

Abstract
Spinal circuits form building blocks for movement construction. In the frog, such building blocks have been described as isometric force fields. Microstimulation studies showed that individual force fields can be combined by vector summation. Summation and scaling of a few force-field types can, in theory, produce a large range of dynamic force-field structures associated with limb behaviors. We tested for the first time whether force-field summation underlies the construction of real limb behavior in the frog. We examined the organization of correction responses that circumvent path obstacles during hindlimb wiping trajectories. Correction responses were triggered on-line during wiping by cutaneous feedback signaling obstacle collision. The correction response activated a force field that summed with an ongoing sequence of force fields activated during wiping. Both impact force and time of impact within the wiping motor pattern scaled the evoked correction response amplitude. However, the duration of the correction response was constant and similar to the duration of other muscles activated in different phases of wiping. Thus, our results confirm that both force-field summation and scaling occur during real limb behavior, that force fields represent fixed-timing motor elements, and that these motor elements are combined in chains and in combination contingent on the interaction of feedback and central motor programs.
The spinal cord of lower vertebrates and mammals may be organized into movement control modules (Grillner and Wallen, 1985; Loeb, 1985; Mortin and Stein, 1989; Rossignol, 1996;Kiehn et al., 1997; Smith and Stein, 1997). Such modules might control specific force patterns in the limb (Bizzi et al., 1991; Giszter et al., 1993; Nichols, 1994). How spinal modules are recruited, combined, and interact in the construction of limb behaviors has implications for motor learning, neural repair and rehabilitation, and the design of prosthetics and robotics. In this paper we present the first direct evidence that force-field primitives, spinal control modules identified in the frog, are combined to produce limb behaviors and their on-line adjustments.
Spinal microstimulation provided the initial evidence that the frog spinal cord may be organized into modules that produce force-field primitives (Bizzi et al., 1991; Giszter et al., 1993; Mussa-Ivaldi et al., 1994). Force fields were constructed by measuring isometric forces at the ankle with the limb held in a range of positions and with the same stimulus applied at each position. Primitives were defined as force fields that exhibited invariant force directions and magnitude balances over time. Only a few force-field types were found for any individual frog, force-field types were similar among frogs, and each force field typically converged to a specific workspace location (Giszter et al., 1993). The effect of coactivating force fields by dual microstimulation could be described as the linear sum of the individually activated force fields (Mussa-Ivaldi et al., 1994). Theoretical studies showed that summation and scaling of a few force-field types could be used to generate a large range of force-field structures and might underlie movement synthesis (Mussa-Ivaldi, 1992, 1997; Mussa-Ivaldi and Giszter, 1992).
Descending and segmental systems might recruit, scale, and sum force-field primitives in a manner similar to microstimulation (Bizzi et al., 1991; Bizzi et al., 1995; d'Avella and Bizzi, 1998). Indeed, force fields produced during certain reflex behaviors in the frog were similar to microstimulation-generated fields (Giszter et al., 1993). However, recent data have indicated that reflex behaviors and phases of behaviors may not correspond in a one-to-one fashion with a muscle synergy or force-field primitive, but instead correspond to combinations of such elements (Kargo et al., 1998; Tresch et al., 1999). Tresch et al. (1999) showed that different withdrawal responses could be described as constructed from different combinations of a basic set of muscle synergies. These synergies were similar to the muscle synergies evoked by spinal microstimulation (Tresch, 1997). How these data, which were measured at a single limb position, relate to the description of limb behaviors as a combination of force-field primitives was not tested directly.
We therefore examined in this paper how force-field mechanisms might be used to generate and control limb behavior. Specifically, we examined how the targeted wiping reflex of the spinal frog is organized and how on-line trajectory corrections are achieved. We found that force-field primitives were recruited in chains and in combination to produce wiping trajectories and their on-line adjustments. Furthermore, each primitive exhibited a common activation waveform of constant duration. Taken together, our data suggest that constant-duration force-field primitives form building blocks for the construction of certain limb behaviors.
MATERIALS AND METHODS
Surgery
Detailed surgical procedures have been outlined in previously published material (Giszter et al., 1993; Kargo and Giszter, 2000). All procedures were approved and in accordance with Institutional Animal Care and Use Committee recommendations. Briefly, bullfrogs received a spinal transection at high cervical levels below the base of the medulla, thereby isolating the spinal cord. The tectum was heat-cauterized. Ten muscles in the wiping limb were implanted with paired intramuscular electrodes for differential recording of muscle electrical activity. Muscles that were implanted included monoarticular hip flexors [iliopsoas (IL) and gluteus (GL)], biarticular hip/knee flexors [biceps (BI) and sartorius (SA)], monoarticular hip extensor [adductor magnus (AD)], biarticular hip flexor/knee extensors [rectus anterior (RA) and vastus externus (VE)], and biarticular hip extensors/knee flexors [semitendinosus (ST), rectus internus (RI), and semimembranosus (SM)]. All skin incisions were sealed with wound clips and cyanoacrylate tissue adhesive (Vetbond, 3m). Frogs were permitted 12–24 hr recovery after the surgery and stored in a damp container in the refrigerator at ∼10°C.
Experimental protocol and design
Fourteen frogs were used in these experiments. We tested the hindlimb-to-hindimb form of wiping. In this form, the wiping limb and target limb both move such that the stimulus and the ankle of the wiping limb (i.e., the wiping tool in this form) meet near the midline of the frog, just caudal to the cloaca [for detailed kinematic descriptions, see Giszter et al. (1989) and Kargo and Giszter (2000)]. In both free limb and isometric trials, the wiping reflex was initiated by a 500–600 msec train of 2 msec, 4–8 V biphasic pulses delivered at 33 Hz. The stimulus was applied via bipolar leads attached to the dorsolateral surface of the foot of the target hindlimb. The stimulus placement, and the parameters used, reliably elicited the hindlimb-to-hindimb form of wiping [for detailed kinematic descriptions, see Giszter et al. (1989) and Kargo and Giszter (2000)]. In this form of wiping, and in our hands, the wiping limb initiates movement 200–300 msec after onset of the stimulus and contacts the stimulus (on the target limb) another 200–300 msec later. Therefore, the 500–600 msec stimulus ends before or near the time of target limb contact and usually evokes single, nonrepetitive movements of the wiping and target limbs. The stimulation parameters therefore approximate the pattern of stimulus successfully removed by the wipe. However, similar wipes can be obtained with shorter pulse trains. The parameters chosen here minimized habituation, sensitization, and decline of response.
Experiments were designed to test (1) the organization of correction responses by the wiping limb to obstacles in its movement path, (2) whether correction responses were produced in the isometric limb, and (3) whether correction responses could be expressed as a summation of force-field primitives.
Free limb testing. To test the organization of free limb corrections, a rod was used as a path obstacle and was attached to a six-axis force transducer (Assurance Technologies, Garner, NC). The rod was placed in the movement path of the wiping limb, and wiping movements were evoked. The initial postures of the wiping and target limbs before the reflex was evoked were similar for all trials and for each tested frog. The distance along the movement path of the wiping limb at which the rod was placed was systematically varied in each of the six frogs. The distance from the initial position of the heel to the rod was measured for all trials. For each distance that was tested in a frog, the rod was placed so that collision with the limb occurred 2–3 mm above the heel.
Isometric testing. To test whether correction responses could be evoked in the isometric limb, we placed the ankle of the wiping limb in a custom-made restraint. The restraint was secured around the perimeter of the ankle, 2–3 mm above the heel, where obstacle collisions occurred during free limb trials. The restraint was attached to the force/torque transducer by a custom-made shaft. We recorded isometric force vectors produced at the ankle after evoking the wiping reflex. The wiping limb was positioned in a number of initial limb postures (10–12) that spanned the reachable workspace of the wiping limb, and wiping was evoked at each posture. In these experiments, although the wiping limb was restrained, the target limb was not restrained. It was free to move and began from a similar initial posture for each trial. We found that wiping reflexes could be evoked in the isometric condition [also see Giszter et al. (1993) andKargo et al. (1998)]. In addition, we found that the restraint acted like a path obstacle and evoked a correction response that was similar to the response evoked by obstacle collision during free limb trials (see Data analysis and Fig. 4).
Cutaneous deafferentation. To test whether correction responses were produced by on-line summation of force fields, we performed a manipulation that selectively eliminated correction responses without disrupting wiping reflexes. Cutaneous feedback was removed from skin areas of the wiping limb in contact with the ankle restraint. Small incisions were made at several sites around the calf and foot, and lower-limb cutaneous nerves were dissected free, frozen with a cold copper rod to reduce injury discharge, and transected. We attempted to evoke flexion withdrawal responses from deafferented skin regions to test the completeness of the deafferentation. Both light (dragging sharp forceps) and deep pressure (pinching skin with blunt forceps) were applied to these skin regions. The skin incisions were sealed with wound clips and Vetbond. Isometric wiping trials were then performed 1–2 hr after this procedure, and isometric force vectors produced during the trials were measured. The wiping limb was held in the same series of initial positions as when cutaneous feedback was intact. Despite the fact that muscle afferent feedback was fully intact, no correction responses could be observed, even after collisions of substantial velocity.
