

Abstract
Neurturin (NRTN) and glial cell line-derived neurotrophic factor (GDNF) are members of a family of trophic factors with similar actionsin vitro on certain neuronal classes. Retrograde transport of GDNF and NRTN was compared in peripheral sensory, sympathetic, and motor neurons to determine whether in vivo these factors are transported selectively by different neuronal populations. After sciatic nerve injections, NRTN was transported by sensory neurons of the dorsal root ganglion (DRG). Competition studies demonstrated only limited cross-competition between NRTN and GDNF, indicating selective receptor-mediated transport of these factors. By using immunohistochemistry, we identified two populations of NRTN-transporting DRG neurons: a major population of small, RET-positive, IB4-positive, non-TrkA-expressing neurons that also show the ability to transport GDNF and a minor population of calretinin-expressing neurons that fail to transport GDNF. Spinal motor neurons in the adult showed relatively less ability to transport NRTN than to transport GDNF, although NRTN prevented the cell death of neonatal motor neurons in a manner very similar to GDNF (Yan et al., 1995) and persephin (PSPN) (Milbrandt et al., 1998). Last, NRTN, like GDNF, was not transported to sympathetic neurons of the adult superior cervical ganglion (SCG) after injection into the anterior eye chamber. These data reveal a high degree of functional selectivity of GDNF family receptor-α (GFRα) coreceptor subtypes for NRTN and GDNFin vivo.
Glial cell line-derived neurotrophic factor (GDNF) originally was identified as a potent survival mediator for mesencephalic dopaminergic neurons in culture (Lin et al., 1993). Three years after the identification of GDNF, a second member of the GDNF family, neurturin (NRTN), was discovered (Kotzbauer et al., 1996). GDNF and NRTN promote the in vitro survival of numerous peripheral neuronal types, including nodose, superior cervical ganglion (SCG), dorsal root ganglion (DRG), and trigeminal ganglion neurons (Buj-Bello et al., 1995; Kotzbauer et al., 1996). The third member of the GDNF family, persephin (PSPN), which has been identified on the basis of sequence homology, has no effect on the peripheral neuronal populations tested in vitro, in contrast to NRTN and GDNF. Yet like NRTN and GDNF (Lin et al., 1993; Henderson et al., 1994;Horger et al., 1998), PSPN can promote the survival of motor neurons and midbrain dopaminergic neurons in vitro and lesioned dopaminergic neurons in the adult animal (Milbrandt et al., 1998). Although the in vitro properties of NRTN have been described (Kotzbauer et al., 1996; Milbrandt et al., 1998), less is known about its in vivo properties. Likewise, although NRTN is a powerful survival factor for developing tissues, its role in the adult organism has been ignored.
The first GDNF family receptor component to be described was the orphan tyrosine kinase receptor RET (Durbec et al., 1996; Trupp et al., 1996). The striking similarity between RET and GDNF knock-out mice led a number of groups to examine potential interactions between GDNF and RET (Schuchardt et al., 1994; Pichel et al., 1996; Sanchez et al., 1996). These studies demonstrated the ability of GDNF to induce tyrosine phosphorylation of the RET receptor. Coincident with the identification of RET, two other groups, using expression-cloning strategies, identified a novel protein with high affinity for GDNF (Jing et al., 1996; Treanor et al., 1996). Both groups have shown that this novel component, GDNF family receptor-α1 (GFRα1), is a GPI-linked protein that acts in concert with RET to transduce the GDNF signal. After the identification of GFRα1, a homologous GPI-linked protein named GFRα2 was identified (Baloh et al., 1997; Klein et al., 1997;Sanicola et al., 1997; Suvanto et al., 1997). Subsequently, additional members of this family have been reported. GFRα3 has a sequence distinct from GFRα1 and GFRα2 and no apparent ability to bind GDNF, NRTN, or PSPN (Jing et al., 1997; Baloh et al., 1998; Naveilhan et al., 1998; Worby et al., 1998). An avian protein named GFRα4 (similar in sequence to GFRα1 and GFRα2) has been identified in the chicken (Thompson et al., 1998), and a recent report suggests that GFRα4 may be the PSPN coreceptor (Enokido et al., 1998).
This study examined the transport of neurturin by adult neuronal populations known to respond to NRTN during development. Competition studies described here demonstrate the selectivity of retrograde transport-mediating receptor sites for NRTN, GDNF, and PSPN. Using retrograde transport in combination with immunohistochemistry, we have identified specific NRTN-responsive populations within the adult DRG. Last, a potent protective effect for neurturin on axotomized motor neurons is demonstrated.
MATERIALS AND METHODS
Reagents, unless stated otherwise, were purchased from Sigma (St. Louis, MO).
Preparation of 125I-labeled GDNF, NRTN, and PSPN. GDNF, NRTN, or PSPN was iodinated to a specific activity (0.6–2 ×107 cpm/μg) with Na-125I and lactoperoxidase by using the methods ofMarchalonis (1969). Radioactive iodine incorporation into the respective proteins (PSPN, GDNF, NRTN) was 5–20%. The reactions, performed at room temperature, used the following quantities: 1–10 μg of protein in 36 μl of 0.2 mNaPO4 buffer, pH 6.0, 3 μl of lactoperoxidase (200–300 μg/ml), 3–5 μl of Na-125I (1 mci/10λ; Amersham, Arlington Heights, IL), and 1 μl of a 1:103 dilution of H2O2 (30%) in a 0.1m NaPO4 buffer, pH 6.0, for a total reaction volume of 50 μl. The reaction was terminated after 10–15 min with the addition of 150 μl of a 0.1 mNaPO4 buffer containing 420 mm NaCl and 0.1 m Na-125I, pH 7.5. Radiolabeled NRTN and GDNF were tested for bioactivity in the SCG survival assay; levels of survival promotion by labeled proteins were comparable to those levels of survival mediated by nonlabeled proteins.
