

Abstract
It is widely accepted that midbrain dopamine (DA) neurons encode actual and expected reward values by phasic alterations in firing rate. However, how DA neurons encode negative events in the environment is still unclear because some DA neurons appear to be depressed and others excited by aversive stimuli. Here, we show that exposing fear-conditioned rats to stimuli predicting electrical shock elicited three types of biphasic responses, each of which contained an inhibitory pause, in neurochemically identified ventral tegmental area (VTA) DA neurons. The duration of the inhibitory pause in these responses of VTA DA neurons was in direct proportion to the increase in respiratory rate reflecting the level of conditioned fear. Our results suggest that the duration of inhibition of VTA DA neurons encodes negative emotional values of signals predicting aversive events in the environment.
Introduction
Dopamine (DA) neurons located in the ventral tegmental area (VTA) project to the amygdala, prefrontal cortex, nucleus accumbens, and hippocampus (Swanson, 1982; Gasbarri et al., 1994), structures known to be involved in the regulation of aversive behavior. Electrophysiological studies in both anesthetized and conscious animals showed that aversive stimuli either phasically excite or inhibit DA neurons (Mirenowicz and Schultz, 1996; Guarraci and Kapp, 1999; Ungless et al., 2004; Brischoux et al., 2009; Matsumoto and Hikosaka, 2009a), and that the type of response of DA neurons depends, to a certain degree, on their location within the midbrain. In particular, experiments in anesthetized rats have demonstrated that nociceptive electrical stimulation elicits inhibitory responses in neurochemically identified DA neurons located predominantly in the dorsal part of the VTA (Ungless et al., 2004), whereas ventral VTA DA neurons are excited by noxious stimulation (Brischoux et al., 2009). In conscious monkeys, excitatory response to an aversive air-puff and associated conditioned stimuli (CSs) were observed in putative DA neurons located in the substantia nigra pars compacta. This group of putative DA neurons also discharged in response to presentations of reward-predicting stimuli (Matsumoto and Hikosaka, 2009a). In contrast, putative VTA DA neurons, which were excited by rewarding stimuli, showed inhibitory responses to aversive stimuli, suggesting that these neurons participate in the encoding of both positive and negative emotional values (Schultz, 2007; Matsumoto and Hikosaka, 2009a).
It is still unclear how VTA DA neurons encode a broad range of aversive stimuli. Since these DA neurons have a low spontaneous firing rate and cease their activity in response to mild negative events (Mirenowicz and Schultz, 1996), there is the possibility that alterations in the duration of inhibition of DA neurons may encode responses to aversive stimuli, as was shown for negative reward prediction errors in monkeys (Bayer et al., 2007). However, previous studies have demonstrated that neurons electrophysiologically identified as VTA DA neurons showed bidirectional responses to aversive stimuli (Mirenowicz and Schultz, 1996; Guarraci and Kapp, 1999, Matsumoto and Hikosaka, 2009a). These reported excitatory and inhibitory responses of VTA DA neurons might be related to their distribution within the midbrain (Ungless et al., 2004; Brischoux et al., 2009) or their electrophysiological misidentification based solely on spike electrophysiological parameters. Juxtacellular Neurobiotin (Nb) labeling of VTA neurons recorded in anesthetized rats has revealed that a large population of cells that does not express tyrosine hydroxylase (TH), a marker of DA neurons, has spike parameters similar to those of TH-positive neurons (Ungless et al., 2004; Luo et al., 2008; Brischoux et al., 2009). These data clearly demonstrate the need to combine electrophysiological and neurochemical criteria for the accurate identification of VTA DA neurons in conscious animals.
We have developed a novel methodological approach that allows juxtacellular labeling of neurons recorded in behaving rats and their subsequent neurochemical identification. By using this approach, we were able to record responses of neurochemically identified VTA DA neurons to aversive CSs paired with electrical shock in fear-conditioned rats and to establish that the duration of inhibition of these DA neurons is modulated by fear in a monotonic manner.
