

Abstract
α2-Adrenergic receptors (α2-ARs) mediate a number of physiological phenomena, including spinal analgesia. We have developed subtype-selective antisera against the C termini of the α2A-AR and α2C-AR to investigate the relative distribution and cellular source or sources of these receptor subtypes in the rat spinal cord. Immunoreactivity (IR) for both receptor subtypes was observed in the superficial layers of the dorsal horn of the spinal cord. Our results suggest that the primary localization of the α2A-AR in the rat spinal cord is on the terminals of capsaicin-sensitive, substance P (SP)-containing primary afferent fibers. In contrast, the majority of α2C-AR-IR was not of primary afferent origin, not strongly colocalized with SP-IR, and not sensitive to neonatal capsaicin treatment. Spinal α2C-AR-IR does not appear to colocalize with the neurokinin-1 receptor, nor is it localized on astrocytes, as evidenced by a lack of costaining with the glial marker GFAP. However, some colocalization was observed between α2C-AR-IR and enkephalin-IR, suggesting that the α2C-AR may be expressed by a subset of spinal interneurons. Interestingly, neither subtype was detected on descending noradrenergic terminals. These results indicate that the α2-AR subtypes investigated are likely expressed by different subpopulations of neurons and may therefore subserve different physiological functions in the spinal cord, with the α2A-AR being more likely to play a role in the modulation of nociceptive information.
- α2-adrenergic receptor
- spinal cord
- immunohistochemistry
- RG10
- RG20
- capsaicin
- dorsal rhizotomy
- confocal
- rat
- substance P
- noradrenaline
- analgesia
- α2A
- α2C
Agonists acting at α2-adrenergic receptors (α2-ARs) mediate a number of physiological and pathophysiological phenomena, including analgesia (Yaksh, 1985; Jänig, 1992; Perl, 1994;O’Halloran and Perl, 1997). α2-Adrenergic receptors belong to the superfamily of seven transmembrane-spanning, G-protein-coupled receptors and share common signal transduction pathways mediated through the pertussis toxin-sensitive inhibitory G-proteins Gi and Go (Crain et al., 1987; Hoehn et al., 1988). Activation of α2-ARs can decrease neuronal excitation by opening inwardly rectifying potassium channels (Surprenant and North, 1988; Shen et al., 1992), by decreasing presynaptic calcium influx (Ewald et al., 1989; Surprenant et al., 1990), and by inhibiting adenylyl cyclase (Andrade and Aghajanian, 1985; Uhlén and Wikberg, 1988; Uhlén and Wikberg, 1989). Three subtypes of α2-ARs have been cloned in human and rat, corresponding to the pharmacological subtypes α2A, α2B, and α2C, respectively (for review, see Bylund et al., 1994). Because of the lack of subtype-selective pharmacological agents, it is unclear which subtype or subtypes contribute to spinal adrenergic analgesia, although a major role has been suggested for the α2A-AR (Millan, 1992; Millan et al., 1994; Stone et al., 1997b).
In situ hybridization studies with probes directed against the α2A-AR have detected mRNA in a subset of dorsal root ganglion (DRG) and spinal cord neurons (Nicholas et al., 1993). In addition, α2A-AR mRNA and immunoreactivity have been detected in the cell bodies of neurons in almost all supraspinal noradrenergic nuclei (Nicholas et al., 1993; Rosin et al., 1993;Scheinin et al., 1994). The α2A-AR may therefore act as an autoreceptor on some noradrenergic terminals. In addition, α2A-AR immunoreactivity (α2A-AR-IR) has been reported in the superficial dorsal horn of the rat spinal cord (Rosin et al., 1993) and in DRG (Gold et al., 1997). Interestingly, although studies have detected α2C-AR mRNA in a large number of DRG neurons (Nicholas et al., 1993; Gold et al., 1997), spinal α2C-AR-IR has only been detected in cell bodies in the ventral horn (Rosin et al., 1996). Based on this information, spinal α2-ARs may originate from three possible sources. First, the receptors may be synthesized in the cell bodies of DRG neurons and trafficked centrally into the spinal cord, where they would serve a presynaptic function to modulate release of transmitter from these terminals. Second, these receptors may be synthesized by second-order spinal neurons in which they may modulate primary afferent activity either postsynaptically or presynaptically within circuits intrinsic to the spinal cord. Third, these receptors may be synthesized by supraspinal neurons and trafficked to axons and nerve terminals of descending noradrenergic fibers in which they would act as inhibitory autoreceptors controlling noradrenaline release in the spinal cord.
