

Abstract
Agonists acting at α2 adrenergic and opioid receptors have analgesic properties and act synergistically when co-administered in the spinal cord; this synergy may also contribute to the potency and efficacy of spinally administered morphine. The lack of subtype-selective pharmacological agents has previously impeded the definition of the adrenergic receptor subtype(s) mediating these effects. We therefore exploited a genetically modified mouse line expressing a point mutation (D79N) in the α2a adrenergic receptor (α2aAR) to investigate the role of the α2aAR in α2 agonist-evoked analgesia and adrenergic–opioid synergy. In the tail-flick test, intrathecal administration of UK 14,304, a nonsubtype-selective α2AR agonist, had no analgesic effect in D79N mice, whereas the analgesic potency of morphine (intrathecal) in this assay was not affected by the mutation. The mutation also decreased α2-agonist-mediated spinal analgesia and blocked the synergy seen in wild-type mice with both the δ-opioid agonist deltorphin II and the μ-opioid agonist [d-ALA2,N-Me-Phe4,Gly-ol5]-Enkephalin (DAMGO) in the substance P behavioral test. In addition, the potency of spinally administered morphine was decreased in this test, suggesting that activation of descending noradrenergic systems impinging on the α2aAR contributes to morphine-induced spinal inhibition in this model. These results demonstrate that the α2aAR subtype is the primary mediator of α2 adrenergic spinal analgesia and is necessary for analgesic synergy with opioids. Thus, combination therapies targeting the α2aAR and opioid receptors may prove useful in maximizing the analgesic efficacy of opioids while decreasing total dose requirements.
- α2 adrenergic receptor
- synergy
- mice
- morphine
- antinociception
- intrathecal
- substance P
- opioid
- isobologram
- gene targeting
- α2aAR
α2 adrenergic receptors (α2ARs) mediate a number of physiological responses, including analgesia (Yaksh, 1985). In addition, α2 adrenergic agonists interact synergistically with opioid agonists (Drasner and Sullivan et al., 1987; Wilcox et al., 1987; Fields, 1988; Ossipov et al., 1989; Monasky et al., 1990), a property important in clinical pain management, because synergy-enabled decreases in dose may minimize side effects (Eisenach et al., 1994) and may be effective in the treatment of chronic, opioid-insensitive pain states (Coombs et al., 1986). α2ARs belong to the superfamily of seven-transmembrane spanning domain G-protein-coupled receptors and share common signal transduction pathways mediated through the pertussis toxin-sensitive inhibitory G-proteins Gi and Go (Crain et al., 1987; Hoehn et al., 1988). Activation of α2ARs can decrease neuronal excitation by opening inwardly rectifying potassium channels (Surprenant and North, 1988), by decreasing presynaptic calcium influx (Surprenant et al., 1990), and by inhibiting adenylyl cyclase (Limbird, 1988; Uhlen and Wikberg, 1988). Three subtypes of α2ARs have been cloned in human and rat, corresponding to the pharmacological subtypes α2a, α2b, and α2c, respectively (Bylund et al., 1994).
Determination of the relative contributions of the three α2AR subtypes to spinal adrenergic analgesia and adrenergic–opioid synergy has been difficult because of the lack of subtype-selective pharmacological agents. In situhybridization studies have localized mRNA for both the α2aAR and α2cAR subtypes in dorsal root ganglion neurons (Nicholas et al., 1993). Thus, one or both subtypes may mediate spinal analgesia at a presynaptic site on primary afferent fibers. Pharmacological studies have suggested that the activation of α2aARs mediates α2agonist-invoked analgesia (Millan, 1992; Millan et al., 1994), whereas others have suggested that the site of action may be at either α2aARs or non-α2aARs depending on the agonist used (Takano and Yaksh, 1992). In addition, adrenergic agonists have been shown to inhibit neurotransmitter release from spinal cord preparations by a prazosin-sensitive receptor, suggesting a role for the α2bAR or α2cAR subtypes (Ono et al., 1991). As is the case for the direct analgesic effects of α2 adrenergic agonists, the α2AR subtype(s) responsible for synergy with the opioid system also has (have) not been established.
We therefore sought to test the involvement of α2aARs in spinal adrenergic analgesia and adrenergic–opioid synergy using a mouse line developed by hit-and-run gene targeting that expresses a point mutation, D79N, in the α2aAR (MacMillan et al., 1996). The mouse line manifests not only an 80% reduction in functional α2aAR binding (MacMillan et al., 1996), but also a lack of coupling to both K+ and Ca2+ channels (Lakhlani et al., 1996), suggesting that D79N mice can be viewed as a functional knockout useful for evaluating the role of the α2aAR subtype in spinal inhibition by α2 adrenoceptor agonists in vivo.
MATERIALS AND METHODS
Animals. All experimental animals were housed in groups of 5–10 in a temperature- and humidity-controlled environment. Animals were maintained on a 12 hr light/dark cycle and had unlimited access to food and water. The D79N mice were generated by hit-and-run gene targeting as described previously (MacMillan et al., 1996) on a hybrid C57BL/6 and 129/Sv background, hereafter designated B6,129. Wild-type (WT) B6,129 mice were used as control animals. Breeding pairs were established, and pups were weaned at between 2 and 3 weeks of age. To control for genetic drift, all studies were performed on generation-matched animals pair-bred in our facility. Animals were used when they were between 6 and 8 weeks of age. Within each experiment, the animals were age- and gender-matched across all groups. All experiments were approved by the Institutional Animal Care and Use Committee of the University of Minnesota.
