In contrast, cortical explants cocultured with agar cubes containing control-transfected 293T cells extended axons in a nondirected manner (F). Scale bar, 200 μm.

Quantification of axon bundles in each type of cortical explant culture. A, Mean number (±SEM) of axon bundles extended from the cortical explants (ctx) toward (▪) or away (░) from test explants. ic, Internal capsule; fp, floor plate; N1–293T, agar cubes of 293T cells transfected with Netrin-1 cDNA; cont 293T, agar cubes of 293T cells transfected with Netrin-1 cDNA in the antisense orientation. The perimeter surrounding each cortical explant was divided into four quadrants (see Fig. 7), and the total number of axon bundles >150 μm in length, found in either the toward quadrant or the away quadrant, was counted. For cortical explants cultured alone (ctx alone), the toward quadrant was to the right and the away quadrant to theleft. B, Mean total number (±SEM) of axon bundles extended both toward and away from the test explants (i.e., sum of the data presented in A). The number of cases scored is the same as the total number of explants indicated in Table 1. These parametric data were statistically analyzed using Student’s t test.
The floor plate and Netrin-1 attract cortical axons
Several lines of evidence suggest that Netrin-1 (Kennedy et al., 1994; Serafini et al., 1994) can act as a soluble molecule to promote the ventrally directed growth of circumferentially projecting axons in the midbrain, hindbrain, and spinal cord (Tessier-Lavigne et al., 1988;Placzek et al., 1990; Shirasaki et al., 1995, 1996; Tamada et al., 1995; Serafini et al., 1996). Because Netrin-1 is expressed in the internal capsule as cortical axons begin to extend toward it (Serafini et al., 1996) and has properties consistent with those of a chemoattractant (Kennedy et al., 1994; Serafini et al., 1994), we have carried out coculture experiments to assess whether Netrin-1 may be involved in the ventrolaterally directed growth of cortical axons toward the nascent internal capsule. In the first set of experiments, floor plate, which expresses Netrin-1 (Kennedy et al., 1994), was isolated from E13 rat spinal cord and cocultured with cortical explants in collagen gels (n = 16; Fig. 7). In these cocultures, cortical axon growth was directed toward the floor plate (Fig.8D). When compared with cultures of cortex alone, or cocultures of cortex with MDT, the directed growth of cortical axons toward the floor plate was significant (p ≤ 0.025 and p ≤ 0.01, respectively; Mann–Whitney test; Table 1). However, there was no significant difference in the directed axon growth exhibited by cortical explants cocultured with the nascent internal capsule and those cocultured with floor plate, suggesting a qualitatively similar response (Table 1). The number of axon fascicles extending from the side of cortical explants facing the floor plate is more than twofold greater than the number extending from the opposite side (Fig. 9A); this difference is statistically significant (p < 0.01, Student’s paired ttest).
A second set of experiments was done to address whether the response of cortical axons to the floor plate may be mediated by Netrin-1. Small cubes of agar-containing HEK 293T cells transfected with a Netrin-1 expression construct were cocultured with cortical explants (n = 33; Fig. 7). As in cocultures with nascent internal capsule or floor plate, cortical explants have greater axon outgrowth from the side facing the cubes of Netrin-1-expressing cells (Fig. 8E). In contrast, cortical explants cocultured with cubes of 293T cells transfected with a control construct (n = 33) exhibited nondirected growth that was not statistically different from cortex cultured alone (Table 1; Fig.8F). When compared with cultures of cortex alone, or cortex cocultured with control-transfected 293T cells, cortical axon growth directed toward the Netrin-1-transfected 293T cells is significant (p < 0.05 and p < 0.05, respectively; Mann–Whitney test; Table 1). Again, counts of axon fascicles support the blind qualitative assessments of cortical explant growth preferences (Fig. 9A). The number of axon fascicles extending from the side of cortical explants facing the Netrin-1-transfected 293T cells is more than twofold greater than the number extending from the opposite side; this difference is statistically significant (p < 0.01, Student’s paired t test). No significant difference was observed in the number of axon bundles extending from the two sides of cortical explants cocultured with 293T cells transfected with a control construct. Because the response of cortical axons to the internal capsule can be mimicked by both the floor plate and Netrin-1-transfected 293T cells, Netrin-1 may be the molecule secreted by the internal capsule that attracts cortical axons in vitro.