The goal of this study was to determine the source or sources of spinal α2A-AR-IR and α2C-AR-IR. We have thus generated anti-peptide antisera directed against the C termini of the predicted sequences of the α2A-AR and α2C-AR. Dorsal rhizotomy, neonatal capsaicin treatment, and double-labeling experiments were used to determine the origin and cytochemical profile of α2-AR immunoreactive fibers and terminals observed in the dorsal horn.
Part of these results have been presented in abstract form (Stone et al., 1996, 1997a,c).
MATERIALS AND METHODS
Generation of antisera. Anti-peptide antisera were generated against the C-terminal portion of the α2A- and α2C-adrenergic receptors in a manner similar to that previously described (Arvidsson et al., 1995a). Briefly, peptides corresponding to the last 15 residues of the rat RG20 (α2A-AR) clone and the last 13 residues of the rat RG10 clone (α2C-AR) (Lanier et al., 1991) were synthesized using an Applied Biosystems (Foster City, CA) 432A solid-phase peptide synthesizer. The peptides (10 mg/ml) were then conjugated to bovine thyroglobulin (Sigma, St. Louis, MO; 40 mg/ml) using 7% glutaraldehyde (Sigma; 30 μl/ml). For initial immunizations, 1 mg of the peptide–thyroglobulin conjugate was emulsified with an equal volume of Freund’s complete adjuvant (Difco, Detroit, MI) and injected into rabbits. Subsequent immunizations consisted of 0.5 mg of the conjugate emulsified with an equal volume of Freund’s incomplete adjuvant administered at 2 week intervals. Serum was obtained 1 week after immunization, beginning after 5 weeks. When guinea pigs were used instead of rabbits, the procedure was identical, with the exception that all immunization volumes were half those used for rabbits. The staining patterns of antisera directed against the α2C-AR derived from rabbits and guinea pigs were indistinguishable, and the antisera were, therefore, used interchangeably throughout the study.
Immunohistochemistry. Madin–Darby canine kidney (MDCK) cells were stably transfected with either the α2A-AR, α2B-AR, or α2C-AR (gift of Dr. Lee Limbird, Vanderbilt University, Nashville, TN) (Wozniak et al., 1997). Cells were grown on coverslips and fixed with 4% paraformaldehyde and 0.2% picric acid in 0.1 m PBS, pH 6.9, for 1 hr. Cells were washed several times in PBS and then were preincubated for 1 hr at room temperature in diluent containing 1% normal donkey serum, 0.3% Triton X-100, and 1% bovine serum albumin. Cells were then incubated overnight at 4°C in a humid chamber with either α2A-AR antisera or α2C-AR antisera (1:1000) in the same diluent. In some cases, the antisera were mixed with unconjugated peptide (10 μg/ml) before application. The cells were then rinsed several times with PBS, incubated for 1 hr at room temperature with cyanine 3.18-labeled donkey anti-rabbit IgG (Jackson ImmunoResearch, West Grove, PA; 1:100), rinsed, and coverslipped with glycerol and PBS containing 0.1% p-phenylenediamine.
Male rats (Harlan Sprague Dawley, Indianapolis, IN; 150–200 gm) were anesthetized with an intramuscular injection of a mixture of 75 mg/kg ketamine, 5 mg/kg xylazine, and 1 mg/kg acepromazine and were fixed with 4% paraformaldehyde and 0.2% picric acid in 0.1 mPBS, pH 6.9, by vascular perfusion as previously described (Wessendorf and Elde, 1985). Spinal cords were removed, rinsed overnight with 10% sucrose in PBS, and prepared for cryostat sectioning. Thaw-mounted cryostat sections (14 μm) were prepared for indirect immunofluorescence histochemistry. Cryostat sections were incubated with primary antisera and processed in a manner similar to that described above. In some instances, fluorescence double-labeling was used as previously described (Wessendorf and Elde, 1985).