Bone pin implantation. To control for nonspecific effects of the cutaneous deafferentation, we performed a number of experiments (n = 5) in which bone pins (Fine Science Tools, Foster City, CA) were implanted into the distal tibia to immobilize the wiping limb. We used the smallest skin incision compatible with the implantation procedure so that cutaneous feedback would be minimally disrupted. In these experiments, we built up a base around the bone pins with carboxylate cement (Durelon, Norristown, PA) into which we secured a metal standoff. The metal standoff could be attached directly to the force/torque transducer without any direct skin contact. Such frogs exhibited typical correction responses in free limb conditions after obstacle collision. However, if held immobile, no correction was evoked in the absence of skin contact.
In three of the frogs in which bone pins were used to immobilize the limb, we examined whether the amplitude and duration of correction responses depended systematically on the magnitude of obstacle collision. We used a stepper motor to drive a mass into the heel of the isometrically held wiping limb with a predetermined velocity (i.e., into skin areas where obstacle collisions occurred during free limb trials). A custom-designed program implemented in DOS controlled the stepper motor. This program was triggered at the onset of target limb stimulation via a signal on the parallel port of the computer controlling the stepper motor. Both the stepper motor velocity and the latency of impact relative to the onset of target limb stimulation were varied.
To summarize, a typical experiment consisted of (1) ∼20 unrestrained wiping trials with and without path obstacles and with sensory feedback intact, (2) ∼10 trials with the wiping limb immobilized in an ankle restraint to record endpoint isometric forces and with feedback intact, (3) ∼10 isometric trials after local cutaneous deafferentation of the wiping limb (so that cutaneous feedback stemming from skin contact with the restraint was absent), and (4) ∼20 unrestrained wiping trials with and without path obstacles and after the deafferentation. In five frogs, the limb was restrained via bone pins, thus eliminating any skin feedback from the restraint without having to deafferent skin regions. Obstacle contact was produced in three of these frogs by driving a mass into the isometric limb by a stepper motor.
Data collection
Limb kinematics, EMGs, and contact forces with the rod were recorded during free limb trials. Movements of the wiping and target limbs were videotaped at 60 fields per second perpendicular to the horizontal plane of the wiping movement. A light-emitting diode was triggered at the initiation of the wiping stimulus and allowed movements and EMGs to be synchronized within 1–16.67 msec. EMGs were amplified 1000× and bandpass-filtered (cutoffs 100 Hz and 10 kHz) using A-M Systems amplifiers (A-M Systems, Everett, WA). EMGs were analog-to-digital converted using a DAS16 A/D converter and stored on computer using custom-made software. Forces were sampled at 250 Hz, and EMGs were sampled at 1000 Hz. During isometric trials, forces generated at the ankle were recorded by using the same force/torque transducer to immobilize the ankle. Figure 1 shows the experimental conditions (with and without local cutaneous deafferentation) and the data types collected (free limb kinematics and isometric forces; EMGs were collected in both restrained and unrestrained trials).

The hindlimb to hindlimb wiping trials described here were evoked in spinalized frogs in two conditions: with sensory feedback intact and with a local cutaneous deafferentation of the calf/foot of the wiping limb (shaded area). Wiping was evoked in all trials by electrical stimulation of the target limb (see Materials and Methods). Two data types were collected: isometric forces and free limb kinematics. Joint angles were measured as shown and converted to engineering format as needed (e.g., for torque calculations). In trials in which the wiping limb was free to move, kinematic data were recorded after the wiping was evoked. In trials in which the wiping limb was restrained, isometric forces were recorded at the ankle. Hindlimb muscle EMGs were recorded during both trial types (free limb and isometric) and during both conditions (feedback intact vs cutaneous deafferentation).
Data analysis
Kinematic analysis. We examined changes in the free limb kinematics that occurred during obstacle collision trials. The hip and knee joint locations of the wiping and target limbs (in the horizontal plane of the limb) were digitized from video recordings using a custom-designed program. The joint coordinates from the digitization procedure were imported into the S-Plus Statistics and Display package (Statistical Science, Seattle, WA). These were used to reconstruct movement of the thigh and calf and to derive the hip and knee joint angles over time. Hip angle was measured as the angle between the longitudinal axis of the frog and the thigh (Fig. 1). Knee angle was measured as the internal angle between the thigh and calf (Fig. 1). Because there was little variation in trial-to-trial duration of the wiping movement for an individual frog, time normalization was not necessary. Thus, averages and SDs of the hip and knee joint angles over time were constructed after simply aligning individual trials at the onset of movement for each condition.
Force analysis (single limb position). We examined differences in the endpoint isometric forces generated during wiping trials and between conditions, i.e., when sensory feedback was intact and when skin regions contacting the ankle restraint were deafferented. We first examined differences in the direction and magnitude of force vectors generated at a single limb configuration. The direction and magnitude of the wiping force vectors were calculated for a 3 sec period after the reflex was evoked (wiping motor pattern typically lasted 600–1000 msec). Endpoint polar force direction was calculated in the plane of the limb (positive x-axis at 0°, longitudinal axis of frog at 90°) from the measured endpoint force at time (t) by:
 Equation 1where Fy andFx are the recorded horizontal force components over time. The magnitude of the force vectors was calculated by:
Equation 1where Fy andFx are the recorded horizontal force components over time. The magnitude of the force vectors was calculated by:
 Equation 2Averages and SDs of the force direction and magnitude at each time point after initiation of wiping were constructed by aligning individual trials at the onset of force. The onset of force for each trial was determined after the averaged baseline force (200 msec before wiping was initiated) was subtracted from the force record. Both force direction and magnitude were determined for active forces, i.e., when resting, baseline forces were subtracted.
Equation 2Averages and SDs of the force direction and magnitude at each time point after initiation of wiping were constructed by aligning individual trials at the onset of force. The onset of force for each trial was determined after the averaged baseline force (200 msec before wiping was initiated) was subtracted from the force record. Both force direction and magnitude were determined for active forces, i.e., when resting, baseline forces were subtracted.
On the basis of analysis of EMG patterns (see EMG analysis), we believed that feedback caused by skin contact with the ankle restraint initiated a corrective response during isometric wiping trials. We tested whether correction responses recruited an isometric force-field primitive and whether this primitive was simply superimposed on the underlying wiping forces. According to the definition used in this paper, a force-field primitive is observed as a structurally invariant force field over time. In its most general form, a force-field primitive would be a function of both position and velocity. In this paper, we confine ourselves to isometric force measurements at which velocity is always zero. Other experiments [for preliminary description, see Giszter et al. (2000)] show, as expected, that velocity plays a role in force-field magnitude. However, because the limb is brought to rest by an obstacle, the isometric measurement corresponds very well to the forces generated for the initiation of the correction movement. Under these isometric conditions, at a single limb position, the force vectors generated by a primitive will increase and decrease in magnitude with activation–deactivation dynamics of the primitive but, importantly, will have a fixed direction (Giszter et al., 1993). At the same time, force magnitude ratios among the sampled limb positions will remain constant, i.e., a primitive's time evolution can be expressed in the form a(t)φ(r) (Eq. 3). That is to say, a constant structure force field is scaled in amplitude through time in a similar way at all locations. To derive the endpoint forces generated by the evoked correction response under the hypothesis that force-field primitives comprised the wiping and corrective patterns, we therefore proceeded as follows. We subtracted the averaged force vectors generated during wiping trials alone, i.e., after cutaneous deafferentation, from the averaged force vectors generated during wiping trials with an evoked correction response, i.e., with cutaneous feedback intact:
 Equation 3
Equation 3
 Equation 4
Equation 4
 Equation 5where FC(r,t) represents the total forces generated in a corrected wipe over time (t) at configuration (r) with cutaneous feedback intact. This field is minimally composed of a sum of primitives θ(r)and φ(r) modulated by time varying scaling parametersb(t) and a(t), where θ(r) represents the corrective primitive. FN(r,t) represents the forces generated after local cutaneous deafferentation (without a correction response). The subtraction produces a time series of resultant force vectors FR(r,t) that represent the endpoint forces specifically associated with the corrective response. This procedure assumes that the endpoint forces sum linearly (Mussa-Ivaldi et al., 1994). The critical test of our hypothesis is the behavior and time evolution of the extracted corrective forces. We expect that the forces FR(r,t)can be expressed in the form b(t)θ(r) if the correction represents a force-field primitive as defined above, i.e., forces at each location have fixed directions, constant magnitude ratios, and similar time evolution.
Equation 5where FC(r,t) represents the total forces generated in a corrected wipe over time (t) at configuration (r) with cutaneous feedback intact. This field is minimally composed of a sum of primitives θ(r)and φ(r) modulated by time varying scaling parametersb(t) and a(t), where θ(r) represents the corrective primitive. FN(r,t) represents the forces generated after local cutaneous deafferentation (without a correction response). The subtraction produces a time series of resultant force vectors FR(r,t) that represent the endpoint forces specifically associated with the corrective response. This procedure assumes that the endpoint forces sum linearly (Mussa-Ivaldi et al., 1994). The critical test of our hypothesis is the behavior and time evolution of the extracted corrective forces. We expect that the forces FR(r,t)can be expressed in the form b(t)θ(r) if the correction represents a force-field primitive as defined above, i.e., forces at each location have fixed directions, constant magnitude ratios, and similar time evolution.