Drug preparation and administration. Drugs used were morphine sulfate (gift of Dr. R. P. Elde, University of Minnesota); UK 14,304 (gift of the Pfizer Drug Company); dexmedetomidine (gift of Zeneca Pharmaceuticals); prazosin, idaxozan, and substance P (SP) from Sigma (St. Louis, MO); deltorphin II and [d-ALA2,N-Me-Phe4,Gly-ol5]-Enkephalin (DAMGO) from Research Biochemicals International (Natick, MA); and clonidine from Boehringer-Ingelheim Ltd. All drugs were dissolved in 0.9% saline and administered intrathecally in a volume of 5 μl according to the method of Hylden and Wilcox (1980) as modified byWigdor and Wilcox (1987).
Thermal nociceptive testing. Thermal nociceptive responsiveness was assessed using the warm water (52.5°C) tail-immersion assay, as described previously (Janssen et al., 1963). Briefly, mice were gently wrapped in a soft cloth such that their tails were exposed, and three-quarters of the length of the tail was dipped into the hot water. Tail-flick latencies were obtained before drug application to establish a baseline response. Drugs were then injected intrathecally, and post-treatment latencies were measured. In some cases, tail-flick latencies were determined every 15 min for 1 hr to determine the time course of the antinociceptive effect. A maximum cut-off of 12 sec was set to avoid tissue damage. The results were then expressed as a percent of the maximum possible effect (%MPE) according to the equation:
 When dose–response relationships were assessed, at least three doses of each agonist were used. With the exception of the time course study, animals were tested 10 min after intrathecal drug administration. Dose–response relationships were determined as described below.
When dose–response relationships were assessed, at least three doses of each agonist were used. With the exception of the time course study, animals were tested 10 min after intrathecal drug administration. Dose–response relationships were determined as described below.
SP behavioral assay. A constant dose (10 ng) of SP was administered intrathecally in a volume of 5 μl, and the number of caudally directed biting, licking, and scratching behaviors was counted for 1 min after the injection as described previously (Hylden and Wilcox, 1981). For each experimental day, a new control count was obtained, and percentage of inhibition was determined relative to that control. Control counts typically ranged from between 30 and 40 behaviors per minute. A minimum of six mice were used for each drug or combination dose. To assess the effect of opioid and adrenergic agonists, agonists were co-administered with SP, and inhibition was expressed as a percent of the mean response of the control group according to the following equation:
 To evaluate interactions between agonists, mixtures were co-administered with SP. In some experiments, antagonists were co-administered with the agonist–SP mixtures. Dose–response relationships were determined as described below. A minimum of six mice were used for each drug or combination dose.
To evaluate interactions between agonists, mixtures were co-administered with SP. In some experiments, antagonists were co-administered with the agonist–SP mixtures. Dose–response relationships were determined as described below. A minimum of six mice were used for each drug or combination dose.
Data analysis. The ED50 values and 95% confidence intervals (CIs) of drugs in nanomoles were calculated using the graded dose–response curve method of Tallarida and Murray (1987). A minimum of three doses were used for each drug or drug combination. In some instances, only the linear portion of a dose–response curve was included in the ED50 calculation. To determine differences in agonist or antagonist potency between treatment groups, nonoverlapping 95% CIs were considered to represent statistically significant differences. When the extent of a potency shift between treatment groups was calculated, a potency ratio representing the ratio of the respective ED50 values was calculated. In the synergy studies, dose–response curves, ED50 values, and 95% CIs were first generated for each agent alone, as described above. The antinociceptive agents were then co-administered at a constant dose ratio based on the potency ratio of the two agents. For example, if Drug A had an ED50 of 1 nmol and Drug B had an ED50 of 10 nmol, the drugs would be co-administered in a 1:10 ratio and a third dose–response curve would be generated for the combination treatment.
Two statistical methods, one graphical and one numeric, are commonly used to test for significance of nonadditive drug combination interactions. Isobolographic analysis, the graphical method, has been described previously (Tallarida et al., 1986; Tallarida, 1992). Briefly, ED50 values, obtained when the two agents are administered separately, represent the x and yintercepts. Agents that interact in an additive fashion should fall on a theoretical additive line connecting these intercepts. Experimental ED50 values and 95% CIs for a drug combination are then superimposed on the isobologram. Values that fall below the theoretical additive line and outside the lower 95% CIs are considered synergistic.
The second, numeric method for evaluating drug interactions has also been described previously by Tallarida et al. (1986) and adapted byOssipov et al. (1997). To test for synergistic interactions via this method, the 95% CIs of all dose–response curves are arithmetically arranged around their respective ED50 values using the equation ln(10) × ED50 × SE(log ED50), where SE is the standard error. A theoretical additive line and its 95% CI are then calculated based on the dose–response curves of the drugs administered separately. This theoretical value is then compared with the experimental combined ED50. An interaction is considered synergistic if the observed ED50 value is significantly less than the theoretical additive (p < 0.05).
For the sake of simplicity, this paper reports only results obtained using the graphical method of analysis; however, the data were processed using the numeric method with similar results.