Primary antisera were used at the following dilutions: rabbit anti-α2A-AR, 1:1000; rabbit anti-α2C-AR, 1:1000; guinea pig anti-α2C-AR, 1:500; mouse anti-calcitonin gene-related peptide (CGRP), 1:2000 (Research Biochemicals, Natick, MA); mouse anti-dopamine-β-hydroxylase, 1:500 (Chemicon, Temecula, CA); rat anti-substance P (SP), 1:1000 (Research Biochemicals); rabbit anti-neurokinin-1 receptor (NK-1R) 1:500 (Vigna et al., 1994; a gift from Dr. P. Mantyh, University of Minnesota, Minneapolis, MN); rabbit anti-somatostatin, 1:1000 (Seybold and Elde, 1980); mouse anti-leuenkephalin, 1:500 (Sera-Lab, Sussex, UK); mouse anti-tyrosine hydroxylase, 1:500 (IncStar, Stillwater, MN); guinea pig anti-preprodynorphin, 1:500 (Arvidsson et al., 1995b); and mouse anti-GFAP, 1:500 (Sigma). Single-labeled preparations were visualized with cyanine 3.18-conjugated secondary antisera 1:200 (Jackson ImmunoResearch). Double-labeled preparations were visualized with a mixture of fluorescein isothiocyanate and lissamine–rhodamine-conjugated secondary antisera 1:200 (Jackson ImmunoResearch). Sections were examined with a Bio-Rad MRC-1000 Confocal Imaging System (Bio-Rad Microscience Division, Cambridge, MA). Digital images were adjusted for brightness and contrast using Photoshop 4.0 (Adobe Systems, San Jose, CA) in a manner similar to that used for traditional darkroom printing of film-based images and were often digitally merged. The examples of colocalization obtained in the higher-magnification images most probably represent colocalization, and not superposition, because the estimated optical section thickness of a confocal image at the higher magnifications used is <1.0 μm, a section size thin enough to minimize the possibility of superimposition of terminals that have dimensions on the order of 1–2 μm. Plates of images were assembled using Confocal Assistant (Bio-Rad) and printed on a Fuji Pictrography 3000 color printer. A minimum of three animals were examined with each antiserum or antisera combination used. Although we confirmed that our observations were consistent at all levels of the spinal cord, representative samples of lumbar cord are used in the figures.
Dorsal rhizotomy. Unilateral dorsal rhizotomies were performed at spinal segments L3 and S1 in sodium pentobarbital-anesthetized animals. Sham operations were performed by exposing the spinal cord without severing the dorsal roots. Five days after surgery, the animals were fixed by vascular perfusion, spinal cords were removed, and the lumbar–sacral segments were processed for immunohistochemistry as described above. The resultant sections were then sampled and stained for SP. The region of maximal effect was determined visually by assessing the decrease in SP immunoreactivity ipsilateral to the lesion. Once this region was identified, serial sections were double-stained with SP and either α2A-AR or α2C-AR antisera. Changes in labeling density were then determined visually. The SP staining was used to indicate that the effect of the rhizotomy (as determined by the decrease in SP-IR) was similar in both the α2A-AR- and α2C-AR-stained sections. A minimum of three animals were examined from each treatment group.
Neonatal capsaicin treatment. Rat pups were injected subcutaneously with 50 mg/kg capsaicin within 48 hr of birth and allowed to survive to 5 weeks of age. Control animals were injected with vehicle only (10% ethanol and 10% Tween 20 in 0.9% saline). Animals were then fixed by vascular perfusion, and spinal cords were removed and treated as above. A minimum of three animals were examined from each treatment group. All experimental procedures were approved by the Institutional Animal Care and Use Committee of the University of Minnesota.
RESULTS
Characterization of antisera
MDCK cells, transfected with the α2A-AR, α2B-AR, or α2C-AR or untransfected, were stained with rabbit-derived antisera generated against the α2A-AR or α2C-AR. α2A-AR-IR was observed only on cells transfected with the α2A-AR (Fig.1A–D), whereas α2C-AR-IR was observed only on cells transfected with the α2C-AR (Fig.1E–H). The α2A-AR-IR was blocked by preabsorption of the antiserum with the peptide corresponding to α2A-AR but not by that corresponding to the α2C-AR (Fig.1I,J). Similarly, α2C-AR-IR was blocked by preabsorption of the antiserum with the peptide corresponding to the α2C-AR but not the α2A-AR (Fig. 1K,L). Guinea pig-derived anti-α2C-AR antisera generated similar results (data not shown). These results demonstrate that both the α2A-AR and α2C-AR antisera recognize the receptors against which they were generated and do not cross-react with other known α2-AR subtypes.