Evidence that increased axon growth toward test explants may be attributable to chemoattraction
As described above, we found a significant difference in the number of axon bundles extended by cortical explants from their toward side, facing test explants, compared with their away side in cocultures with internal capsule or Netrin-1-transfected 293T cells, but not in control cultures of cortex cultured alone or with control-transfected 293T cells. In contrast, the total number of axon bundles extended from the two sides in cocultures of cortex with internal capsule or Netrin-1-transfected 293T cells did not differ significantly from the two types of control cultures (Fig. 9B; Student’s unpaired t test). Thus, these explants extend similar total numbers of axon bundles, which are not increased by coculturing in the presence of the internal capsule or Netrin-1-transfected 293T cells. Therefore, the differences between toward and away sides in the number of axon bundles extended by cortical explants cocultured with either internal capsule or Netrin-1-transfected 293T cells is attributable to an increase in the proportion of axon bundles directed toward these test explants, out of a constant total number of axon bundles. This finding is consistent with a chemoattractive mechanism promoting cortical axon growth toward these test explants.
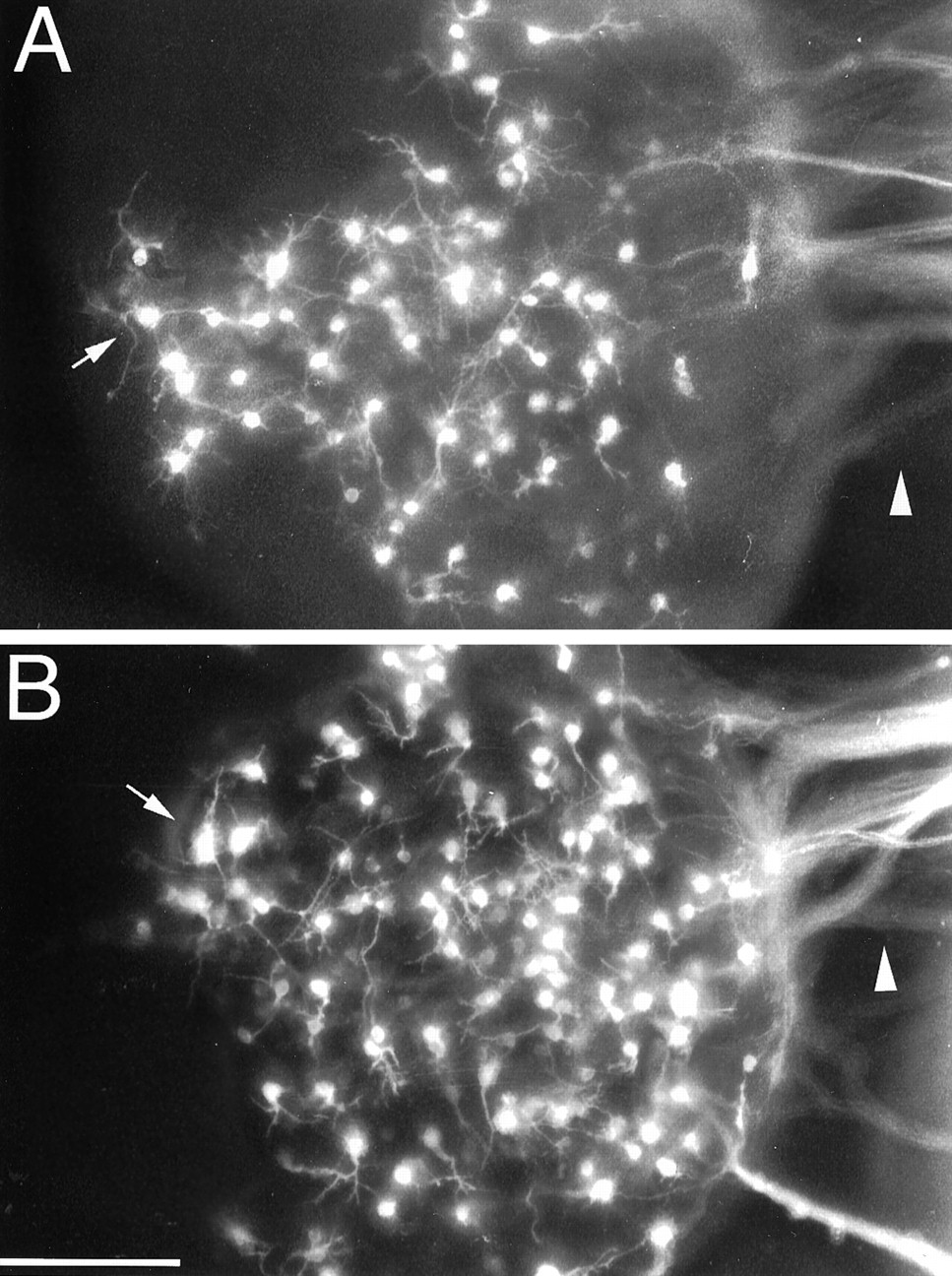
The increase in the number of axonal processes on the toward side of cortical explants cocultured with internal capsule, floor plate, or Netrin-1-transfected 293T cells could possibly be attributed to a trophic effect of the test explant that is highly localized to the side of the cortical explant facing it (e.g., a strong bias for survival or axon extension by neurons on the side closest to the test explant). To further investigate this possibility, we retrogradely labeled neurons within the cortical explants by injecting DiI into the collagen gel between the cortical and test explants. Figure 10 shows our typical findings, using as examples an explant of cortex cultured alone (Fig. 10A) and one cocultured with floor plate (Fig. 10B). In both cultures, retrogradely labeled neurons are distributed over the entire explant. This finding suggests that the increased cortical axon outgrowth exhibited on the toward side versus the away side in the cocultures is not attributable to a localized trophic effect.