Retrograde transport of 125I-labeled ligands. Adult male Sprague Dawley (Harlan, Indianapolis, IN) rats (250–300 gm) were anesthetized. The sciatic nerve was exposed, and firm pressure was applied to the nerve for 30 sec to deliver a crush. An amount of 1–5 μl (25–100 ng) of radiolabeled protein plus vehicle or radiolabeled protein plus cold competitor (within an experiment the injections were matched for volume) was injected directly into the nerve at the crush site, and 14–18 hr later (14–16 hr for maximal transport into DRG and 18 hr for maximal transport to motor neurons) the animals were perfused transcardially with PBS, followed by 4% paraformaldehyde. Ipsilateral and contralateral L3–L6 DRG as well as lumbar spinal cord segments were removed and counted. Autoradiography was performed on ipsilateral L4 and L5 DRG as well as on spinal cord cross sections. Tissues were dehydrated and paraffin-embedded; serial sections (10 μm) were coated with Kodak NTB-2 emulsion and exposed for 4–5 weeks at 4°C before being developed. Ipsilateral L4–L5 DRG were taken from parallel experiments after paraformaldehyde fixation and sucrose immersion and were used for immunocytochemistry (see below). Identical autoradiographic procedures were performed after antibody treatments.
Immunocytochemistry. Frozen sections of DRG (10 μm each) were dried and incubated in blocking buffer containing 1.5% normal goat serum, 1% porcine gelatin, and 0.2% Triton X-100 in Superblock buffer (Pierce, Rockford, IL). The RET antibody (Molliver et al., 1997) was used at 1 μg/ml; α-internexin (Chemicon, Temecula, CA) was used at 1:400; rabbit anti-TrkA antibody, kindly provided by Dr. Louis Reichardt (University of California, San Francisco), was used at a dilution of 1:10,000. Calretinin (Chemicon) and IB-4 conjugated to HRP were used at 10 μg/ml. Primary and secondary antibodies were diluted 1:1 with Superblock buffer (Pierce)/1.5% normal serum (Vector, Burlingame, CA). Sections were incubated in primary antibody overnight, washed three times in PBS, and placed in secondary antibody for 30 min; then the sections were visualized by using the Vectastain peroxidase substrate kit (Vector, Burlingame, CA). Control sections were processed without a primary antibody. Comparing antibody staining and autoradiographic labeling, we calculated and then confirmed the population percentages by comparison with counts generated by a second blinded observer.
Size/frequency analysis. With camera lucida the neuronal profiles containing nuclei and labeled with silver grains or a RET antibody or both were traced under bright-field or dark-field conditions at 200× magnification; micrometer tracings were taken at the same time. Retrogradely labeled neurons were identified on the basis of the presence of a twofold or greater density of silver grains (as compared with background) clustered over neuronal somata. Camera lucida drawings were scanned, and somal areas were quantified with SigmaScanPro4 (SSPS, Chicago, IL). Size/frequency histograms were generated by using SigmaPlot; they represent all RET-stained and NRTN- or GDNF-labeled neurons from at least five DRG slices taken from five different animals (RET, n = 11; NRTN, n= 6; GDNF, n = 5).
For size analysis in the competition experiment, the paraffin-imbedded ganglia were used. After being developed, the sections were stained with crystal violet (EM Diagnostic Systems, Gibbstown, NJ), and somal areas were measured with camera lucida as described above. Paraffin immersion resulted in some cell shrinkage, so cell-size data from frozen preparations could not be pooled with paraffin data. Competition data were assessed for L4 and/or L5 ganglia from four different animals.
Sciatic axotomy. The sciatic nerve was transected unilaterally in anesthetized postnatal day 2 (P2) rats (Sprague Dawley, Harlan). Approximately 2 μl of bovine serum albumin (BSA; in controls) or the neurotrophic factor tested was applied onto the proximal nerve stump by means of a small, closed tube to keep the solution on the nerve tip. To label the sciatic motor neuron pool in a highly specific manner, we simultaneously added fluorogold, a fluorescent retrograde tracer (Fluorochrome, Englewood, CO), at 2.5% in the tube. The effects of NRTN and GDNF were compared directly in littermates; the concentrations used were 0.14 or 1.65 mg/ml. After a survival period of 10–11 d the rats were given an overdose of pentobarbital and perfused transcardially with 4% paraformaldehyde in PBS. The lumbosacral spinal cord was removed, cryoprotected in sucrose-PBS solution, and cut at 30 μm on a cryostat. To determine the number of surviving motor neurons, we viewed serial sections coverslipped with Eukitt under a Reichert–Jung fluorescent microscope with a UV filter. Profiles of fluorogold-labeled sciatic motor neurons, localized in the ventral horn of spinal segments from L3–L6, ipsilateral to the nerve lesion, were counted on each section; no correction factor was applied to the counts. The values shown are means ± SEM (n = 3–7 per group).
In situ hybridization. Riboprobe preparation andin situ protocols have been described in detail previously (Golden et al., 1998).