Characterization of α2A-AR-IR and α2C-AR-IR in transfected MDCK cells. MDCK cells stably transfected with either the α2A-AR, α2B-AR, or α2C-AR or untransfected, wild-type (WT) cells were stained with antisera generated against the C-terminal portions of the α2A-AR and α2C-AR. The α2A-AR antisera stained the membranes of MDCK cells transfected with the α2A-AR (A) but not untransfected cells (D) or cells transfected with the α2B-AR (B) or the α2C-AR (C). Similarly, α2C-AR-IR was observed on the membranes of MDCK cells transfected with the α2C-AR (G) but not untransfected cells (H) or cells transfected with the α2A-AR (E) or the α2B-AR (F). The α2A-AR-IR observed on the membranes of α2A-AR-transfected cells was blocked by preabsorption of the antiserum with the peptide corresponding to the α2A-AR (I) but not the α2C-AR (J). The α2C-AR-IR observed on the membranes of α2C-AR-transfected cells was blocked by preabsorption of the antiserum with the peptide corresponding to the α2C-AR (L) but not that of the α2A-AR (K). These results demonstrate that both the α2A-AR and α2C-AR recognize their respective receptors and do not cross-react with the other α2-AR subtypes.
Distribution of α2A-AR and α2C-AR in the spinal cord
In rat spinal cord, both α2A-AR-IR and α2C-AR-IR were observed in nerve terminals and varicosities. No definitive evidence for cell body labeling was obtained. These immunoreactive terminals were most highly concentrated in laminae I and II of the dorsal horn at all levels of the spinal cord (Fig.2A,C). In addition, α2A-AR-IR was observed in the area surrounding the central canal and in the intermediolateral cell column of the thoracic cord (data not shown). Strong α2C-AR-IR was observed in the lateral spinal nucleus and was more prevalent in deeper layers of the dorsal horn than α2A-AR-IR. Staining was blocked in both cases by preincubation of each antisera with its cognate peptide (Fig.2B,D).

Characterization of α2A-AR-IR and α2C-AR-IR in rat spinal cord. Immunoreactivity for α2A-AR (A) and α2C-AR (C) were observed in the superficial dorsal horn. In addition, α2A-AR-IR was observed in the area surrounding the central canal and in the intermediolateral cell column of the thoracic cord (data not shown). Strong α2C-AR-IR was observed in the lateral spinal nucleus and was more prevalent in deeper layers of the dorsal horn than α2A-AR-IR. Staining was blocked in both cases by preincubation of antiserum with the cognate peptide (B, D).
To determine whether the spinal α2A-AR-IR and α2C-AR-IR were of primary afferent origin, dorsal rhizotomies were performed to deplete the spinal cord of primary afferent input. We observed a dramatic reduction in α2A-AR-IR ipsilateral to dorsal rhizotomy (Fig.3A). In contrast, dorsal rhizotomy yielded only a slight reduction in α2C-AR-IR (Fig. 3C). Double-labeling each section with antiserum directed against SP confirmed that SP-IR was also reduced by dorsal rhizotomy to approximately the same extent (Fig.3B,D) in both sections, indicating that the efficacy of the treatment was similar in each case.

Reduction in α2A-AR-IR and α2C-AR-IR after dorsal rhizotomy. Dorsal rhizotomies were performed to determine whether the α2-ARs were synthesized in dorsal root ganglia neurons and trafficked centrally into the spinal cord. After rhizotomy, a dramatic reduction in α2A-AR-IR was observed ipsilateral to the lesion (A). A reduction in α2C-AR-IR was also observed (C), although to a lesser extent than for the α2A-AR. SP-IR, however, was reduced to a similar extent in both tissue sections, indicating that the efficacy of the treatment was similar in each case (B,D).
Neonatal capsaicin treatment reduces α2AAR-IR but not α2CAR-IR
To determine whether one or both subtypes is expressed in capsaicin-sensitive neurons, we treated neonatal rats with capsaicin and examined their spinal cords at 5 weeks of age. Capsaicin, the active ingredient in hot peppers, acts as a neurotoxin on small diameter primary afferents, many of which are SP-containing (Jancso et al., 1977; Nagy et al., 1981). Sensitivity of sensory neurons to capsaicin has been associated with a role in nociception (Nagy et al., 1980; Jancso et al., 1987; Hammond and Ruda, 1991). We observed a dramatic reduction in α2A-AR-IR but not α2C-AR-IR (Fig. 4), indicating that the subset of DRG cells that express the α2A-AR is likely to consist of small diameter, capsaicin-sensitive nociceptors. In addition, these results suggest that the two subtypes may exist on different populations of neurons.

Reduction in α2A-AR-IR but not α2C-AR-IR after neonatal capsaicin treatment. Animals were treated as neonates with capsaicin to determine whether the α2-ARs were expressed on the terminals of capsaicin-sensitive neurons. α2A-AR-IR was reduced in animals treated with neonatal capsaicin (A) compared with controls (B). In contrast, the α2C-AR-IR was not noticeably reduced by this treatment (C) compared with controls (D), suggesting that the two receptor subtypes are expressed by independent subpopulations of neurons and that the α2A-ARs in particular are expressed on the subset of primary afferents that are capsaicin-sensitive.