Materials and Methods
Subjects.
Experiments were performed in 23 male Sprague Dawley rats. Two plastic microdrive hubs, bipolar stimulating, and indifferent electrodes were fixed on the skull under general anesthesia (Equithesin, 3.3 ml/kg, i.p.). Coordinates of the VTA and gigantocellular reticular nucleus were determined with the rat brain atlas of Paxinos and Watson (1997). Chronic EEG and neck EMG electrodes were additionally implanted in six control rats to assess the sleep–wakefulness states during habituation. All procedures and experiments were approved by the Animal Care and Use Committee of the National Institute on Drug Abuse and complied with the Public Health Service Policy on use of laboratory animals.
Training procedures.
Before learning, rats were habituated to light restraint with a fabric hammock for 2–3 d (3–4 h daily). Our criteria for habituation were as follows: low frequency of defecation, lack of strong motor activity and freezing episodes, normal respiratory rate, and ability to fall asleep during restraint. Seventeen rats were fear conditioned using two different tones (10 kHz or 600 Hz, 70 dB, 3 s). The first tone (CS+), paired with subsequent electrical tail shock (0.5–1.2 mA, 60 Hz, 1 s), was delivered during a 30 min session with 50–60 s intertrial intervals. The second tone (CS−), unpaired with shock, was presented 20–30 s after the CS+. Auditory frequencies for the CS+ and CS− were counterbalanced across rats. For comparison, the same tones unpaired with electrical shock were presented to six naive rats habituated to light restraint. The respiratory rate was recorded to evaluate levels of fear evoked by the CSs or control tones (Frysztak and Neafsey, 1991; Ley, 1999; Gallego et al., 2001). For this purpose, a respiratory transducer (a miniature headphone without a cover) was placed between the body of a rat and the fabric hammock. EMG electrodes (50 μm stainless steel wires) were implanted bilaterally in the gastrocnemius muscles to record acoustic startle responses to the CS+ and CS−. Training was stopped when rats accelerated breathing to 10 consecutive CS+ presentations and did not respond to the CS− delivery. On the next day, the additional one or two conditioning sessions (10–15 min) were performed before neuronal recording.
Recording and electrical stimulation.
Recording of spikes and juxtacellular labeling of neurons were performed using Axoclamp 900A (Molecular Devices) and Model 1700 (A-M Systems) amplifiers. Micropipettes were filled with 4% Nb dissolved in 0.5 m potassium acetate. To obtain stable recording and juxtacellular labeling of neurons, we used micropipettes with thick tips (2.0–3.0 μm in diameter) and a sinusoidal shape of injection currents (3–15 nA).
Electrical stimulation of the medial part of the gigantocellular reticular nucleus (Mileykovskiy et al., 2002) immobilized the rat for filling a cell with Nb after a recording session. As an application of standard rectangular stimulating pulses evoked strong electrical artifacts, which impeded the control of cell labeling, we used sinusoidal currents (100–250 μA, 25 Hz, 5–15 min) for electrical stimulation of the gigantocellular reticular nucleus. Agilent 33220A generator (Agilent Technologies) was chosen for stimulation because its output, isolated from the ground, substantially decreased electrical artifacts. Recording of spikes was performed in the frequency band from 100 Hz to 5 kHz. An additional filter (300 Hz–5 kHz) was connected during juxtacellular labeling. The signal/noise ratio >5/1 allowed successful juxtacellular labeling of a single neuron. One or two neurons with similar discharge patterns were labeled on each side of the brain.
Immunohistochemical identification of neurons.
VTA sections (20 μm) were incubated in cyanine 2-conjugated streptavidin (1:200; Jackson ImmunoResearch) overnight. Nb-labeled neurons were visualized by epifluorescence using a Leica DMLB microscope. Sections containing Nb-labeled neurons were incubated for 3 h in a mouse monoclonal anti-TH antibody (1:200; Millipore Bioscience Research Reagents), followed by 3 h incubation in a Texas Red dye-conjugated donkey anti-mouse antibody (1:50; Jackson ImmunoResearch).