RESULTS
Transport of factors in DRG sensory neurons
Previous studies show an effect of NRTN on rat embryonic DRG cultures (Kotzbauer et al., 1996). To examine the ability of NRTN to be transported within the adult rat DRG, we exposed the right sciatic nerve and delivered a 30 sec crush by applying firm pressure. After the crush, a single injection of 25–100 ng of125I-NRTN or125I-GDNF was injected into the nerve at the crush site. At 14–18 hr after the sciatic nerve injection, the L3–L6 DRG were removed and examined for the presence of125I-labeled factor. The accumulation of labeled protein in ipsilateral, but not contralateral, L4 and L5 DRG indicated that both 125I-GDNF and125I-NRTN were retrogradely transported, suggesting the presence of functional receptor complexes in sensory neurons (Fig. 1). As expected, radioactivity was generally undetectable within the L3 or L6 DRG (data not shown). Radiolabeled PSPN showed no ability to be transported to DRG neurons as assessed by comparison with contralateral counts (data not shown). The transport data for NRTN and PSPN correlated within vitro survival data from DRG cultures (Kotzbauer et al., 1996; Milbrandt et al., 1998).

125I-NRTN and125I-GDNF are transported into the dorsal root ganglia (DRG) of adult rats. DRG were removed from the rats 16 hr after the injection of iodinated ligand into the sciatic nerve (see Materials and Methods) and were imbedded in paraffin. Sections of 10 μm each were prepared, exposed to emulsion for 3–5 weeks, and then stained with cresyl violet. A, Microphotograph shows NRTN-labeled cells within the DRG. B, Microphotograph shows GDNF-labeled cells within the DRG. Scale bar, 100 μm.
Competition experiments were performed to determine the specificity and the relative affinities of NRTN and GDNF for receptor-mediated transport within DRG sensory neurons.125I-NRTN or125I-GDNF was injected in the presence or absence of excess unlabeled factors. The appearance of labeled NRTN within neurons of the DRG was blocked by 50- to 100-fold excess of unlabeled NRTN. Likewise, GDNF transport was blocked by using 50-to 100-fold excess of unlabeled GDNF (Fig.2). Neither a 100× excess of persephin (PSPN) nor NGF had any ability to compete with either GDNF or NRTN transport (data not shown).

Quantitation of retrograde transport and competition by unlabeled factor. Subsequent to the transport of iodinated ligand, DRG were removed, and the radioactivity was quantified. Excess cold GDNF (50- to 100-fold) reduces the detectable radioactivity caused by 125I-GDNF to background levels; 50- to 100-fold excess cold NRTN blocks the majority of labeled NRTN counts. A 100-fold excess of unlabeled NRTN, however, shows far less ability to compete with 125I-GDNF (the same being true for competition between excess unlabeled GDNF and labeled NRTN), suggesting a significant degree of receptor specificity. Contralateral counts are at background level (data not shown).
The ability of NRTN or GDNF to block transport of the other factor was examined also. A 100× excess of unlabeled GDNF blocked only 54% of radioactive NRTN counts and was clearly unable to block NRTN transport as efficiently as unlabeled NRTN itself. Likewise, a 100× excess of unlabeled NRTN only partially blocked the transport of GDNF, blocking 48% of radioactive counts (Fig. 2). These data demonstrate that even high concentrations of cold GDNF or NRTN only partially blocked transport of the other ligand, thus indicating a significant degree of selectivity in the receptor-mediated transport of NRTN and GDNF.
Size distribution of NRTN-transporting populations in the adult DRG
Recent publications describe the distribution of mRNA for RET, GFRα1, and GFRα2 within the adult DRG (Molliver et al., 1997;Bennett et al., 1998). Previous size/frequency analyses reveal that 40% of the total GFRα1-expressing population are large neurons that lack GFRα2. Coreceptor subpopulations of small DRG neurons can be divided into those that express GFRα2 but not GFRα1, those that express GFRα1 but not GFRα2, and a third subpopulation that express both GFRα1 and GFRα2 (Bennett et al., 1998). We hypothesized that if GDNF had a receptor preference for GFRα1 and NRTN showed a similar preference for GFRα2, then the size/frequency distributions of the transported ligands would reflect the size/frequency distributions of the receptors and would be distinct from each other. To test this hypothesis, we determined the size/frequency distributions of neuronal populations transporting GDNF or NRTN to allow for comparison to the known distributions of neurons expressing RET, GFRα1, and GFRα2. As expected, the distribution of GDNF- or NRTN-labeled neuronal populations fell within that of RET-immunoreactive neurons (Fig.3). Furthermore, both NRTN and GDNF were transported into a population of small cells reported to coexpress GFRα1 and GFRα2 (Bennett et al., 1998). A striking observation, particularly in light of the competition results (see Fig. 2), is that GDNF also was transported into a population of larger cells, whereas NRTN was transported into many fewer of these larger cells (Fig. 3). This subset likely represents the large GFRα1-expressing population previously reported (Bennett et al., 1998). As a result of this differential transport, the average size of the GDNF population (1011 μm2 ± 728; n = 339 cells) was larger than the average NRTN-transporting cell (746 μm2 ± 504; n = 189 cells). This difference in mean size of the cells transporting the factors is highly significant (p < 0.0001). The pattern of cell-size distributions reported here for GDNF is very similar to the pattern of cell-size distributions reported after125I-GDNF injection into the footpad of the hindlimb of P1 rats (Matheson et al., 1997). These data and our immunohistochemical data (see Fig. 5) are consistent with a model in which GFRα1-expressing cells selectively transport GDNF, GFRα2-expressing cells selectively transport NRTN, and cells that express both coreceptors transport both ligands.