Differential distribution of α2A-AR and α2C-AR in the rat spinal cord
Double-labeling experiments were conducted to determine the relationship between α2A-AR-IR and α2C-AR-IR in the superficial dorsal horn. Sections of rat spinal cord double-labeled with anti-α2A-AR and anti-α2C-AR antisera produced robust immunostaining for each receptor in the superficial dorsal horn (Fig.5A,B). The resultant digital images were then digitally merged (Fig.5C). In merged images, the appearance of yellow may indicate colocalization, although higher resolution is required to distinguish colocalization from close apposition or superimposition. When examined at high resolution, instances of receptor colocalization were rare (Fig. 5D). This observation further suggests that the two subtypes are located primarily on different neuronal populations.

Characterization of α2A-AR-IR fibers in rat spinal cord. Single sections of lumbar spinal cord were double-labeled with α2A-AR antisera in combination with various other antisera. The first column(A, E, I,M, Q, U) depicts α2A-AR-IR (red). The second column (B, F, J,N, R, V) depicts immunoreactivity obtained with other antisera used in double-labeling (green). The third column(C, G, K,O, S, W) depicts digitally merged images of the first two columns. In merged images, the appearance of yellow may indicate colocalization, although higher magnification is required to distinguish colocalization from close apposition. The fourth column(D, H, L,P, T, X) depicts merged images obtained at higher magnification. Row 1(A–D) demonstrates that α2A-AR -IR and α2C-AR-IR are rarely colocalized. Row 2(E–H) demonstrates that α2A-AR-IR and SP-IR are highly colocalized. Row 3 (I–L) demonstrates that α2A-AR-IR and CGRP-IR fibers are highly, but not exclusively, colocalized. Row 4(M–P) demonstrates that α2A-AR-IR and ENK-IR are rarely colocalized. Row 5 (Q–T) demonstrates that α2A-AR-IR and DβH-IR are closely apposed, but not colocalized. Similarly, row 6(U–X) demonstrates that instances of colocalization between α2A-AR-IR and TH-IR could not be detected. SP, Substance P; CGRP, calcitonin gene-related peptide; ENK, leuenkephalin; DβH, dopamine β-hydroxylase; TH, tyrosine hydroxylase.
Characterization of α2A-AR-IR fibers in the rat spinal cord
Double-labeling with anti-α2A-AR and anti-SP antisera revealed extensive colocalization in the spinal cord (Fig.5E–H). Thus, α2A-ARs appear to be present on the spinal terminals of SP-containing primary afferent fibers. Examination of sections double-labeled for α2A-AR-IR and CGRP-IR also revealed a significant degree of colocalization (Fig. 5I–L). In contrast, staining for the neuropeptide leuenkephalin failed to colocalize with α2A-AR-IR (Fig. 5M–P).
To determine whether α2A-ARs exist as autoreceptors on descending noradrenergic terminals, we used antibodies directed against dopamine-β-hydroxylase (DβH) and tyrosine hydroxylase (TH), enzymes involved in the biosynthesis of noradrenaline. Spinal cord sections double-labeled for either α2A-AR-IR and DβH-IR or α2A-AR-IR and TH-IR showed no instances of colocalization (Fig. 5Q–X). This trend was observed throughout the spinal cord. However, examples of close apposition between DβH-IR or TH-IR and α2A-AR-IR fibers and terminals were occasionally observed. Whether such close appositions represent axoaxonic synaptic relationships remains to be established.
