

Abstract
Cocaine-induced plasticity persists during abstinence and is thought to underlie cue-evoked craving. Reversing this plasticity could provide an opportunity for therapeutic intervention. Converging evidence suggest that zeta inhibitory peptide (ZIP) eliminates memories for experience-dependent behaviors, including conditioned drug associations. However, the effect of ZIP on reward seeking and drug-induced plasticity is unknown. The current study examined the effect of ZIP administration in the nucleus accumbens on reinstatement (RI) of cocaine seeking, a rodent model of relapse. We demonstrate that intra-accumbal ZIP administration blocks cocaine-primed RI in rats when administered 24 h or 1 week before testing. These effects of ZIP on drug seeking are specific, as we did not see any effect of ZIP on RI of sucrose seeking. ZIP is a synthetic compound designed to inhibit the atypical PKC, PKMζ, a protein implicated in learning and memory. However, recent evidence from PKMζ-knock-out (KO) mice suggests that ZIP may function through alternative mechanisms. In support of this, we found that ZIP was able to block cue-induced RI in PKMζ-KO mice. One possible mechanism underlying addictive phenotypes is the ability of cocaine to block further plasticity. We hypothesized that ZIP may be working to reverse this anaplasticity. Although ZIP has no effect on accumbal LTD in slices from naive or yoked saline mice, it is able to restore both NMDA-dependent and mGluR5-dependent LTD in animals after cocaine self-administration and withdrawal. These findings demonstrate that intra-accumbal ZIP persistently reverses cocaine-induced behavioral and synaptic plasticity in male and female rodents.
SIGNIFICANCE STATEMENT Zeta-inhibitory peptide (ZIP) has been shown to disrupt memory maintenance for experience-dependent behaviors. We examined the effect of ZIP infused into the nucleus accumbens on the reinstatement (RI) of cocaine seeking. We found that intra-accumbal ZIP blocked RI of cocaine seeking 24 h and 1 week later. This effect was specific to RI of cocaine seeking as ZIP did not disrupt RI of food seeking. In conjunction with these behavioral studies we examined the ability of ZIP to reverse cocaine-induced deficits in LTD. We found that ZIP was able to rescue two forms of LTD in cocaine-experienced mice. These studies demonstrate that ZIP is able to reverse cocaine-induced behavioral and synaptic plasticity in a persistent manner.
Introduction
Cocaine addiction is a chronic, relapsing disorder characterized by continued use in the face of adverse consequences. Despite decades of research, a safe, efficacious pharmacological treatment for cocaine addiction remains to be discovered. Relapse to cocaine use is partially mediated by persistent cue-driven, drug memories (Childress et al., 1993; Dackis and O'Brien, 2001). These memories for drug-paired cues elicit craving in human addicts and reinstate extinguished cocaine-seeking behavior in rodent models (Everitt et al., 2018). During abstinence memories associated with drug-taking persist and the inability to eliminate these drug memories is thought to underlie addiction. Although many studies have demonstrated temporary pharmacological inhibition of cue-driven cocaine seeking (Cornish et al., 1999; Mahler et al., 2013, 2014; Knackstedt et al., 2014; Reissner et al., 2015; LaCrosse et al., 2017; Manuszak et al., 2018), a more persistent disruption has been elusive.
Zeta-inhibitory peptide (ZIP) is a cell-penetrating peptide derived from the pseudosubstrate domain of the PKC isoform, PKMζ, a constitutively active splice variant of PKCζ (Pastalkova et al., 2006). This peptide has been used extensively in memory studies to support a role for PKMζ in maintaining memories via long-term potentiation (Hrabetova and Sacktor, 1996; Pastalkova et al., 2006; Sacktor, 2011; Zhang et al., 2016). Additionally, ZIP has also been shown to disrupt cocaine-conditioned reward in the nucleus accumbens (NAc) (Li et al., 2011; Shabashov et al., 2012), cocaine-induced synaptic potentiation in the ventral tegmental area (VTA) (Ho et al., 2012; Vaquer-Alicea et al., 2018), and cocaine sensitization when injected into the ventricle (Howell et al., 2014). However, recent studies using mice deficient in PKMζ show that these isoforms are not necessary for synaptic plasticity or memory (Lee et al., 2013; Volk et al., 2013). ZIP is still able to impair LTP maintenance and several types of memory, including conditioned drug associations, in mice deficient in PKCζ/PKMζ, suggesting that the actions of ZIP are independent of PKMζ (Lee et al., 2013).
Here, we examined the ability of ZIP to alter cocaine-induced behavioral and synaptic plasticity. To do this, we examined the ability of intra-accumbal ZIP administration to disrupt reinstatement (RI) of cocaine seeking in both rats and mice. Synaptic plasticity within the NAc is associated with cue driven cocaine seeking. Specifically, cocaine self-administration leads to blunted long-term depression (LTD), a form of meta plasticity that is thought to underlie reduced behavioral flexibility in individuals with substance use disorder (Martin et al., 2006; Moussawi et al., 2009; Kasanetz et al., 2010). To determine whether ZIP is able to rescue this blunted plasticity after cocaine use, we examined the effect of ZIP on LTD in the NAc after cocaine self-administration. Our results suggest that ZIP is able to disrupt cocaine-induced behavioral and synaptic plasticity in a persistent manner.
Materials and Methods
Subjects.
