

Abstract
Most unmyelinated nociceptive neurons that mediate pain and temperature sensation from the skin bind isolectin B4 (IB4)-lectin and express Ret, the common signaling component of glial cell line-derived neurotrophic factor (GDNF) family. One of these factors, neurturin, is expressed in the epidermis, whereas its GDNF family receptor α2 (GFRα2) is expressed in the majority of unmyelinated Ret-positive sensory neurons. However, the physiological roles of endogenous neurturin signaling in primary sensory neurons are poorly understood. Here, we show that the vast majority (∼85%) of IB4 binding and P2X3 purinoreceptor-positive neurons, but virtually none of the calcitonin gene-related peptide (CGRP) or vanilloid receptor transient receptor potential vanilloid 1-positive neurons in mouse dorsal root ganglion (DRG) express GFRα2. In GFRα2 knock-out (KO) mice, the IB4-binding and P2X3-positive DRG neurons were present but reduced in size, consistent with normal number but reduced caliber of unmyelinated axons in a cutaneous nerve. Strikingly, nonpeptidergic (CGRP-negative) free nerve endings in footpad epidermis were >70% fewer in GFRα2-KO mice than in their wild-type littermates. In contrast, the density of CGRP-positive epidermal innervation remained unaffected. In the formalin test, the KO mice showed a normal acute response but a markedly attenuated persistent phase, indicating a deficit in inflammatory pain response. Behavioral responses of GFRα2-KO mice to innocuous warm and noxious heat were not blunted; the mice were actually markedly hypersensitive to noxious cold in tail immersion test. Overall, our results indicate a critical role for endogenous GFRα2 signaling in maintaining the size and terminal innervation of the nonpeptidergic class of cutaneous nociceptors in vivo.
- cold hyperalgesia
- cutaneous C-fiber innervation
- formalin test
- IB4-lectin
- neurturin
- noxious heat sensation
Introduction
Cutaneous nociceptors are polymodal primary afferent neurons, the mainly unmyelinated (C-fiber) axons of which terminate as free nerve endings between the epidermal keratinocytes to mediate thermal, mechanical, and pain sensation from the skin. Most of these small sensory neurons are selectively labeled by isolectin B4 (IB4) and express the Ret receptor tyrosine kinase (Plender-leith and Snow, 1993; Molliver et al., 1997; Bennett et al., 1998b). Ret is the common signaling component of glial cell line-derived neurotrophic factor (GDNF) family ligands that bind a GDNF family α-receptor (GFRα) (Baloh et al., 2000; Airaksinen and Saarma, 2002). GFRα1, GFRα2, and GFRα3, the receptors for GDNF, neurturin (NRTN), and artemin, respectively, are expressed in partially overlapping subpopulations of Ret-positive dorsal root ganglion (DRG) neurons (Bennett et al., 1998b). Ret and GFRα receptor expression in DRG starts during embryonic development and continues to adulthood (Golden et al., 1999; Baudet et al., 2000). Of the GDNF family ligands, NRTN mRNA is known to be expressed in embryonic skin epidermis as well as in adult skin (Luukko et al., 1998; Golden et al., 1999), suggesting that NRTN acts as a physiological target-derived factor for the development, maintenance, and/or function of the cutaneous Ret/GFRα2-positive sensory neurons.
Although exogenous NRTN promotes the survival and neurite outgrowth of DRG neurons in vitro (Kotzbauer et al., 1996; Rossi et al., 1999; Paveliev et al., 2004), GFRα2 knock-out (KO) mice have normal numbers of sensory axons in the saphenous nerve (Stucky et al., 2002), suggesting that the primary sensory neurons do not need GFRα2 signaling for survival or axon growth into the peripheral target area. Yet, whether the peripheral target innervation of the neurons (i.e., free nerve endings in the skin epidermis) is affected in these mice has not been investigated. Previous electrophysiological studies using acute DRG neuron cultures and skin–nerve preparations from adult GFRα2-KO mice suggest that GFRα2 signaling regulates noxious heat transduction but not mechanical response properties of the IB4-binding subpopulation of unmyelinated C-fiber sensory neurons (Stucky et al., 2002). However, a potential physiological role of endogenous GFRα2 signaling in somatosensory neuron function in vivo has not been studied. Here, we address the role of GFRα2 signaling in DRG neuron morphology, innervation, and function in mice. We show that GFRα2 is expressed predominantly by the unmyelinated IB4-binding subpopulation of mouse DRG neurons and that their size but not number is reduced in GFRα2-KO mice. We found that the KO mice have a profoundly reduced density of calcitonin gene-related peptide (CGRP)-negative free nerve endings in footpad skin and a markedly attenuated formalin-induced inflammatory pain response. Surprisingly, temperature sensitivity of the mice in different behavioral tests was either normal or increased. Our results indicate that endogenous NRTN signaling via GFRα2 is needed to maintain the size, inflammatory pain responses, and terminal innervation of small nonpeptidergic cutaneous neurons. A preliminary report of this work has been published (Hiltunen et al., 2003).
Materials and Methods
Animals.
The local ethics committee for animal research at the University of Helsinki approved all animal experiments. GFRα2-KO mice were produced and genotyped as described previously (Rossi et al., 1999). The mutation was maintained by backcrossing heterozygous animals for at least 10 generations separately for two inbred strains (129S2/SvHsd and C57BL/6JOlaHsd; Harlan, Bicester, UK), resulting in congenic B6 and 129 strains. Littermate wild-type (WT) and GFRα2-KO mice in (129 × B6) F1 hybrid (129B6) backgrounds were obtained by intercrossing heterozygotes from the two congenic lines (Banbury Conference on Genetic Background in Mice, 1997).
