

Abstract
The development of olfactory bulb projections that form the lateral olfactory tract (LOT) is still poorly understood. The septum and the olfactory cortex have been shown to secrete diffusible factors repelling olfactory axons in vitro and are likely to cause the axons to avoid the septum region in vivo. Slit2, a member of the Slit gene family, has been proposed to be this septal factor based on its expression in the embryonic septum and its ability to repel and collapse olfactory axons. However, this issue is still controversial, and recent in vitro studies have questioned the role of the septum and Slit proteins in organizing LOT projections. We therefore decided to examine directly the role of Slit proteins in mediating olfactory axon guidance in vivousing mice with targeted deletions in the Slit1 andSlit2 genes. When olfactory bulb explants are cocultured with septum from Slit1- and/orSlit2-deficient mice, the septum repulsive activity for olfactory bulb axons is progressively abolished in a gene dose-dependent manner. Anterograde tracing of olfactory bulb axons showed that the LOT develops normally in Slit1 orSlit2 single-deficient mice but is completely disorganized in Slit1/Slit2double-deficient embryos, with many axons reaching the midline and entering the septum region. Therefore, our study showed that the septum chemorepellent is a combination of Slit1 and Slit2 and that these molecules play a significant role in olfactory bulb axon guidancein vivo.
The organization of axonal projections in the rodent olfactory system has been extensively characterized. Axons from olfactory receptor neurons in the olfactory epithelium project ipsilaterally to glomeruli in the main olfactory bulb (OB), in which they synapse on the dendrites of the mitral and tufted cells. These neurons project ipsilaterally to the anterior olfactory nucleus and to higher olfactory centers collectively referred to as the primary olfactory cortex (Shipley and Ennis, 1996; Zou et al., 2001). The axons of the mitral and tufted cells are located immediately under the pial surface (Schwob and Price, 1984) and form the lateral olfactory tract (LOT). The development of bulbofugal projections is still poorly understood (Zou et al., 2001). Isolated LOT axons start leaving the OB by embryonic day 14 (E14) in rat or E12.5 in mouse, and the following day, an LOT has clearly formed (Pini, 1993;Lopez-Mascaraque et al., 1996; Sugisaki et al., 1996). The precise pathfinding of LOT axons in the telencephalon is thought to be controlled by a combination of short-range and long-range cues (Sugisaki et al., 1996; Hirata and Fujisawa, 1997; Sato et al., 1998;Soussi-Yanicostas et al., 2002). In this regard, the septum and the olfactory cortex have been shown to secrete diffusible factors that repel LOT axons in vitro (Pini, 1993; Hu and Rutishauser, 1996) and that could prevent them from approaching the septum region in vivo. The identity of this septal-derived repulsive activity is still unknown. Recently, Slit2, a member of the Slit gene family, has been hypothesized to be this septal factor based on its expression by the embryonic septum and its ability to repel and collapse OB axons (Li et al., 1999; Nguyen-Ba-Charvet et al., 1999). Slit proteins are a family of chemotropic factors (Brose and Tessier-Lavigne, 2000) first identified in Drosophilaembryo, in which it regulates midline crossing by commissural axons and the fasciculation of longitudinal axons (Rothberg et al., 1990; Kidd et al., 1999; Rajagopalan et al., 2000a; Simpson et al., 2000b). In mammals, three Slit genes (Slit1–Slit3) have been cloned (Holmes et al., 1998; Itoh et al., 1998; Brose et al., 1999; Yuan et al., 1999b). In Drosophila, Slit function is mediated by the Roundabout (Robo) receptors (Kidd et al., 1999). So far, three robo genes have been found in flies (Kidd et al., 1998; Rajagopalan et al., 2000b; Simpson et al., 2000b) and mammals (Brose et al., 1999; Li et al., 1999; Yuan et al., 1999a).
Some recent in vitro studies have questioned the role of the septum and Slit proteins in organizing LOT projections because, in organotypic cultures, mitral cell axons elongate along their normal pathway in the absence of septum (Sugisaki et al., 1996) or in the presence of a Robo1 ectodomain (Robo1-Fc), which should block Slit protein function (Hirata et al., 2001). In addition, Robo1-Fc or Robo2-Fc do not abolish the septum repulsive activity in repulsion assays (Patel et al., 2001). Therefore, direct evidence for a role of Slit proteins in directing OB axon guidance in vivo is still missing. We addressed this controversial issue by analyzing LOT development in mice with targeted deletions in the Slit1 andSlit2 genes (Plump et al., 2002). Here we demonstrate that the septum chemorepellent is a combination of Slit1 and Slit2, and, in their absence, the LOT does not form properly.