Male Sprague Dawley rats (Rattus norvegicus) weighing 250–275 g were obtained from Taconic Laboratories. Animals were individually housed with food and water available ad libitum in their home cages. Male and female heterozygous PKMζ-knock-out (KO) mice on a C57BL/6 background (as described previously in Volk et al., 2013) were mated resulting in mutant and wild-type littermates. Additional male and female C57BL/6J mice were bred in house for electrophysiology experiments. Mice (2–6 months old, 20–40 g; age matched across group) were group housed until the start of the behavioral experiments at which point they were individually housed. All animals were housed in a temperature- and humidity-controlled animal care facility with a 12 h light/dark cycle (lights on at 7:00 A.M.). All procedures were approved by the Temple University Animal Care and Use Committee.
Drugs.
Cocaine was obtained from the National Institutes of Drug Abuse Drug Supply Program (Bethesda, MD) and dissolved in sterile 0.9% saline. ZIP (catalog #2549) and a scrambled, inactive form of the peptide (Serrano et al., 2008), SCR-ZIP (catalog #3215), were obtained from Tocris Bioscience. For the in vivo drug administration, ZIP and SCR-ZIP was dissolved in artificial CSF (ACSF). For the electrophysiological recordings, a stock solution was made by dissolving ZIP in 80% H2O and 20% ETOH. The final concentration of ETOH in the bath was 0.19%.
Surgery.
Rats and mice were anesthetized with 80 mg/kg ketamine and 12 mg/kg xylazine. An indwelling SILASTIC catheter was placed into the right jugular vein and sutured in place. Then the catheter was threaded subcutaneously over the shoulder blade and was routed to a mesh back mount platform (Strategic Applications) that secured the placement. The catheters were flushed daily with 0.1 ml of antibiotic (Timentin, 0.93 mg/ml) dissolved in heparinized saline. The catheters were sealed with plastic obturators when not in use. After catheter insertion, rats and mice were immediately mounted in a stereotaxic apparatus (Kopf Instruments). Guide cannulas (rat: 14 mm, 24 gauge; mouse; 9 mm, 26 gauge) for microinjections were implanted bilaterally 2 mm (rat) or 1 mm (mouse) dorsal to the accumbens. Guide cannulas were cemented in place by affixing dental acrylic to stainless steel screws secured in the skull. The coordinates for the ventral ends of the guide cannulas, relative to bregma, were as follows: Rat: 1.2 mm A/P, ±1.0 mm M/L, and 5.2 mm D/V; mouse: +1.5 A/P, +/− 1.0 M/L, −4.0 D/V. An obturator (14 mm (rat), 9 mm (mouse), 33 gauge) was inserted into each guide cannula to prevent occlusion.
Rat cocaine and sucrose self-administration, extinction and RI of cocaine seeking.
After surgery, rats were allowed 7 d to recover before behavioral testing commenced. Initially, rats were placed in operant chambers and allowed to lever press for intravenous infusions of cocaine (0.25 mg of cocaine/59 μl of saline, infused over a 5 s period) on a fixed-ratio 1 (FR1) schedule of reinforcement. When a cocaine infusion was delivered, a cue light above the active wheel was illuminated for 5 s and the house light turned off. Each operant session began with the intravenous administration of 0.25 mg of cocaine (dissolved in 59 μl of saline) to fill the catheter. Rats were allowed to self-administer a maximum of 30 injections per 120 min operant session. Once an animal achieved at least 20 infusions of cocaine in a single daily operant session under the FR1 schedule, the subject was switched to an FR5 schedule of reinforcement. The maximum number of injections was again limited to 30 per daily self-administration session under the FR5 schedule. For both FR1 and FR5 schedules, a 20 s time-out period followed each cocaine infusion, during which time the house light remained off, active lever responses were tabulated but had no scheduled consequences. Responses made on the inactive lever, which had no scheduled consequences, were also recorded during both the FR1 and FR5 training sessions. After 18 d of daily cocaine self-administration sessions, drug seeking behavior was extinguished by replacing the cocaine with 0.9% saline. Daily extinction sessions continued until responding on the active lever was <15% of the total active lever responses completed on the last day of cocaine self-administration maintained on a FR5 schedule of reinforcement. Typically, it took ∼7 d for rats to meet this criterion. Once cocaine self-administration was extinguished, animals entered the RI phase of the experiment. To control for any natural variation in RI behavior, all animals underwent a RI pretest in which a systemic priming injection of cocaine (10 mg/kg, i.p.) was administered immediately before the test session. During the RI test sessions, satisfaction of the response requirement (i.e., five presses on the active lever) resulted in an infusion of saline rather than cocaine. Rats then underwent 1–2 d of extinction to return to the extinction criterion. Two hours after the final extinction session, rats received bilateral intra-accumbal microinfusions of ZIP (30 nmol) or SCR-ZIP. They then received another cocaine-primed RI session either 24 h or 1 week later. Groups were balanced based upon their cocaine self-administration, extinction and RI data. Catheter patency was checked at the end of the cocaine self-administration phase and any animals that were no longer patent were removed from the study (n = 2).
Rat sucrose self-administration, extinction, and RI of sucrose seeking.
A separate group of rats underwent a similar procedure with 18 d of sucrose self-administration (FR1 transitioning to FR5 as described above), extinction of sucrose seeking, and sucrose-primed RI. For the sucrose RI, the experimenter remotely administered one sucrose pellet every 2 min for the first 10 min of the RI test session. During this session, active lever presses had no scheduled consequences. Rats were only tested for sucrose RI at the 24 h postmicroinfusion time point.