Immunohistochemistry.
Littermate mice of the same gender were perfused transcardially with 4% paraformaldehyde in PBS under deep anesthesia and postfixed overnight. For P2X3 purinoceptor staining, some mice were perfused with calcium-free Tyrode’s solution, followed by 4% paraformaldehyde and 0.2% picric acid in 0.1 m phosphate buffer, pH 6.9. Lumbar L3 and L4 ganglia, a length of lumbar spinal cord, and forepaw and hindpaw skin including the footpads were dissected, cryoprotected in sucrose, and cut into cryosections. The slides were incubated with specific polyclonal primary antibodies and biotin-labeled IB4 lectin (Sigma, St. Louis, MO) overnight at room temperature. Primary antibodies were against CGRP (rabbit; Chemicon, Hampshire, UK), GFRα2 (goat; R & D Systems, Abingdon, UK), P2X3 (guinea pig; Chemicon), protein gene product 9.5 (PGP9.5; rabbit; Chemicon), peripherin (rabbit; Chemicon), and transient receptor potential vanilloid 1 (TRPV1) (rabbit; a gift from Dr. Julius, University of California, San Francisco, CA). Cyanine 3 (Cy3)- and Cy2-conjugated streptavidin and donkey secondary antibodies were obtained from Jackson ImmunoResearch (Cambridgeshire, UK). The embedding medium included 4′,6-diamidino-2-phenylindole (DAPI) to aid nuclear identification. Histological quantification was performed by an experimenter who was blind to the genotype of the samples.
Neuronal counts, colocalization, size distribution, and densitometry.
To estimate the density of P2X3-positive neurons in lumbar DRG, we used the unbiased Abercrombie Method 2 (http://www.nervenet.org/papers/Aberfp.html#Method2) for sections double-stained with P2X3 and the pan-neuronal marker PGP9.5. Briefly, immunopositive neuronal profiles (with a clear nucleus) were counted in every second pair of adjacent 10 and 20 μm sections cut through the L4 ganglion. Subtraction of the sum of 10 μm section counts from the sum of 20 μm section counts results in an unbiased estimate of neuron number in the ganglion from which the average density of P2X3-positive neurons (relative to total PGP9.5-positive neurons) was derived. Colocalization of GFRα2 with different DRG neuronal markers was estimated from four to eight sections per L4 ganglia from three to four wild-type mice using systematic random sampling. All GFRα2-positive profiles with a clear nucleus were analyzed from the images, and the percentage of colocalization was calculated. For soma size analysis in L4 DRG, at least 100 labeled neuronal profiles per mouse and marker were selected randomly and outlined from digital images using ImageProPlus software (Media Cybernetics, Wokingham Berkshire, UK). The intensity of IB4-binding in wild-type and GFRα2-KO lumbar spinal cords (level L3-L5) was estimated from several digital images using AIDA/2D densitometry software (Raytest Isotopengeräte, Straubenhardt, Germany). The results are given as intensity per area (in arbitrary units/pixel) after subtraction of background.
Epidermis innervation.
Adjacent transverse or horizontal (15–30 μm) cryosections of interdigital footpads were stained for the pan-neuronal marker PGP9.5 and for CGRP. The average thickness of footpad epidermis did not differ between the genotypes (data not shown). To estimate the density of free nerve endings in the epidermis, digital images of randomly selected areas from the interdigital footpad skin were taken at 40× magnification, and the number of immunopositive nerve fibers in the epidermis within each image area was counted by carefully focusing throughout the section thickness to identify all of the fibers. The dermis-epidermis borderline within the image area was traced and its length measured using the ImageProPlus software. At least 10 areas were analyzed for each footpad and marker. Similar results were obtained from the forepaws and hindpaws, so the results were pooled. The average number of PGP9.5- and CGRP-positive fibers per unit length of epidermis was then calculated for each animal. To estimate the percentage of peptidergic (CGRP-positive) and nonpeptidergic (CGRP-negative) free nerve endings in the epidermis, 20 μm footpad sections from a separate set of animals were double stained for PGP9.5 (sheep polyclonal; Serotec, Oxford, UK) and CGRP. One hundred randomly selected PGP9.5-positive fibers were analyzed per animal. Confocal microscopy images were taken using a 40× water immersion objective, and maximum intensity projections were generated using Zeiss (Thornwood, NY) LSM5 software.
Behavioral studies.
Littermate mice (of both genders), 3–6 months of age, were used in all behavioral tests. In addition to mice in a 129B6 hybrid background, congenic mice in B6 and 129 backgrounds were used as indicated. The mice were housed in groups of two to four per cage in standard specific pathogen-free conditions under a constant dark-light cycle. Food and water were provided ad libitum. Ambient room temperature was a constant 22.0 ± 0.5°C. The experimenter was blind to the genotype of the mice during testing.
Hot plate test.
A standard hot plate (TSE, Bad Homburg, Germany), adjusted to 49.0 or 52.0°C (±0.2), was used to assess nociceptive sensitivity. The mouse was confined on the plate by a Plexiglas cylinder (diameter, 19 cm; height, 26 cm), and the latency to the first hindpaw response (licking or shaking) was recorded.
Tail withdrawal test.
The mice were gently restrained by hand, and the distal half of the tail was inserted into a water bath. The latency to withdraw the tail was recorded. The test was repeated four times with 1 h intertrial intervals at each temperature. The water temperature was controlled either thermostatically (49 and 52°C) or by a digital thermometer (4°C; bath surrounded with ice).
Temperature choice test.