MATERIALS AND METHODS
Animals. Slit-deficient mice were generated and genotyped by PCR as described previously (Plump et al., 2002). Actin-green fluorescent protein (GFP) mice and embryos [strain TgN(GFPU)5Nagy; The Jackson Laboratory, Bar Harbor, ME] (Hadjantonakis et al., 1998) were selected using a MZFLIII fluorescence stereomicroscope equipped with a GFP filter (Leica, Rueil-Malmaison, France). The day of the vaginal plug was considered as E0, and the day of birth was considered as postnatal day 0 (P0). Pregnant dams were anesthetized with chloral hydrate (40 mg/kg).
Embryos were fixed by immersion in 4% paraformaldehyde in 0.1m phosphate buffer, pH 7.4 (PFA). Postnatal mice were perfused transcardially with 4% PFA. Embryo heads or brains were postfixed overnight in the same fixative.
Immunocytochemistry. After fixation, E15 embryo heads were cryoprotected in 10% sucrose and frozen in isopentane (−55°C). Coronal cryostat sections (20 μm) were blocked 1 hr at room temperature (RT) in PBS containing 0.2% gelatin (Prolabo, Fontenay-sous-Bois, France) and 0.25% Triton X-100 (Sigma, St. Louis, MO) and then incubated overnight at RT with a rabbit anti-neuropilin-1 antiserum (1:500). This antibody was generated as described previously (Kolodkin et al., 1997), using a GST-fusion neuropilin-1 construct (a king gift from Dr. A. Kolodkin, Johns Hopkins University, Baltimore, MD). Sections were then incubated in a biotinylated goat anti-rabbit antibody (1:200; Vector Laboratories, Burlingame, CA) and an HRP-conjugated streptavidin (1:400; Amersham Biosciences, Orsay, France). The sections were developed with a diaminobenzidine reaction. The same sections were then incubated with a goat anti-GFP antibody (1:500; Molecular Probes, Eugene, OR), followed by a Cy3- or FITC-conjugated goat anti-rabbit antibody (1:200; Jackson ImmunoResearch, West Grove, PA).
OB explant assays. OB explants from E14 rat embryos were cocultured with aggregates of Slit1-transfected COS cells as described previously (Nguyen-Ba-Charvet et al., 1999). The full-length mouse Slit1 expression construct was a kind gift from Dr. Yi Rao (Washington University, St. Louis, MO) (Li et al., 1999). The Slit1-N expression construct was obtained by PCR amplification of the N-terminal portion of Slit1 (encoding amino acids 32–1118), stopping before epidermal growth factor (EGF) repeat 6. The amplified cDNA was then subcloned in the pSecTag2C expression construct (Invitrogen, Carlsbad, CA). Constructs were sequenced, and their expression was tested by transient transfection in COS cells, followed by Western blotting using anti-myc antibody (clone 9E10; Santa Cruz Biotechnology, Santa Cruz, CA). After 24 hr, cocultures were fixed for 1 hr in 4% PFA and processed for immunocytochemistry with a neuron-specific anti-class III β-tubulin antibody (TUJ-1; Babco, Richmond, CA) (Moody et al., 1989) as described previously (Nguyen-Ba-Charvet et al., 1999).
The cocultures of septum from E14–E15 Slit-deficient mice with OB explants from E14–E15 actin-GFP mice were made in Neurobasal medium containing B27 additives (Invitrogen), 0.5 mm glutamine (Invitrogen), and 0.1 μg/ml heparin (Sigma). After 24 hr, cocultures were fixed and processed for immunocytochemistry with a rabbit polyclonal anti-GFP (1:1000; Molecular Probes) as described above.
Repulsive activity is measured by the neurite outgrowth ratio P/D, where P is the extent of neurite outgrowth on the side proximal to the cell aggregate or the septum explant, and D is the extent of neurite outgrowth on the side distal to the cell aggregate or the septum explant (Chen et al., 2000). This quantification was performed blind with the Metamorph image analysis system (Universal Imaging Corporation, Downington PA). The statistical significance was determined by ANOVA with a Fisher's PLSD test, using Statview (Abacus Concepts, Cary, NC).
DiI tracing. The olfactory bulb in E14 to P0 brains was labeled with the lipophilic tracer DiI (Molecular Probes) by injection with 2 μl of a 1% solution of DiI in 300 mmsaccharose or by placing a crystal of DiI in the olfactory bulb as described previously (de Castro et al., 1999). Some injected brains were cut in 100 μm sections with a vibratome (Leica). The sections were then counterstained in a solution of 10 μg/ml Hoechst 33258 (Sigma) in PBS for 30 min and rinsed three times in PBS.