We first examined the direction of the resultant force vectorsFR(r,t) produced at a single limb position (direction calculated by Eq. 1) and in some frogs expanded this to full field descriptions (see below). We measured the variance of the resultant force directions for each frog tested. For each frog, there were 6–12 trials, trials were ∼800 msec long, and forces were sampled at 250 Hz. Thus the population of force vector samples for an individual frog comprised from 1200 to 2400 samples (0.8 × 250 × 12). Previous data showed that the forces generated at the ankle during microstimulation recruitment of a synchronously activated group of muscles exhibited 12° or less variance in direction (Giszter et al., 1993, their Table 1). Therefore, we chose this degree of variance (12°) as our criterion for data to support the hypothesis that the resultant forces (i.e., correction response forces) have a fixed direction over time. Force vectors were examined in these tests in both two and three dimensions. The results in three dimensions are qualitatively similar to those in two. We have therefore confined our presentation to the simpler two-dimensional data with the exception of force-field inner product measures (Eq. 7–10) in which measures that are presented apply to the three-dimensional force.
We also examined whether the x and y components of the resultant forces were caused by multi-joint or single-joint torque components in the plane of the limb. The static joint torques of the hip and knee were determined from the endpoint forces, limb position, and link lengths as below:

where lt andlc are lengths of the thigh and calf and Θh and Θk are the hip and knee angles using the measurement convention in Asada and Slotine (1986), Fx andFy are the derived resultant force components in the plane of the limb, andTH andTK are the hip and knee torques acting in the horizontal plane of the limb, i.e., omitting elevator and rotator hip torques acting out of this plane.
Force-field analysis. Isometric force fields generated with and without cutaneous feedback intact were examined to test whether a structurally invariant force field, i.e., a corrective response, was superimposed on the underlying wiping force field. Force fields were constructed for each condition as thoroughly described in Giszter et al. (1993). Data were time-aligned using the applied stimulus train. No time dilation was necessary because durations of responses did not vary across the workspace. Briefly, after a Delauney tesselation, a piecewise, linear interpolation procedure was applied to the sparsely sampled field of forces (10–12 wiping trials, each trial at a different limb position) at each time point after initiation of wiping (Giszter et al., 1993). This procedure allows estimation of force vectors across the convex region of the sampled workspace over time.
We subtracted the force fields generated after local cutaneous deafferentation from the force fields generated with all feedback intact at each time point after initiation of wiping (Eq. 3 and 4 applied at each position in the workspace). The force-field subtraction produces a time series of corrective, or resultant, force fields. We quantified the similarity of corrective force fields over time. A detailed description of the comparison procedure is presented inMussa-Ivaldi et al. (1994) (Eq. 3–5). Briefly, an inner product measure is calculated between two force fields:

where “●” represents the inner product of two vectors.Ft andFt+m denote the two resultant force vectors that are compared at times t and t +m at N locationsx1, x2, … xN.
The cosine of the angle between the two sampled fields is calculated:
 Equation 8where:
Equation 8where:
 Equation 9represents the norm of a sampled force field. If the correction is generated by a field, which can be expressed in the form of Equation5 above, then at two chosen time points (t) and (t + m):
Equation 9represents the norm of a sampled force field. If the correction is generated by a field, which can be expressed in the form of Equation5 above, then at two chosen time points (t) and (t + m):
 Equation 10
Equation 10
 because the scalars b(t) andb(t + m) determining force magnitude do not affect the result of the cosine operator in Equation 8. The value of 1 indicates that the two fields tested are simply scaled versions of one another. If the value of the cosine measure from Equation 8 was 0.9 for two test fields, these were considered to have a similar force-field structure, as described in Mussa-Ivaldi et al. (1994). A consistent value of 0.9 over time thus supports a stable force-field structure over time, as in Equation 5.
because the scalars b(t) andb(t + m) determining force magnitude do not affect the result of the cosine operator in Equation 8. The value of 1 indicates that the two fields tested are simply scaled versions of one another. If the value of the cosine measure from Equation 8 was 0.9 for two test fields, these were considered to have a similar force-field structure, as described in Mussa-Ivaldi et al. (1994). A consistent value of 0.9 over time thus supports a stable force-field structure over time, as in Equation 5.
EMG analysis. To identify motor pattern changes during trajectory corrections, EMG signals for individual trials were rectified and filtered in S-Plus using a moving average filter [30 point, tapered boxcar filter, (Basmajian and DeLuca, 1985)]. The rectified, filtered signals were averaged for unobstructed and obstructed wiping trials. Trials were aligned at the time of onset of the first muscle (ST) in each frog's respective motor pattern. Although laborious, onset and offset were determined visually from rectified and filtered data in S-Plus. We found this more reliable than fixed algorithms using a baseline and noise-based threshold. Within a frog, EMG bursts determined in this way had low variance (see Table 2). The magnitude of the EMG envelope (the integrated area under the rectified, filtered signal) from the time of EMG onset to offset was determined for each of the 10 muscles. When EMGs were subtracted between conditions, the variance of the EMG differences was estimated as the summed variance of the individual conditions. Statistical differences in the magnitude and onset and offset times of EMG envelopes between conditions were tested by using standard, pairedt tests implemented in the S-Plus environment.
As stated earlier, we hypothesized that correction responses recruited a force-field primitive. Force-field primitives are characterized by structurally invariant force fields over time. Either a single muscle or a group of coactive and linearly covarying muscles may generate invariant force-field structures over time (Giszter et al., 1993;Pellegrini and Flanders, 1996; Loeb et al., 2000). We examined whether multiple muscles were recruited during correction responses and whether these muscles' activations exhibited significant covariation. To test this, we correlated muscles' rectified and filtered EMG waveforms over time for correction trials using a standard linear regression algorithm in S-Plus Statistical Software.
To examine the waveform shape of EMGs and their temporal evolution, we proceeded as follows. We aligned individual rectified and filtered EMGs at their peak or initial peak if they exhibited multiphasic activity. We normalized each EMG to its own peak value and then averaged the normalized waveforms for each muscle. SDs of the normalized waveforms were calculated at each time point. As a result of normalization, SD at peak was always zero.
RESULTS
Kinematic observations of the trajectory correction response
We first analyzed the kinematic features of corrective responses during the hindlimb-to-hindlimb form of wiping. These corrections have never been described in any detail. This kinematic analysis will form the basis of subsequent analyses. In this form of wiping, when an irritant is applied to the foot of the target limb, the wiping limb attempts to reach and remove this stimulus. Figure2A shows the normal, free limb kinematics of the wiping and target limbs (i.e., the thigh and calf of each limb) after we evoked this reflex behavior using our standard electrical stimulation (500 msec duration, 2 msec biphasic pulses, 33 Hz, 5 V). The ankle of the wiping limb followed a fairly direct trajectory to a region caudal to the cloaca where target limb contact occurred. In the trial shown, target limb contact occurred 233 msec after the onset of wiping limb movement. After contact with the stimulus on the target limb, the ankle of the wiping limb followed a trajectory that was directed away from the body of the frog. This final portion of the trajectory [“whisk”; see Berkinblit et al. (1986)] normally acts to remove the stimulus.

Path obstacles evoked trajectory correction responses during hindlimb wiping movements.A–D, Digitized stick figures of the wiping and target limbs (thigh and calf) and the ankle path of both limbs (traced by a bold line; arrows mark direction of movement along path) are shown after initiation of the wiping reflex in four experimental manipulations. Each stick figure is separated in time from the next by 16.67 msec (
 sec).Bold stick figures mark the initial limb configurations and their configuration at target limb contact. The frog's body would be above each stick figure. A, The unobstructed kinematics of the wiping limb with feedback intact consisted of a knee flexion and hip extension that resulted in the ankle moving along a trajectory directed to a midline region of stimulus contact. A final knee extension (“whisk”) occurred on stimulus contact.B, When the wiping limb collided with a path obstacle en route to stimulus contact, a hip flexion was evoked that enabled the limb to clear the obstacle. Once clear there was an enhanced knee flexion. Target limb contact occurred at a similar midline site and at a time similar to unobstructed trials (see Results). C, After local removal of cutaneous feedback from the wiping limb, unobstructed free limb kinematics showed no apparent differences from the kinematics seen with feedback intact (compare A,C). D, When obstacles were placed in the path of the locally deafferented limb, corrective responses were no longer evoked, and the limb was driven further into the obstacle (compare B,D).
sec).Bold stick figures mark the initial limb configurations and their configuration at target limb contact. The frog's body would be above each stick figure. A, The unobstructed kinematics of the wiping limb with feedback intact consisted of a knee flexion and hip extension that resulted in the ankle moving along a trajectory directed to a midline region of stimulus contact. A final knee extension (“whisk”) occurred on stimulus contact.B, When the wiping limb collided with a path obstacle en route to stimulus contact, a hip flexion was evoked that enabled the limb to clear the obstacle. Once clear there was an enhanced knee flexion. Target limb contact occurred at a similar midline site and at a time similar to unobstructed trials (see Results). C, After local removal of cutaneous feedback from the wiping limb, unobstructed free limb kinematics showed no apparent differences from the kinematics seen with feedback intact (compare A,C). D, When obstacles were placed in the path of the locally deafferented limb, corrective responses were no longer evoked, and the limb was driven further into the obstacle (compare B,D).