Statistical analysis.
The duration of inhibitory pauses of DA neurons, which showed inhibitory responses to the CSs, was calculated from the onset of the inhibitory pause to the appearance of the first spike (Bayer et al., 2007). In three DA neurons showing biphasic excitatory/inhibitory responses, the duration of inhibitory pauses was calculated from the last short latency spike to the appearance of the first spike. The firing rate following the CS+ or CS− termination was determined for 1 s using peristimulus time histograms (100 ms bin) constructed for each DA neuron and compared with the baseline firing rate calculated for 3 s preceding the CS onset. The respiratory rate and firing rate of DA neurons were calculated for 3 s before (baseline) and after turning on CS+, CS−, or control auditory tones. The alterations in respiratory rate evoked by the CSs were indicated as a percentage of the baseline value. Freezing was defined as complete immobility of rats, excepting respiratory movements. Every second following the CS+ or CS− onset, an experimenter evaluated the freezing or not freezing state of the rat. Two-tail probability value was used in all statistical tests. All data are presented as mean ± SEM.
Results
After five to eight training sessions, 12 of the 17 fear-conditioned rats developed differential respiratory responses to the CS+ and CS−, showing a higher increase in respiratory rate to the CS+ than to the CS− (Fig. 1A,C; supplemental Fig. S1A,B, available at www.jneurosci.org as supplemental material). These discriminating rats expressed short freezing episodes after the CS+ onset (7.5 ± 0.6 s, minimum = 4.2 s, maximum = 11.6 s, n = 12) and a restored normal breathing rhythm (1.81 ± 0.04 cycle/s, minimum = 1.54 cycle/s, maximum = 2.2 cycle/s, n = 12) before the subsequent CS− presentation. Five of the 17 rats did not discriminate between the CS+ and CS− and exhibited strong breathing acceleration in response to both stimuli (Fig. 1B,D; supplemental Fig. S1C,D, available at www.jneurosci.org as supplemental material). During intertrial periods, these non-discriminating rats expressed continuous freezing behavior that was characterized by a slow respiratory rhythm (1.17 ± 0.03 cycle/s, minimum = 0.85 cycle/s, maximum = 1.48 cycle/s, n = 5) alternating with short episodes of accelerated breathing.

Alterations in respiratory rate and DA neuronal firing elicited by the CS+ and CS−. A, Differential respiratory responses recorded during the initial six trials in rats (n = 12) discriminating between the CS+ and CS−. Paired t test, t = 5.588, ***p < 0.0002, df = 11. B, Similar increases in respiratory rate evoked by the CS+ and CS− in non-discriminating rats (n = 5) during the initial six trials. Wilcoxon matched-pairs signed-rank test, p = 0.4375. C, D, Representative respiratory and DA neuronal inhibitory responses to the CS+ and CS− recorded in discriminating (C) and nondiscriminating (D) rats. Electromyograms of the left (EMG Gc-L) and right (EMG Gc-R) gastrocnemius muscles. Auditory conditioned stimuli paired (CS+) and unpaired (CS−) with electrical tail shock.
Of 49 VTA neurons recorded during 9–13 trials and submitted to juxtacellular labeling, 32 cells (65%) were successfully entrained by iontophoretic currents, filled with Nb, and visualized in the brain sections. Twenty-three of the 32 VTA neurons expressed TH immunoreactivity, a selective marker of DA, and were predominantly located within the parabrachial pigmented and paranigral nuclei (Fig. 2A–D). These DA neurons were recorded in 23 rats: 15 fear-conditioned rats (10 discriminating and five non-discriminating), two discriminating rats after fear extinction, and six naive rats habituated to light restraint. Nine of the 32 VTA neurons were TH-negative (supplemental Fig. S2, available at www.jneurosci.org as supplemental material).