RESULTS
Slit1 and Slit2 expression in the septum
Slit1- and Slit2-deficient mice were generated with an internal ribosomal entry site element inserted between the Slit promoter and a tau-GFP reporter gene (Plump et al., 2002). Therefore, GFP expression should reflect the expression of the endogenous Slit genes. We showed previously that, in rat embryos, the septum expresses mRNAs for both Slit1 andSlit2 but not Slit3 during LOT development (Li et al., 1999; Nguyen-Ba-Charvet et al., 1999; Marillat et al., 2002). Similarly, we found that, in E15 Slit1- andSlit2-deficient mice, GFP was highly expressed in the septum (Fig. 1). A robust GFP expression was also detected in the neocortex and ganglionic eminence ofSlit1-deficient mice (Fig. 1C) in which high levels of Slit1 mRNAs have been detected (Marillat et al., 2002;Whitford et al., 2002). Likewise, in Slit2-deficient mice, GFP was expressed in the septum (Fig.1B,D) and also in the internal capsule (Fig. 1D). Therefore, at E15, GFP expression in the telencephalon of Slit1- andSlit2-deficient mice was well correlated withSlit1 and Slit2 mRNA expression patterns. At this stage, no obvious difference in the GFP expression pattern was detected between Slit1+/− and Slit1−/− or Slit2+/− and Slit2−/− mice, although, as expected, GFP expression was stronger in homozygous animals than in heterozygotes (data not shown). This suggests that septal neurons were not affected by the absence of eitherSlit1 or Slit2. In some cases, the LOT was visualized using immunocytochemistry for the class III semaphorin receptor neuropilin-1, which is expressed in LOT axons (Fig. 1) (Kawakami et al., 1996). Overall, these results confirm that there is a strong expression of Slit1 and Slit2 in the septum during LOT development.

Expression of Slit1 (S1) and Slit2 (S2) in the embryonic septum assessed by GFP staining. Immunocytochemistry against GFP was performed on 20 μm coronal cryostat sections of heterozygous E15 Slit1(A, C) and Slit2(B, D) -deficient embryos to assess Slit1 and Slit2 expression. The LOT (arrowhead) is stained with anti-neuropilin-1 (red). Aand B show a rostral section, whereas Cand D represent a more caudal section of the forebrain. At this stage, GFP is highly expressed in the septum (Se) of both Slit1 andSlit2-deficient mice. D, GFP is also present in the internal capsule (arrow) ofSlit2-deficient mice. ge, Ganglionic eminence. Scale bars: A, C, 200 μm;B, 150 μm; D, 300 μm.
Both Slit1 and Slit2 can repel OB axonsin vitro
In vitro, recombinant Slit2 can repel OB axons (Li et al., 1999; Nguyen-Ba-Charvet et al., 1999) (Fig.2B). However, because Slit1 and Slit2 are both expressed in the septum of rodents, we wanted to determine whether Slit1 could also repel OB axons. Therefore, we cultured OB explants from E15 rat embryos at a distance from COS cell aggregates expressing full-length Slit2 or full-lengthSlit1 for 24–36 hr (Itoh et al., 1998) (see Materials and Methods). Mock-transfected COS cells were used for control experiments. Immunostaining of the explants with anti-class III-β-tubulin showed that, with control COS cells, axon outgrowth was radial (16 of 16 explants) (Fig. 2A). In contrast, axonal outgrowth was asymmetric, with COS cells expressing Slit2 (20 of 20 explants) or Slit1 (nine of nine explants), with axons growing almost exclusively on the distal side of the explants, away from Slit1 (0.094 ± 0.034, P/D ratio ± SEM) (Fig. 2C) or Slit2-expressing cells (0.157 ± 0.032) (Fig. 2B). Therefore, both Slit1 and Slit2 can repel E15 rat OB axons (Patel et al., 2001).

Illustration of the cocultures of E14–E15 rat olfactory bulb explants next to aggregates of control COS cells (A) or COS cells transfected withSlit2 (B), Slit1(C), or the N-terminal portion ofSlit1 (D). All explants were fixed and stained with anti-β-tubulin antibodies. Olfactory bulb axons grow symmetrically in the case of control cells (A), whereas they are strongly repelled by Slit1- orSlit-2-expressing COS cell aggregates (B–D). Scale bars, 100 μm.