Spinal frogs exhibited trajectory correction responses when obstacles were placed in the movement path of the wiping limb. In Figure2B such a trial is shown. The trajectory of the ankle, after contact with the obstacle, was redirected toward the head of the frog. This rostral redirection of the ankle trajectory was caused by a hip flexion, which was not seen in the unimpeded movement. Once clear of the obstacle, there was an enhanced knee flexion, and the ankle trajectory exhibited an increased velocity toward the target region (compare distance between digitized ankle positions). Interestingly, the time of target limb contact as determined from the video record was similar to the unimpeded trial. In the frog shown in Figure 2, the averaged difference in the time of target limb contact between unimpeded and impeded wiping trials (in successful obstacle avoidance) was only 30 msec.
In this portion of the study we examined nine frogs that all exhibited successful correction responses when path obstacles were placed close to the ankle of the wiping limb, i.e., with obstacles placed ∼2–3 cm or less from the initial position of the ankle. The straight-line distance from the initial position of the ankle to the site of target limb contact ranged from 10 to 13 cm for the nine frogs. In each frog, two common kinematic features characterized successful correction responses (Figs. 2,3A,B,bottom panels): (1) an evoked hip flexion that allowed the ankle to clear the obstacle and (2) an enhanced knee flexion once the obstacle was cleared. The enhanced knee flexion allowed the limb to extend at the hip without further collision and also resulted in an increased ankle velocity toward the target region. Taken together, these effects acted to maintain target limb contact times that were close to contact times in unimpeded wiping trials. In five of nine frogs there was only a small increase of ∼30 msec (significant atp < 0.05) in the mean latency to target limb contact (mean of 283 ± 20 msec SD with path obstacles compared with 250 ± 13 msec SD without obstacles) (Table1). Thus, although there was a small and significant difference, these frogs were able to maintain contact times within the extremes of the range of variations seen during unobstructed trials. However, in the other four frogs there were larger increases (p < 0.01) in the mean latency to target limb contact after movement initiation (mean 316 ± 25 msec SD with path obstacles compared with 245 ± 12 msec SD without). The increased latency to target limb contact in these four frogs was mainly attributable to an increase in the time to clear the obstacle (mean duration of 125 msec in contact with the obstacle in this group compared with a mean duration of 66 msec in the five frogs with almost normal movement durations).

Wiping motor patterns and evoked correction responses were similar for free limb and isometric trials. Rectified EMGs are shown for six wiping limb muscles after the wiping reflex was evoked in four separate conditions: (A) unobstructed free limb trial, (B) free limb trial with obstacle collision, (C) isometric trial, and (D) isometric trial after local cutaneous deafferentation of the wiping limb. Below the EMG panels inA and B are shown obstacle contact forces and hip and knee joint kinematics (in A, there was no obstacle collision). First and second vertical lines mark movement onset and target limb contact, respectively. Below the EMG panels in C andD are shown the recorded ankle force components (Fx, Fy, and Fz) and the derived hip and knee torques. A, The wiping motor pattern consisted of three phases: initial knee flexor (ST), subsequent hip extensor (RI), and final knee extensor (VE) activation. Hip flexors (IL) exhibited phasic activity near the time of stimulus contact. B, The timing of wiping motor phases was unaltered by obstacle collision. Hip flexor muscles (IL, GL, and SA) showed a strong synchronous activation after obstacle contact (see contact force) that ended near the onset of RI. Their activation was associated with a reversal of hip joint motion to flexion. After obstacle clearance there was an enhanced knee flexion and resumed hip extension. C, The time of onset of knee flexor (ST) and hip extensor phases (RI) was unaltered during isometric trials. The knee extensor phase (VE) was absent. Hip flexor muscles showed a strong activation after the onset of force, which represented the correction response that ended near the onset of RI. The time of hip flexor activation was associated with production of a hip flexor torque (bolded line below), whereas RI activation was associated with the onset of hip extensor torques. D, The time of onset of knee flexor and hip extensor phases was unaltered after local cutaneous deafferentation. Hip flexor activity and hip flexor torque associated with the correction response were eliminated by the deafferentation.
Target-to-target limb contact
The movement path of the contralateral (i.e., target) limb did not exhibit any major differences during wiping trials with and without the path obstacle (Fig. 2), and in this study, we focus only on corrective changes within the wiping limb.
Trajectory correction responses are evoked by cutaneous feedback
The sensory basis of the evoked correction response in frogs has not been described. Muscle, joint, or skin feedback could all contribute to initiating and controlling the response. We determined that the correction response in the wiping limb was initiated primarily by feedback from cutaneous receptors. We could demonstrate this by local removal of cutaneous feedback from the calf and foot of the wiping limb alone (see Materials and Methods). The successful correction responses depended on cutaneous feedback stemming from the obstacle collision. In the frog shown in Figure 2, after cutaneous deafferentation, the wiping limb did not exhibit a successful corrective response and was in fact driven further into the path obstacle by the extending hip.
Cutaneous feedback, although necessary for evoking successful correction responses, was not necessary for controlling the unimpeded free limb trajectory. In Figure 2C, the unimpeded kinematics of the wiping limb are shown after the local cutaneous deafferentation. The kinematics of the wiping limb did not exhibit any major differences, e.g., in the latency to target limb contact (233 msec), in its position at target limb contact (knee angle 50°, hip angle 42°), and in the shape of the endpoint path, from the trials in which cutaneous feedback was intact [see also Kargo and Giszter (2000)]. In summary then, spinal frogs exhibited smoothly integrated trajectory corrections during hindlimb wiping that were initiated by skin contact with an obstacle. The removal of skin feedback eliminated successful correction responses but did not interfere with control of the unobstructed wiping trajectory.
To show that loss of corrections was not attributable to nonspecific effects of the loss of cutaneous feedback, we performed a second series of experiments. Rather than using cutaneous deafferentation, we used bone pins placed in the tibia to immobilize the limb during the wiping reflex just as an obstacle would, but without skin contact. At the same time, skin sensory fields were largely undisturbed. We were unable to initiate corrective EMGs during isometric holds (see below) in this arrangement in the absence of cutaneous contact. However, true collisions evoked corrections. These approaches allowed us to manipulate wiping and corrective responses in isometric experiments.
Basis for isometric examination of wiping and correction responses
In the following sections, we tested whether the observed correction response forcesFC(r,t) were generated by the on-line superposition of a structurally invariant, corrective force field with an underlying wiping force field. Specifically:
 Equation 11where FN represents the normal pattern of forces over time (t) at position (r) activated by the skin stimulus on the contralateral limb andFR represents the force-field primitive activated by ipsilateral obstacle contact. To test this hypothesis we recorded the endpoint isometric forces generated during wiping and trajectory correction responses. Previous studies have shown that forces and EMGs appropriate for wiping can be evoked in the isometric limb (Giszter et al., 1993; Giszter and Kargo, 2000). We wished to test the organization of correction responses using isometric methods. To do this we needed to establish the similarity of motor patterns for wiping and correction responses in free limb and isometric conditions. We therefore analyzed the motor patterns evoked under each condition in the same frog using EMG recordings.
Equation 11where FN represents the normal pattern of forces over time (t) at position (r) activated by the skin stimulus on the contralateral limb andFR represents the force-field primitive activated by ipsilateral obstacle contact. To test this hypothesis we recorded the endpoint isometric forces generated during wiping and trajectory correction responses. Previous studies have shown that forces and EMGs appropriate for wiping can be evoked in the isometric limb (Giszter et al., 1993; Giszter and Kargo, 2000). We wished to test the organization of correction responses using isometric methods. To do this we needed to establish the similarity of motor patterns for wiping and correction responses in free limb and isometric conditions. We therefore analyzed the motor patterns evoked under each condition in the same frog using EMG recordings.
Free limb EMG patterns generated during wiping and trajectory corrections
We first examined the basic motor pattern that occurs during free limb wiping trials. The normal wiping motor pattern consisted of three periods of muscle activity (for detailed analysis, also see Kargo and Giszter (2000)]: an initial phase dominated by knee flexor activity (ST and BI), a second phase during which the hip extensors (RI, SM, and AD) become activated, and a final phase during which the knee extensors (for example, VE) become activated. The first two phases contribute to the hindlimb trajectory up to target limb contact (i.e., the aimed portion of the wipe), and the final phase is initiated around the time of target limb contact. Figure 3A shows the rectified EMGs of six muscles in the wiping limb during individual trials without a path obstacle. The corresponding hip and knee joint angles of the wiping limb are shown below the EMGs. Figure 3B shows the effect of an obstacle on this motor pattern and joint coordination.