Cell size distributions of L4 and L5 DRG neurons immunocytochemically stained for RET and retrogradely labeled with125I-NRTN or 125I-GDNF. Both GDNF- and NRTN-labeled cells are mostly small, consistent with the significant proportion of small RET-stained cells. However, GDNF also labels a sizable percentage of larger cells that fail to transport NRTN to the same extent. The total number of cells measured was 189 cells for NRTN, 339 cells for GDNF, and 983 cells for RET from at least five different sections from five different animals. All cells containing nuclei and with 10 or more silver grains visible under bright-field conditions were determined to be labeled and were traced by using camera lucida. The areas of these cells were determined later by using the SigmaScanPro4 program.
The apparent coreceptor selectivity of the size/frequency data raised the question: How can NRTN and GDNF be competing with one another if they are being transported by different coreceptors? To begin to address this question, we performed a size/frequency analysis on the competition experiments to determine whether competition was limited to a specific size subpopulation or represented decreases in every area bin across the population. Figure 4 shows the cell-size distributions observed in such a competition experiment. A total of 186 cells were counted in the125I-GDNF control and 119 cells in the NRTN competition experiments. NRTN competition caused a selective decrease in the number of small cells transporting GDNF such that the average size of 125I-GDNF-labeled cells in the presence of NRTN increased 145% (p < 0.0001). Thus, NRTN primarily competes with GDNF for transport into a subpopulation of small neurons. One possible model explaining this observation is that of indirect competition of ligands via RET. This model would propose that cells contain a limiting amount of RET that is necessary for ligand internalization leading to transport. If a cell contains multiple coreceptors (as does a subpopulation of small neurons in the DRG), whichever ligand is present in excess will bind to its specific coreceptor and essentially “win” the competition for the limited amount of RET, resulting in the transport of that ligand. This model implies that competition will occur primarily in cells containing both coreceptors. This mechanism is consistent with the observation that the size/frequency distribution of125I-GDNF-transporting cells in the presence of excess unlabeled NRTN revealed a discrete loss of a subpopulation of small cells, with other subpopulations remaining relatively unaffected.

Unlabeled NRTN changes the size/frequency distribution of 125I-GDNF. Size/frequency analysis of a competition experiment reveals a significant decrease in a population of small GDNF-transporting cells. The rest of the distribution remains primarily the same. Competition results in an increase in the average size of GDNF-transporting cells. For the125I-GDNF-transport control experiment, 186 cells were measured; for the competition experiment, 119 cells were measured. Cell areas were determined as described above.
Immunocytochemical analysis of NRTN-transporting populations in adult DRG
The DRG can be divided into roughly three main classes of neurons: small nociceptive neurons that express the NGF receptor TrkA (Carroll et al., 1992; Crowley et al., 1994; Smeyne et al., 1994), middle-sized neurons of undefined function that express TrkB and respond to BDNF (Schecterson and Bothwell, 1992), and large, primarily proprioceptive neurons that express TrkC and bind NT-3 (Ernfors et al., 1994; Farinas et al., 1994). These size/function categories represent a generalized division, but many smaller classes of DRG neurons do not fall cleanly along these dividing lines. For example, whereas all small DRG neurons express TrkA early in rat development, one-half of these neurons go on to downregulate TrkA after birth and subsequently express RET (Molliver et al., 1997). This population of small RET-expressing DRG neurons can be visualized by labeling with the plant lectin IB4 (Bennett et al., 1996; Molliver and Snider, 1997). Although soma-size measurements allowed for a preliminary determination of NRTN-responsive neuronal subsets, immunocytochemical experiments enabled us to determine the neurochemical identity of NRTN-transporting populations. After transport of labeled factors, DRG were removed and stained with antibodies identifying different neuronal populations. These data are presented (Fig. 5) as both dark-field and bright-field images so as to clearly distinguish among the four classes of cells: stained and unlabeled cells, labeled and unstained cells, cells that are both stained and labeled, and cells that are neither labeled nor stained. Labeled GDNF protein was detected in three distinct populations of DRG neurons: a small subset of TrkA-expressing cells; a large proportion of the small, IB4/non-TrkA cells; and a population of large, TrkC/noncalretinin-staining cells (see Table1; Fig. 5) (Molliver et al., 1997). In contrast, the majority of NRTN-labeled cells belonged to the IB4/non-TrkA-expressing population, with a significant minority of calretinin-staining cells (Table 1; Fig. 5). NRTN was virtually absent from TrkA-expressing cells (Table 1). Immunocytochemistry also was performed for the DRG subpopulation marker α-internexin, a neurofilament protein that shows almost complete colocalization with IB4 and labels small neurons lacking TrkA (Molliver and Snider, 1997). α-Internexin colocalized extensively with NRTN and to a lesser, but still significant, degree with GDNF (Table 1; Fig. 5). These data agree with reported receptor distributions in which GFRα1 is present in small IB4-labeled neurons, a subpopulation of TrkA neurons, as well as a significant fraction of larger neurons, whereas GFRα2 is restricted mainly to the small IB4 cells (Bennett et al., 1998). As expected, antibodies to the RET receptor stained all NRTN- and GDNF-transporting neurons (Fig. 5).