Cortical axons extended toward test explants arise from neurons distributed throughout the cortical explants. DiI injected into the collagen gel to the right side (i.e., toward side) of the cortical explants retrogradely labels neurons distributed across cortical explants cultured alone (A) or cocultured with floor plate (B).Arrowheads point to bundles of retrogradely labeled cortical axons extended into the collagen gel. Arrowsmark retrogradely labeled neurons at the edge of the side of the cortical explants opposite the injection site (i.e., the away side). InB, the injection was placed between the cortical explant and the floor plate. In each case, only a proportion of the axons extending into the collagen contact the DiI injection site. Scale bar, 200 μm.
Interestingly, we found that the total number of axon bundles extended by cortical explants cocultured with floor plate was approximately twofold greater than the number extended by cortex cultured alone or cocultured with internal capsule or Netrin-1-transfected 293T cells (the difference is significant, p < 0.01, Student’s unpaired t test; Fig. 9B). Thus, floor plate appears to release a soluble activity that enhances cortical axon outgrowth. The twofold increase in total cortical axon bundles reflects an increase in the number of axon bundles extending from each of the two sides of the cortical explants, toward and away from the floor plate, compared with the baseline condition of cocultures of cortical explants with internal capsule or Netrin-1-transfected cells (Fig.9A). This observation suggests that the apparent trophic effect of the floor plate is not localized to the side of the cortical explant nearest it, and as described above seems to uniformly influence the entire cortical explant. Given this generalized chemotrophic effect, it is reasonable to suggest that the trophic effect is superimposed on the chemoattractive effect discussed above.
As described above, we found no evidence that explants of MDT exert a chemorepellent effect on cortical axons. However, we do find an approximately 50% decrease in the total number of axon bundles in cortical explants cocultured with MDT compared with cortex cultured alone (p < 0.02, Student’s unpairedt test; Fig. 9B). As for the generalized trophic effect of the floor plate, this opposite effect of MDT appears to have a similar effect on axon growth from both sides of the cortical explant (Fig. 9A). Thus, MDT appears to release a soluble activity that decreases cortical axon outgrowth, a phenomenon termed chemosuppression (Wang et al., 1996).
DISCUSSION
The mechanism of chemotropism, or the action at a distance by diffusible signals on the directional growth of axons, has recently gained prominence through the demonstration of physiological roles for such a mechanism (Tessier-Lavigne and Placzek, 1991; Serafini et al., 1996) and the cloning of the floor plate-derived chemoattractant Netrin-1 (Kennedy et al., 1994; Serafini et al., 1994). Previousin vitro studies have suggested that a target-derived chemoattractant activity can provide a directional cue for the growth of cortical layer 5 axons in vitro (Heffner et al., 1990;Joosten et al., 1994; Sato et al., 1994), and because the activity can promote the collateral branching of cortical axons (Sato et al., 1994), it has been implicated in the process of target selection by layer 5 efferent axons. Evidence presented here suggests that at a much earlier stage in the development of cortical efferent projections, a chemoattractant released by an intermediate target of cortical efferent axons, the internal capsule, may influence the directed growth of cortical efferent axons out of the cortex into their subcortical pathway through the ventral forebrain.
The finding that Netrin-1 is expressed in the mouse striatum/internal capsule at early stages of cortical axon extension [Serafini et al. (1996), their Fig. 9] suggests that it may have a role in directing early cortical axon growth toward the internal capsule en route to more caudally located targets. By examining cocultures of cortical explants with floor plate, a tissue that also expresses Netrin-1 (Kennedy et al., 1994), or with Netrin-1-transfected 293T cells, we have found activities similar to those observed in cocultures using the internal capsule. Similar results have been obtained by C. Metin and D. Deleglise (personal communications). These findings suggest that Netrin-1 may be involved in directing cortical axon growth toward and/or through the internal capsule.