The animals were tested in a two-temperature choice (thermotaxis) test modified from Lee et al. (2005) and Moqrich et al. (2005). The testing apparatus consists of two square aluminum plates (20 × 20 × 2 cm thermally isolated with a 2-mm-thick plastic spacer) enclosed within a Plexiglas chamber. The temperature of the plate was controlled with a block heater (UBD2; Grant Instruments, Cambridge, UK) and measured with a digital thermometer. One plate was kept at room temperature (22°C), and the other was heated to 35°C. Alternatively, one plate was heated to 34°C and the other to 36°C. Mice were placed in the chamber and allowed to acclimate to the testing environment for 20 min, and the total time spent on the warm surface during the following 10 min was recorded with a video system (EthoVision, Noldus, The Netherlands).
Mechanosensitivity.
A separate set of 2- to 4-month-old mice (in B6 background) were tested for behavioral responses to mechanical stimuli using calibrated von Frey nylon filaments (tip diameter, 0.8 mm). The mice were placed in an elevated plastic cage on a wire mesh floor. Von Frey hairs were applied from below to the plantar surface of both hindpaws in ascending order of stimulus force (0.28, 0.39, 0.65, 1.17, 1.88, and 3.29 g). At threshold, the mouse withdraws its paw away from the hair. Both hindpaws were tested 10 times, and the interval between two consecutive trials on the same paw was at least 1 min. The results are presented as the average percentage of responses against a given stimulus force.
Formalin test.
Formalin-induced pain behavior was assessed as described previously (Tjolsen et al., 1992; Mogil et al., 1998) using a short anesthesia during the injection, which has been reported to reduce variability (Murray et al., 1988). Mice (4- to 6-month-old littermates in B6 background) were allowed to acclimate for 15 min in a plastic box, anesthetized briefly with isoflurane vapor (in a glass container), and injected subcutaneously with 20 μl of 1.85% formaldehyde in 0.9% saline (corresponds to 5% formalin) into the plantar right hindpaw. The animals woke up within 1 min (average, 15–30 s) after the injection. The mice were observed for 60 min after the formalin injection, and the time spent licking or shaking the injected paw was measured (with a stop watch) in 5 min intervals. The acute phase (phase 1) was defined as 0–10 min after injection, and the persistent (tonic) phase (phase 2) was defined as 10–60 min after injection. Formalin-induced footpad edema was estimated by comparing injected and noninjected (contralateral) paw thickness and weight as described by Mogil et al. (1998). Briefly, 60 min after the formalin injection, animals were given a lethal dose of anesthetic, and the dorsoventral thickness of both hindpaws was measured with a micrometer caliper from the middle of the hindpaw. After the animals were killed, the hindpaws were cut at the ankle joint and weighed on a microbalance.