Slit2 is known to be cleaved into two fragments, a large N-terminal fragment and a short C-terminal fragment (Wang et al., 1999). The N-terminal portion of Slit2 (Slit2-N) is sufficient for binding of Slit2 to its receptors Robo1 and Robo2 and for mediating Slit2 repulsive activity (Chen et al., 2001; Nguyen-Ba-Charvet et al., 2001). The axon of cortical neurons can also be repelled by Slit1 but not by the N-terminal portion of Slit1 (Whitford et al., 2002). To determine whether Slit1 repulsion of OB axons is mediated by the same domain than Slit2, we generated a deletion construct encoding a truncated protein (Slit1-N) lacking EGF repeats 6–9 and the ALPS (for Agrin, Laminin, Perlecan, and Slit) domain (which are cleaved for Slit2-N). Although, our constructs did not allow us to determine whether Slit1 is normally cleaved (but see Whitford et al., 2002), we found that Slit1-N-expressing cells were as efficient as those expressing full-length Slit1 in repelling OB axons (eight of eight cases; 0.204 ± 0.16) (Fig. 2D). Thus, the repulsive activity of both Slit proteins for olfactory axons does not require their C-terminal portion and are probably mediated by similar receptors and signaling pathways.
Absence of chemorepulsive activity from the septum ofSlit1- and Slit2-deficient mice
We next examined whether the previously identified septum-derived repellent activity (Pini, 1993) is mediated by either of these Slit proteins. We used E14 OB explants from actin-GFP transgenic mice (Hadjantonakis et al., 1998) and cultured them in collagen gels near septum explants from E14 wild-type mice, homozygousSlit1-deficient mice, homozygous Slit2-deficient mice, or compound heterozygous or homozygousSlit1/2-deficient mice. OB axons were easily visualized either directly by observing GFP expression or after immunostaining with anti-GFP or anti-βIII-tubulin. As described previously (Pini, 1993; Hu and Rutishauser, 1996), a very robust repulsion was observed with control septum, with OB axons growing away from the septal explants (0.316 ± 0.06, P/D ratio ± SEM from four independent experiments) (Fig.3A). Septum repulsive activity was not significantly changed in Slit1 (0.365 ± 0.11) or Slit2 (0.311 ± 0.15) single heterozygotes (Slit1+/− or Slit2+/−) or in compound heterozygotes (Slit1+/−;Slit2+/−) (0.215 ± 0.12) (Fig. 3D). In contrast with septum explants from homozygous Slit1-deficient mice (Slit1−/−; 0.697 ± 0.19) or Slit2-deficient mice (Slit2−/−; 0.681 ± 0.11), the repulsion was significantly reduced (Fig. 3B,C). Homozygous Slit2-deficient mice are lethal at birth (Plump et al., 2002); therefore, we could mate onlySlit1/2-deficient mice heterozygous for the Slit2deficiency. Unfortunately, in the four different litters used for these experiments, no double homozygous Slit1/2-deficient mice were present. However, the repulsive activity was almost completely abolished in septum explants from embryos in which three of fourSlit1 and Slit2 alleles were mutant, i.e., fromSlit1−/−;Slit2+/− mutant embryos (P/D ratio of 0.81 ± 0.18) or Slit2−/−;Slit1+/− mutant embryos (P/D ratio of 0.787 ± 0.14) (Fig.3E,F), with OB axons growing symmetrically around the explants. Together, these results indicate that the septum-derived repulsive activity for OB axons detectablein vitro involves Slit1 and Slit2 proteins.

Olfactory axons have a reduced repulsion response to septum from Slit mutant mice. Explants of olfactory bulb were dissected from E14–E15 actin-GFP transgenic mice and cultured with septum explants from wild-type (A),Slit1−/− (B),Slit2−/− (C),Slit1+/−;Slit2+/− (D),Slit1−/−;Slit2+/− (E), orSlit1+/−;Slit2−/− (F) E15 embryos. Dotted linesrepresent the position of septum explants. G, Quantitation of the repulsion experiments. The repulsive activity is measured by the axon outgrowth ratio proximal/distal (see Materials and Methods). SEM for each condition is indicated with an error bar.n is the number of olfactory bulb explants. Scale bars: 100 μm.
LOT axon defects in Slit1/2-deficient mice
A role for the septum and Slit proteins in the development of LOT projections in vivo has been questioned recently because, in cultures of whole-telencephalon, the LOT develops normally in the absence of septum or after the addition of Robo1 or Robo2 ectodomains (Sugisaki et al., 1996; Hirata et al., 2001; Patel et al., 2001). Thus, we next analyzed the development of the LOT inSlit1-deficient mice, Slit2-deficient mice, andSlit1/2-deficient mice. We injected DiI in the olfactory bulb of E15 to P1 mice, but most of the injections were done on E15–E16 embryos. In wild-type mice (n = 8) (Fig.4A), the injections lead to the anterograde tracing of a single axon tract that runs rostrocaudally just under the pial surface and corresponds to the LOT (Fig. 4A,F). In homozygousSlit1-deficient (10 of 10 cases) andSlit2-deficient (nine of nine cases) mice, the LOT runs normally as a thick axon bundle, similar to wild type (Fig.4B,E). The same pattern was observed in double heterozygous Slit1/2-deficient mice (20 of 20 cases; data not shown). In contrast, in double homozygousSlit1/2-deficient mice (seven of seven cases), OB axons were still projecting caudally, but the LOT was totally disrupted and multiple axonal fascicles were fanned all over the ventral side of the telencephalon (Fig.4C,D,G), assuming a shape reminiscent of California sea lion whiskers. A mild defect was observed in some Slit1−/−;Slit2+/−mice, with a few defasciculated axons running ventrally (two of six cases; data not shown).