We have not been able to provide evidence that the internal capsule-derived activity or Netrin-1 has a tropic effect on cortical axons (as defined by a reorientation of axons toward the source). We attempted to do this by retrogradely labeling cortical axons with DiI injections into the collagen gel, but the resolution of axon labeling within the cortical explants was inadequate to perform a definitive analysis on axonal trajectories. However, the widespread distribution of the retrogradely labeled cells across the cortical explants suggests that the enhanced outgrowth of cortical axons toward the internal capsule, floor plate, or Netrin-1-transfected cells is not attributable to a trophic effect localized to the toward side of the cortical explant. In addition, our findings show that although neither the internal capsule nor the Netrin-1-transfected 293T cells increase the total number of axon bundles extended by cortical explants, both promote a significant increase in the number of axon bundles that extend specifically from the side of the cortical explant toward the test explant. Taken together, these results suggest that the internal capsule-derived activity and Netrin-1 have chemoattractive effects on cortical axons. In this discussion, for simplicity we will consider that Netrin-1 is indeed the internal capsule-derived chemoattractant activity, but the same considerations would apply to the internal capsule-derived activity whether it is attributable entirely or in part to Netrin-1 or to an unidentified agent.
In principal, an internal capsule-derived activity could act at a distance or, alternatively, only locally. For example, Netrin-1 may be secreted by cells in the internal capsule and then diffuse through the overlying cortex, thereby producing a graded distribution of the molecule that could direct cortical axon growth from the outset. Consistent with this possibility, the trajectories of early cortical axons approximate the shortest path to the internal capsule; a path that in principal should parallel the steepest gradient of a diffusible signal emanating from the internal capsule. For example, neurons in the dorsolateral cortex send their axons directly laterally and ventrally, whereas neurons in the rostromedial cortex send their axons around the rostral end of the lateral ventricle, rather than over the dorsal cortical surface (see Fig. 3).
Alternatively, Netrin-1 may diffuse only a short distance in vivo, if at all, and may act to maintain directed axon growth after initial axon extension has begun. The fact that Netrin-1 has a signal sequence suggests that it is secreted (Serafini et al., 1994), and in vitro evidence presented here and elsewhere (Kennedy et al., 1994) demonstrates that Netrin-1 can act in a soluble manner. However, Netrin-1 can be tightly associated with cellular membranes because during its purification and isolation, Netrin-1 activity was identified in salt extracts of the membrane fraction prepared from E13 chicken brain rather than in the soluble fraction (Serafini et al., 1994), and in Netrin-1-transfected COS cells most of the activity was associated with high salt extracts of COS cell membranes rather than in COS cell conditioned medium (Kennedy et al., 1994). This evidence could indicate that, in vivo, Netrin-1 produced by cells within the internal capsule may remain in this region and affect axon growth locally rather than at a distance. In this scheme, other agents may establish the initially directed extension of cortical axons toward the internal capsule, and Netrin-1 may serve to promote the turning of cortical axons into the internal capsule and/or to maintain their directed growth through it, by directly guiding the axons or by creating a favorable environment for their extension.
If the diffusion of Netrin-1 is limited, gradients of other potential guidance cues, for example, cell adhesion molecules (CAMs) or extracellular matrix molecules (ECMs), may direct the initial growth of cortical axons toward the internal capsule. A decreasing peripheral-to-central maturational gradient of the ECM, chondroitin-sulfate proteoglycan (CSPG), has been implicated in directing retinal ganglion cell axons toward the optic disk (Brittis et al., 1992). In this instance, axons grow “down” a CSPG gradient, which parallels the central-to-peripheral maturational gradient in the retina. Although this is an attractive idea that merits further consideration, there are no reports of CAMs or ECMs distributed in gradients that parallel the directed growth of early cortical axons (Fushiki and Schachner, 1986; McKenna and Raper, 1988; Sheppard et al., 1991; Bicknese et al., 1994). In addition, unlike retina, the trajectories of many cortical axons to the internal capsule do not seem to parallel the maturational gradient, which in cortex proceeds from rostrolateral to caudomedial (Bayer and Altman, 1991).