Immunohistochemistry reveals colocalization of RET, α-internexin, calretinin, and TrkA, with transported NRTN and GDNF. All panels are matched dark-field and bright-field images.A and B show RET-stained125I-GDNF-transporting cells. C andD show α-internexin-stained125I-NRTN-transporting cells. E andF show calretinin-stained125I-NRTN-transporting cells. G andH show TrkA-stained 125I-GDNF-transporting cells. Large arrowheads indicate examples of stained but nontransporting (unlabeled) cells; split arrowheadsindicate examples of labeled (transporting) but unstained cells;small arrows indicate examples of co-stained and labeled cells; asterisks indicate examples of unlabeled and unstained cells. All images were taken with a 20× magnification objective. Scale bar, 50 μm.
Size and immunocytochemical characteristics of125I-NTN- and 125I-GDNF-transporting populations of adult rat DRG neurons
Retrograde transport in sympathetic neurons
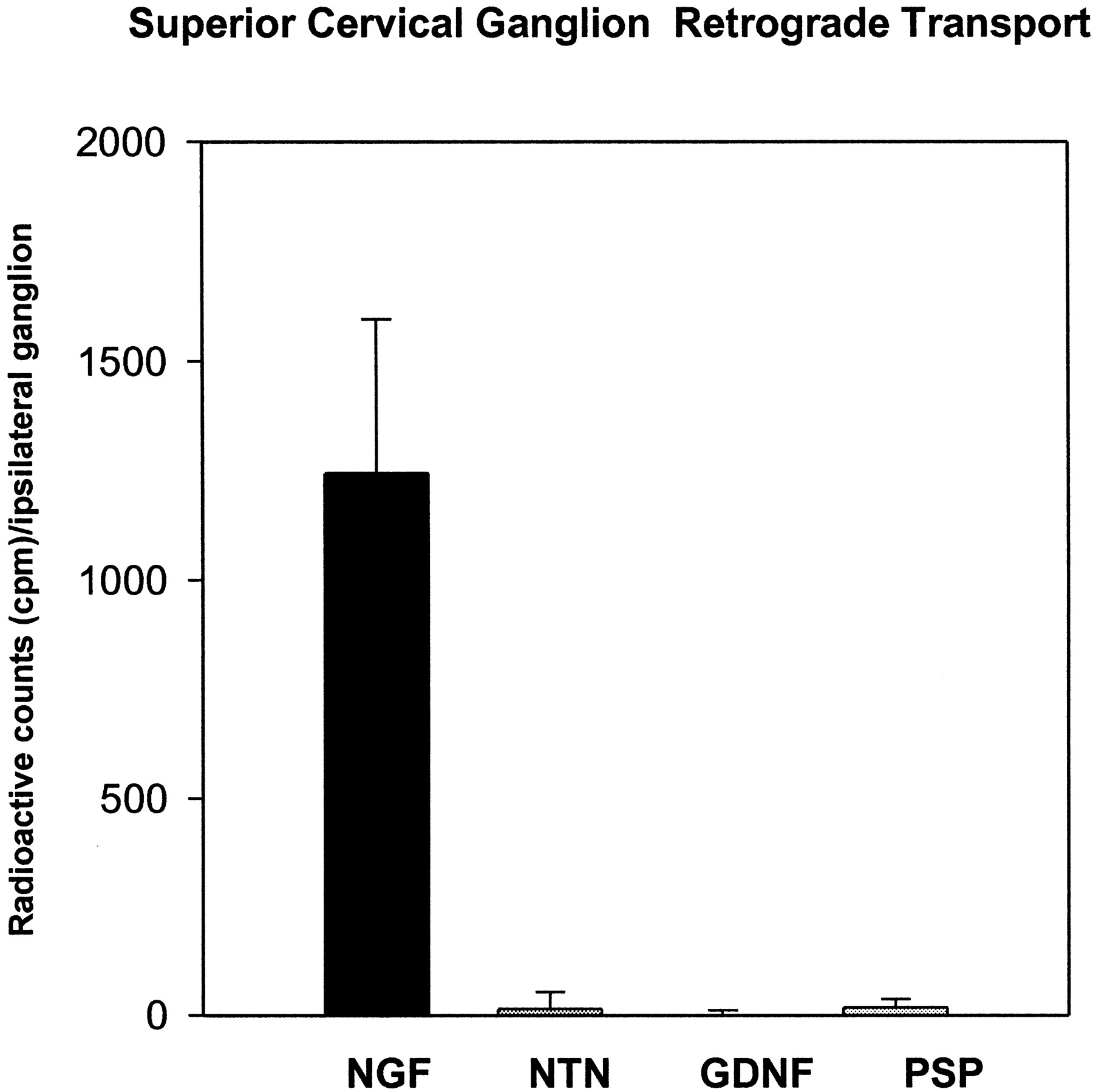
On the basis of the reported ability of GDNF (Buj-Bello et al., 1995) and NRTN (Kotzbauer et al., 1996) to rescue embryonic and early postnatal SCG neurons in the rat, we examined the retrograde transport of factors to the SCG after injection into the anterior eye chamber. Although nerve growth factor (NGF) was transported efficiently into adult SCG neurons 14–20 hr after injection into the anterior eye chamber, at no time was NRTN, GDNF, or PSPN detected in the SCG (Fig.6; time course not shown). These data are consistent with previously reported results on the lack of GDNF transport into the SCG of adult rats (Yan et al., 1995) and suggest that, unlike the DRG, the functional significance of NRTN and GDNF for the sympathetic system may be limited to embryonic and early postnatal development. Interestingly, a recent in situ hybridization report on GFRα1 and GFRα2 expression in the adult mouse reveals that, whereas RET remains expressed at high levels in the SCG, GFRα1 and GFRα2 mRNA levels are very low (Golden et al., 1998). The lack of an SCG phenotype observed in the NRTN and GFRα2 adult knock-out mice (Heuckeroth et al., 1999; Rossi et al., 1999) is consistent with the lack of retrograde transport we observed in the adult rat. Again, the lack of PSPN transport correlated closely with previous experiments indicating that PSPN has no ability to promote the survival of SCG neurons (Milbrandt et al., 1998).