Characterization of α2C-AR-IR fibers in the rat spinal cord
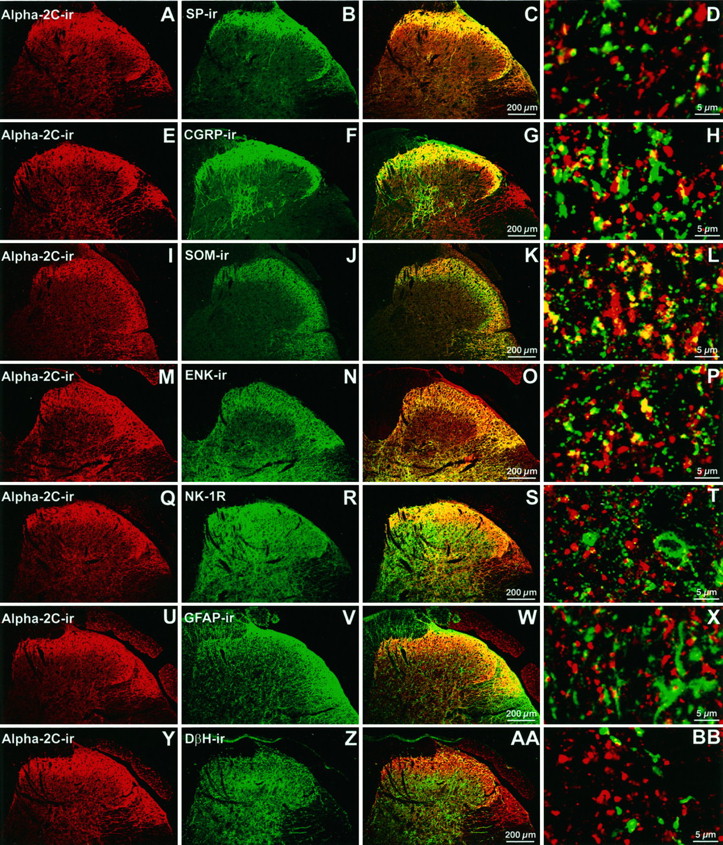
Double-labeling experiments indicated that α2C-AR-IR and SP-IR are rarely colocalized in the spinal cord (Fig.6A–D). However, α2C-AR-IR was present on some CGRP-IR (Fig.6E–H) and somatostatin-IR (Fig.6I–L) fibers. Interestingly, some overlap was detected between enkephalin-IR and α2C-AR-IR (Fig.6M–P). Because enkephalin is not believed to be present in primary afferent fibers (Hokfelt et al., 1977;Johansson et al., 1978; Seybold and Elde, 1980), this result is consistent with a spinal source of α2C-AR. No colocalization was observed between α2C-AR-IR and the endogenous opioid peptide preprodynorphin-IR (data not shown).

Characterization of α2C-AR-IR fibers in rat spinal cord. Single sections of lumbar spinal cord were double-labeled with α2C-AR antisera in combination with various other antisera. The first column(A, E, I,M, Q, U,Y) depicts α2C-AR-IR (red). The second column(B, F, J,N, R, V, Z) depicts immunoreactivity obtained with other antisera used in double-labeling (green). The third column (C, G, K,O, S, W,AA) depicts digitally merged images of the first two columns. In merged images, the appearance of yellow may indicate colocalization, although higher magnification is required to distinguish colocalization from close apposition. The fourth column (D, H, L,P, T, X,BB) depicts merged images obtained at higher magnification. Row 1(A–D) demonstrates that α2C-AR-IR and SP-IR are rarely colocalized. Row 2 (E–H) demonstrates that α2C-AR-IR and CGRP-IR fibers are moderately colocalized.Row 3 (I–L) demonstrates that α2C-AR-IR and somatostatin-IR fibers are moderately colocalized. Row 4 (M–P) demonstrates that α2C-AR-IR and ENK-IR are moderately colocalized. Row 5(Q–T) demonstrates that α2C-AR-IR and NK-1R are not colocalized. Row 6 (U–X) demonstrates that α2C-AR-IR and GFAP-IR are very rarely colocalized.Row 7 (Y–BB) demonstrates that α2C-AR-IR and DβH-IR are not colocalized. A similar lack of colocalization was observed between α2C-AR-IR and TH-IR (data not shown). SP, Substance P; CGRP, calcitonin gene-related peptide; SOM, somatostatin; ENK, leuenkephalin; NK-1R, NK-1 receptor; GFAP, glial fibrillary acidic protein; DβH, dopamine β-hydroxylase.
In an effort to further characterize the phenotype of spinal cells that express the α2C-AR, double-labeling was performed with antisera directed against the NK-1 receptor (Fig.6Q–T). We observed that the α2C-AR is not detected on either NK-1-expressing neurons or astrocytes, indicated by the rare colocalization of the glial marker GFAP with α2C-AR-IR (Fig.6U–X). α2C-AR-IR failed to colocalize with either DβH-IR (Fig. 6Y–BB) or TH-IR (data not shown), suggesting that neither the α2A-AR nor the α2C-AR is likely to be present on the terminals of descending supraspinal noradrenergic neurons.