Mouse food training, cocaine self-administration, extinction, and RI.
Before catheterization, mice were trained to perform an operant response for sucrose pellets. The mice were placed in operant chambers (Med Associates) where they learned to spin a wheel manipulandum to receive the sucrose pellet. When the pellet was delivered, a cue light above the active wheel was illuminated, a 2900 Hz tone played, and the house light turned off. This was followed by an 8 s time-out where the house light remained off and spinning the wheel had no programed consequences. Mice were able to self-administer up to 50 pellets per 60 min operant session. The mice were food restricted to ∼90% of their free feeding weight throughout the course of the operant training. They were returned to ad libitum feeding 3 d into the cocaine self-administration sessions. After 10 d of food self-administration, mice underwent jugular catheterization surgery as described above. Mice were given 3–4 d to recover from surgery before resuming behavioral testing. The cocaine self-administration testing was measured over 2 h sessions (6 d per week) in the same chamber used for the operant food training. During testing, responding on the active wheel delivered an intravenous cocaine injection (0.6 mg/kg/infusion,) paired with the same cues as the food training. After 10 d of cocaine self-administration, cocaine seeking was extinguished by replacing the cocaine with 0.9% saline. During extinction, the light and tone cues were not present. Daily 2 h extinction continued until the mice met the extinction criterion of <25% of their responding during the self-administration (average of the last 3 d). Two hours after the extinction session mice received microinjections of either ZIP or Scr-ZIP (see below for more details). Twenty-four hours after meeting the extinction criterion animals underwent a cue-induced RI session. The light and tone were presented noncontingently for 20 s every 2 min during the first 10 min of the session. For the remaining 110 min, the cues were presented after responses on the active wheel, just as was done during the cocaine self-administration sessions. During RI, the mice received saline infusions after active responses. For the physiological measurements, mice were exposed to 10–12 d of cocaine self-administration or yoked saline experience. Saline-yoked mice were placed in identical operant chambers however their responding had no programmed consequences. Instead, they received saline infusions along with cue presentations when their partner mouse received cocaine. Mice were killed 10–14 d after the final day of self-administration. As previous work has shown LTD to be disrupted in rodents both with and without extinction training (Moussawi et al., 2009; Briand et al., 2014), these mice were exposed to forced abstinence rather than active extinction.
Microinjection procedures.
Obturators were removed from the guide cannulas and 33-gauge, 16 mm (rat) or 10 mm (mouse) stainless steel microinjectors were inserted. Bilateral infusions were performed simultaneously over 2 min in a total volume of 0.5 μl (rat) or 0.4 μl (mouse) per hemisphere. After infusion, microinjectors were left in place for an additional 1 min to allow for diffusion of the drug solution away from the tips of the microinjectors.
Verification of cannula placements.
After completion of all microinjection experiments, rats were given an overdose of pentobarbital (100 mg/kg) and perfused with 0.9% saline followed by 10% formalin. Brains were removed and coronal sections (100 mm) were taken at the level of the NAc with a vibratome. The sections were mounted on gelatin-coated slides and stained with cresyl violet. An individual blind to behavioral responses confirmed cannula placements and removed any animals that had excessive cannula-induced damage. Two rats were removed from the studies due to inaccurate cannula placements and one was removed for excessive cannula-induced damage.
Nucleus accumbens slices.
C57/B6J mice were decapitated after cervical dislocation. The brain was removed and coronal slices (250 μm) containing the NAc were cut with a Vibratome (VT1000S, Leica Microsystems) in an ice-cold ACSF, in which NaCl was replaced by an equiosmolar concentration of sucrose. ACSF consisted of 130 mm NaCl, 3 mm KCl, 1.25 mm NaH2PO4, 26 mm NaHCO3, 10 mm glucose, 1 mm MgCl2, and 2 mm CaCl2 (pH 7.2–7.4 when saturated with 95% O2/5% CO2). Slices were incubated in ACSF at 32–34°C for 25 min and kept at 22–25°C thereafter, until transfer to the recording chamber. The osmolarity of all solutions was 300–315 mOsm. Slices were viewed using infrared differential interference contrast optics under an upright microscope (Slice Scope Pro; Scientifica) with a 40 × water-immersion objective. For the ZIP bath manipulation ZIP (5 μm) was added to the bath after 10 min of stable field EPSP (fEPSP) recording and another 10 min of EPSP recordings were taken to confirm the peptide did not alter baseline responding.
Electrophysiology.
The recording chamber was continuously perfused (1–2 ml/min) with oxygenated ACSF heated to 32 ± 1°C using an automatic temperature controller (Warner Instruments). Picrotoxin (100 μm) was added to all solutions to block the GABAA receptor-mediated currents. Recording pipettes were pulled from borosilicate glass capillaries (World Precision Instruments) to a resistance of 4–7 MΩ when filled with the intracellular solution (whole-cell recordings) or to a resistance of 1–2 MΩ when filled with extracellular solution (field recordings). All recordings were conducted with a MultiClamp700B amplifier (Molecular Devices).
LTD.