Neurochemical identification of DA neurons and their location in the VTA. A, A VTA neuron juxtacellularly labeled with Nb. B, This neuron expresses TH immunoreactivity. C, The location of this TH-positive neuron in the ventral part of the VTA. D, Cumulative data showing the location of 23 recorded DA neurons in the VTA. Blue dots, DA neurons inhibited by the CS+; red dots, DA neurons showing biphasic excitatory/inhibitory responses; brown dots, DA neurons recorded in rats after fear extinction; green dots, DA neurons recorded in naive rats habituated to light restraint. E, A triphasic waveform of a DA neuronal spike. F, Modulation of the activity of a DA neuron during juxtacellular labeling with Nb.
All TH-positive neurons had broad triphasic spikes (Fig. 2E) and slow irregular or bursting activity that clearly distinguished them from putative GABA neurons having narrow spikes and high firing rates. These discharge parameters, however, were insufficient to distinguish TH-positive neurons from TH-negative neurons that had slower firing rates and broader spikes than putative GABA neurons (supplemental Fig. S3A–D, available at www.jneurosci.org as supplemental material). Durations of spike negative deflections (Fig. 2E), however, were significantly different between these two cell groups (supplemental Fig. S3E, available at www.jneurosci.org as supplemental material).
Presentations of the CS+ that evoked respiratory acceleration in fear-conditioned rats elicited three types of responses in VTA DA neurons (n = 15). In particular, nine of the 15 DA neurons ceased their discharges in response to the onset of the CS+ (Fig. 3A). The average firing rate of these nine neurons during the CS+ delivery significantly decreased from 4.1 ± 0.5 Hz to 0.75 ± 0.35 Hz (paired t test, t = 9.87, p < 0.00001, df = 8), with the subsequent increase by 42 ± 14% (minimum = 4%, maximum = 65%, n = 9; paired t test, t = 2.58, p = 0.01557, df = 8) compared with baseline. In response to the CS−, the average firing rate of these DA neurons altered insignificantly (4.1 ± 0.5 Hz vs 3.6 ± 0.4 Hz, paired t test, t = 1.01, p = 0.17061, df = 8). Three of the 15 DA neurons decreased their activity to both the CS+ onset and offset (Fig. 3B). The inhibitory responses in these 12 DA neurons to the CS+ onset had an average latency of 78.1 ± 7.0 ms (minimum = 43 ms, maximum = 139 ms) and an average duration of inhibitory pauses of 2.8 ± 0.4 s (minimum = 0.9 s, maximum = 5.9 s). The other three of the 15 DA neurons showed biphasic excitatory/inhibitory responses consisting of two to five short-latency (18–45 ms) spikes followed by inhibitory pauses with durations from 0.5 to 3.7 s (Fig. 3C). The occurrence of spikes in these three neurons correlated with the acoustic startle responses evoked by the CS+ onset.

Three types of responses of VTA DA neurons to the CS+ delivery. A, Spike raster plot and peristimulus time histograms (100 ms bin) illustrating continuous inhibition of DA neurons to the onset of the CS+. B, Spike raster plot and peristimulus time histograms (100 ms bin) illustrating biphasic inhibition of DA neurons to the CS+ onset and offset. C, Spike raster plot and peristimulus time histograms (100 ms bin) illustrating biphasic excitatory/inhibitory responses of DA neurons to the CS+.
To determine whether different levels of fear modulate inhibitory responses of VTA DA neurons, we analyzed their activity in 10 discriminating rats. We found that the CS+ evoking a higher increase in respiratory rate than the CS− elicited significantly longer inhibition of DA neurons (Fig. 4A) and longer inhibitory pauses in their activity (2.47 ± 0.45 s, minimum = 1.28 s, maximum = 3.67 s vs 0.92 ± 0.25 s, minimum = 0.33 s, maximum = 2.09 s; paired t test, t = 3.87, p = 0.0038, df = 9). The DA cell inhibition to the CS+ was followed by a significant (paired t test, t = 7.791, p < 0.0001, df = 9) increase in firing rate (42 ± 6%, minimum = 15%, maximum = 69%, n = 10) compared with the baseline level. This rebound excitation of DA neurons lasted ∼1 s after the CS+ offset (Fig. 4A) and was not observed after the CS− termination (paired t test, t = 1.748, p = 0.1144, df = 9).