RESULTS
The α2aAR is required for α2AR agonist-mediated thermal antinociception
To assess whether reducing α2aAR function would influence the potency or efficacy of α2AR agonists in spinal analgesia, we evaluated the effect of α2AR agonist UK 14,304 (bromonidine) administered intrathecally in the hot water tail-flick assay (Janssen et al., 1963) in WT and D79N mice (Fig.1). UK 14,304 (3 nmol, i.t.) produced long-lasting antinociception in the WT animals that was not apparent at this same dose (3 nmol, i.t.) or at a much higher dose (100 nmol, i.t.) in the D79N mice. Tail-flick latencies were slightly shortened by UK 14,304 in the D79N mice. These findings not only demonstrate that spinal α2aARs play an important role in the antinociceptive effect of UK 14,304 in the hot water tail-flick test, but they also suggest that other UK 14,304-binding receptors may contribute to nociceptive effects in WT animals that are masked by the dominant antinociceptive effects of the α2aARs.

Inhibition of thermal nociceptive behaviors by α2AR agonists in D79N and WT mice. A, Comparison of WT and D79N mice in the hot water tail-flick test. Administration of the α2AR agonist UK 14,304 (3.0 nmol, i.t.) produced long-lasting antinociception in WT animals. In D79N mice, however, neither a 3.0 nor a 100 nmol dose of UK 14,304 was antinociceptive. Baseline tail-flick latencies did not differ between the two strains (see time = 0). Error bars represent ±SEM for each dose point (n = 6–10 animals/dose).
To ascertain whether the D79N animals were responsive to the antinociceptive actions of opioids in this assay, we examined the effects of morphine administered intrathecally on the tail-flick latencies in WT and D79N mice. In contrast to the lack of antinociceptive efficacy observed for UK 14,304 in D79N mice, no difference in morphine potency was observed in these animals as compared with WT (Fig. 2). This result demonstrates that one nonadrenergic antinociceptive pathway is unchanged in the D79N animals and furthermore that α2aARs are not required for morphine to produce antinociception in this test.

Inhibition of thermal nociceptive behaviors by morphine in D79N and WT mice. Comparison of WT and D79N mice in the hot water tail-flick test. Intrathecal administration of morphine produced dose-related antinociception in both WT and D79N animals. The ED50 for morphine in WT animals (0.52 nmol; 95% CI = 0.36–0.74) was not significantly different from that observed in D79N animals (ED50 = 0.53 nmol; 95% CI = 0.27–1.0). Error bars represent ±SEM for each dose point (n = 6–10 animals/dose).
The α2aAR mediates α2AR agonist-induced inhibition of SP-elicited behavior
To determine whether lack of efficacy observed in the tail-flick assay was specific for thermal stimuli, we also examined the effects of α2AR agonists in the SP behavior test (Hylden and Wilcox, 1981). SP is an excitatory neuropeptide that mediates nociceptive transmission and serves as a co-transmitter with glutamate in small-diameter primary afferent neurons and their terminals (Battaglia and Rustioni, 1988; De Biasi and Rustioni, 1988). After intrathecal administration, SP elicits a stereotypical, caudally directed, biting and scratching behavior in mice (Hylden and Wilcox, 1981). Like most spinally acting analgesics, agonists acting at α2ARs have been shown to inhibit the excitatory action of the neurokinin SP (Hylden and Wilcox, 1983). Behavior elicited by intrathecally administered SP has been shown to be a reliable, indirect measure of nociception (Wilcox, 1988). The α2AR agonists UK 14,304 (Fig. 3A) and dexmedetomidine (Fig. 3B) inhibited SP-elicited behavior in a dose-dependent manner in both D79N mice and the corresponding WT control mice. The ED50 values for UK 14,304 and dexmedetomidine were increased ∼250- and 2500-fold, respectively, in D79N mice compared with WT animals. At supramaximal doses of agonist, however, near-maximal efficacy was achieved in the mutant animals.

Inhibition of SP-elicited behavior by α2AR agonists in D79N and WT mice. A, UK 14,304 inhibited SP-elicited behavior in a dose-dependent manner in both D79N and WT mice. The ED50 for UK 14,304 increased >250-fold in D79N mice (95 nmol; 95% CI = 58–158) compared with WT (0.37 nmol; 95% CI = 0.21–0.65). B, SP-elicited behavior was inhibited by dexmedetomidine in a dose-dependent manner in both WT (ED50 = 0.014 nmol; 95% CI = 0.008–0.025) and D79N (ED50 = 35 nmol; 95% CI = 24–51) mice; however, a 2500-fold decrease in agonist potency was observed in the D79N animals. Error bars represent ±SEM for each dose point (n = 6–10 animals/dose).