A bipolar tungsten stimulating electrode was placed within 100–300 μm from the recording electrode and used to stimulate excitatory afferents at 0.1 Hz. The field recordings were performed within the core of the NAc. The amplitude of current pulses was set at the intensity required to evoke a 70% maximal response. After 10 min of stable responding, LTD was induced using a paired-pulse protocol (50 ms interpulse interval) consisting of a 1 Hz train of paired stimuli for 7.5 min. Both the fEPSP slope amplitude were measured (graphs depict slope) from fEPSPs recorded at 0.1 Hz for 50 min following the pairing protocol. To induce mGluR5-dependent LTD, after recording a stable baseline for at least 10 min (S)-3,5-dihydroxyphenylglycine (DHPG; 100 μm) was applied via bath application for 10 min, followed by ACSF. Field EPSPs were recorded for 60 min after the start of DHPG application. For the ZIP recordings, ZIP was present in the bath throughout the entire recording protocol.
Whole-cell recordings.
Intracellular solution contained the following (in mm): 100 CsCH3O3S, 50 CsCl, 3 KCl, 0.2 BAPTA, 10 HEPES, 1 MgCl2, 2.5 phosphocreatine-2Na, 2 Mg-ATP, 0.25 GTP-Tris, 1 QX-314 (pH 7.2–7.3 with CsOH, osmolarity 280–290 mOsm). Currents were low-pass filtered at 2 kHz and digitized at 20 kHz using a Digidata 1440A acquisition board and pClamp10 software (both from Molecular Devices). Access resistance (10–30 MΩ) was monitored throughout the recordings by injection of 10 mV hyperpolarizing pulses and data were discarded if access resistance changed by >25% over the course of data acquisition. Evoked responses were triggered by 100 μs constant-current pulses generated by an A310 Accupulser (World Precision Instruments) and delivered at 0.1 Hz via a bipolar tungsten stimulation electrode positioned within 100 μm of the recorded cell. The amplitude of the current pulses was controlled by a stimulus isolator (World Precision Instruments Linear Stimulus Isolator A395) and was adjusted to elicit monosynaptic responses in the range of 100–300 pA (the required stimulus intensity ranged from 15 to 80 μA). AMPA/NMDA current ratios were computed by dividing the mean peak eEPSC at −70 mV (AMPA-mediated) by the mean amplitude at +40 mV, 35 ms after the peak over a 2 ms window (NMDA-mediated). For all measures, cells from at least 3 animals, within each group, were used. Recordings were taken from cells within the accumbens core.
Data analysis.
All analyses were performed using GraphPad Prism 7.0 software. Data were analyzed using two-tailed Student's t test or two-way ANOVA with Sidak's post hoc test as appropriate. Statistical significance for all tests was set at α = 0.05.
Results
Intra-accumbal infusion of ZIP 24 h or 1 week before testing blocks cocaine-primed RI of cocaine seeking in rats
Rats underwent 18 d of cocaine self-administration followed by 7–10 d of extinction until reaching the extinction criterion (<15% of their cocaine responding). Twenty-four hours after reaching the extinction criterion, all rats were given a cocaine-primed RI test in which they received an injection of cocaine (10 mg/kg, i.p.) immediately before being placed into the box under extinction conditions. Cocaine-primed RI was chosen because it induces the most robust RI in rats and allows for testing of multiple RI sessions without much decay in responding across tests. Rats in both groups exhibited a significant RI of cocaine seeking (Fig. 1A; two-way repeated-measures ANOVA (including all extinction 1, RI pretest, extinction 2, and RI posttest): main effect of test, F(3,30) = 46.19, p < 0.0001; interaction, F(3,30) = 10.0, p < 0.0001, Sidak's post hoc test, SCR-ZIP and ZIP extinction vs pretest, p < 0.0001, n = 6/group). Two hours after rats again reached the extinction criterion they were given a microinjection of either ZIP (30 nmol) or SCR-ZIP. Twenty-four hours after this peptide administration, rats were again tested on cocaine-primed RI. Rats that had received a microinfusion of ZIP into the accumbens failed to exhibit RI of cocaine seeking (Fig. 1A; Sidak's post hoc test, SCR-ZIP posttest vs ZIP posttest, p < 0.0001; n = 6/group). To determine the longevity of ZIP's effects, an additional cohort of animals were injected with ZIP (30 nmol) or SCR-ZIP 1 week before the second cocaine-primed RI test. Although rats receiving the SCR-ZIP infusion exhibited RI of cocaine seeking after cocaine injection, rats that received intra-accumbal infusion of ZIP 1 week before the RI test did not (Fig. 1B; two-way repeated-measures ANOVA (including extinction, RI pretest, and RI posttest): main effect of test, F(2,20) = 14.1, p = 0002; interaction, F(2,20) = 6.3, p = 0.008; Sidak's post hoc test, SCR-ZIP posttest vs ZIP posttest, p = 0.0025; n = 6/group).

Intra-accumbal ZIP administration leads to persistent inhibition of cocaine seeking during RI. A, Dots indicate the location of the bilateral injections of 0.5 μl of ZIP (black) or Scr-ZIP (gray) into the nucleus accumbens. B, Despite both groups exhibiting equivalent levels of RI of cocaine (Coc) seeking during the pretest, intra-accumbal infusion of ZIP (30 nmol) 24 h before test blocked RI of cocaine seeking (bars represent mean ± SEM; ***p < 0.001 Sidak post hoc ZIP vs SCR-ZIP on posttest, n = 6/group). C, Similar blunted RI behavior was seen in rats that received intra-accumbal ZIP administration 1 week before the RI test (c, **p < 0.01 Sidak post hoc ZIP vs SCR-ZIP on posttest, n = 6/group). D, Intra-accumbal infusion of ZIP did not alter RI of sucrose seeking behavior 24 h after infusion (n = 7–8/group).