The duration of inhibition of VTA DA neurons depends on a level of conditioned fear. A, Population peristimulus time histograms (100 ms bin) illustrating the responses of 10 DA neurons to the CS+ and CS− during the initial six trials that were characterized by stable respiratory responses to the CS+ in discriminating rats. B, The correlation between the duration of inhibitory pauses in the activity of DA neurons (n = 10) and the increase in respiratory rate during presentations of the CS+ (7–13 trials) evoking partial extinction of respiratory responses in discriminating rats. The correlation is expressed by a linear regression: y = 0.3398 + 0.0792x; F = 160.05, p < 0.0001, df = 1, 67. C, The correlation between the duration of inhibitory pauses in the activity of a DA neuron and the increase in respiratory rate in one non-discriminating rat (y = 0.92070 + 0.03207x; F = 13.02, p = 0.0112, df = 1, 8). D, The lack of correlation between the duration of inhibitory pauses in the activity of DA neurons (n = 4) and the increase in respiratory rate in four non-discriminating rats (y = 1.694 + 0.007x; F = 1.180, p = 0.2837, df = 1, 41). Each black square represents one trial without shock delivery.
To study the relationship between the within-session fear extinction and inhibition of DA neurons, we estimated the correlation between respiratory rates and the duration of inhibitory pauses in the activity of DA neurons during consecutive presentations of the CS+ evoking a gradual decrease in respiratory responses (supplemental Fig. S1A, trials 7–13, available at www.jneurosci.org as supplemental material). We found that the correlation between the duration of inhibitory pauses in DA neural activity and the increase in respiratory rate elicited by the delivery of the CS+ was well described by a linear function with a positive slope (Fig. 4B). In the group of rats (n = 5) that did not discriminate between the CSs and expressed continuous freezing behavior, one rat showed a positive correlation between the duration of inhibitory pauses of a DA neuron and the increase in respiratory rate during CS+ presentations (Fig. 4C). In the other four non-discriminating rats, this correlation was insignificant because of the lack of extinction of respiratory responses to the CS+ delivery and relatively stable inhibitory pauses in responses of DA neurons (Fig. 4D). Importantly, the average duration of inhibitory pauses in responses of DA neurons to the CS+ during the initial six trials was significantly shorter in discriminating rats (n = 10) than in rats (n = 5) that did not distinguish between the CSs (2.47 ± 0.45 s, minimum = 1.28 s, maximum = 3.67 s vs 3.2 ± 0.32 s, minimum = 2.8 s, maximum = 4.72 s; Mann–Whitney test, p = 0.037).
To further validate a correlation between inhibition of DA neurons and conditioned fear, we recorded the activity of VTA DA neurons in two rats after fear extinction and six naive rats habituated to light restraint. We found that presentations of the CS+ after extinction of respiratory responses did not elicit inhibition of DA neurons (Fig. 5). Similarly, the delivery of auditory stimuli unpaired with electrical shock did not change either the breathing rhythm or firing rate of DA neurons recorded in naive rats after habituation to light restraint (Fig. 6).