Data are expressed as mean ± SEM. Statistical analysis between genotypes was done either using unpaired Student’s t test assuming unequal variances or with one-way ANOVA with genotype as a grouping variable, as appropriate. The size distribution data were analyzed using Kolmogorov–Smirnov test.
Results
Most IB4-lectin binding and P2X3 receptor-positive neurons express GFRα2
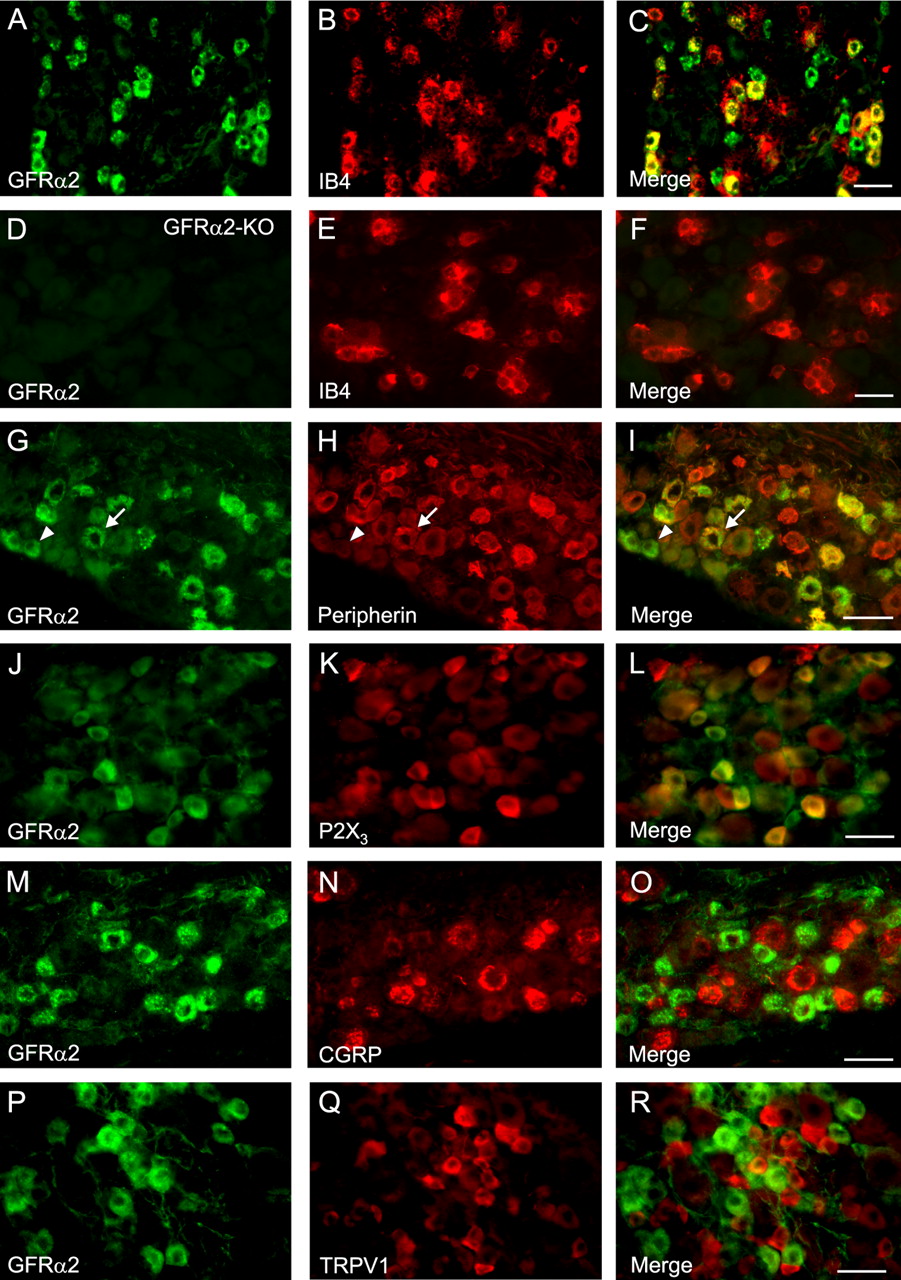
To estimate the degree of GFRα2 colocalization with various markers of DRG subpopulations in mice, we performed quantitative immunohistochemical analysis of DRG sections (Table 1). In mouse lumbar DRGs, 83% of the IB4-binding neurons expressed GFRα2, whereas of the GFRα2-positive neurons, 71% bound IB4 (Fig. 1A–C). Of GFRα2-positive neurons, 68% colocalized with peripherin, a marker for unmyelinated neurons, and 64% of the unmyelinated, peripherin-positive neurons expressed GFRα2 (Fig. 1G–I). For comparison, ∼15% of the GFRα2-positive neurons in the rat express a marker for myelinated neurons (Bennett et al., 1998b). The average size of GFRα2-expressing neuronal profiles that were peripherin-positive (380 ± 9 μm2) was not significantly different from those that were peripherin-negative (344 ± 11 μm2), suggesting that the peripherin-negative GFRα2-neurons represent thinly myelinated nociceptors. Most (85%) of the P2X3-expressing neurons were GFRα2-positive, and vice versa, 66% of GFRα2-positive neurons expressed P2X3 (Fig. 1J–L). In contrast, few (1.5%) of the GFRα2-positive neurons coexpressed CGRP (Fig. 1M–O). Moreover, GFRα2 was not colocalized with the vanilloid receptor TRPV1 in mouse DRG neurons (Fig. 1M–O). In summary, a large majority of the small nonpeptidergic neurons in mouse DRG express GFRα2.
Percentage of colocalization of GFRα2 with various markers in mouse lumbar (L3 and L4) DRGs

Colocalization of GFRα2 protein with markers of unmyelinated neurons in mouse DRG. Sections of wild-type (A–C, G–R) and GFRα2-KO (D–F) mouse lumbar L4 DRG are stained for IB4-binding (B, E), peripherin (H), P2X3 (K), CGRP (N), and TRPV1 (Q) to estimate colocalization with GFRα2 protein (left; green). The yellow color in the merged images (right) indicates colocalization. No specific GFRα2 staining is seen in the DRG section from the GFRα2-KO mouse (D). Note that GFRα2-expressing neurons that are peripherin negative (arrowheads) appear to be similarly sized to neurons that are peripherin positive (arrows). Scale bars, 50 μm.
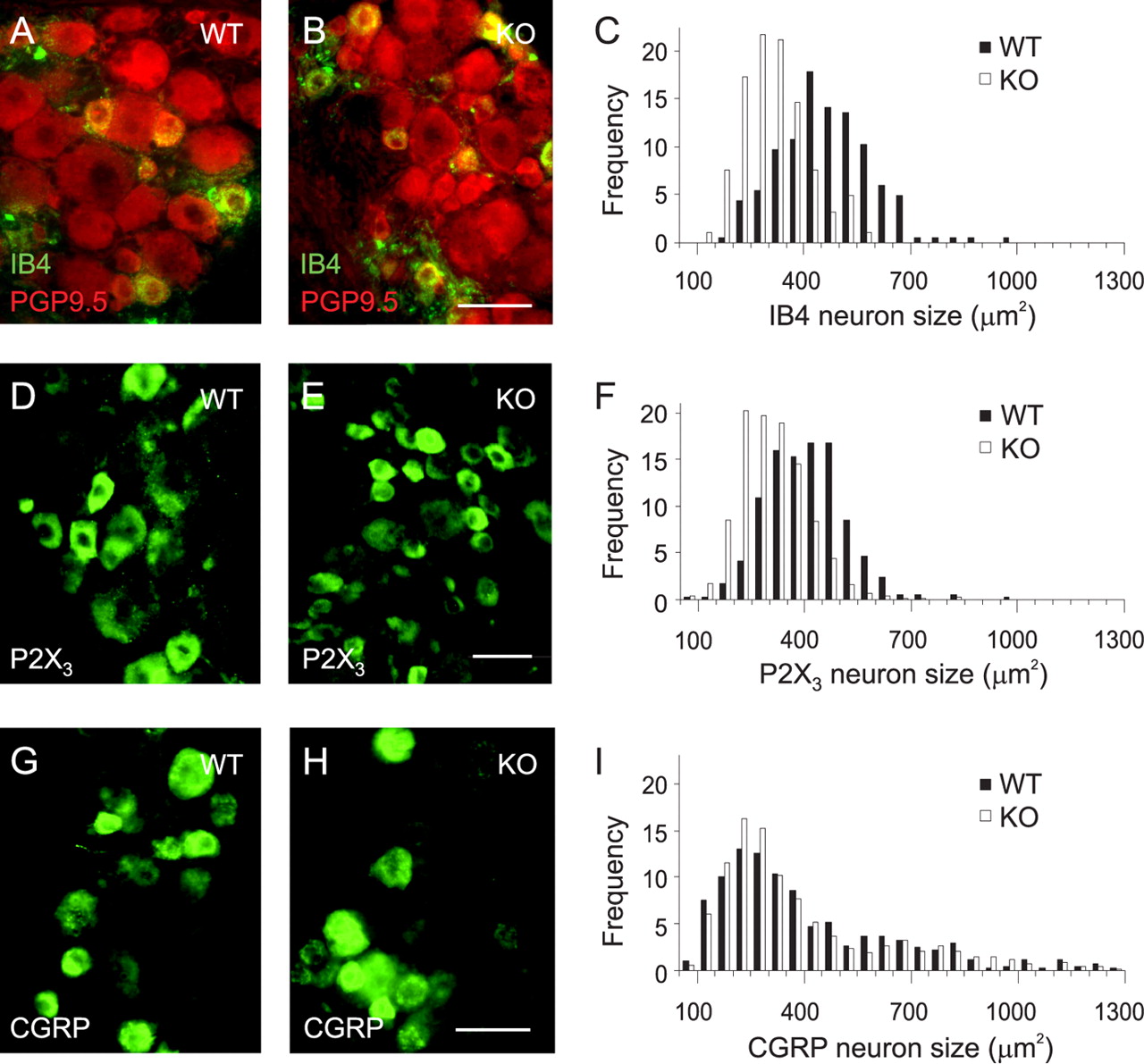
IB4-binding and P2X3-positive neurons are smaller in GFRα2-KO mice