Intra-accumbal infusion of ZIP 24 h before testing does not alter sucrose-primed RI of sucrose seeking in rats
To determine whether the effects of ZIP generalized to natural reinforcers, we ran a separate cohort of rats to test RI of sucrose seeking. After 18 d of sucrose self-administration and extinction, all rats were exposed to a sucrose-primed RI pretest to allow for appropriate balancing of the groups. Two hours after rats again reached the extinction criterion they were given a microinjection of either ZIP (30 nmol) or SCR-ZIP. Twenty-four hours after this peptide administration, rats were again tested on sucrose-primed RI. Both the animals that received ZIP and SCR-ZIP exhibited significant RI on the posttest (Fig. 1C; two-way repeated-measures ANOVA (including all extinction 1, RI pretest, extinction 2, and RI posttest): main effect of test, F(3,39) = 31.9, p < 0.0001; interaction, F(3,39) = 0.33, p = 0.08, Sidak's post hoc test, SCR-ZIP and ZIP extinction vs posttest, p < 0.0001; n = 7–8/group).
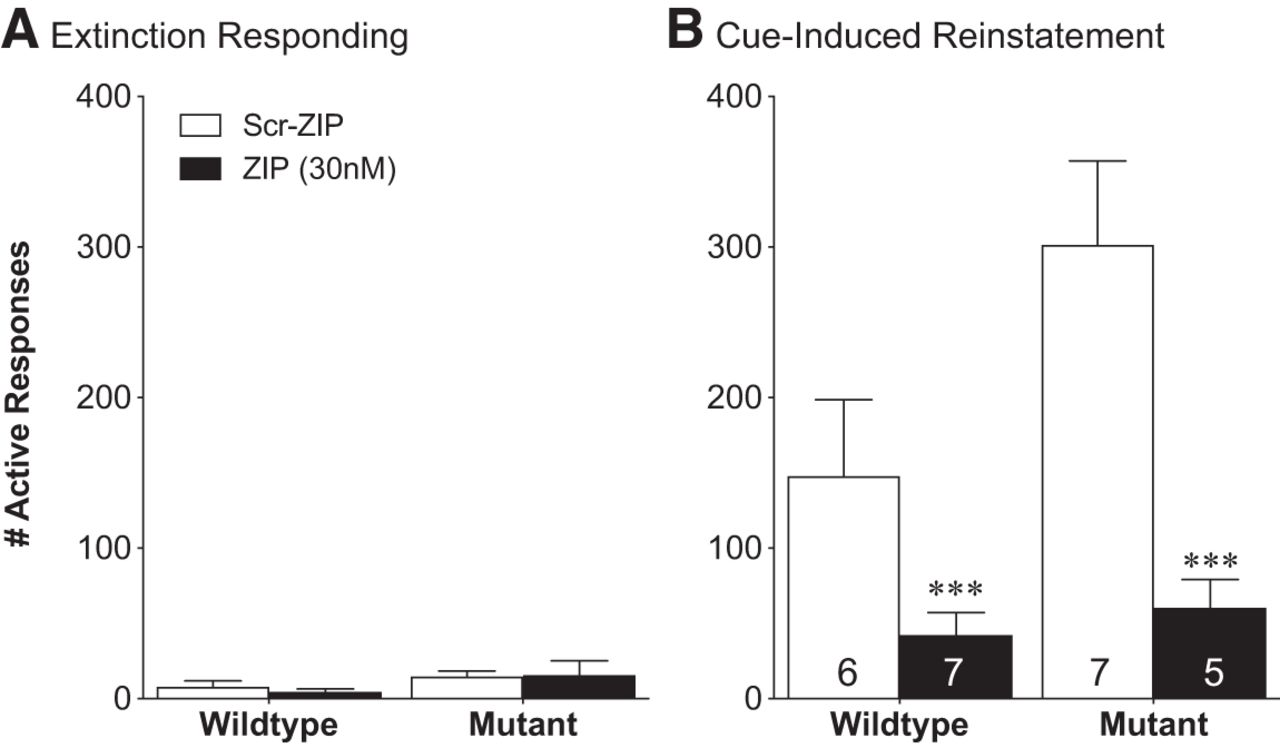
Intra-accumbal infusion of ZIP 24 h before testing blocks cue-induced RI of cocaine seeking in PKMζ-KO mice
For cross-species validation and to determine whether the effects of ZIP were dependent upon PKMζ, we examined the ability of ZIP to block RI of cocaine seeking in male and female wild-type and constitutive PKMζ-KO mice. As the role of glutamatergic circuits is similar in different forms of RI (McFarland and Kalivas, 2001; Kalivas and McFarland, 2003; McFarland et al., 2004) and cues provide the most robust driver of RI behavior in mice, we chose to examine cue-induced cocaine seeking in these studies. Wild-type and constitutive PKMζ KO mice underwent 10 d of food training to acquire operant responses for food, followed by 10 d of cocaine self-administration (FR1, 0.65 mg/kg/infusion) and an average of five extinction sessions. Mice were then divided into groups to normalize their cocaine intake, cocaine responding, and extinction responding. As described previously (McGrath et al., 2018), PKMζ KO mice exhibit higher levels of responding during cocaine self-administration so the groups were balanced between infusion groups but not across genotype. Two-hours after their last extinction session mice were given microinfusions of either ZIP (30 nmol) or SCR-ZIP into their NAc. Twenty-four hours later they underwent a cue-induced RI session. Although mice given the SCR-ZIP infusion exhibited significant RI of responding during the session, mice given the ZIP infusion did not (effect of test, F(1,10) = 16.76, p = 0.002; interaction, F(1,10) = 9.54, p = 0.01; Sidak's multiple-comparisons test, SCR-ZIP vs ZIP RI test, adjusted p = 0.0009; n = 5–7/group; Figure 2).