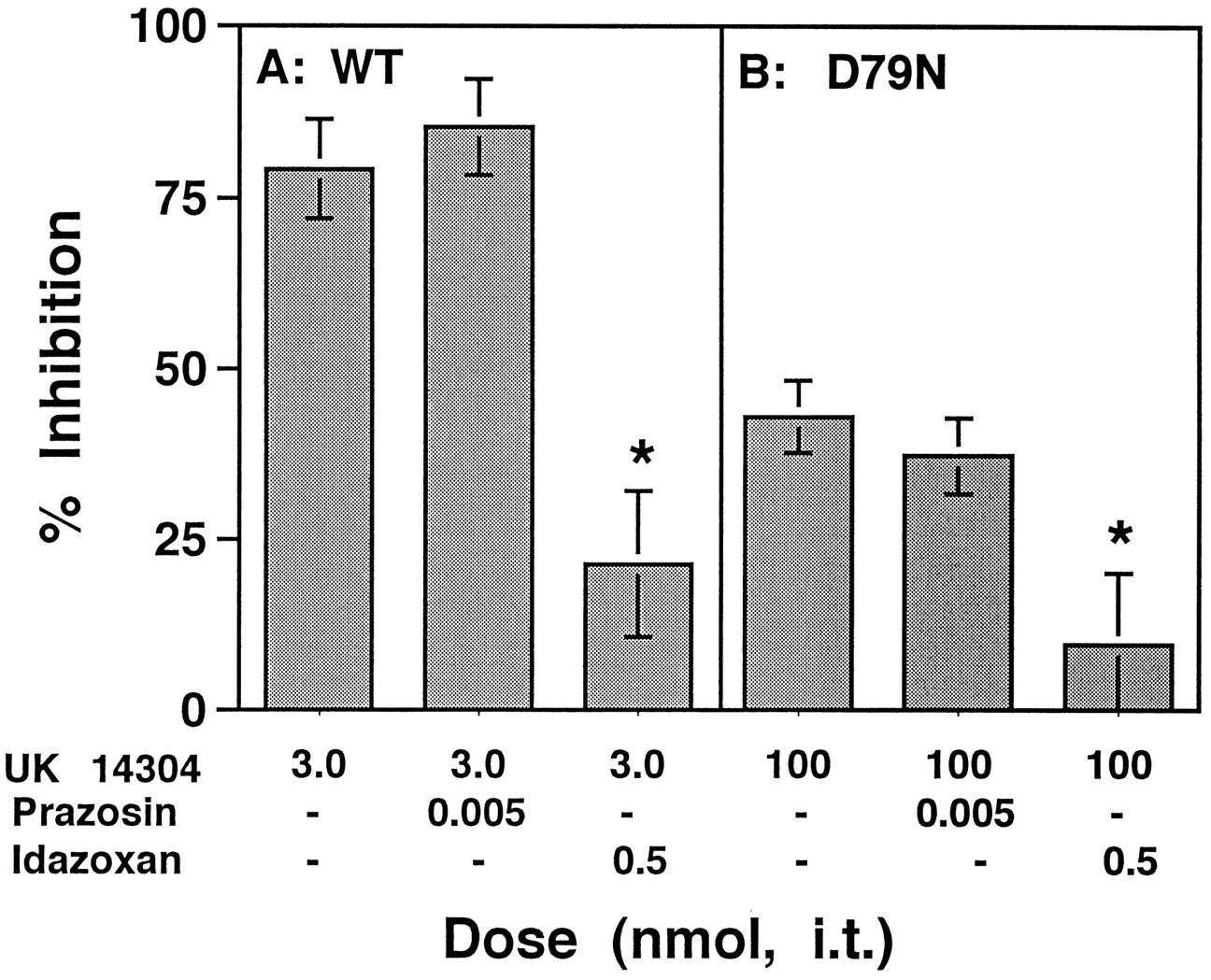
To further clarify which α2AR subtype mediates the inhibition of SP-elicited behavior, we co-administered UK 14,304 with prazosin, which blocks α1ARs as well as the α2bAR and α2cAR subtypes (Bylund et al., 1994). The presence of prazosin (0.5 pmol, i.t.) failed to antagonize UK 14,304 in either D79N or WT animals (Fig.4), corroborating the finding that the effects of UK 14,304 are α2aAR-mediated. This dose of prazosin is in the range used in other studies (Howe et al., 1983). Idazoxan, an antagonist effective at all α2AR subtypes (Bylund et al., 1994), attenuated the effect of UK 14,304 in both WT and mutant animals (Fig. 4). The antagonism by idazoxan was dose-related in both strains. The IC50 values were 0.14 nmol (95% CI = 0.07–0.31) in WT and 0.013 nmol (95% CI = 0.001–0.32) in D79N, values that were not significantly different. Idazoxan alone had no effect over the entire dose range tested (data not shown). These results verify the interpretation that activation of the α2aAR subtype is sufficient for inhibition of SP-elicited behavior in vivo and suggests that residual activity of the mutant α2aAR may be responsible for the efficacy of these ligands at higher doses in the D79N animals; alternatively, analgesic effects of UK 14,304 at the α2bAR and α2cAR subtypes, at the supramaximal concentrations used in the D79N mice, may have surmounted any antagonism by prazosin present at these sites.

Selective antagonism of the analgesic effects of UK 14,304 by idazoxan but not by prazosin. A, UK 14,304 (3.0 nmol, i.t.) inhibited SP-elicited behavior in WT animals (left column). Prazosin, an antagonist at α1AR as well as the α2bAR and α2cAR subtypes, failed to antagonize the inhibitory effects of UK 14,304 (middle column), but the nonsubtype-selective α2AR antagonist idazoxan significantly attenuated the action of UK 14,304 (right column) in these animals. B, The inhibitory action of 100 nmol UK 14,304 (left column) was not altered by co-administration of prazosin in D79N mice (middle column), whereas idaxozan antagonized UK 14,304 in these animals (right column). Antagonism by idazoxan was dose-related in both WT and D79N mice. The IC50 values were 0.14 nmol (95% CI = 0.07–0.31) in WT and 0.013 nmol (95% CI = 0.001–0.32) in D79N and were not significantly different. Error bars represent ±SEM for each dose point (n = 6–10 animals/dose).