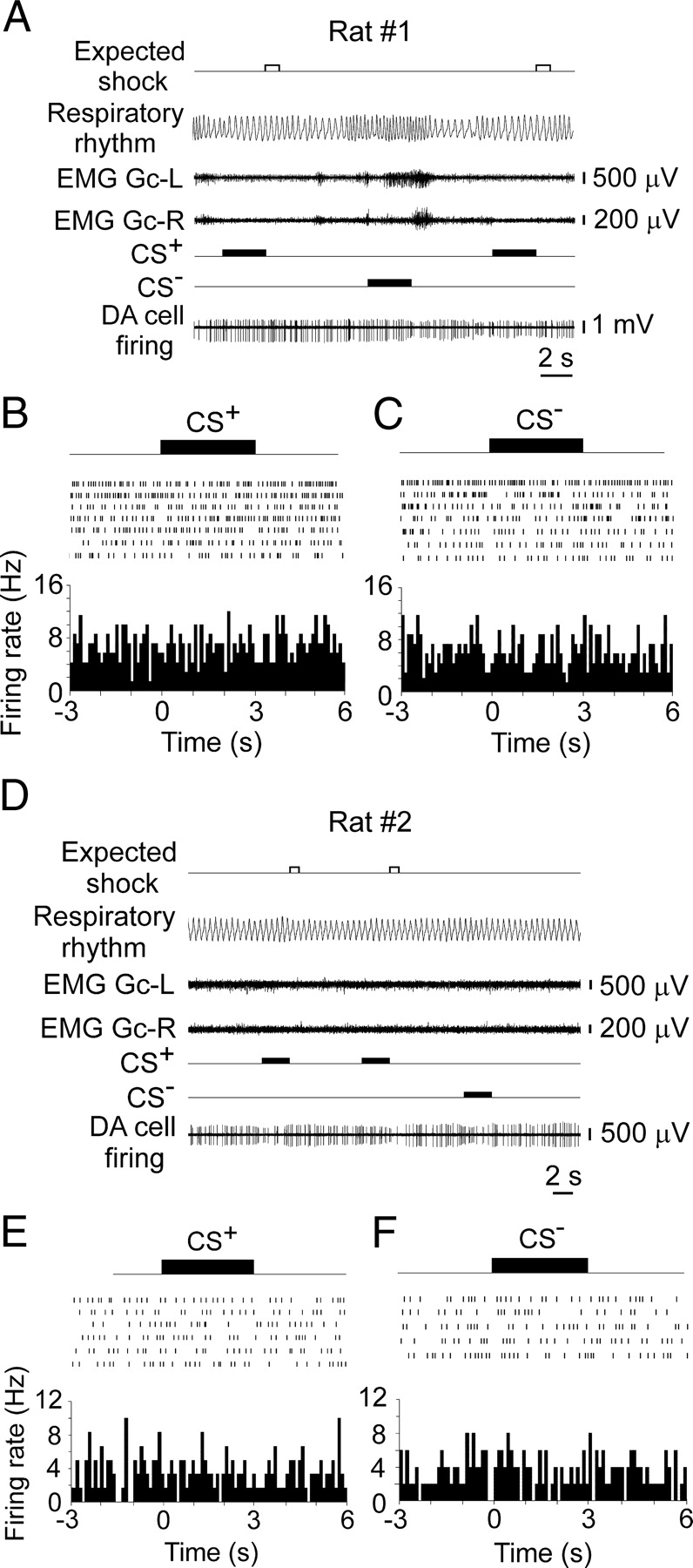
After fear extinction, neither the CS+ nor CS− presentations altered the discharge activity of VTA DA neurons or the respiratory rate in rats. A, D, Lack of respiratory acceleration and inhibitory responses of DA neurons to the CS+ or CS− presentations. B, C, E, F, Spike raster plots and peristimulus time histograms illustrating the lack of responses of these neurons to the CS+ or CS− during consequent trials. EMG Gc-L, Electromyograms of the left gastrocnemius muscles; EMG Gc-R, electromyograms of the right gastrocnemius muscles.