The density of P2X3-positive neurons in lumbar DRGs was similar in wild-type and GFRα2-KO mice (WT, 47 ± 7%; KO, 45 ± 4% of PGP9.5-positive neurons in L4 DRG; n = 4 mice for both genotypes) consistent with identical numbers of unmyelinated axons in cutaneous nerves and IB4-binding neurons in acute DRG cultures isolated from wild-type and GFRα2-KO mice (Stucky et al., 2002). However, the IB4-binding (WT, 463 ± 10 μm2; KO, 316 ± 7 μm2; p < 0.0001) (Fig. 2A–C) and P2X3-positive (WT, 407 ± 6 μm2; KO, 309 ± 4 μm2; p < 0.0001) (Fig. 2D–F) DRG neuronal profiles were markedly smaller in GFRα2-KO mice than in their wild-type littermates, compatible with the smaller caliber of unmyelinated nerve fibers in the KO mice (Stucky et al., 2002). These results agree with the reduced size of sensory neurons expressing GFRα2 mRNA in NRTN-deficient mice (Heuckeroth et al., 1999). In apparent contrast to our data, the number of GFRα2-expressing DRG neurons is also reduced in the NRTN-deficient mice, but whether this is attributable to cell death or downregulation of GFRα2 expression remains unclear. As expected from the lack of GFRα2 expression in CGRP-positive DRG neurons in wild-type mice, the CGRP-positive neuronal profiles in GFRα2-KO mice were of normal size (WT, 411 ± 13 μm2; KO, 390 ± 9 μm2; p = 0.12) (Fig. 2G–I).

IB4-lectin binding and P2X3-positive neurons are smaller in GFRα2-KO mice. Representative images of IB4-binding (A, B), P2X3-positive (D, E), and CGRP-positive (G, H) neurons in adult lumbar DRG from wild-type (A, D, G) and GFRα2-KO (B, E, H) mice, which were used to count the neurons and to generate the size distribution histograms (C, F, I), are shown. Note the atrophy of IB4-binding and P2X3-positive but not CGRP-positive neurons in GFRα2-KO mice. Scale bars, 50 μm.
Many CGRP-negative free nerve endings are missing in GFRα2-KO mouse footpad epidermis
Free nerve endings in the glabrous footpad skin were visualized using antibodies against CGRP and pan-neuronal marker PGP9.5. Almost all nerve fibers in the epidermis are unmyelinated (either CGRP-positive or nonpeptidergic) free nerve endings (Zylka et al., 2005). Fiber density and labeling intensity of the subepidermal nerve plexus in the footpad dermis appeared similar between the genotypes (Fig. 3A–E). In contrast, the density of PGP9.5-positive nerve fibers in the epidermis appeared clearly reduced in GFRα2-KO mice compared with wild-type mice (Fig. 3A,B), whereas the density of CGRP-positive nerve fibers appeared similar between the genotypes (Fig. 3D,E) (data not shown). Quantification of the fiber density per unit length indicated that the epidermal density of CGRP-positive nerve fibers was unchanged in GFRα2-KO mice (Fig. 3C) (p = 0.2; n = 4 for both genotypes). In contrast, there was ∼70% less PGP9.5-positive fibers in epidermis per unit length in KO mice than in wild-type mice (Fig. 3C) (p < 0.001 between genotypes; n = 8 for both).