The α2aAR is necessary for synergy to occur between UK 14,304 and μ- or δ-opioid receptor agonists
When agonists to both α2AR and opioid receptors are co-administered with SP, they act synergistically to inhibit SP-elicited behavior (Hylden and Wilcox, 1983; Roerig et al., 1992). Previous work has shown that activation of δ-opioid receptors is necessary for this synergy in the mouse (Roerig et al., 1992). Because the relative contributions of the α2AR subtypes to α2–opioid synergy are unclear, we tested whether α2aAR activation is necessary and sufficient for α2 adrenergic–δ-opioid synergy. We administered either deltorphin II, a δ-opioid receptor agonist, or an α2aAR-selective cocktail (UK 14,304 + 5 pmol prazosin; hereafter referred to as UK + P) or both, and constructed dose–response curves for inhibition of SP-elicited behavior (Fig.5A). In WT mice, application of either UK + P or deltorphin II alone inhibited the behavior in a dose-dependent manner, and the combination treatment (1:1 molar agonist ratio) was 10-fold more potent than either drug given alone. Isobolographic analysis of the dose–response data from WT animals indicated a synergistic interaction (Fig. 5B), manifested by the effect of the combined agents falling significantly below the predicted line for an additive drug interaction. In D79N mice, the deltorphin II dose–response curve was indistinguishable from that in WT mice, whereas (as shown in Fig. 3) the potency of UK + P was 100-fold lower in D79N than WT mice. Because of the fact that UK + P was 100-fold less potent in the D79N animals, we used a 1:100 (deltorphin II/UK + P) dose ratio in these animals to maintain an equal potency ratio between compounds. The co-administration of deltorphin II with UK + P (1:100 molar agonist ratio) in D79N mice did not significantly alter the potency of either drug when given alone (Fig.5C). Isobolographic analysis of these dose–response curves revealed that the interaction between the two agonists in mutant mice is not synergistic but additive (Fig. 5D). This observation indicates that decreasing the functional efficacy of the α2aAR eliminates its ability to synergize with δ-opioid ligands. These findings also show that selective activation of the α2aAR subtype is sufficient to mediate the synergistic effect of the α2AR agonist UK 14,304 on δ-opioid-mediated antinociception.

Co-administration of UK 14,304 (+ 5 pmol prazosin) and deltorphin II is synergistic in WT but not in D79N mice.A, SP-elicited behavior was challenged by intrathecal administration of either deltorphin II or UK 14,304 + 5 pmol prazosin (UK + P) or both in WT mice. UK + P (squares) and deltorphin II (circles) inhibited the behavior in a dose-dependent manner with similar potency and efficacy. When both UK 14,304 and deltorphin were co-administered, a constant potency ratio (1:1 molar agonist ratio) was maintained. The combination treatment (triangles) was ∼10-fold more potent than either drug given alone, an indication of a synergistic interaction. The abscissa for the combined treatment dose–response curves represent the dose of UK 14,304 in the presence of an equal potency ratio of deltorphin II. B, Isobolographic analysis was applied to the data from Figure 5A. They-intercept represents the ED50 (0.24 nmol; 95% CI = 0.09–0.63) for UK + P, and thex-intercept represents the ED50 (0.42 nmol; 95% CI = 0.21–0.87) for deltorphin II when each was administered alone for inhibition of SP-elicited behavior in WT mice. Theheavy line connecting the intercepts is the theoretical additive line. Coordinates for drug combinations falling below this line and outside the confidence limits indicate synergy. When the two compounds were co-administered in WT animals, the resultant ED50 (0.021 nmol; CI = 0.016–0.028) of UK + P in the presence of deltorphin II fell well below the additive line, indicating a synergistic interaction. Error bars parallel to each axis represent the lower 95% CI for each compound. The error bars on the combined dose point represent the upper and lower 95% CIs. C, SP-elicited behavior was challenged by intrathecal administration of either deltorphin II (circles) or UK + P (squares) or both (triangles) in D79N mice. The combination treatment (100:1 molar agonist ratio) failed to shift the UK + P dose–response curve in D79N animals, even though deltorphin II was otherwise effective at those doses. The abscissa values for the combined treatment dose–response curves represent the dose of UK 14,304 in the presence of an equal potency ratio of deltorphin II. D, Isobolographic analysis was applied to data from Figure 5C. The ED50 values for the drugs given alone were 51 nmol (95% CI = 22–118) for UK + P and 0.33 nmol (95% CI = 0.20–0.57) for deltorphin II. The ED50 for UK + P when co-administered with deltorphin II was 12 nmol (95% CI = 8.6–17). The 95% CI of the combined ED50 fell within the lower confidence 95% CIs of the theoretical additive line, indicating that the interaction between these two compounds in D79N mice was not significantly different from additive. This study has been repeated blind with similar results (data not shown).
To determine whether the μ-opioid receptor interacts with the α2aAR, we administered either DAMGO, a μ-opioid agonist, or UK + P or both, and constructed dose–response curves for inhibition of SP-elicited behavior in WT (Fig.6A) and D79N animals (Fig. 6C). Isobolographic analysis revealed that DAMGO and UK interacted synergistically in the WT animals (Fig.6B). Consistent with the results observed with deltorphin II, this synergy was absent in the D79N animals (Fig.6D). Taken together, these results suggest that α2 adrenergic agonist activation of the α2aAR is sufficient for synergy of UK 14,304 with either of the μ- or δ-opioid receptor subtypes to occur.