A specific group of hindlimb muscles was commonly activated in response to obstacle collision during free limb wiping trials. With obstacles placed 2–3 cm from the initial position of the ankle, collisions occurred during the initial knee flexor phase (see contact force below EMGs in Fig. 3B). In the frog shown in Figure3A,B, a group of hip flexor-related muscles (IL and GL, monoarticular hip flexors, and SA, a hip/knee flexor) were reflexively activated ∼15 msec after obstacle contact. The duration of their activation was ∼150 msec. The reflex activation of these muscles did not appear to interfere with or cause variation of the onset times of subsequent wiping motor phases (see section entitled Correction responses are generated by addition of a muscle synergy to the basic pattern and Table 2). None of the comparisons of motor pattern latency differed significantly between correction and free limb, unobstructed trials.
Latency in milliseconds of the first two motor phases of wiping in four different conditions
Motor patterns are similar under isometric conditions
After evoking a series of free limb wiping trials in each frog, we immobilized the wiping limb using our ankle restraint. This apparatus allowed us to examine isometric forces and force-field patterns. We again recorded EMGs and also isometric forces generated at the ankle after evoking the wiping reflex. We found that the initial knee flexor phase and the subsequent hip extensor phases of the wiping motor pattern were initiated in all frogs and remained similar to free limb patterns (see Fig. 3C, restrained trial, and compare with Fig. 3B, free limb trial with correction). The hip extensor phase maintained a similar time of onset relative to knee flexor onset in eight of nine frogs (Table 2). With the exception of bf74, the ST to RI latency did not differ significantly between free limb, correction, and isometric tests. However, in the majority of frogs (eight of nine), the usual, final phase of knee extensor activation was absent (Fig. 3C,D). In the one exception, the knee extensor phase was much reduced, although present. Loss of knee extensor activation was often associated with a more abrupt termination of knee flexor and hip extensor EMG activity (compare ST and RI EMGs in Fig. 3C,D, isometric, with Fig. 3A,B, free limb). The lack of the knee extensor burst representing initiation of the final motor phase suggests that either triggering or aborting of this phase of the motor program may occur based on successful or unsuccessful achievement of target limb contact and/or a specific wiping limb configuration. These observations are compatible with the kinematic observations reported in Giszter et al. (1989). Despite the similarity of timing between isometric and free limb conditions, there were some amplitude variations in EMGs in isometric conditions. These are analyzed elsewhere.
We also examined the EMGs recorded during isometric wiping trials to assess whether the ankle restraint evoked a correction response during wiping. The ankle restraint was placed in contact with the same skin areas (∼2–3 mm above the ankle) upon which obstacle collision occurred during the free limb trials. We found that the same hip flexor-related muscles that were activated after obstacle collision during free limb wiping were activated during isometric wiping in our tested frogs. In the frog shown in Figure3C,D, these hip flexor muscles were activated during the initial knee flexor phase, ∼90 msec after ST onset and 65 msec after force onset (average onset time of 55 ± 11 msec relative to first measured force onset for the nine frogs taken together). The evoked EMG activity always ended close to the onset of the hip extensor phase. In fact, in eight of nine frogs of this study, the duration of activation of these muscles during isometric trials was similar to their duration of activation during collisions in free limb trials, e.g., the averaged duration of the evoked IL EMG during free limb trials was 154 ± 16 msec, whereas in isometric trials it was 165 ± 20 msec (Table 2) (p > 0.1).
In addition to the EMG similarities, correction responses initiated during isometric wiping could be abolished by cutaneous deafferentation (Fig. 3D), similar to free limb trials. The deafferentation eliminated pertinent skin feedback caused by contact with the ankle restraint. In the frog shown in Figure 3D, the IL, SA, and GL EMGs no longer exhibited a strong, phasic activation after cutaneous deafferentation. However, the knee flexor and hip extensor phases of the wiping motor pattern were clearly unaffected and had similar onset and offset times. An examination of the derived hip and knee torques generated during isometric trials showed the torque patterns produced when cutaneous feedback was intact and when this feedback was eliminated by the deafferentation procedure (Fig.3C,D, bottom panels). With feedback intact, a hip flexor torque was initiated soon after the initial knee flexor torque and was associated with the onset of IL, GL, and SA. After removal of cutaneous feedback, this strong hip flexor torque was absent, as were the enhanced activities of IL, GL, and SA. The wiping motor pattern also involved a substantialz-direction force (Fig. 3C,D). Variations in the z-force associated with corrections were small (Fig. 3, compare C and D,Fz ).
In summary, our analysis showed that wiping motor patterns can be initiated in the isometric limb, and these differ primarily only in amplitude from free limb EMGs. The first two phases of the motor pattern, the initial knee flexor (ST and BI) and subsequent hip extensor (RI, SM and AD) phases, had similar onset times as in the free limb. A different report (Giszter et al., 2000) documents in detail the differences between free limb and isometric wiping trials and the role of proprioceptive feedback in force-field structure within a phase of the motor pattern. When cutaneous feedback was left intact during isometric trials, the ankle restraint acted like a path obstacle and evoked a correction response. The same set of muscles activated during free limb trajectory corrections was activated during isometric corrections and with a similar duration (Fig. 3, compare Band C). Last, the correction response could be eliminated by local cutaneous deafferentation of the ankle/calf region without affecting the timing of the underlying wiping motor phases.
In the following sections we exploit our observation that wiping and correction motor patterns appear conserved in the isometric limb. We used this to test the hypothesis that on-line superposition of force fields underlies the generation of trajectory corrections during wiping. We test this first at a single limb position and then at multiple limb positions.
Superposition of isometric endpoint forces underlies trajectory corrections
We examined the endpoint forces generated during wiping trials with cutaneous feedback intact, i.e., when correction responses were evoked, and after cutaneous deafferentation at a single, standard limb position (hip 90°, knee 105°) in nine frogs. In each frog, when cutaneous feedback was intact we observed a temporal pattern of endpoint forces similar to those shown in Figure4B. We plotted both the angular directions and amplitudes of force vectors over time. Figure4C shows the directions, expressed as an angle, over time in each condition. Figure 4D shows the magnitudes. We also expressed the angular variations of force direction as distributions in circular histograms (Fig.5). In the frog shown in Figure4B, the force vectors generated by the wiping limb after evoking the wiping reflex were initially directed toward the midline (∼160°), where target limb contact normally occurs, and remained in this direction for ∼75 msec (Fig. 4B,WCF). The forces then began to rotate toward the head of the frog over the next 150 msec. This continuous rostral rotation immediately followed the activation of IL, a hip flexor, and SA, a hip/knee flexor (averaged period of activations shown in Fig.4E as bars). At the onset of the hip extensor burst (RI), the endpoint forces rotated in an opposite direction, away from the frog, and back toward and through the target region. They stabilized at a final direction of ∼260° (measured counterclockwise and relative to horizontal, 0°) (Fig.4C).