Loss of PGP9.5-positive but not CGRP-positive free nerve endings in the footpad epidermis in GFRα2-KO mice. A, B, Conventional microscopic images of sections through the forepaw footpad skin from wild-type (A) and GFRα2-KO (B) mice stained for pan-neuronal marker PGP9.5. C, Quantification of PGP9.5- and CGRP-positive nerve fibers per unit length of epidermis indicates profound loss of PGP9.5 fibers (***p < 0.001; n = 8 mice in both genotypes), whereas the density of CGRP fibers is unchanged (n = 4 mice in both genotypes). Error bars represent SEM. D, E, Confocal microscopic images from sections through the footpad skin from wild-type (D) and GFRα2-KO (E) mice double stained for pan-neuronal marker PGP9.5 (green) and CGRP (red). Nuclei are counterstained with DAPI (blue). Arrowheads indicate PGP9.5 and CGRP double-positive fibers. Scale bars, 20 μm.
Because direct identification of the nonpeptidergic fibers in skin is difficult (Zylka et al., 2005), we quantified the extent of colocalization of CGRP within the PGP9.5-positive fibers in the footpad epidermis from additional wild-type and KO mice (n = 3 for both genotypes) as an independent estimate of the fiber loss. As expected, all CGRP fibers were PGP9.5 positive (Fig. 3D,E). In the wild-type mouse epidermis, CGRP was expressed by 27% of PGP9.5-positive nerve endings, whereas in the KO mouse epidermis, 56% of the remaining PGP9.5-positive nerve fibers expressed CGRP. From this, assuming that the CGRP-positive fibers are unaffected, it can be estimated that 52% of all PGP9.5-positive nerve fibers were missing in the KO epidermis. [For 27 CGRP-positive fibers in KO mice, 21 (44% of 27 + 21) fibers are CGRP negative, and thus 52 (= 100 − 27 − 21) fibers are lost.] This corresponds to a loss of 71% (= 21/73 − 1) of the CGRP-negative nerve fibers. Together, the results show that the vast majority of nonpeptidergic (and ∼50–70% of all) free nerve endings are missing in the GFRα2-KO mouse footpad epidermis.
To study whether GFRα2 signaling is critical for the spinal projections of nonpeptidergic neurons (Snider and McMahon, 1998; Hunt and Mantyh, 2001), we analyzed lumbar spinal cord sections for IB4 binding. The density of IB4 binding in inner lamina II of dorsal horn was comparable between the genotypes (WT, 2.8 ± 0.1; KO, 3.0 ± 0.5 arbitrary units; p = 0.6) indicating that central target innervation of the nonpeptidergic neurons is not affected in GFRα2-KO mice (Fig. 4).

IB4 binding is unaltered in the GFRα2-KO spinal cord. Cryosections of wild-type (A) and GFRα2-KO (B) mouse spinal cords were labeled with IB4 (green) and CGRP (red). Scale bar, 100 μm.
GFRα2-KO mice show normal behavioral responses to innocuous warm in the temperature choice test and to noxious heat in the hot plate test but are hypersensitive to hot and cold in the tail immersion test
Neurons from GFRα2-KO mice have a defect in heat transduction in vitro (Stucky et al., 2002). To assess noxious temperature sensitivity in vivo, we first used the standard hot plate and tail withdrawal tests of noxious pain. In the hot plate test, no significant difference was present in the escape latency between GFRα2-KO and wild-type mice in a 129B6 hybrid background at plate temperatures of 52°C (Fig. 5A) or 49°C (data not shown). Similar results were obtained using mice in congenic B6 (Fig. 5A) and 129 (data not shown) backgrounds. In the tail immersion test, withdrawal latency at a water bath temperature of 52°C was similar between the genotypes (Fig. 5B). However, at a water temperature of 49°C, the GFRα2-KO mice exhibited significantly shorter withdrawal latencies than the wild-type controls (Fig. 5C). Moreover, the mutant mice exhibited much shorter latencies in the cold water (4°C) tail withdrawal test (Fig. 5D). To assess sensitivity to innocuous warm temperatures, we used the two-temperature choice test. In accord with a previous report (Lee et al., 2005), wild-type mice did not distinguish between the temperatures of 34 and 36°C, whereas they clearly preferred 35°C over room temperature (Fig. 5E). However, GFRα2-KO mice did not differ from their wild-type littermates in this test.

Behavioral analysis of cutaneous sensory responses in GFRα2-KO mice. Results using mice in 129B6 hybrid and in congenic B6 or 129 backgrounds are shown. The number of mice in each test is shown in parentheses. A, B, GFRα2-KO mice do not differ from their wild-type littermates in the 52°C hot plate (A; 129B6, F(1,70) = 1.26, p = 0.3; B6, F(1,20) = 0.21, p = 0.7) or tail-withdrawal tests (B; 129B6, F(1,23) = 0.17, p = 0.7; B6, F(1,20) = 0.73, p = 0.4). C, D, Response latencies in GFRα2-KO mice are shorter than in their wild-type littermates at 49°C (C; 129B6, F(1,48) = 5.15, *p = 0.03; 129, F(1,12) = 5.49, *p = 0.04) and 4°C (D; 129B6, F(1,23) = 42.58, ***p < 0.001; B6, F(1,20) = 7.95, **p = 0.01). E, GFRα2-KO mice do not differ from their wild-type littermates in responses to the warm two-temperature choice test (36 vs 34°C, F(1,14) = 0.05, p = 0.8; 35 vs 22°C, F(1,14) = 0.24, p = 0.6). F, The second but not the first phase of formalin-induced pain behavior is impaired in GFRα2-KO mice. Hindpaw licking and shaking responses in wild-type (n = 10) and GFRα2-KO mice (n = 9) in a B6 background after intraplantar injection of 5% formalin are shown. ***p = 0.0001. Error bars represent SEM.