Ventrolateral views of whole-mount brains fromSlit-deficient mice at E15 (A,C–G) or E16 (B) showing the localization of the LOT. Embryos have been injected in the olfactory bulb with DiI. The LOT extends caudally and laterally along the pial surface of the telencephalon, forming a single axonal bundle in wild-type brain (A) but also inSlit1−/− (B) andSlit2−/− (E) mice. In contrast in Slit1−/−;Slit2−/− mutants (C, G), OB axons are defasciculated and form multiple fascicles on the ventral side of the brain.D is a higher magnification of the ventral side of the brain shown in C. Scale bars: A–C,E–G, 350 μm; D, 90 μm.wt, Wild type; S1, Slit1;S2; Slit2.
To determine whether OB axons of Slit1/2-deficient mice reached or even crossed the telencephalic midline, we analyzed vibratome sections of brains injected with DiI in the olfactory bulb and counterstained with Hoechst 33258. As described previously, in wild-type embryos, the LOT was located laterally at an approximately equal distance between the neocortex and the septum (Figs.1A, 5A). In contrast, in Slit1/2-deficient mice, the LOT was enlarged and extended ventrally reaching the septum area (Fig. 5B), with many axons reaching the midline (Fig. 5C).
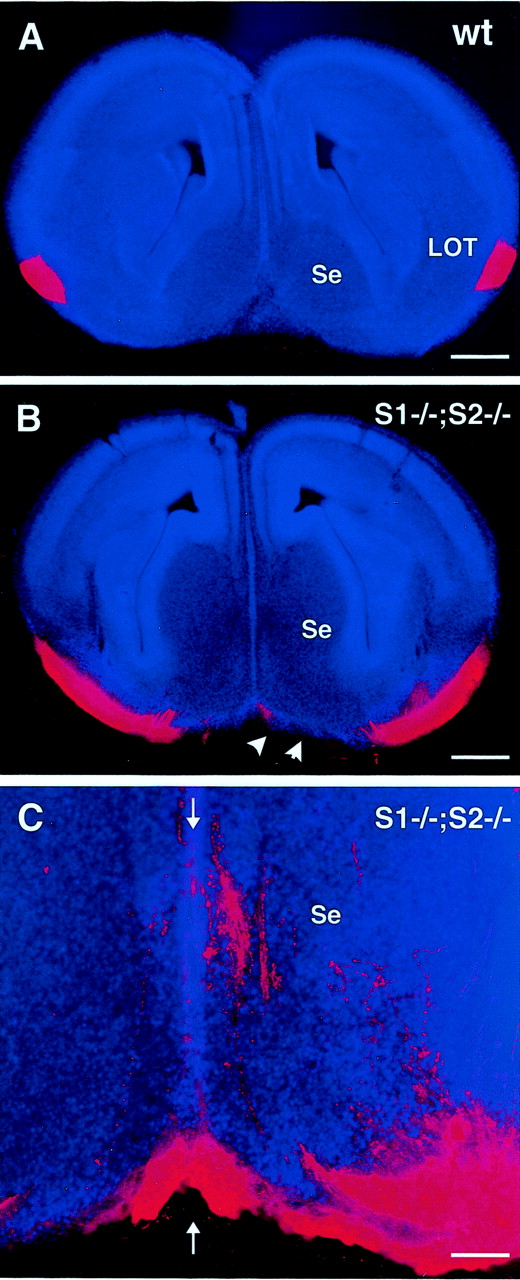
Coronal brain sections from E15Slit1/2-deficient mice injected with DiI in the olfactory bulb. Vibratome sections (100 μm) are counterstained with of Hoechst 33258. The LOT is thick and concentrated just under the pial surface on both ventrolateral sides of the telencephalon in wild-type embryos (A). In Slit1/2-deficient mice (B, C), the LOT isspreading all over the ventral side of the telencephalon. At higher magnification (C), it appears that some OB axons reach the telencephalic midline (arrows) and septum region (Se). Scale bars: A,B, 375 μm; C, 95 μm.wt, Wild type; S1, Slit1;S2, Slit2.