Intra-accumbal ZIP administration blocks cue-induced RI of cocaine seeking in constitutive PKMζ-KO mice. A, Two hours after meeting the extinction criteria, wild-type and PKMζ KO mice received intra-accumbal infusions of either ZIP or SCR-ZIP. B, Twenty-four hours later, while both wild-type and PKMζ KO mice that received SCR-ZIP infusions exhibited significant cue-induced RI of cocaine seeking, both groups that received ZIP infusions did not. Error bars indicate mean ± SEM; ***p < 0.001 main effect of infusion on cue-induced RI. Numbers inside bars represent ns for each group.
Cocaine-induced disruption in NMDAR-dependent LTD is rescued by ZIP administration
After demonstrating that ZIP blocks cocaine-induced behavioral plasticity, we next examined whether ZIP could reverse cocaine-induced synaptic plasticity in wild-type C57BL/6J mice. As blunted accumbal LTD is seen after cocaine experience (Martin et al., 2006; Moussawi et al., 2009; Kasanetz et al., 2010; Briand et al., 2014; Heinsbroek et al., 2017), we hypothesized that bath application of ZIP (5 μm) would rescue this deficit. We found that slices from yoked control mice exhibited robust LTD within the NAc core (NAcc) and this response was not altered by bath application of ZIP (Fig. 3A). Consistent with what we have seen previously (Briand et al., 2014), LTD was absent in mice after 10–12 d of cocaine self-administration and 10–12 d of withdrawal (Fig. 3B). However, after bath application of ZIP, a low-frequency train of stimuli (1 Hz, 10 min) induced robust LTD in NAcc slices from cocaine-experienced mice (effect of cocaine, F(1,32) = 11.72, p = 0.002; effect of ZIP F(1,32) = 6.86, p = 0.01; interaction, F(1,32) = 3.85, p = 0.05; Sidak's multiple-comparisons test, cocaine ACSF vs cocaine ZIP, adjusted p = 0.008; n = 7–12 slices (3–4 mice)/group; Figure 3B–D).

Bath application of ZIP rescues blunted NMDAR-LTD after cocaine self-administration. A, Yoked saline control mice exhibit reliable long-term depression after low-frequency stimulation (LFS) (1 Hz, 10 min) and this is not altered by bath application of ZIP. B, Ten to 14 d of cocaine self-administration followed by 10–14 d of withdrawal leads to blunted NMDAR-LTD that is rescued by bath application of 5 μm ZIP (**p < 0.01 main effect of bath infusion after LFS). C, Example traces demonstrate the rescue of LTD by ZIP in cocaine-experienced mice with baseline in black and post-LFS in red. D, Change in fEPSP slope across the 30 min after LFS demonstrates robust LTD in control mice, blunted LTD after cocaine, and the rescue of LTD after bath application of ZIP in cocaine-experienced mice (**p < 0.01 Sidak post hoc ACSF vs ZIP in cocaine-treated mice).
Cocaine-induced disruption in mGluR5-dependent LTD is rescued by ZIP administration
To determine whether ZIP's ability to rescue cocaine-induced deficits in LTD was specific to NMDAR-mediated plasticity, we examined mGluR5-dependent LTD within the NAcc of cocaine-experienced and yoked control mice. Previous work has shown disruptions in mGluR5-dependent LTD with the accumbens shell after experimenter-administered cocaine (Huang et al., 2011, 2015). We found that 10–14 d of cocaine self-administration followed by 10–14 d of withdrawal led to blunted DHPG induced LTD (effect of cocaine, F(1,18) = 5.62, p = 0.03; effect of ZIP, F(1,18) = 4.41, p = 0.05; interaction, F(1,18) = 6.34, p = 0.02; Sidak's multiple-comparisons test, control ACSF vs cocaine ACSF, adjusted p = 0.004; n = 5–6 slices (3–4 mice)/group; Fig. 4). Further, bath application of ZIP rescued DHPG-LTD in cocaine experienced mice (Sidak's multiple-comparisons test, cocaine ACSF vs cocaine ZIP, adjusted p = 0.009; Fig. 4).

Bath application of ZIP rescues blunted mGluR5-LTD after cocaine self-administration. A, Yoked saline control mice exhibit reliable long-term depression after DHPG administration (100 μm, 10 min) and this is not altered by bath application of ZIP. B, Ten to 14 d of cocaine self-administration followed by 10–14 d of withdrawal leads to blunted mGluR5-LTD that is rescued by bath application of 5 μm ZIP (*p < 0.05 main effect of bath infusion after DHPG). C, Example traces demonstrate the rescue of mGluR5-LTD by ZIP in cocaine-experienced mice with baseline in black and post-DHPG in red. D, Change in fEPSP slope across the 30 min after DHPG demonstrates robust LTD in control mice, blunted LTD after cocaine, and the rescue of LTD after bath application of ZIP in cocaine-experienced mice (*p < 0.05 Sidak post hoc ACSF vs ZIP in cocaine-treated mice).