DISCUSSION
We have developed subtype-specific anti-peptide antisera in rabbits and guinea pigs directed against the C-terminal portions of the α2A-AR and α2C-AR. The resultant antisera recognized their corresponding receptors and did not cross-react with other known α2-AR subtypes. In addition, labeling was blocked by preabsorption with the cognate peptide in both transfected cells and tissue sections. Immunoreactivity for both α2A-AR-IR and α2C-AR-IR was observed predominantly in fibers in the superficial layers of the dorsal horn of the spinal cord.
Both dorsal rhizotomy and neonatal capsaicin treatment resulted in a dramatic decrease in α2A-AR-IR. The present results suggest that the majority of spinal α2A-ARs are synthesized by DRG neurons and trafficked centrally. These observations are consistent with previous work showing a significant decrease in α2-AR agonist-binding in cat spinal cord ipsilateral to dorsal root ganglionectomies (Howe et al., 1987b). In contrast, the decrease in α2C-AR-IR ipsilateral to dorsal rhizotomy was small, and no change was detected in α2C-AR-IR after neonatal capsaicin treatment, suggesting that the majority of spinal α2C-AR is not of primary afferent origin. These results are surprising in light of evidence from in situhybridization studies demonstrating α2C-AR mRNA in numerous DRG cell bodies (Nicholas et al., 1993; Gold et al., 1997). It is possible that many DRG neurons express and transport α2C-ARs centrally at undetectable levels or in a form that is not recognized by our antisera. Alternatively, many α2C-AR-expressing DRG neurons may not transport the receptor to central terminals. Thus, the α2C-AR may be processed in a manner similar to the NPY-Y1 receptor, which has been shown to be present in a functional form on DRG cell bodies yet does not appear to be trafficked into the spinal cord (Zhang et al., 1994).Zhang et al. (1994) have proposed that the natural ligand for the NPY-Y1 receptor on DRG neurons may be blood-borne NPY released from sympathetic nerves or the adrenal medulla. A similar situation might occur for one or more α2-AR subtypes that may respond to circulating catecholamines, as was recently suggested to be the case in aberrant pain syndromes such as causalgia (O’Halloran and Perl, 1997).
The decrease in α2A-AR-IR after neonatal treatment of rats with capsaicin suggests that the α2A-AR may be primarily expressed by nociceptive neurons, because capsaicin-sensitive fibers have been associated with nociception (Nagy et al., 1980; Jancso et al., 1987; Hammond and Ruda, 1991). In addition, the fact that α2A-AR-IR but not α2C-AR-IR was affected by this treatment indicates that the two subtypes may not be present in the same neurons. This is supported by our observation in double-labeling studies that overlap between the two subtypes is rare. Interestingly, Wikberg and Hajós (1987) have reported that the number of [3H]clonidine-binding sites in the spinal cord was not significantly altered by neonatal capsaicin treatment. Their failure to detect a decrease in binding may be related to their use of a ligand that is not subtype-specific, leading to a “masking” of the decrease in the α2A-AR by the lack of effect on other clonidine-binding sites. Thus, a decrease in only one of two or more possible binding sites may not have been detectable under the experimental conditions used in their study.
In an effort to further identify the cellular sources of α2ARs in the superficial dorsal horn, we used a number of cytochemical markers. The α2A-AR was found to be highly colocalized with SP-IR fibers in the superficial dorsal horn, suggesting that α2A-ARs are synthesized and trafficked into the spinal cord almost exclusively by SP-containing neurons, some of which are likely to be nociceptors (for review, see Levine et al., 1993). A fair degree of colocalization was also observed with the neuropeptide CGRP. This result further supports our assertion that the majority of spinal α2A-ARs are of primary afferent origin, because CGRP-IR in the dorsal horn is believed to have exclusively primary afferent origins (Tuchscherer and Seybold, 1989). Consistent with this hypothesis are our observations that α2AAR-IR did not coexist with either leuenkephalin-IR or preprodynorphin-IR (data not shown), nor did it appear to be expressed at detectable levels by astrocytes, as evidenced by the lack of colocalization with the glial marker GFAP (data not shown).
In contrast to the α2A-AR, the α2C-AR did not extensively colocalize with either SP or CGRP. We did observe, however, a moderate extent of overlap between α2C-AR-IR and somatostatin-IR. Because the SP-IR and somatostatin-IR populations of DRG neurons are reported to be largely exclusive (Hökfelt et al., 1976; Tuchscherer and Seybold, 1985), this result suggests that the α2C-AR may be present in higher proportion on the spinal terminals of the latter. However, somatostatin in the rat dorsal horn is mainly of spinal origin (Nagy et al., 1981). Thus, the α2C-AR may be expressed by a population of somatostatin-IR positive cells intrinsic to the spinal cord.