Co-administration of UK 14,304 (+ 5 pmol prazosin) and DAMGO is synergistic in WT but not in D79N mice. A, SP-elicited behavior was challenged by intrathecal administration of either DAMGO or UK 14,305 + 5 pmol prazosin (UK + P) or both in WT mice. UK + P (squares) and DAMGO (circles) inhibited the behavior in a dose-dependent manner. When both UK 14,304 and DAMGO were co-administered, a constant potency ratio (10:1 molar agonist ratio) was maintained. The combination treatment (triangles), expressed in terms of UK + P, was ∼10-fold more potent than either drug given alone.B, Isobolographic analysis was applied to the data fromA as described in Figure 5. They-intercept represents the ED50 (0.09 nmol; 95% CI = 0.07–0.11) for UK + P, and thex-intercept represents the ED50 (0.006 nmol; 95% CI = 0.004–0.01) for DAMGO when each is administered alone for inhibition of SP-elicited behavior in WT mice. When the two compounds were co-administered in WT animals, the ED50 for UK + P in the presence of DAMGO (0.004 nmol; CI = 0.003–0.005) fell well below the additive line, indicating a synergistic interaction. C, SP-elicited behavior was challenged by intrathecal administration of either DAMGO (circles) or UK + P (squares) or both (triangles) in D79N mice. The combination treatment (10,000:1 molar agonist ratio) failed to shift the UK + P dose–response curve in D79N. Abscissa values for the combined treatment dose–response curves represent the dose of UK 14,304 in the presence of an equal potency ratio of DAMGO.D, Isobolographic analysis was applied to data fromC. The ED50 values for the drugs given alone in D79N mice were 97 nmol (95% CI = 52–180) for UK + P and 0.008 nmol (95% CI = 0.004–0.015) for DAMGO. The ED50 for UK + P when co-administered with DAMGO was 70 nmol (95% CI = 40–123). The 95% CI of the combined ED50 crossed the theoretical additive line, indicating that the interaction between these two compounds in D79N mice is not synergistic.
The α2aAR modulates morphine-induced inhibition of SP-elicited behavior
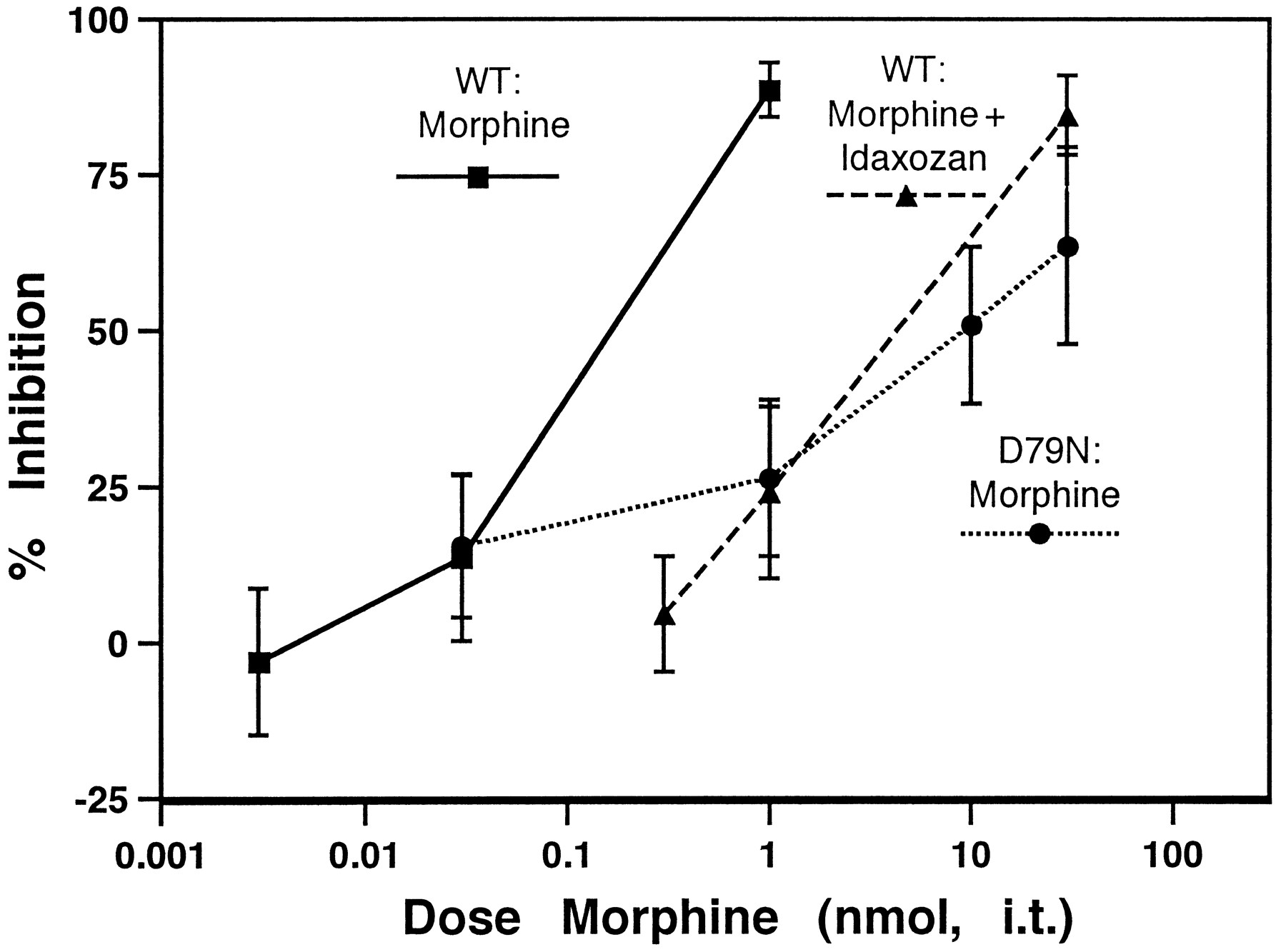
Endogenous norepinephrine (NE) released in the spinal cord from descending fibers contributes to the inhibitory effects of morphine, possibly through a synergistic interaction between opioid and adrenergic receptor systems; for example, the potency of morphine can be attenuated by spinal administration of adrenergic antagonists, presumably by blocking the action of endogenously released NE (Yaksh, 1979). We therefore hypothesized that the D79N mutation would result in a decrease in the potency of spinal morphine. To examine this hypothesis, we assessed the ability of morphine to inhibit SP-induced behavior in both WT and D79N animals. We observed a 75-fold increase in the morphine ED50 in the mutant animals as compared with WT (Fig. 7). To confirm that endogenous NE was contributing to inhibition by morphine, we co-administered morphine and the α2AR antagonist idaxozan in WT animals. The presence of idaxozan (0.1 nmol, i.t.) increased the ED50 of morphine in WT animals by 35-fold, confirming the involvement of the adrenergic system in morphine-induced inhibition in this assay. Co-administration of morphine and idaxozan in the D79N animals failed to further shift the morphine dose–response curve (data not shown). These results suggest that the α2aAR mediates the adrenergic component of morphine-induced inhibition in the SP assay.