GFRα2-KO mice show normal behavioral responses to tactile stimulation
We used calibrated von Frey filaments of increasing force to evoke paw withdrawal but did not observe any difference between the genotypes (supplemental material, available at www.jneurosci.org). This is consistent with the previous report using an in vitro skin-nerve preparation that neurons from GFRα2-KO mice show normal electrophysiological responses to mechanical stimulation (Stucky et al., 2002).
Formalin-induced persistent pain is reduced in GFRα2-KO mice
We measured nociceptive responses to chemogenic inflammatory pain using the formalin test (Murray et al., 1988) (Fig. 5F). Subcutaneous injection of formalin causes local tissue damage and acute activation of both myelinated and unmyelinated nociceptors (Puig and Sorkin, 1995). During the first acute phase, both wild-type and GFRα2-KO mice spent equally long licking and shaking the injected paw (WT, 72 ± 11 s; KO, 56 ± 9 s; p = 0.3). However, in the second persistent phase, the formalin-induced pain behavior was significantly reduced in KO mice compared with their wild-type littermates (WT, 330 ± 30 s; KO, 140 ± 20 s; p = 0.0001). Formalin-induced swelling of paws was similar between the genotypes (data not shown), indicating that the degree of inflammation was comparable.
Discussion
In this study, we show that nonpeptidergic C-fiber nociceptive neurons require GFRα2 for their size and cutaneous target innervation. We also demonstrate that GFRα2-deficiency in mice results in specific abnormalities in sensory tests.
We show that the average size of IB4-binding and P2X3-positive DRG neurons is reduced (by ∼30%) in GFRα2-KO mice in vivo, consistent with the smaller caliber of unmyelinated axons in a cutaneous nerve in KO mice (Stucky et al., 2002). We assume that the shift in cell size distribution in the GFRα2-KO mice represents general atrophy of the nonpeptidergic DRG neurons that express GFRα2 in wild-type mice. However, it cannot be excluded that this shift is attributable to a more dramatic atrophy of only a subset of large nonpeptidergic neurons. The density of P2X3-positive neurons is not reduced in GFRα2-KO mice, consistent with our previous report of a normal number of IB4-binding neurons and unmyelinated axons in the cutaneous nerve (Stucky et al., 2002). Thus, similar to many autonomic cholinergic neurons (Hiltunen and Airaksinen, 2004), GFRα2 signaling is required for cell size but not survival of a major subpopulation of cutaneous primary sensory neurons.
The most important result of this study is the profound (>70%) reduction of nonpeptidergic epidermal nerve endings in adult GFRα2-KO mouse footpad. The epidermis receives robust innervation by small-caliber (mainly C- but also Aδ) fibers, the free nerve endings of which mediate thermal, mechanical, and pain sensation. Approximately half of the C-fiber nociceptors are CGRP positive, but only a subpopulation of them project to skin. Our estimate that ∼30% of all PGP9.5-positive nerve fibers in wild-type mouse foot pad epidermis are CGRP positive is comparable with another recent study (Zylka et al., 2005). The peptidergic class of cutaneous nociceptor expresses tyrosine receptor kinase A and requires its ligand nerve growth factor (NGF) postnatally from the skin for terminal arborization and sensitivity to thermal and chemical stimuli (Bennett et al., 1998a). As shown here, the peptidergic neurons do not express GFRα2 and remain unaffected in GFRα2-KO mice. Thus, our results indicate that most nonpeptidergic cutaneous sensory neurons require GFRα2 signaling for target innervation. The ability to genetically label this innervation (Zylka et al., 2005) should help to address whether GFRα2 signaling is required for the development or maintenance of epidermal nerve endings in the footpad, and whether epidermal innervation of other cutaneous areas have a similar requirement for GFRα2 signaling. A similar analysis of NRTN-KO mice could also be helpful.
Both GDNF and NRTN are expressed in developing mouse skin (Golden et al., 1999), and GFRα1 and GFRα2 are expressed in distinct but partly overlapping subpopulations of IB4-binding neurons (Bennett et al., 1998b), suggesting that the remaining CGRP-negative nerve endings in GFRα2-KO mouse epidermis may be GDNF dependent. Interestingly, the morphological changes found in the cutaneous sensory neurons of GFRα2-KO mice, namely the reduced caliber of cutaneous axons (Stucky et al., 2002) and density of nonpeptidergic epidermal innervation (this study), are opposite to those found in mice overexpressing GDNF in skin keratinocytes (Zwick et al., 2002). Because in vivo signaling for GDNF and neurturin in sensory neurons is GFRα receptor-specific (Leitner et al., 1999), it is possible that the morphological changes in GDNF-overexpressing mice are restricted to the GFRα1-expressing subpopulation of cutaneous sensory neurons. It would be informative to compare sensory phenotype of these mice to mice overexpressing NRTN in skin.
We show here that behavioral responses to innocuous warm and noxious heat are not reduced in the GFRα2-KO mice in vivo, and thus the physiological relevance of the heat transduction deficit in vitro (Stucky et al., 2002) remains unclear. Transient receptor potential receptors are temperature-sensitive ion channels mainly expressed in different sensory neurons (Jordt et al., 2003; Patapoutian et al., 2003). We show that TRPV1 was not coexpressed with GFRα2 in wild-type mouse DRG, and the density of TRPV1-positive neurons was unchanged in GFRα2-KO mouse DRG (data not shown). Consistent with this, IB4-binding neurons do not express TRPV1 in mouse DRG (Zwick et al., 2002; Woodbury et al., 2004), suggesting that receptors other than TRPV1 transduce noxious heat in GFRα2/IB4-positive nociceptors. However, because neurotrophic factors (including GDNF) and peripheral inflammation increase TRPV1 expression and function in the IB4-positive sensory neurons (Amaya et al., 2004; Breese et al., 2005), a role for GFRα2 signaling in TRPV1-mediated thermal hyperalgesia remains possible.