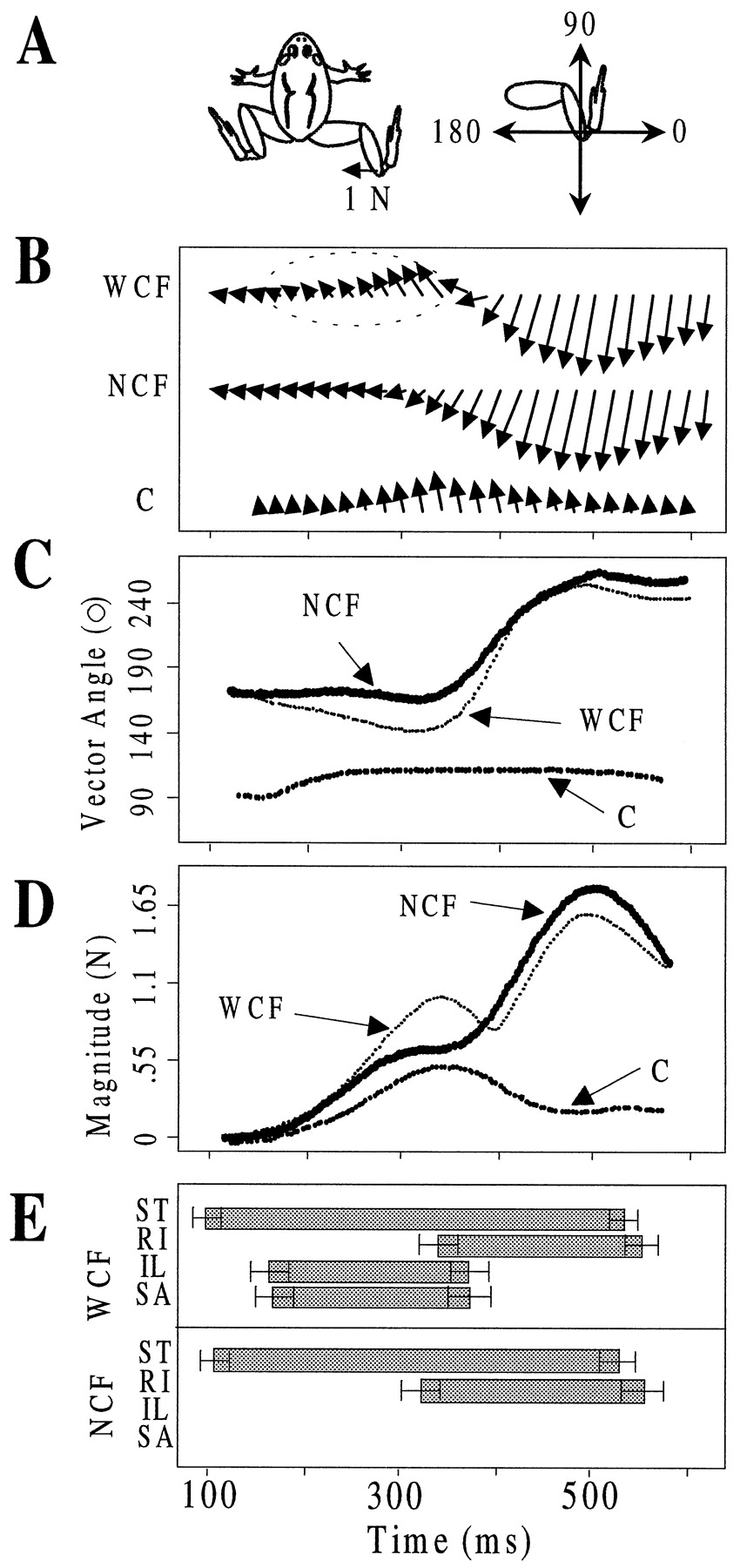
Cutaneous reflex pathways recruited a fixed-direction force component that summed with an underlying sequence of forces generated during wiping. In this figure, time is referenced to stimulus onset. A, Force vectors in Bare plotted with respect to the frog shown. Thus, vectors pointing to the left of the page are pointing to the midline of the frog. The force vector shown here represents the scale forB. Vector directions in C and Figures 5and 6 are referenced relative to the axes in this panel.B, Isometric force vectors are shown at 20 msec intervals after initiation of wiping for two conditions (top two rows; WCF, with feedback intact;NCF, no cutaneous feedback). Forces in each condition represent an average of seven trials. In a correction response (WCF), force vectors were initially directed to the midline, rotated rostrally toward the hip, and then rotated back to and through the midline to caudal directions. In the absence of a correction (NCF), force vectors were initially directed to the midline for 200 msec and then rotated caudally. In the third row, vectors, C, produced by a subtraction of NCF vectors from WCF vectors are shown. These vectors represent the correction response. Note that these appear uniformly directed. C, Angular directions (see A) of the force vectors for the two conditions and for the correction vectors are shown at 4 msec intervals. In all trials (WCF and NCF), force vectors were initially similarly directed to 180°. In corrections (WCF), force vectors rotated to ∼140°, whereas NCF vectors remained fixed in direction. In both cases (WCF and NCF), force vectors then rotated caudally at around the same time to a direction of 260°. Corrective, C, vectors as noted in Bremained stable in time. D, Vector magnitudes are shown over time. With no correction (NCF), force vectors exhibited a biphasic pattern of growth. Each phase is associated with a period of fixed-direction force production as shown in B. In corrections (WCF), initial phase of vector growth is increased because of the summation of C vectors along a similar direction. The second phase is decreased because correction and wiping vectors are oppositely directed. C vectors show a monotonic increase and decline. E,Bar plots representing the averaged period of activation of ST, RI, IL, and SA EMGs are shown. The horizontal bars at each end represent ±1 SD. With feedback intact, the onset of IL and SA activation was associated with the onset of the rostral force rotation. After cutaneous deafferentation, IL and SA activation were absent, as was the rostral rotation of forces. RI onset was similar in both conditions and associated with the onset of caudally rotated forces.

Corrective force components are directionally stable. A–C, Circular histograms of the directions of force vectors generated over wiping in five trials (600 msec duration trials, 250 Hz sampling rate; thus ∼750 samples total) are shown for each condition: with feedback and correction response (A) and without cutaneous feedback or a correction response (B). The derived corrective vectors are shown in C. Each bin represents 12°. SeeD for orientation of the circular histograms to the frog's body and wiping limb. A, With feedback intact, vector directions exhibited two clusters: an initial cluster directed toward the midline and a second cluster directed away from the frog. The first cluster was broadly spread and spanned four bins.B, Without cutaneous feedback, the initial cluster of vector directions was more focally distributed to the midline, i.e., mainly in one bin (78–90°), whereas the second cluster of forces was similar to that with feedback intact (252–264°). C, Corrective vectors were obtained by subtracting time-matched forces inB from A. The directions of the corrective vectors clustered tightly in a single bin from 108 to 120° and were directed to the hip joint. Thus, the corrective vectors summed with the initial set of wiping vectors directed to the midline and resulted in the rostral spread of these midline-directed force clusters when feedback was intact.
After local removal of cutaneous feedback, the pattern of forces generated by the wiping limb consisted of only two sequentially activated and uniformly directed force components (Fig.4B, NCF). These could correspond to stable force fields in each phase. In the absence of a corrective response, the initial forces after the wiping reflex was evoked grew in magnitude along a uniform direction (∼160°) for ∼200 msec (Fig.4B,C). As the hip extensor burst was activated, the forces rotated away from the frog and reached a second set of uniformly directed forces (∼260°) for the remainder of the period of RI activation. There was no period of IL/SA activation and no corresponding rostral rotation of endpoint forces without cutaneous feedback.
We analyzed the distribution of the force directions expressed as angles (as in Fig. 4C) that were generated during wiping trials both with and without cutaneous feedback intact. After cutaneous deafferentation, i.e., no correction response, the force directions clustered into two discrete subgroups (Fig. 5B). The forces in the initial (midline-directed) subgroup were contained mainly within a single 12° bin directed to 168–180° (mean direction of 175 ± 5 SD). Forces in the second (caudally directed) subgroup were mainly contained within two bins directed to 252–276° (mean direction of 261 ± 7 SD). When feedback was intact (Fig. 5A), the initial subgroup of forces directed toward the midline was more broadly distributed than the corresponding initial subgroup of forces produced when cutaneous feedback was removed (average direction of 156 ± 13 SD) (Fig. 5A). The subsequent caudal subgroup of forces with feedback intact was similarly directed to the corresponding caudal subgroup produced with cutaneous feedback removed, although the variance was greater (mean direction of 258 ± 13 SD).
We hypothesized (Eq. 10; also see Eq. 4 and 5) that differences in the variance and direction of the initial subgroup of forces produced with and without cutaneous feedback intact were caused by differences in the contribution of two fixed-direction force components (i.e., of low directional variance). When cutaneous feedback was intact, a corrective force component was activated and summed through time with an initial wiping component. This summation resulted in the rostral rotation and increased variance of the midline-directed force vectors. To test this, we subtracted the time series of force vectors that were generated when cutaneous feedback was removed from the force vectors that were generated at the corresponding times with feedback intact. We examined the time-varying pattern of resultant (corrective) forces, i.e., direction and magnitude. These are shown in Figure4B–D (series marked C). In the frog shown in Figures 4 and 5, the corrective force vectors exhibited a monophasic increase and decrease in amplitude and were uniformly directed to ∼115° during the period of IL/SA activation (Fig.4C,D). The distribution of the corrective force directions over time showed low variance (Fig. 5C). Eighty-one percent of the corrective vectors (n = 675 vectors; 5 trials × 135 force samples; 540 msec long trials) were contained in a single bin (118–130°). The mean direction was 120°, and the SD was only 3°.
To summarize, cutaneous feedback triggered the activation of a fixed-direction force component associated with a correction response. This corrective component summed with an ongoing force component produced during the initial phase of wiping. The subsequent onset of the hip extensor phase of wiping was associated with the production of a final, fixed-direction force component. Thus these data indicate that three fixed-direction force components are activated both synchronously and sequentially to generate correction responses during wiping. This applies to isometric conditions and when testing at a single configuration.
We examined how the force component recruited during correction responses varied among frogs. Polar plots of the distributions of corrective force directions are shown for four frogs in Figure6A. The limb posture at which the wiping trials were evoked is shown in the inset of Figure 6A (90° hip and 105° knee angles) and was the same for each frog. For six of nine frogs tested, a large percentage (>80%) of the corrective forces over time (and for multiple trials) were contained in a single bin. For each of these frogs, the variance of the corrective force directions was <12°. In addition to the uniformity of corrective force directions over time for each frog, the corrective vectors were similarly directed among frogs (the modal bin for all frogs lay within a 30° segment).

Cutaneous reflex pathways that signal obstacle contact recruited similar corrective force components among frogs.A, Circular histograms of the directions of corrective forces over time are shown for four frogs (obtained as described in Figs. 4 and 5 and Materials and Methods). For each frog, the collection of corrective force directions clustered mainly within one bin and the modal bins for each frog in the group were localized within a 36° segment (from 84 to 120°). B, The averaged corrective hip and knee torques are shown for each of the four frogs shown inA. Knee and hip torques are normalized to the maximal hip torque produced for each frog. For each frog, the corrective forces were produced by a multi-joint torque response. In addition, in each frog the knee to hip torque ratios were constant over time (linear up to the peak corrective force). The small variability among frogs in the direction of corrective forces was related to the magnitude of the ratio of knee flexor torque to hip flexor torque, and in some frogs (bf65) also to the direction of the knee torque component, e.g., extensor versus flexor.