In addition to acting within the forebrain, Netrin-1 may also act in the guidance of cortical efferent axons farther along their subcortical pathway. For example, after cortical layer 5 axons pass through the internal capsule, they deviate toward the midline in the caudal diencephalon and midbrain, and then extend caudally through the hindbrain in a pathway adjacent to the ventral midline, apposed to the floor plate. Thus, Netrin-1 (or another floor plate-derived molecule) may be involved in guiding corticospinal axons along their spinally directed pathway. However, at the junction between the hindbrain and the spinal cord, corticospinal axons turn dorsally, cross the midline, and project caudally down the spinal cord in a dorsally located midline tract. Therefore, additional molecular mechanisms must contribute to defining the corticospinal axon pathway.
Given that the neocortex has two major efferent pathways, through the internal capsule to subcortical targets and through the corpus callosum to the contralateral cortex, it was surprising to find that early cortical axon extension is predominantly directed along a trajectory toward the internal capsule. This directed axonal growth is observed from E12, the earliest time of axon extension in cortex, to E16. Only later, beginning at E17, do we observe substantial numbers of cortical cells extending axons medially toward the midline corpus callosum. This apparent delay in cortical axon extension toward the midline cannot be explained by a later generation of neurons with medially directed axons compared to those with ventrolaterally directed axons. The earliest generated cortical neurons are subplate neurons, which are also the population that pioneers the internal capsule (McConnell et al., 1989; De Carlos and O’Leary, 1992). Although subplate neurons rarely extend axons across the corpus callosum, a substantial proportion of them do extend axons medially at later ages (Koester and O’Leary, 1994). Furthermore, at E15 and E16, cortical axon growth is still predominantly directed toward the internal capsule at a time when axons have begun to be extended by cortical plate neurons that will form the deep cortical layers, which contain callosally projecting and subcortically projecting neurons in approximately equal proportions and are generated at the same time (Koester and O’Leary, 1993).
This leaves three potential explanations for the delay in the extension of cortical axons toward the midline: first, cells that will eventually extend axons medially, initially extend axons laterally, and only later direct axons toward the midline. However, we have found no evidence from present or previous (Koester and O’Leary, 1993, 1994) axonal labeling studies to suggest that this is a major phenomenon. Thus, the delay is more likely attributable to one of two other scenarios. First, neurons that will project medially initiate axons, but their axons either grow very slowly, or stall and wait in a deeper layer (e.g., subplate or intermediate zone) before extending medially for a distance detectable by the techniques used here. Second, neurons that will project medially do not initiate axogenesis until many days after their laterally projecting counterparts with which they are cogenerated.
The lag in medially directed axon growth could be attributable to the late development of an attractant signal, either soluble or membrane-associated, that directs axon growth toward the midline. Another potential cause for the observed delay in medially directed axon extension is that a repellent or inhibitor of axon growth could be transiently expressed. The coculture experiments presented here revealed no evidence that the medial dorsal telencephalon, a midline cortical structure, secretes a chemorepellent activity for early cortical axons. However, we did find evidence that the medial dorsal telencephalon releases a soluble activity that decreases or inhibits cortical axon extension. Thus, this chemosuppressive activity could contribute to the delay in cortical axon extension toward the midline. In addition, repellents or inhibitors could develop locally within the cortex and specifically inhibit the extension of the medially projecting axons. For example, collapsin/Sema III (Kolodkin et al., 1993; Luo et al., 1993), a protein that has been shown to repel or inhibit axon growth (Luo et al., 1993), is expressed in developing rodent cortex (Messersmith et al., 1994) and could operate in this manner. The molecular basis of the early directed growth of cortical axons toward the internal capsule and the delay in medially directed axon extension toward the midline must be due to multiple cues expressed in temporally and spatially defined sequences. Such complexity is necessary to account for the multiple directions of axon growth by specific subsets of cortical neurons in the same region of cortex during overlapping periods of development.
Footnotes
This work was supported by National Institutes of Health Grant NS31558 (D.D.M.O.). L.J.R. is a Lucille P. Markey Fellow and was supported in part by a grant from the Lucille P. Markey Charitable Trust. We thank A. Frankfurter for providing the TuJ1 antibody, M. Tessier-Lavigne for Netrin-1 cDNA, J. Majors for the internal ribosomal entry site cDNA, C. Lucidi-Phillipi for preparing some of the TuJ1-immunostained material, S. Boutsaboualoy for technical assistance, G. Goodhill for blind scoring of growth preferences and help with statistical analyses, and D. Borngasser for help with figure preparation.
Correspondence should be addressed to Dennis D. M. O’Leary, MNL-O, The Salk Institute, 10010 North Torrey Pines Road, La Jolla, CA 92037.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}