Presentations of auditory stimuli unpaired with electrical shock do not elicit the acceleration of a respiratory rhythm or inhibitory responses in DA neurons recorded in naive rats habituated to light restraint. A, The first presentation of the 10 kHz tone (70 dB) during drowsiness elicited EEG desynchronization and weak acceleration of the respiratory rhythm without virtual inhibition of a DA neuron. The subsequent presentation of the 600 Hz tone (70 dB) did not change either the respiratory rhythm or the baseline DA cell firing. B, The average respiratory rates recorded in six control rats before (baseline) and during the presentation of 10 kHz or 600 Hz tones. One-way ANOVA, F = 0.6472, p = 0.5375, df = 2, 15. C, The average interspike intervals calculated for 3 s before (baseline) and after the onset of 10 kHz or 600 Hz tones for six DA neurons recorded in six control rats. One-way ANOVA, F = 0.5281, p = 0.6003, df = 2, 15.
Discussion
We found that the CSs associated with electrical shock evoked three types of biphasic responses in neurochemically identified VTA DA neurons. The presence of inhibitory pauses in these responses suggests that the transient suppression of DA neural activity is specifically related to acute fear evoked by the delivery of the CS+. Indeed, we found that the CS+ elicited significantly longer inhibition of DA neurons and higher acceleration of breathing than the CS−. Furthermore, the duration of inhibitory pauses of DA neurons was in direct proportion to the increase in respiratory rate. It has been demonstrated that acceleration of breathing as well as the elevation in blood pressure and heart rate reflect an increased level of fear in rats during anticipation of electrical shock (Frysztak and Neafsey, 1991, 1994). Thus, our results suggest that VTA DA neurons are involved in distinguishing between the CSs having different aversive values, and that the duration of inhibition of DA neurons encodes a level of fear evoked by aversive stimuli. In four of the five freezing rats, which did not discriminate between the CS+ and CS−, the linear correlation between the duration of inhibitory pauses of DA neurons and the increase in respiratory rate was not revealed because of the lack of within-session fear extinction. These rats expressed continuous freezing behavior and strongly accelerated breathing in response to each CS+ delivery. We suggest that the relatively stable duration of inhibitory pauses, which we detected in the responses of DA neurons to consequent CSs in rats expressing continuous freezing behavior, encodes intense persisting fear in this group of animals. It was previously shown that rats highly sensitive to stress are characterized by prolonged freezing behavior in response to a tone predicting electrical shock, and that this freezing behavior is resistant to extinction (Shumake et al., 2005). In our study, fear conditioning was performed in lightly restrained rats; therefore, we cannot exclude the possibility that additional stress related to partial immobilization might modulate behavioral, respiratory, and DA neural responses to the CSs. Indeed, a body of evidence demonstrates that the preexposure of animals to stressful events, including restraint, substantially facilitates subsequent classic fear conditioning (for review, see Shors, 2004).
In our experiments, the inhibition to the CS+ observed in the majority of VTA DA neurons was followed by a transient increase in their firing rate. Short-term excitation of neurochemically confirmed DA neurons was also reported in anesthetized rats after termination of nociceptive stimulation inhibiting their discharge activity (Brischoux et al., 2009). The transient increase in DA cell firing that follows inhibition may explain the elevation in DA release detected by microdialysis in the medial prefrontal cortex and limbic structures in response to aversive unconditioned and conditioned stimuli (Abercrombie et al., 1989; Puglisi-Allegra et al., 1991; Kalivas and Duffy, 1995). Since the microdialysis technique lacks high temporal resolution, brief reductions in DA release corresponding to inhibitory DA cell responses to aversive stimuli have not been detected in these experiments. A recent study using fast scan cyclic voltammetry have shown that aversive taste stimulation elicited a sharp decrease in the DA level in the nucleus accumbens for 4–5 s (Roitman et al., 2008).