Quantification of retrograde transport of125I-labeled NRTN, GDNF, PSPN, and NGF into the superior cervical ganglion after injection in the anterior eye chamber. Injection of labeled NGF results in the clear transport of the labeled material, whereas neither NRTN nor PSPN nor GDNF shows transport above contralateral background.
Retrograde transport to motor neurons
Both NRTN and GDNF rescue embryonic motor neurons in culture (Henderson et al., 1994; Oppenheim et al., 1995; Kotzbauer et al., 1996; Klein et al., 1997); however, little analysis has been performed in the adult. Retrograde transport into spinal lumbar motor neurons of the adult rat was determined by using autoradiography. The spinal cords from the injected animals (described previously) were removed and counted to determine motor neuron transport for125I-NRTN and125I-GDNF. After sciatic injection, autoradiographic analysis of the spinal cords showed that125I-GDNF was localized to many ventral motor neurons, whereas NRTN labeled only a rare motor neuron (Fig.7). These data indicate that GDNF was retrogradely transported to a greater extent than NRTN in adult motor neurons and again suggest physiologically relevant differences in the actions of GDNF and NRTN on these neurons.
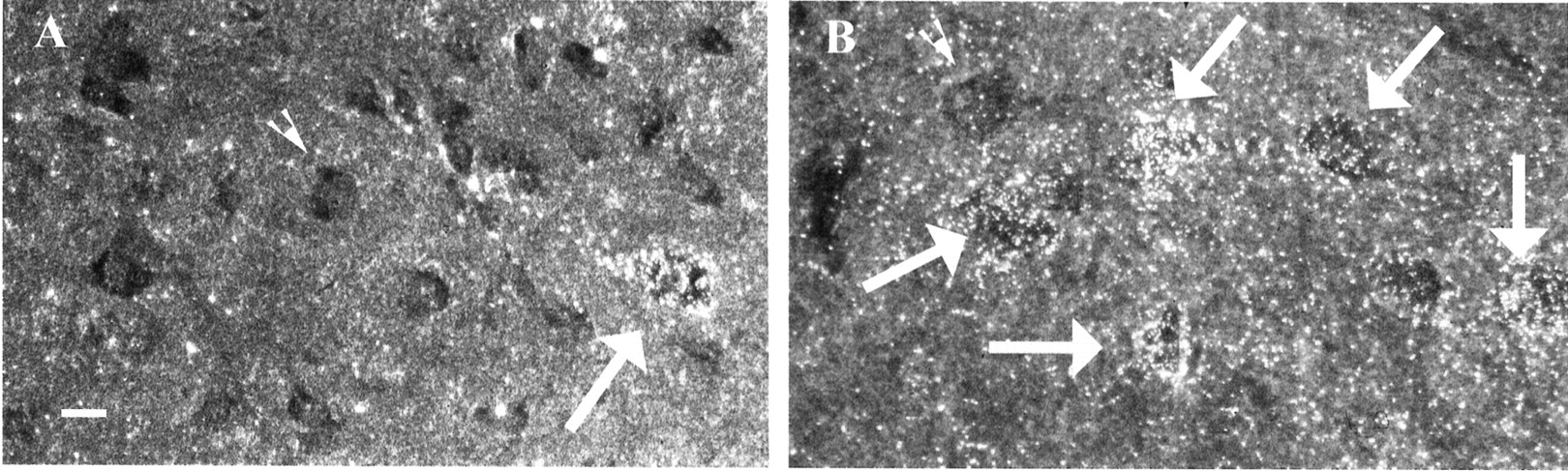
Autoradiographic analysis of transport of GDNF and NRTN into motor neurons reveals that GDNF is transported into motor neurons at greater levels than is NRTN. A, Single125I-NRTN-labeled motor neuron. B,Representative field of 125I-GDNF-labeled motor neurons.Large arrows point to labeled cells;arrowheads point to unlabeled cells. Scale bar, 50 μm.
NRTN rescues immature motor neurons
Besides its in vitro survival effects, GDNF prevents the death of motor neurons in the neonate after axotomy (Oppenheim et al., 1995; Yan et al., 1995). To determine whether NRTN exhibited similar effects, we performed sciatic nerve axotomies on P2 rats, and NRTN, GDNF, or BSA was applied at the site of the lesion. At 10 d after axotomy the animals treated with 0.14 mg/kg NRTN had a greater number of living motor neurons than animals treated with the BSA control (Student's t test, p < 0.00002). At the higher dose of 1.65 mg/kg, NRTN was able to rescue ∼80% of the motor neurons (Fig. 8). One hundred percent survival had been determined previously by Vejsada et al. (1998). The dose dependency of motor neuron rescue was similar to that seen for GDNF (Fig. 8) and PSPN (Milbrandt et al., 1998) in this paradigm.

NRTN promotes the survival of neonatal motor neurons after axotomy. Comparison of NRTN- and GDNF-mediated rescue reveals that NRTN possesses equal efficacy and potency for preventing death 10 d after the sciatic axotomy of P2 motor neurons.