Inhibition of SP-elicited behavior by morphine is reduced in D79N mice. Morphine potency (nmol, i.t.) is decreased in D79N animals (ED50 = 9.5 nmol; 95% CI = 1.4–62) as compared with WT animals (ED50 = 0.13 nmol; 95% CI = 0.05–0.30). This decrease in potency was mimicked by co-administration of the nonsubtype-selective α2AR antagonist idazoxan (ED50 = 4.2 nmol; 95% CI = 1.9–9.0). This result, together with those shown in Figures 5 and 6, suggests that a lack of synergy between descending noradrenergic and spinal opioid analgesia in D79N animals is mediating the decreased potency observed in the mutant mice. Supporting this conclusion, co-administration of idazoxan did not further alter the potency of morphine in D79N mice (ED50 = 0.83 nmol; 95% CI = 0.08–8.6). Error bars represent ±SEM for each dose point (n = 6–10 animals/dose).
DISCUSSION
Our results indicate that the analgesic effects of spinally administered α2AR agonists are mediated primarily by the α2aAR subtype as assessed using both the tail-flick and SP assays. In addition, the synergistic interaction observed in WT animals between the adrenergic agonist UK 14,304 and both μ- and δ-opioid agonists was abolished in the D79N mice, indicating that the α2aAR subtype is the primary mediator of adrenergic–opioid synergy. Furthermore, the presence of the α1, α2b, and α2c antagonist prazosin in these studies suggests that activation of the α2aAR is sufficient to mediate adrenergic–opioid synergy in WT animals. Interestingly, we found that the analgesic potency of morphine was decreased in the D79N animals in the SP assay. The ability of the nonsubtype-selective α2AR antagonist idaxozan to mimic the effect of the mutation on morphine potency in WT animals suggests that, at least in the SP assay, endogenously released noradrenaline from descending fibers likely modulates spinal morphine through an action at α2aARs.
In contrast to the lack of α2AR-mediated analgesia observed in thermal nociceptive tests in D79N mice, some agonist-induced effects on spinal analgesia were observed in the SP assay, albeit at supramaximal doses of agonists. This remaining activity may be explained in several ways. (1) At the supramaximal doses of agonist used in the D79N animals, the antagonist used to suppress possible antinociceptive actions of UK 14,304 on the α2bAR and α2cAR subtypes may not have been sufficient to antagonize agonist action at these receptors. (2) The α2aAR may retain some ability to activate residual signal transduction pathways independent of coupling to K+and Ca2+ channels. For example, α2ARs have been shown to couple to adenylyl cyclase in the spinal cord (Uhlen and Wikberg, 1988), and whether this inhibitory pathway remains intact in the mutant animals has not yet been established. The importance of residual coupling is unclear, however, because the inhibitory actions of spinal α2ARs at adenylyl cyclase may not be linked to their antinociceptive properties (Uhlen et al., 1990). (3) The residual effects of the α2AR agonists UK 14,304 and dexmedetomidine may be attributable to an action at another receptor population, such as imidazoline receptors, which exhibit a high affinity for many adrenergic ligands. A role, or lack thereof, for imidazoline receptors in antinociception has yet to be clearly determined (Codd et al., 1995). It is clear from our data, however, that the α2aAR is the primary mediator of spinal adrenergic analgesia in the mouse.