Bath administration of ZIP does not disrupt evoked field potentials
To confirm that ZIP did not disrupt the evoked field potentials, we examined responses during the 10 min before administration of ZIP and the 10 min after the start of bath application. We did not see any differences in average fEPSP slope (Control: 0.21 ± 0.026; ZIP: 0.20 ± 0.024; n = 8/group) or average fEPSP amplitude (control: 0.42 ± 0.047; ZIP: 0.43 ± 0.056; n = 8/group) during these two 10 min windows.
Cocaine-induced increase in AMPA/NMDA ratio is reversed by ZIP administration
To determine whether ZIP is able to reverse other forms of cocaine-induced plasticity, we examined the AMPA/NMDA ratio within the NAcc of cocaine-experienced and yoked control mice. Previous work has shown increases in AMPA/NMDA ratio within the NAcc after cocaine self-administration (Moussawi et al., 2009). We found that 10–14 d of cocaine self-administration followed by 10–14 d of withdrawal led to an increase in the AMPA/NMDA ratio in the NAcc and this increase was reversed after bath application of ZIP (effect of cocaine, F(1,62) = 10.93, p = 0.001; effect of ZIP, F(1,62) = 2.85, p = 0.096; interaction, F(1,62) = 5.44, p = 0.02; Sidak's multiple-comparisons test, cocaine vs cocaine + ZIP, adjusted p = 0.016; n = 13–18 cells from 5–6 mice per group; Fig. 5).

Bath application of ZIP reversed cocaine-induced increases in AMPA/NMDA ratio in the NAcc. Ten to 14 d of cocaine self-administration followed by 10–14 d of withdrawal leads to increased AMPA/NMDA ratio that is reversed by bath application of 5 μm ZIP. Error bars indicate mean ± SEM. Numbers in bars represent number of cells (number of animals); *p < 0.05 Sidak post hoc cocaine vs cocaine + ZIP.
Discussion
The current studies demonstrate that a single intra-accumbal infusion of ZIP can block RI of cocaine seeking up to 1 week later. Consistent with what has been seen for the effects of ZIP on hippocampal dependent memory and long-term potentiation (Lee et al., 2013; Volk et al., 2013), the ability of ZIP to block cocaine seeking is PKMζ independent. However, the effects of ZIP on cocaine seeking do not generalize to natural rewards, as RI of sucrose seeking is not influenced by intra-accumbal infusion of ZIP. The ability of ZIP to disrupt cocaine-induced behavioral plasticity was accompanied by the ability to disrupt cocaine-induced synaptic plasticity. Our electrophysiological data revealed that ZIP rescues cocaine-induced deficits in NMDA-dependent and mGluR5-dependent LTD without affecting LTD in saline-yoked controls.
Intra-accumbal administration of ZIP blunts RI of cocaine seeking in a persistent manner
The present study shows that ZIP infusion into the NAc 24 h or even 1 week before testing can block RI of cocaine seeking, whereas the inactive form (SCR-ZIP) has no behavioral effect. To date, no other study has shown a compound that is able to block cocaine RI in such a long-lasting manner. In contrast to traditional pharmacological manipulations, ZIP is cleared from the brain before the 24 h test (Kwapis et al., 2012); therefore, its effect on RI does not depend upon continuous activity at its binding site. Previous studies have demonstrated long-lasting disruption of spatial memories after ZIP infusion into the hippocampus (Pastalkova et al., 2006; Serrano et al., 2008), taste memory after ZIP infusion in the insular cortex (Shema et al., 2007), fear memory after ZIP infusions in the amygdala (Serrano et al., 2008; Migues et al., 2010) and morphine, cocaine, and food conditioned place preference, after ZIP infusions into the NAc (Li et al., 2011).
However, the effect of ZIP in our experiments is not simply due to the erasure of reward-associated memories, as we do not see effects on sucrose-primed RI. At first, this may seem contradictory to previous findings demonstrating that ZIP is able to disrupt other forms of memory. However, these studies have administered ZIP into different brain regions that play a more general role in memory consolidation, including the hippocampus and the amygdala (Serrano et al., 2008; Madroñal et al., 2010; Migues et al., 2010; Kwapis et al., 2012; Evuarherhe et al., 2014). Previous studies examining infusions of ZIP into the NAc have found disruptions in conditioned place preference for high fat food (Li et al., 2011). Although it is possible these differences are due to the different behavioral paradigms used to examine reward, if we examined RI of food seeking for a more highly palatable food we may find broader effects of intra-accumbal ZIP administration. However, the cocaine-specific effects seen in the current study may provide support for ZIP affecting glutamate plasticity in the NAc. ZIP has been shown to disrupt AMPAR trafficking (Yao et al., 2008; Migues et al., 2010; Yu et al., 2017), and manipulations of AMPAR trafficking in the NAc preferentially affect cocaine-associated behaviors leaving natural reward seeking intact (Anderson et al., 2008; Famous et al., 2008; Briand et al., 2014, 2016).