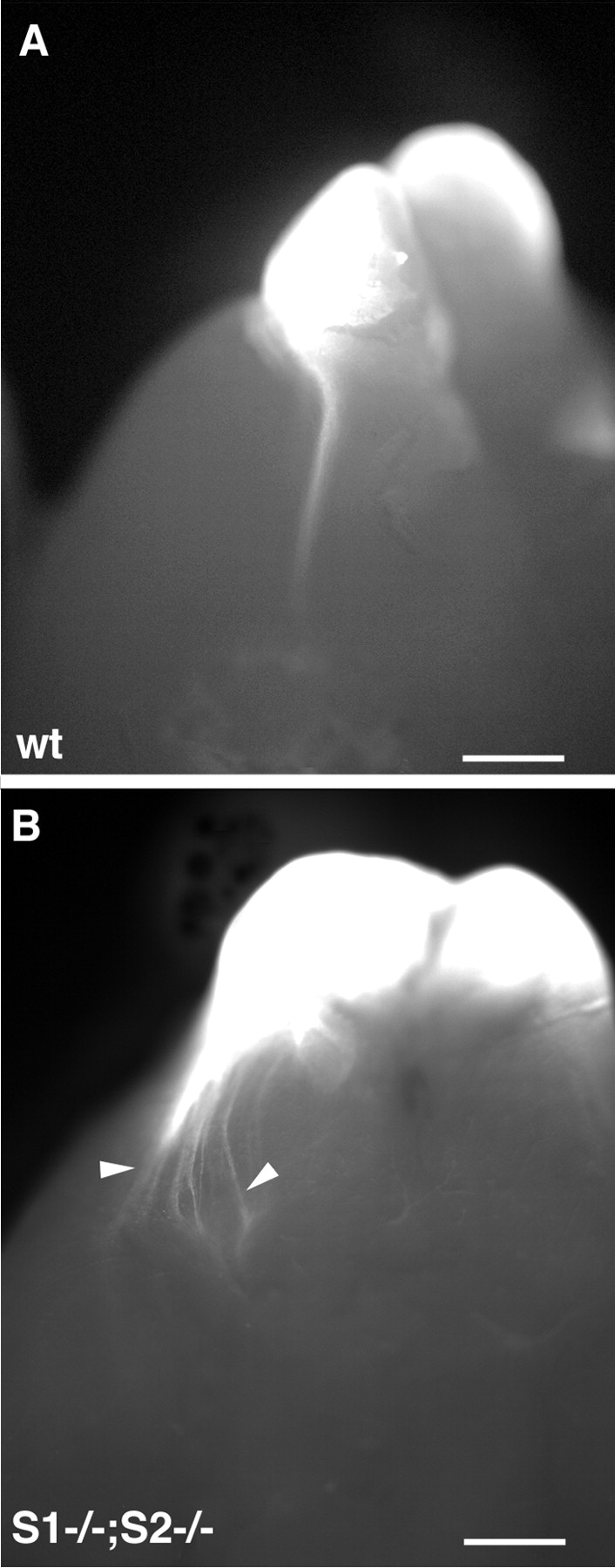
At birth, it was still possible to see the LOT in whole-mount brains using DiI tracing (Fig.6A). This labeling showed that, in P0 Slit1/2-deficient mice, the LOT was very disorganized, similar to E15 embryos (Fig. 6B), and therefore that no correction of this pathfinding defect had occurred.

Ventrolateral views of whole-mount brains from postnatal Slit1/2-deficient mice injected with DiI in the olfactory bulb. At this stage, the LOT is still visible at the pial surface of the telencephalon. In wild-type mice at P1 (A), the axons extend as a single bundle on the lateral side. In double homozygous Slit1/2-deficient mice (B) at P0, LOT axons are still spreading on the ventral side of the brain (arrowheads). Scale bars: 750 μm. wt, Wild type; S1, Slit1;S2, Slit2.