The other three frogs showed deletions of the extensor phase. Extensor phase deletions will not be further pursued here but are reported in more detail in Giszter and Kargo (2000).
A hip flexion torque was primarily responsible for generating the corrective endpoint forces. Figure 6B shows the balance of corrective knee torque versus hip torque, displayed from the onset of corrective force up to the peak corrective force. In each frog, there was an almost constant ratio of hip flexion torque to knee torque over time. This constant joint torque ratio resulted in the uniformly directed corrective forces. However, the direction (flexor vs extensor) and relative magnitude of the corrective knee torque exhibited some variability among frogs (knee torques are normalized to peak hip torque in Fig. 6B). In four of six frogs, a corrective knee flexion torque accompanied the hip flexion torque. In the other two frogs, there was a small knee extensor torque component. This variability may be related to differences among frogs in the coactivation of other muscles in addition to monoarticular hip flexors (IL and GL) during correction responses. For instance, SA, a hip flexor/knee flexor, and RA, a hip flexor/knee extensor, were both commonly activated during corrective responses. The balance of these muscles' activations would then determine the resulting knee torque. We examine in more detail the EMGs underlying corrective responses (in the sections entitled Correction responses are generated by addition of a muscle synergy to the basic pattern and Muscles recruited during correction responses exhibit covariation).
Superposition of isometric force fields underlies trajectory corrections
Although the correction responses at a single location showed clear superposition of forces, this guarantees nothing about the forces generated at other configurations or the stability over time of a force-field primitive. We therefore examined the force fields generated during wiping trials with and without cutaneous feedback intact, i.e., with and without evoked correction responses. Force fields were constructed by measuring the isometric forces generated at a range of initial limb positions. Thus, force-field measurements describe the configuration-dependent effects of both muscle actions and changes in muscle activation (caused by changes in proprioceptive feedback). Conceivably, responses might differ radically across the workspace, and field structure might be complex or divergent.
In Figure 7, a time series of force fields generated by the wiping limb with cutaneous feedback intact (Fig. 7A) and after cutaneous deafferentation (Fig.7B) are shown for one frog. The force fields were initially similar between conditions. Approximately 90 msec after the onset of force, the force fields began to exhibit apparent differences. With feedback, intact force fields rotated rostrally toward the hip joint, whereas force fields produced with no cutaneous feedback continued to grow in magnitude and remained directed toward the midline region of stimulus contact. The time points at which the force fields are shown, i.e., 130–330 msec at 40 msec intervals, are marked as successive vertical lines on an EMG record collected at a single, central location in the workspace (Fig. 7; EMG record to the right of each force-field series). The EMG record includes four muscles that represent the knee flexor phase (BI), the hip extensor phase (RI), and the corrective response (IL and RA). The alteration in force-field structure between conditions was associated with the reflex activation of IL and RA when feedback was intact, i.e., a correction response.

Cutaneous reflex pathways signaling obstacle contact recruited a corrective force field that summed with an underlying force-field sequence produced during wiping. Time series of force fields are shown after initiation of wiping both with feedback intact and after cutaneous deafferentation. A typical EMG pattern of representative muscles associated with field generation is shown to theright of each time series. For the fields, a single vector represents the force generated at that position at the specified latency after wiping initiation. The black dot marks the location of the hip joint. The lightly shaded ellipseindicates the region of contact with the target limb in free limb trials. A, With feedback intact, force fields were initially directed to the midline where target limb contact normally occurred (bold line overlaid on the 130 msec field shows the unrestrained ankle path up to limb contact. At 210 msec up to 330 msec, force fields rotated rostrally toward the hip and away from the target region. At 330 msec, force fields began to rotate back to the target region. The hip extensor burst began around this time (see EMG record to the right; rectified EMGs fromBI knee flexor, RI hip extensor, andIL/RA-hip flexors are shown for a single trial taken from the middle of the workspace). B, After removal of cutaneous feedback, initial force fields were directed to the midline. Subsequent fields (210–330 msec) converged to the target region and did not rotate to the hip joint. The strong IL/RA activation was absent without cutaneous feedback (see EMGs to the right). At 330 msec, the field began to rotate caudally because of hip extensor activation. C, Procedure used to produce corrective force fields. Time-matched fields in B were subtracted from corresponding fields in A. The procedure was performed at each time point after wiping initiation. D, Corrective fields are shown at three times, at 40 msec intervals. Corrective fields were structurally similar (similarity measures > 0.95) over a 100 msec period starting shortly after IL EMG onset. These rostrally directed fields summed with the underlying wiping force fields.
We hypothesized that the alteration in force-field structure was caused by the reflex activation and superposition of a structurally invariant corrective field on a wiping field. To examine the time-varying structure of the corrective force fields, we subtracted vector fields between conditions (feedback intact fields − feedback removed fields; procedure graphically represented in Fig. 7C). In Figure 7D, corrective fields are shown at three successive time points (130, 170, and 210 msec after force onset). We used a correlation method to examine the similarity of corrective force fields over a 200 msec interval (IL and RA were activated for ∼200 msec) starting at 90 msec after force onset [see Materials and Methods,Mussa-Ivaldi et al. (1994)]. Over this time period, the correlations between corrective field structures at any two time points were 0.95 or larger. This high correlation measure between field structures suggests that the corrective force fields over time represent scaled versions of a structurally similar force field. This is in keeping with the low variance of corrective force directions at a single limb position described in Superimposition of isometric endpoint forces underlies trajectory corrections. In addition, it shows that the force magnitude ratios across the limb's workspace are relatively constant during the period of the corrective response. The constant structure of the extracted, corrective force field over time implies uniform temporal dynamics at all positions. This and the smooth unimodal rise and fall of its force magnitude at each location (Fig. 4) are both consistent with the hypothesis of force-field summation. All tested frogs showed the high correlations (>0.9) for corrective force-field structure over time (i.e., structure was conserved in the field resulting from the vector subtraction procedure). Finally, we found that the corrective and wiping force fields were similar among tested frogs (n = 4).
In summary, we addressed configuration-based variations in forces and corrective responses by measuring entire force fields. Examination of these force fields showed that trajectory corrections were generated by a dynamically evolving force-field structure. Our subtraction procedure showed that this dynamic force-field evolution was generated by combining the force-field primitive evoked in the correction response with the force-field primitives associated with the wiping reflex.
Correction responses are generated by addition of a muscle synergy to the basic pattern
The data above suggest that cutaneous feedback triggered the activation of a force-field primitive. This corrective force field sums with and does not disrupt the underlying wiping force-field sequence. In this section, we examine directly whether muscle activation triggered by obstacle collision sums with and does not disrupt the ongoing wiping motor pattern.
We found that two to three hip flexor-related muscles were activated in response to obstacle collision in each of the nine frogs tested in free limb conditions. The activation of these muscles was added to and did not interfere with the underlying motor pattern (Table 2; see list of reflexly activated muscles). In Figure8A,B, rectified, filtered, and averaged EMGs (n = 11 trials per condition) are shown for one frog during unobstructed, free limb wiping trials (bold lines, thin lines, 1 SD above and below average, respectively) and for obstacle collision trials, respectively (similar convention). The obstacle was placed next to but not touching the wiping limb. Collision occurred immediately after movement onset. Both movement onset and obstacle collision are indicated by the dotted vertical line. SA, GL, and RA muscles that all generate a hip flexor torque (Loeb et al., 2000) were activated ∼20 msec after obstacle contact (marked by the dotted vertical line). Reflex activation of these muscles during the correction appeared not to interfere with the timing or magnitude of the knee flexor phase (characterized by ST), hip extensor phase (characterized by RI), or knee extensor phase (characterized by VE) that comprised the wiping motor pattern (Fig.8A–C). We directly examined this by subtracting the individual muscles' averaged EMG waveforms in trials without obstacle collision from averaged EMGs obtained in trials with collision. The EMG differences are shown in Figure8D. Only SA, RA, and GL showed significant EMG changes during obstacle collision trials. Horizontal linesmark the maximum calculated SD for the difference for each muscle between the two conditions. EMG differences at peak for SA, GL, and RA were significant (p < 0.05).

Evoked muscle activities during correction responses sum with the underlying wiping motor pattern. On obstacle trials, the obstacle was placed next to but not touching the wiping limb. Collision occurred immediately after movement onset.A, Rectified, filtered, and averaged EMGs for unobstructed wiping trials. Averages were constructed from 10 trials in each condition. Lines indicating 1 SD above and below the mean value are shown (fine lines). The first vertical line (dotted) marks the averaged time of movement onset and obstacle contact, and the second vertical line (solid) marks the averaged time of target limb contact for both conditions. B, EMGs for obstacle collision trials are shown. EMGs were calculated as inA. C, The EMGs for both conditions are overlaid (bold lines/no shading: unobstructed trials; hairlines/shaded envelopes: obstacle collision trials). D, The EMG differences obtained by subtracting the unobstructed EMGs from the collision trial EMGs are shown. Horizontal lines mark the maximum SDs for the EMG differences between conditions (see Materials and Methods). Obstacle contact evoked the synchronous activation of hip flexor-related muscles SA, GL, and RA without significantly disrupting other EMG bursts of the wiping motor pattern.E, The peak EMGs of GL, SA, and RA muscles recruited during the correction response are plotted versus one another for multiple correction trials (n = 10). Peak EMGs were significantly correlated (all p < 0.01) over multiple trials for each of the three muscles. F, The normalized SA EMG is plotted versus the normalized GL EMG over time from the onset of the correction response up to peak activation for multiple trials (n = 10). The EMG activation waveforms up to peak EMG were highly correlated (p < 0.001).