When we investigated the relationship between α2C-AR-IR and that of the opioid peptides leuenkephalin and preprodynorphin, we observed some degree of colocalization between α2C-AR-IR and leuenkephalin-IR but not between α2C-AR-IR and preprodynorphin-IR. Although the physiological relevance of α2C-AR expression on leuenkephalin-containing neurons is not clear, this observation suggests that α2C-ARs are in a position to modulate enkephalinergic but not dynorphinergic neurons in the spinal cord. The α2C-AR does not appear to be expressed on neurons positively labeled for the NK-1 receptor nor by astrocytes.
Interestingly, immunoreactivity for both DβH and TH enzymes involved in the biosynthesis of noradrenaline did not colocalize with immunoreactivity for either of the α2-ARs investigated in this study. This result is puzzling in light of the evidence for strong expression of α2A-ARs in most supraspinal noradrenergic nuclei (Nicholas et al., 1993; Rosin et al., 1993; Scheinin et al., 1994). It has therefore been assumed that the α2A-ARs serve as presynaptic autoreceptors on these terminals. In contrast to this assumption, however, is the observation by Howe et al. (1987a) that the removal of descending noradrenergic inputs results in a decrease in total noradrenaline content in the spinal cord with no corresponding decrement in the density of α2-AR-binding sites. In addition, at least one study has shown that spinal administration of α2-AR agonists does not lead to agonist-induced inhibition of noradrenaline release, suggesting a lack of inhibitory adrenergic autoreceptors in the spinal cord (Klimscha et al., 1997).
Although it is possible that α2-ARs expressed by supraspinal noradrenergic cell groups are simply not trafficked to the spinal terminals of these neurons, it is also possible that they are present at levels below the detection limits of our antisera or that they exist in a conformation that precludes recognition. Furthermore,Nicholas et al. (1993) observed small α2A-AR mRNA-positive clusters in lamina II and III, yet we have not detected any α2A-AR-IR-positive cell bodies in these regions. Therefore, we cannot discount the possibility that other sources of α2-ARs may exist in addition to those that we have described in this study.
Evidence from functional studies supports many of our observations. It has been shown that α2-AR agonists can inhibit peptide release from spinal cord preparations (Kuraishi et al., 1985, Ono et al., 1991), suggesting an action on primary afferent fibers. Because α2A-AR-IR is observed on the terminals of many peptide-containing primary afferents, the α2A-AR may be mediating such phenomena. Adrenergic agonists have also been shown to directly hyperpolarize neurons in the dorsal horn, suggesting a postsynaptic action (Fleetwood-Walker et al., 1985; Surprenant and North, 1988). We report here that the majority of spinal α2C-AR-IR appears to be expressed by local spinal neurons; hence activation of spinal α2C-ARs may lead to the adrenergic agonist-induced hyperpolarizations observed in the spinal cord.
In summary, our results suggest that the major localization of the α2A-AR in the rodent spinal cord is on the terminals of capsaicin-sensitive, SP-containing primary afferent fibers. The importance of these primary afferents to nociception is firmly established. Therefore, it is likely that the analgesic action of spinal adrenergic agonists may be attributable, in part, to presynaptic inhibition of the release of excitatory transmitters such as glutamate and SP by α2A-ARs after peripheral noxious stimuli. In contrast, the majority of spinal α2C-ARs in the dorsal horn may be produced by neurons intrinsic to the spinal cord or on the terminals of non-noradrenergic brainstem neurons that project to the spinal cord. The differential expression of these subtypes suggests that they subserve different physiological roles in the spinal cord. It may therefore be possible to design subtype-selective pharmacological agents capable of targeting these functions independently for the purpose of modulating spinal nociceptive transmission.
Footnotes
This work was supported by National Institutes of Health Grants R01DA01933 to G.L.W. and DA06299 to R.E., Swedish Medical Research Council Grants 04X-2887 and ECBMH4-CT95-0172 to T.H. and DK43879 to Lee E. Limbird, Vanderbilt University. We thank J. Wang and G. Kalyuzhnaya for technical support, Dr. Lee Limbird for the gift of the transfected cells, and Shannon Wright for thoughtful reading of this manuscript.
Correspondence should be addressed to Dr. R. Elde, University of Minnesota, Department of Cell Biology and Neuroanatomy, 4-135 Jackson Hall, 321 Church Street SE, Minneapolis, MN 55455.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}