Receptor subtypes involved in synergy
Previous attempts to determine receptor subtypes necessary for adrenergic–opioid synergy have focused on opioid receptor subtypes. It has been shown, for example, that the δ-opioid receptor mediates this synergistic interaction in the mouse spinal cord, whereas co-administration of adrenergic and μ-opioid agonists results in an antagonistic or subadditive interaction (Roerig et al., 1992). Electrophysiological studies in the rat have concluded in one case that the δ-opioid receptor is required (Omote et al., 1991), whereas in another that the μ-opioid receptor is necessary (Sullivan et al., 1992). In this study, we have shown that both the μ-opioid agonist DAMGO and the selective δ-opioid agonist deltorphin II synergize with the α2 adrenergic agonist UK 14,304. Furthermore, this synergy is absent in the D79N mice, indicating that the α2aAR subtype is necessary for adrenergic–opioid synergy with either opioid receptor subtype. To confirm that the lack of synergy observed in D79N was not specific for the SP test, we co-administered ineffective doses of the adrenergic agonist clonidine with low doses of morphine in the tail-flick test. The presence of clonidine resulted in a significant increase in morphine potency in WT but not in D79N mice (data not shown). We are confident, therefore, that the lack of synergy observed in the SP test generalizes to other tests. Our observation that μ-opioid receptor activation results in a synergistic rather than an additive or antagonistic interaction with adrenergic agents can be explained in two ways. First, Roerig et al. (1992) used ICR mice and others used rat, whereas our study was performed an a B6,129 mixed genetic background. Thus, species or strain differences could explain the apparent differences in synergy with δ- versus μ-opioid receptors. Second, those studies that failed to show a role for the μ-opioid receptor used clonidine as their adrenergic agonist, which in many settings behaves as a partial agonist. In addition, clonidine is also a ligand at both α1ARs and imidazoline receptors, and these nonselective actions may account for the differences between reports.
Modulation of morphine antinociceptive action by spinal α2aARs
We observed that the potency of spinal morphine is significantly reduced in the D79N animals in the SP test, suggesting that activation of the α2aAR by endogenous NE contributes to spinal morphine potency in this assay. In support of this, we found that co-administration of idazoxan with morphine in WT animals also decreased morphine potency. Interestingly, we did not see a difference in the potency of spinal morphine in the tail-flick assay. Differences in the nature of the stimuli may lead to differential activation of descending NE pathways, such that endogenous NE plays a greater role in the SP test. If, for example, the tail-flick response at the temperature used in this study (52.5°C) evokes a largely spinal reflex, descending systems may not be sufficiently activated to contribute a measurable effect; however, the belief that the tail-flick response is a purely spinal reflex has been tempered in light of evidence linking brainstem activation to the onset of tail withdrawal (Heinricher et al., 1989). Exogenously applied SP may simply lead to a stronger activation of descending pathways than the thermal stimuli under the conditions used, leading to increased NE release and hence modulation of morphine effects in the spinal cord.
The response times measured in the two assays also may provide an explanation for the difference between the tail-flick and SP test results. Tail-flick withdrawals approximate a few seconds, whereas SP-elicited behavior encompasses a full 60 sec after intrathecal injection. If the activation of descending systems requires several seconds to evoke, then detection of the contributions of descending noradrenergic fibers may be difficult in the briefer tail-flick response. Alternatively, the tail-flick assay may lack the sensitivity necessary to detect the contribution of descending pathways. Although our data do not distinguish between these possibilities, they do indicate that endogenous NE acting at the α2aAR modulates spinal morphine action in the SP test. This observation strongly suggests that the α2aAR is the site of analgesic action of endogenous NE as well as of exogenous adrenergic agonists.
Implications for mechanisms of synergistic interactions
Synergistic interactions between classes of agonists have been reported frequently in the literature, yet the underlying mechanisms of such supra-additive interactions remain unknown. Although this study does not directly address the biochemical substrates necessary for synergy to occur, it does provide some preliminary insights into the issue. First, the observation that the D79N mutation leads to an uncoupling of the receptor to both K+ and Ca2+ channels (Lakhlani et al., 1996) suggests that ion channel activation may be necessary for synergistic interactions to occur. Second, it has been proposed that synergy can occur only when two receptor populations, acting through common signaling systems, are anatomically located at different locations in the pathway (Honore et al., 1996). If, for example, receptor pairs that couple similarly are co-expressed in single Xenopus oocytes, receptor co-activation yields an additive rather than supra-additive interaction (Birnbaum et al., 1995). Additive interactions have also been reported in locus ceruleus neurons that hyperpolarize in response to both opioid and adrenergic agonists (Andrade and Aghajanian, 1985). Data presented in this study, indicating that the α2aAR subtype is both necessary and sufficient for adrenergic–opioid synergy, open the door for investigations into the spatial relationships between synergistic receptor pairs for the first time.
Our results provide strong evidence that the α2aAR subtype is responsible for α2AR agonist-mediated analgesia in the mouse spinal cord. In addition, absence of synergy between α2aAR and both μ- and δ-opioid agonists in the D79N mice indicates that the α2aAR subtype is necessary for this interaction. Furthermore, α2aAR–opioid synergy may contribute to the potency of spinal morphine in situations in which descending noradrenergic pathways are activated. These synergistic interactions are important in clinical pain management. Low doses of combined adrenergic–opioid medications produce analgesic efficacy at minimal doses of the two analgesic agents, thus decreasing total drug requirements in patients. Our findings emphasize that agents capable of selective α2aAR activation should prove therapeutically useful when used alone or in combination with opioid analgesics in the treatment of pain.
Footnotes
This work was supported by National Institutes of Health (NIH) Grants R01-DA-01933, R01-DA-04274, and K02-DA-00145 to G.L.W., and NIH Grant R01-HL-43671 and a National Alliance for Research on Schizophrenia and Depression Established Investigator award to L.E.L. We thank Dr. Robert Elde for support and encouragement, Drs. S. Roerig and M. Ossipov for advice on the analysis of isobolograms, the Pfizer Drug company for the gift of UK 14,304, and Zeneca Pharmaceuticals for the dexmedetomidine.
Correspondence should be addressed to Dr. George L. Wilcox, Department of Pharmacology, University of Minnesota, 3-249 Millard Hall, 435 Delaware Street SE, Minneapolis, MN 55455.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}