The ability of NRTN to rescue axotomized motor neurons in the neonate suggests that functional transport was occurring, at least at this developmental stage. Because the retrograde transport studies in adult sensory neurons suggested a relatively selective pairing of NRTN transport with GFRα2, one possible explanation for the apparent discrepancy between the survival-promoting ability of NRTN for developing motor neurons and the reduced transport by mature motor neurons is that the expression of GFRα2 was downregulated developmentally on maturation. Consistent with this hypothesis,in situ hybridization analysis reveals a robust expression of GFRα2 in motor neurons during development, whereas minimal expression was observed in the adult (Fig.9). In contrast, GFRα1 expression was retained in the adult. This strongly suggests that the lack of NRTN transport observed in the adult animal reflected the low expression levels of GFRα2 and a decreased ability of NRTN to bind GFRα1. These results provide further support for the hypothesis that the transport of GDNF and NRTN in the adult animal is mediated functionally by GFRα1 and GFRα2, respectively.

Adult motor neurons downregulate the expression of GFRα2 but maintain the expression of GFRα1. In situhybridization for GFRα1, GFRα2, and RET was performed on adult spinal cord (A–C) and E18 (D–F). Although levels of GFRα1 and RET remain detectable throughout the adult ventral spinal cord, levels of GFRα2 decline to trace amounts. DH, Dorsal horn;VH, ventral horn.
DISCUSSION
NRTN and GDNF are related trophic molecules that signal via the receptor tyrosine kinase RET in combination with one of several GPI-linked coreceptors belonging to the GFRα family. To date, conflicting in vitro evidence exists as to whether NRTN and GDNF have preferred coreceptors. We examined the retrograde transport of NRTN and GDNF in sensory neurons of the DRG, motor neurons of the lumbar spinal cord, and sympathetic neurons of the SCG to provide insight into which neurons responded to NRTN in vivo. Our data suggest that some neuronal subpopulations showed a clear factor preference. Furthermore, a comparison of our findings with previously reported expression patterns of the GFRα1 and GFRα2 coreceptors indicate that, in the adult animal, NRTN and GDNF were transported selectively by different GFRα coreceptors.
GFRα coreceptor selectivity within the DRG
NRTN, like GDNF, was retrogradely transported to the DRG. The transport of 125I-NRTN into the DRG was blocked fully by a 100-fold excess of unlabeled NRTN but blocked only partially by a similar excess of GDNF. Likewise, GDNF transport was blocked only partially by a 100-fold excess of unlabeled NRTN. Closer examination reveals that this partial competition appeared to be occurring primarily in a population of small cells that we predict to be the dual receptor-containing population. We hypothesize that coreceptor selectivity was being maintained in this subpopulation but that ligand-bound coreceptors were competing for limited amounts of RET and only the winner of this competition, presumably the ligand/coreceptor complex in excess, was being transported. Other models are possible as well, and this model will have to be tested more rigorously in vitro to ascertain its validity. PSPN, the third member of the GDNF family, was unable to block the retrograde transport of either NRTN or GDNF and was not transported by DRG neurons. These findings correlate with earlier in vitroreports that PSPN does not bind to either GFRα1 or GFRα2 (Milbrandt et al., 1998).
Immunohistochemical analysis indicated that NRTN-transporting neurons expressed RET but only rarely expressed the NGF receptor TrkA (Table1). The cells that retrogradely transport NRTN were composed of at least two populations: a substantial group of small neurons (87% of NRTN-transporting neurons), which expressed α-internexin and bound IB4, and a small group (13%) of medium-to-large neurons, which expressed calretinin. Although many IB4 neurons also transported GDNF, calretinin neurons appeared to be selective for NRTN. Little is known about the function of these calretinin-expressing neurons.
It is instructive to consider our data with respect to previous descriptions of the distributions of GFRα1 and GFRα2 in the DRG.In situ hybridization analyses of GFRα1 and GFRα2 expression indicate that GFRα1 mRNA is expressed by both small and large neurons, whereas GFRα2 is expressed primarily by small neurons: 40% of GFRα1-expressing cells have a somal area >1000 μm2; only 17% of GFRα2 cells are this large (Bennett et al., 1998). Notably, the size distribution of NRTN-transporting neurons (see Fig. 3) was very similar to that reported for GFRα2-expressing cells, whereas the distribution of GDNF-transporting cells matched that of neurons expressing GFRα1. Our results strongly suggest that in vivo GDNF preferentially bound GFRα1, whereas NRTN preferentially bound GFRα2. These results are somewhat in contrast to in vitro studies (Baloh et al., 1997; Sanicola et al., 1997; Suvanto et al., 1997) that suggest a significant degree of receptor–ligand cross-talk. At pharmacological doses NRTN and GDNF apparently can interact with either GFRα1 or GFRα2, whereas at lower and perhaps more physiological doses ligand–receptor selectivity is a major determinant of ligand function. An additional possibility is that coreceptor selectivity changes throughout the lifetime of the animal, being less selective during development but becoming more selective as the animal reaches maturity. All of the experiments suggesting coreceptor promiscuity have been performed in developing animals, developing cells, or immortalized cells in culture (Baloh et al., 1997; Jing et al., 1997; Sanicola et al., 1997; Suvanto et al., 1997; Cacalano et al., 1998; Enomoto et al., 1998; Trupp et al., 1998). Maturation may be associated with a process (e.g., changes in RET isoforms or changes in GFRα splice forms) that leads to increased selectivity.