DISCUSSION
The first evidence for chemorepulsion in the vertebrate brain came from studies on the rat olfactory system in which the septum was shown to secrete a diffusible factor repelling OB axons (Pini, 1993). Despite its early discovery, the molecular identity of this septum repellent remained an enigma until recently, when it was found thatSlit2 mRNA is expressed in the septum, embryonic OB output neurons express Robo2 mRNA, and Slit2 protein can repel OB axons in the collagen gel assay (Li et al., 1999; Nguyen-Ba-Charvet et al., 1999). This suggested that Slit2 was the septum repellent. However, some recent data have shown that, in the collagen gel assay, conditioned medium containing human Robo1-Fc or Robo2-Fc is unable to block septum repulsion, although it blocks the repulsive activity of recombinant human Slit1 or Slit2 (Patel et al., 2001). This contrasts with our results, which clearly show that Slits are mediating septum repulsion not only in vitro but also in vivo. It is possible that human Robo-Fc conditioned medium does not efficiently block the endogenous rodent Slit or that, in septum explants, Slits are bound to other molecules or possible cofactors and are not efficiently blocked by human Robo-Fc conditioned medium (see below). In contrast to those in vitro assays, our study is consistent with the septum-derived repellent for OB axons being a combination of Slit1 and Slit2 proteins, which are essential for proper formation of the LOTin vivo. It is possible that in vitro the septum releases other repulsive factors, but they are not sufficient for normal LOT development in vivo. Among the known chemorepulsive molecules for developing axons, Slits were the last to be characterized. They have been shown to repel motor axons, commissural axons upon crossing of the midline (Brose et al., 1999; Zou et al., 2000), several classes of telencephalic axons from the hippocampus (Nguyen-Ba-Charvet et al., 1999), the retina (Erskine et al., 2000; Niclou et al., 2000; Ringstedt et al., 2000), and the neocortex (Shu and Richards, 2001). The detailed analysis of the spatiotemporal expression pattern of Slit genes in rat embryos has shown that they are detected in almost all developing neurons and that neurons frequently coexpress several Slits, for instance in the septum (Nguyen-Ba-Charvet et al., 1999; Marillat et al., 2002). Moreover, the present and previously published results (Patel et al., 2001) indicate that both Slit1 and Slit2 can simultaneously repel OB axons, suggesting that there is some functional redundancy among Slit proteins. This conclusion is supported by the results of the collagen gel cocultures showing that the septum repulsion, although reduced with septum from single Slit1-or Slit2-deficient mice, only disappears with septum from embryos deficient for one Slit and heterozygous for the other one. Likewise, the LOT is normal in Slit1- orSlit2-deficient mice and only shows major defects in doubleSlit1/2-deficient mice, suggesting that the repulsive action of Slit proteins on OB axons is dose dependent. Interestingly, very similar observations have been made recently in other systems. Thus, the analysis of the visual system of Slit1- andSlit2-deficient mice and double Slit1/2-deficient mice has shown that the two Slit gene product act synergistically to control retinal axon guidance, particularly around the region of the developing optic chiasm (Plump et al., 2002) and also in many telencephalic axonal tracts (Bagri et al., 2002).
Although the exact mechanism of Slit function in the development of LOT and retinal axons is still unclear, it seems that, in the telencephalon, Slit proteins play a major role in preventing axons from approaching the midline, a function similar to the one exercised by Slit in Drosophila embryo (Kidd et al., 1999). Among diffusible axon guidance molecules, functional redundancy is not specific to Slits. For instance, hippocampal axons and commissural axons are simultaneously repelled by several secreted semaphorins (Chédotal et al., 1998; Zou et al., 2000; Pozas et al., 2001), and the maxillary attractant for trigeminal sensory axons is composed of two neurotrophins, neurotrophin-3 and brain-derived neurotrophic factor (O'Connor and Tessier-Lavigne, 1999).
The receptor mediating the repulsive action of Slit proteins on OB axons has not been identified. So far, the best candidate is the roundabout family member Robo2, which is known to bind Slit1 and Slit2 and is the only Robo member expressed in embryonic mitral cells (Nguyen-Ba-Charvet et al., 1999; Marillat et al., 2002; A. Chédotal unpublished data). In Drosophila embryos, genetic approaches have directly shown that Robos mediate Slit function (Kidd et al., 1999; Rajagopalan et al., 2000b; Simpson et al., 2000a). Finally, retinal axon guidance defects at the optic chiasm in the zebrafish mutant astray (Fricke et al., 2001; Hutson and Chien, 2002), which is deficient in the robo2 homolog, show similarities to those in Slit1/2-deficient mice (Plump et al., 2002). However, additional extracellular proteins could be involved, such as the glycosylphosphatidylinositol-anchored heparan sulfate proteoglycan glypican-1, which also bind Slits (Liang et al., 1999; Ronca et al., 2001). This is supported by the observation that the repulsive activity of Slit on migrating subventricular zone (SVZ) neuroblasts requires cell-surface heparan sulfate (Hu, 2001). Interestingly, the caudal septum secretes a chemorepulsive factor orienting the migration of SVZ cells in vitro (Hu and Rutishauser, 1996), and Slit1 and Slit2 can mimic this effect (Hu, 1999; Wu et al., 1999). It will be interesting to useSlit1/2-deficient mice to determine whether additional factors are involved and whether Slit repulsive activity has an important role in vivo in the orientation of the migration of SVZ neuroblasts.