Recent studies of innocuous warm sensation suggest TRPV3 and TRPV4 receptors in skin keratinocytes as the sensors of warm stimulus, which transmit the signal to the epidermal sensory nerve endings (Lee et al., 2005; Moqrich et al., 2005). Our demonstration that mice lacking 50–70% of the epidermal nerve endings have no gross deficits in innocuous warm sensation may seem at odds with such a model. However, the remaining innervation may be sufficient for the KO mice to respond normally in the two-temperature choice test. The hypothesis that normal behavior can be exhibited despite dramatic anatomical changes is supported by a study in which NGF-deficient mice were rescued by transgenic expression of NGF in the skin. The response of these mice to noxious thermal stimuli was fully recovered, although the IB4-positive sensory neuron population was only modestly restored (Harrison et al., 2004).
NGF and the peptidergic nociceptors are important mediators of inflammatory pain (Snider and McMahon, 1998; Woolf and Costigan, 1999; Hunt and Mantyh, 2001), but recent studies suggest that the nonpeptidergic nociceptors are also involved (Breese et al., 2005). Consistent with this idea, we show that GFRα2-KO mice have reduced formalin-induced inflammatory pain response. The intact first-phase response in GFRα2-KO mice suggests that functional chemonociceptor terminals mediating the acute phase are present in KO mouse skin. Inflammation-induced hyperactivity of C-fibers generates and sustains an elevated discharge of dorsal horn neurons (Pitcher and Henry, 2002), which is thought to be critical for the second, persistent phase formalin response (Tjolsen et al., 1992; Puig and Sorkin, 1995; Taylor et al., 1995). Thus, a parsimonious explanation for the reduced second-phase response in GFRα2-KO mice would be weaker activation of the nonpeptidergic DRG neurons. However, the behavioral pain response involves a network of neurons and other cells and is modulated at several levels. Whether the reduced formalin-induced inflammatory pain response in GFRα2-KO mice involves peripheral and/or central component mechanisms and whether pain responses in the KO mice are altered in other models of inflammatory pain remain to be studied.
Interestingly, formalin-induced inflammatory pain is inhibited by specific P2X3 blockers (Tsuda et al., 1999; McGaraughty et al., 2003) and in mice lacking P2X3 (Cockayne et al., 2000; Souslova et al., 2000). The P2X3-deficient mice also have a remarkably similar temperature sensation phenotype to the GFRα2-KO mice, including shorter withdrawal latencies at temperatures above and below thermoneutrality in the tail flick test (Shimizu et al., 2005). The extensive colocalization between GFRα2 and P2X3 in DRG neurons suggests that the loss of nonpeptidergic nerve fibers in the GFRα2-KO mouse epidermis represents predominantly P2X3-positive nerve endings. Thus, an intriguing possibility is that attenuated inflammatory pain response (and enhanced thermal avoidance) in GFRα2-KO mice is a result of reduced activation of P2X3. Another candidate mediator for the reduced inflammatory pain is bradykinin B1 receptor, the expression of which in nociceptive neurons can be upregulated by neurturin (Vellani et al., 2004). A number of other molecules contribute to inflammatory pain, including immune and glial cell factors involved in the inflammatory response (Marchand et al., 2005).
In conclusion, we have shown that the majority of nonpeptidergic cutaneous free nerve endings in the mouse footpad require GFRα2 signaling. In addition, this signal is required to maintain the size of the corresponding unmyelinated nociceptive sensory neurons. We propose that NRTN, the cognate GFRα2 ligand expressed in the epidermis (Golden et al., 1999), may act as a short-range factor that helps the C-fiber terminals to branch, grow, and remain in the relatively repulsive environment of the epidermis (Cahoon and Scott, 1999). In addition to being distinct in terms of trophic requirements, molecular markers, and spinal projections (Snider and McMahon, 1998; Hunt and Mantyh, 2001), recent studies suggest that the peptidergic and nonpeptidergic subsets of polymodal nociceptors differ in their intracellular mechanisms of sensitization (Hucho et al., 2005), skin innervation patterns (Zylka et al., 2005), and ascending pathways to the brain (Braz et al., 2005). GFRα2-KO mice provide a unique model to test the role of nonpeptidergic cutaneous innervation in pain and other sensory functions.
Reduced density of epidermal nerve endings is a general finding in human neuropathic syndromes (Chien et al., 2001), including diabetic and idiopathic small fiber neuropathy (Holland et al., 1997) and postherpetic neuralgia (Fields et al., 1998; Oaklander, 2001), and is correlated with clinical severity. Remarkably, loss of epidermal innervation is often found without obvious defect in the peripheral nerve (Periquet et al., 1999; Herrmann et al., 1999), raising the possibility of altered NRTN/GFRα2 signaling in these conditions. Because NRTN treatment has been shown to stimulate growth and branching of cutaneous axons in diabetic mice (Christianson et al., 2003), the GFRα2-signaling pathway appears to be a potential target of therapy for sensory regeneration and persistent pain.
Footnotes
-
This work was supported by grants from the Finnish Academy and the Sigrid Jusélius Foundation (M.S.A.). We thank Kaija Berg for technical assistance and Heikki Rauvala for comments.
- Correspondence should be addressed to Matti S. Airaksinen, Neuroscience Center, Viikinkaari 4, University of Helsinki, 00014 Helsinki, Finland. Email: mairaksi{at}operoni.helsinki.fi





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}