Developmental changes in GFRα expression account for differential transport
Both NRTN and GDNF promote the survival of fetal motor neuronsin vitro (Henderson et al., 1994; Oppenheim et al., 1995;Kotzbauer et al., 1996; Klein et al., 1997). We examined the in vivo actions of GDNF and NRTN in both mature and immature motor neurons. Given the reported similarity of action in cultures of immature motor neurons, initially we were somewhat surprised to find that, in the adult animals, 125I-NRTN was transported less efficiently to motor neurons than was125I-GDNF. In vivo experiments indicate that GDNF can rescue motor neurons in the neonate after axotomy (Oppenheim et al., 1995; Yan et al., 1995). The ability of GDNF to prevent death after axotomy is confirmed in Figure 8, and we show that NRTN acted in a manner very similar to GDNF in that paradigm. Like the DRG competition data, this apparent dichotomy also may be explained on the basis of the expression and selectivity of GFRα1 for GDNF and GFRα2 for NRTN. In situ hybridization analysis shows that GFRα1 and GFRα2 were both expressed in developing motor neurons, whereas GFRα2 expression was reduced significantly in the adult (Fig.9). These data suggest that GDNF may be more effective than NRTN in adult models of motor neuron injury and degeneration.
PRSP, like NRTN, failed to be transported extensively into motor neurons after a sciatic injection. The lack of PSPN transport into adult motor neurons conceivably could reflect a reduction in the bioactivity of PSPN after radioactive labeling. However, the failure of excess unlabeled PSPN to reduce the transport of labeled NRTN or GDNF clearly supports in vitro results indicating that PSPN lacks any capacity to bind to GFRα1 or GFRα2 (Milbrandt et al., 1998). Intriguingly, PSPN resembles NRTN in being able to prevent the death of axotomized immature motor neurons while apparently lacking the ability to be transported into adult motor neurons. These data suggest that the PSPN-binding receptor component GFRα4 may be regulated developmentally in motor neurons.
Knock-out phenotypes suggest that cross-talk is limited even during development
Recent descriptions of the similar phenotypes of the GDNF (Moore et al., 1996; Pichel et al., 1996; Sanchez et al., 1996; Cacalano et al., 1998; Enomoto et al., 1998) and GFRα1 (Cacalano et al., 1998;Enomoto et al., 1998) knock-out animals and the NRTN and GFRα2 (Heuckeroth et al., 1999; Rossi et al., 1999) knock-out animals support the hypothesis that, even in the developing animal, GDNF and NRTN exhibit a significant degree of receptor and functional specificity. NRTN appears to be unable to compensate for the loss of GDNF in either kidney or enteric neuron development although NRTN can act on both of these systems in vitro. Similarly, GDNF cannot compensate for NRTN in enteric, parasympathetic, or trigeminal neuronal development. Most strikingly, the NRTN knock-outs demonstrate a 45% reduction in GFRα2-expressing neurons in the DRG (suggesting that only a subpopulation of GFRα2-expressing neurons can be rescued by GDNF). This reduction correlates exactly with the 54% of transported NRTN counts that can be competed by excess unlabeled GDNF. However, some degree of cross-talk appears to be occurring because the deficits reported in the DRG, SCG, and nodose ganglia in the GDNF knock-out are greater than those observed in the GFRα1 knock-out, suggesting that GDNF is mediating some of its survival-promoting ability via an additional receptor complex.
Regional differences in NRTN transport correlate with coreceptor expression
Although NRTN has potent effects on embryonic DRG, SCG, and motor neurons in culture, its effects in vivo on the adult DRG, SCG, and spinal motor neuron systems are strikingly different. Discrete populations of neurons within the adult rat DRG were capable of transporting both NRTN and GDNF at significant levels. In contrast, adult motor neurons were capable of only low levels of NRTN transport while retaining the ability to transport GDNF at higher levels. Last, neurons of the adult SCG showed no ability to transport NRTN or GDNF. The differences between NRTN and GDNF in these transport paradigms appeared to correlate highly with the presence or absence of GFRα1 and GFRα2 coreceptors in the regions that were examined, suggesting that coreceptor expression and selectivity are crucial predictors of GDNF family function.
Conclusion
In summary, an examination of the retrograde transport of125I-NRTN and125I-GDNF indicates that the transport of these factors is mediated selectively by GFRα2 and GFRα1, respectively. The ability of GDNF and NRTN to compete for transport in sensory neurons appears to require the expression of both coreceptors and is restricted mainly to a subpopulation of small neurons. The decrease of GFRα2 expression in adult motor neurons suggests that GDNF is likely to have a more important physiological role in motor neurons than NRTN does in the adult animals and that GDNF may be a more effective pharmacological agent in adult motor neurons.
Footnotes
This work was supported by the Swiss National Science Foundation, the Association Francaise contre les Myopathies, and the Swiss Foundation of Neuromuscular Diseases (to A.C.K.); by National Institutes of Health Grants AG13729 and AG13730 (to E.M.J. and J.M); and by Genentech, Incorporated (South San Francisco, CA). M.L.L. was supported by a Lucille P. Markey fellowship and is a Howard Hughes Medical Institute Predoctoral Fellow.
Correspondence should be addressed to Dr. Eugene M. Johnson, Jr., Department of Molecular Biology and Pharmacology, Washington University School of Medicine, 4566 Scott Avenue, Box 8103, St. Louis, MO 63110. E-mail: ejohnson{at}pcg.wustl.edu.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}