In addition to Slits, developing LOT axons are influenced by other long-range factors from the semaphorin family. Mitral cell axons were shown to highly express the semaphorin receptor neuropilin-1. In organotypic cultures, the olfactory epithelium releases a diffusible factor that repels OB axons (de Castro et al., 1999). This repulsive effect is mimicked by the secreted semaphorin Sema3F, whose mRNA is expressed in the olfactory epithelium at the time LOT axons grow. Moreover, OB axons grow preferentially toward aggregates of COS cells secreting Sema3B, suggesting that chemoattraction could also be involved in the formation of the LOT. However, an important role for secreted semaphorins in LOT axon pathfinding in vivo is not supported by the analysis of mice deficient for their receptors neuropilin-1 (Kitsukawa et al., 1997) or neuropilin-2 (Chen et al., 2000), which have a normal LOT.
Several studies have shown that short-range cues are involved in the pathfinding of LOT axons from the OB to the olfactory cortex. These cues are thought to be at the surface of specialized cells, so-called LOT cells or “guide post cells” by analogy with insects. These cells are stained by the monoclonal antibody LOT1, which recognizes only neurons around the LOT position (Sato et al., 1998). LOT cells are generated at approximately E10.5 in the neocortex and migrate tangentially and ventrally to the future location of the LOT (Tomioka et al., 2000). 6-OHDA ablation of LOT cells prevents the formation of LOT in organotypic cultures (Sato et al., 1998). However, LOT cues seem to play a role in the positioning of LOT axons in the LOT pathway but lack directional (rostrocaudal) information (Sugisaki et al., 1996). In a recent report, it was proposed that the septum repellent and Slit proteins are not essential to guiding LOT axons, and they only act in parallel with LOT cells. Thus, in whole telencephalon cultures, the LOT is normally positioned even in the absence of septum or when a Robo1 ectodomain is added to the culture medium. This contrasts with our results, which clearly show that long-range guidance by Slit molecules is preponderant over short-range cues for LOT axon pathfinding. As suggested by Hirata et al. (2000), one major drawback of the Robo blocking experiments could be that endogenous Slit was not abolished because of a limited diffusion of Robo proteins in the cultured telencephalic hemispheres. It will be interesting to determine whether the expression of other putative guidance cues for LOT axons, in particular LOT1, is perturbed in Slit1- andSlit2-deficient mice, to address the question of the physiological role of LOT short-range cues in vivo.
In conclusion, the detailed molecular mechanism of LOT formation is very complex and far from being fully understood. For instance, the factors controlling the growth of LOT axons along the rostrocaudal axis are still unknown. This behavior is not affected inSlit1/2-deficient mice. Similarly, LOT axons are known to wait (for ∼2 d in the mouse) before sending collateral branches to the olfactory cortex (Hirata and Fujisawa, 1999), which they invade in a precise rostrocaudal order (Devor, 1976; Derer et al., 1977; Scott et al., 1980; Luskin and Price, 1982; Schwob and Price, 1984; Shipley and Ennis, 1996). These collateral branches are the only connections of mitral and tufted cell axons with the olfactory cortex. Anosmin-1, a secreted protein, defective in the X chromosome-linked form of Kallmann syndrome, has been shown recently to promote the branching of OB axons (Soussi-Yanicostas et al., 2002). However, Slit proteins are also possible candidates because Slit2 regulates the branching of sensory axons from the dorsal root ganglia (Wang et al., 1999) and Slit1 the branching of pyramidal cell dendrites in the neocortex (Whitford et al., 2002). This possible branching activity of Slits on OB axons is difficult to assess in the Slit1/2-deficient mice because the growth of LOT axons is very perturbed. Our results also indicate that some guidance cues act on all LOT axons, but a recent study (Zou et al., 2001) has shown clearly that the projection from the OB to the olfactory cortex is topographically organized and that mitral cells innervated by olfactory receptor neurons from distinct zones of the olfactory epithelium (expressing distinct olfactory receptors) project to distinct regions in the olfactory cortex. The factors that control the formation of this complex projection map are completely unknown.
Footnotes
This work was supported by Institut National de la Santé et de la Recherche Médicale (A.C. and K.N.-B.-C.), French Ministry for Research Action Concertée Incitative Grant ACI 5116, and Association pour la Recherche sur le Cancer Grant 5249 (A.C.). K.N.-B.-C. was funded by the Fondation de France and the Fondation de la Recherche Médicale. A.P. was funded by National Institute of Neurological Disorders and Stroke Grant K08, a Howard Hughes Medical Institute (HHMI) Physician Scientist Award, and the Sarnoff Society. M.T.L. is an investigator of the HHMI.
Correspondence should be addressed to Alain Chédotal, Institut National de la Santé et de la Recherche Médicale U106, Bâtiment de Pédiatrie, Hôpital de la Salpêtrière, 47 Boulevard de l'Hôpital, 75013 Paris, France. E-mail: chedotal{at}infobiogen.fr.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}