Effect of ZIP on RI of cocaine seeking is independent of PKMzeta
ZIP was initially designed to inhibit PKMζ and therefore any actions of ZIP were attributed to its actions at PKMζ (Yao et al., 2013). However, recent studies have called the specificity of ZIP into question, demonstrating that ZIP is able to disrupt LTP and memory in PKMζ-KO mice (Lee et al., 2013; Volk et al., 2013). Our results support a PKMζ-independent mechanism for ZIP's actions on cocaine seeking. Others have suggested that the effect of ZIP in PKMζ-KO mice is due to compensatory actions of PKC ι/lambda (Tsokas et al., 2016). Although we do not see any upregulation of PKC ι/lambda (McGrath et al., 2018), we cannot rule this out as a potential mechanism. The current study does provide insight into the mechanism underlying the actions of ZIP as ZIP acts to rescue both NMDAR- and mGluR-mediated LTD, pointing to a mechanism common to these two forms of plasticity. Both forms of LTD ultimately lead to the removal of AMPA receptors from the synapse. Therefore, ZIP may be playing a role in this trafficking.
ZIP application rescues-cocaine induced deficits in synaptic plasticity
Here, we show that the bath application of ZIP leads to recovery of cocaine deficits in LTD within the NAc core. Furthermore, the ability of ZIP to rescue LTD is nonspecific in that we see recovery of both NMDA- and mGluR-mediated forms of LTD. The lack of LTD within the accumbens after cocaine suggests an inability to rescale synaptic transmission when contingencies are updated (Yin et al., 2009; Kasanetz et al., 2010). The ability of ZIP to restore the capacity for plasticity provides a mechanism by which ZIP may allow for the weakening of cue-induced craving. Consistent with this, optogenetic protocols that induce LTD within the accumbens block RI of cocaine seeking (Pascoli et al., 2014).
Both NMDA- and mGluR-mediated LTD require the internalization of the AMPA receptors (Kim et al., 2001; Snyder et al., 2001; Xiao et al., 2001). The trafficking of AMPA receptors after exposure to cocaine, is mediated, in part, by PKC (Famous et al., 2008). Furthermore, inhibition of PKC blocks cocaine-induced behavioral phenotypes, including RI of cocaine seeking (Steketee, 1993; Cervo et al., 1997; Schmidt et al., 2013). ZIP has been shown to inhibit multiple isoforms of PKC and interfere with the subcellular targeting of PKC (Bogard and Tavalin, 2015). Therefore, ZIP may be reversing cocaine-induced increases in PKC-mediated insertion of AMPA receptors. This is supported by the current finding that ZIP reverses cocaine-induced increases in AMPA/NMDA ratio. ZIP is known to disrupt the interaction between atypical PKCs and their protein scaffold, p62, which in turn disrupts the interaction between GluA1 and GluA2 AMPA subunits and p62 (Tsai et al., 2015). As interactions between p62 and GluA1 can mediate AMPA receptor trafficking and subsequent log-term plasticity (Jiang et al., 2009), ZIP's ability to disrupt this interaction could mediate the behavioral and physiological changes we see in the current studies. Although it is not known how cocaine self-administration affects the interaction between atypical PKCs, p62, and AMPARs, morphine and methamphetamine increase the expression of p62, suggesting that drugs of abuse could recruit these AMPAR trafficking mechanisms (Lazzeri et al., 2018; Rodriguez et al., 2019).
The current electrophysiological studies used animals that underwent forced abstinence rather than active extinction of cocaine responding. Although both behavioral procedures are known to disrupt LTD (Moussawi et al., 2009; Briand et al., 2014), extinction does lead to alterations in glutamate plasticity that are not present after forced abstinence. Abstinence leads to an increase in GluA2 expression in the nonsynaptic fraction, potentially suggesting differences in AMPAR trafficking between the two protocols (Knackstedt et al., 2010). However, as there are no differences in AMPAR subunit expression in the synaptic fraction these differences may not influence the forms of plasticity examined in the current studies. In contrast, there are clear differences in mGluR5 surface expression, as well as proteins involved in mGluR5 trafficking, after extinction that are not present after abstinence (Knackstedt et al., 2010). This would suggest that DHPG-induced LTD would also be disrupted after extinction and future work could confirm that ZIP would reverse this plasticity to the same extent as it does in the current study.
At high concentrations, ZIP has been shown to be toxic to cells and lead to cell death in culture (Sadeh et al., 2015). Although a lesion of the NAc could lead to the effects we see on RI behavior, we did not see any histological evidence for this and the specificity of our effect to cocaine versus sucrose suggests this is not the case. Furthermore, we did not see any effect of ZIP on LTD in drug-naive animals, suggesting that the effects of ZIP are more specific than simply generalized toxicity.
Conclusion
The current study establishes that intra-accumbal ZIP administration blocks cue-induced cocaine seeking in a persistent manner while not affecting sucrose seeking. Furthermore, ZIP can rescue two forms of LTD that are disrupted by cocaine self-administration. We suggest that the ability of ZIP to restore the capacity to weaken synaptic input may underlie its behavioral effects.
Footnotes
This work was supported by the National Institute on Drug Abuse (Grants #DA033372 and #DA047265 to L.A.B.) and the Deutsche Forschungsgemeinschaft (Grant #DE 2828/1-1 to A.U.D). We thank Richard L. Huganir for providing the initial breeders for the PKMζ-KO mouse studies.
The authors declare no competing financial interests.
- Correspondence should be addressed to Lisa A. Briand at lbriand{at}temple.edu





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}