Different salient events, such as novel, unexpected, and high-intensity sensory stimuli, evoke fast phasic excitatory responses in DA neurons (Horvitz et al., 1997; Horvitz, 2000; Schultz, 2007; Redgrave et al., 2008). We found that a small group of VTA DA neurons (n = 3) exhibited biphasic excitatory/inhibitory responses to aversive CSs. Since spikes elicited in these DA neurons had very short latencies (18–45 ms), excitatory responses might reflect sensory or motivational salience of the CSs, whereas inhibitory pauses following short-latency excitation may encode the negative emotional value of the CSs (for review, see Bromberg-Martin et al., 2010). Two of the three DA neurons were located in the parabrachial pigmented nucleus, and the third neuron was situated in the paranigral nucleus of the VTA. Brischoux et al. (2009) showed the locations of four of five DA neurons excited by stimulation of nociceptive inputs in anesthetized rats in the paranigral pigmented and parainterfascicular nuclei of the VTA and one neuron in the parabrachial pigmented nucleus. However, the sample sizes in both studies are insufficient to make any conclusion about the predominant locations of these groups of DA neurons in the VTA.
It is well documented that DA neurons are involved in forming positive and negative prediction error signaling in reward-related paradigms (Mirenowicz and Schultz, 1996; Bayer et al., 2007; Schultz, 2007; Matsumoto and Hikosaka, 2009a). These neurons transiently increase their activity if the reward exceeds expectations and decrease or stop firing if the reward is omitted or has less value than expected. Bayer et al. (2007) reported that pauses in the activity of putative DA neurons became longer for more negative reward prediction errors and that this correlation had a largely linear character. Together with our results, these data suggest the involvement of similar neural circuits in monotonic encoding of both the magnitude of negative reward prediction error and the negative emotional value of the CSs. Indeed, recent studies show that inhibition of DA neurons during reward omission and presentation of aversive stimuli may be mediated by neurons of the lateral habenula (Matsumoto and Hikosaka, 2009b) and rostromedial tegmental nucleus (Jhou et al., 2009). Neurons in these structures showed excitatory responses to both reward omission and aversive stimulation and decreased their firing in response to rewarding stimuli.
It was hypothesized that the increase in DA signaling is associated with the motivated go process or action (Romo and Schultz, 1990; Schultz and Romo, 1990; Ljungberg et al., 1992). If so, the transient decrease in DA concentration evoked by inhibition of VTA DA neurons in response to aversive stimuli might facilitate behavioral arrest during processing of sensory signals associated with fear. In contrast, long-lasting inhibition of DA cell activity evoked by events with excessive negative values might reflect transient disintegration of behavior and the loss of adaptive responding to environmental stimuli.
Inhibition of VTA DA neurons by negative CSs also suggests the participation of these neurons in forming an aversive teaching signal mediating avoidance learning (Johansen and Fields, 2004) via interaction with the lateral habenular (Matsumoto and Hikosaka, 2009b) and rostromedial tegmental nucleus (Jhou et al., 2009). A recent study (Brinschwitz et al., 2010) showed that lateral habenular glutamate neurons may convey aversive/negative information to rostromedial tegmental GABA neurons that inhibit VTA DA neurons (Jhou et al., 2009). Transient reduction in DA signaling, in turn, may affect different brain systems mediating aversive learning.
Thus, our results show that the duration of inhibition of VTA DA neurons encodes a level of fear evoked by the CSs in a monotonically increased manner in fear-conditioned rats. The ability of VTA DA neurons to encode stimuli with different negative emotional values suggests the participation of these neurons in the analysis of signals that predict the degree of aversiveness of events in the environment.
Footnotes
This research was supported by the Intramural Research Program of the National Institute on Drug Abuse. We thank Dr. Roy Wise for comments to this manuscript.
The authors declare no competing financial interests.
- Correspondence should be addressed to either Boris Y. Mileykovskiy or Marisela Morales, National Institute on Drug Abuse, Intramural Research Program, Neuronal Network Section, 251 Bayview Boulevard, Baltimore, MD 21224. mileykob{at}nida.nih.gov or mmorales{at}intra.nida.nih.gov
This article is freely available online through the J Neurosci Open Choice option.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}