Muscles recruited during correction responses exhibit covariation
The extracted corrective fields described earlier (see sections entitled Superposition of isometric endpoint forces underlies trajectory corrections and Superposition of isometric force fields underlies trajectory corrections) were structurally invariant over time. Structurally invariant fields are produced either by activation of single muscles or by the synchronous and linearly covarying activation of a group of muscles. We examined EMG patterns related to the correction. These were extracted as described in the preceding section (Fig. 8D). We analyzed these to confirm that muscle activation covaried as expected. Such a pattern of activation could result from a common motor drive that is distributed to a group of motor pools. In the following section, we examined whether muscles recruited during correction responses exhibited correlated activation, which could be associated with a common motor drive.
We correlated the SA and GL EMG waveforms from the onset of the corrective response up to their peak amplitude for the 11 correction response trials (Fig. 8F). The EMGs were normalized so the peak amplitude for each trial and for each muscle was equal to one. The GL to SA EMG correlation was highly significant (p < 0.001), and ther2 value was 0.85. This high correlation over time shows that the recruitment of muscles comprising the correction response was consistent with a common motor drive. SA and GL exhibited quite different EMG waveforms during unobstructed trials, and the muscles were recorded at distant sites (ventral/distal thigh, SA; dorsal/proximal thigh, GL). The high correlation of these muscles in correction trials could not therefore be attributed to EMG cross-talk or to their very similar mechanical actions.
Our normalization procedure in the preceding analysis showed that muscles recruited in a correction response exhibited within-trial coupling. This does not guarantee an invariant gain or coupling ratio of muscle activations between trials. However, we found that the peak amplitude of the non-normalized EMG of recruited muscles covaried between correction trials. This indicated that there was little trial-to-trial variation in the coupling of muscles recruited in a correction response. The peak EMG amplitudes of SA, GL, and RA, the muscles activated during the correction response, are plotted versus one another in three separate panels in Figure 8E. The peak EMG (within a 100 msec time window after obstacle contact; thus the knee extensor burst in RA was not considered in calculation of the peak) of an individual muscle varied from trial to trial. However, the peak EMGs of SA, GL, and RA covaried together and were significantly correlated (p < 0.01;r2 values ranged from 0.80 to 0.90).
In summary, EMG analyses showed that the several muscles that were recruited during the correction response linearly covaried through time and behaved as a regulated group. Thus, in the frogs tested here, muscle groups and not single hindlimb muscles produced the fixed-structure corrective fields.
Correction response amplitude is regulated based on phase and impact force
We examined what factors might regulate the peak amplitude and duration of the correction response from trial to trial. We found that the peak amplitude of the corrective motor response depended on the magnitude of obstacle collision, i.e., the contact force. In Figure9A, rectified and filtered SA EMGs evoked during successive obstacle collision trials are shown plotted on top of one another. Each wiping trial started from the same initial limb position with the path obstacle at the same distance (∼2 mm) from the ankle. Obstacle collision occurred at a similar time for each trial, within 16.67 msec of movement onset, and collisions evoked correction responses at a similar time. There was a 15 msec range of variation in the onset of the initial EMG response. We correlated the peak of the SA EMG evoked by obstacle contact with the highest initial contact force exerted against the obstacle before the onset of the SA EMG. For most frogs (five of six) as shown for the frog in Figure9B, there was a significant (p < 0.05) correlation between initial rise of contact force before the corrective response was evoked and the peak of the ensuing corrective response (here represented by the SA EMG). However, the duration of the corrective motor response showed no correlation with contact force. The total range of variation in the duration of the corrective response was ±5 msec. Variations in corrective response amplitude could be caused by variations in the magnitude of the feedback signal related to strength of impact. However, we expect that contact force in this experiment will vary with the magnitude of the initial knee flexor burst. Fluctuations in the overall strength of the motor program could cause variations in the strength of this burst. If corrective response strength covaried with overall motor program strength, this could therefore also account for the covariation of corrective response amplitude and contact force.

Correction response amplitude was regulated both by motor phase and by feedback arising from obstacle contact.A, Rectified and filtered SA EMGs are overlaid for multiple trials (n = 11) where obstacle collision occurred at a fixed latency. For each trial, obstacle collision occurred within 16.67 msec after movement onset (the mean time of movement onset and the SDs from the mean are shown asbold and dotted vertical lines, respectively). Collision occurred slightly after thesecond dotted line. Evoked responses represent scaled versions of one another. B, The peak magnitude of the evoked SA EMG is plotted versus the highest contact force before SA onset as the limb collided with the obstacle. The peak SA EMG was significantly correlated with the highest preceding contact force (p < 0.05). This relationship could indicate regulation of corrective strength by contact force or covariation of corrective strength with wiping motor pattern strength.C, SA EMG duration at half peak showed no correlation with the magnitude of obstacle collision. D, In a second frog held under isometric conditions, we generated impacts of varying strength using a stepping motor to accelerate a mass into the normal site of obstacle contact. Rectified and filtered peak IL EMG (n = 34 trials) significantly covaried with the applied force (p < 0.01) independently of motor program strength, strength of correction, and time of collision.E, The rectified and filtered EMGs of evoked IL response are shown for multiple trials (n = 12) in which obstacle collision occurred at different distances (and therefore times) along the movement path of the wiping limb. The IL EMG was evoked at progressively longer latencies from movement onset because contact occurred at later times. F, The peak magnitude of the evoked IL EMG is plotted versus the time of obstacle contact. Successful responses (●) were positively correlated with the time of obstacle contact (r2 = 0.73). After RI onset (second solid vertical line), IL responses were no longer evoked. Close to RI onset, some evoked correction responses failed to successfully clear the obstacle (○; see Results). Evoked IL responses had reduced amplitudes in these trials, departing from the linear relationship with impact force. G, The duration of evoked IL responses showed no correlation with the time of obstacle contact and varied by <10 msec. H, The contact force showed a significant linear correlation with the time of obstacle contact (p < 0.001).
In three frogs, we directly tested whether the magnitude of contact force affected the amplitude of the ensuing correction response by varying contact force independently of the motor program. In these frogs, we held the wiping limb immobile by placing bone pins in the distal portion of the tibia and attaching this arrangement to the force transducer (see Materials and Methods). Thus, skin contact did not occur with any restraint device during the wiping reflex, yet the limb was held immobile. To evoke correction responses in this condition, we used a stepper motor to drive a mass into the heel of the wiping limb with a known velocity. We varied the speed of the collision with the limb and measured the force transient produced by the impact. The times at which the mass collided with the limb were randomly varied within a 100 msec time window starting at the onset of the wiping motor pattern. We found a significant correlation between the peak impact force produced by the accelerated mass and the ensuing peak correction response. In Figure 9D, data are shown for one frog (n = 34 trials). Peak impact force varied between 0.5 and 1.0 N, and the peak-evoked IL EMG was significantly (p < 0.01) correlated with this force. The correlation between the peak IL EMG and the time of mass impact was not significant in this experiment. However, we suspect that the correlation between amplitude of the correction response and peak impact force would be higher if the time of mass impact was also taken into account. In the next section we show that motor phase, in addition to impact force, does in fact regulate the amplitude of the correction response.
In five frogs, we systematically varied the position of the path obstacle so that collisions occurred at different times in the motor pattern. Collision times varied from just after movement onset, i.e., near the onset of the knee flexor burst, to just before target limb contact, i.e., near the middle or end of the hip extensor burst and onset of the knee extensor burst. The onset of limb motion was typically 45–65 msec after ST onset, and limb contact occurred ∼200–250 msec after movement onset. We found in each of these frogs that there was a time window (100–150 msec in duration) in which correction responses could be evoked. This window spanned from the time of movement onset up to the time of RI onset, i.e., the hip extensor burst.
Figure 9E shows a set of rectified and filtered IL EMGs evoked in this type of experiment. Correction responses were evoked by placing obstacles at successively further distances from the initial position of the ankle. The evoked corrective responses shown in Figure9E had a duration that varied as little as 10 msec [duration measured at half peak (Fig. 9G)]. Thus, the IL EMGs shown in Figure 9E may represent time-shifted and similar duration (∼120 msec) EMG waveforms. Figure 9Fshows that these IL EMGs were evoked when obstacle contact occurred between the time of movement onset (the first set ofvertical lines denote average and SDs of movement onset) and RI onset (second solid vertical line), a time window of ∼100 msec. However, not all of the evoked correction responses successfully cleared the obstacle. In particular, when obstacle contact occurred close to RI onset, the corrective response often failed (Fig.9F–H, circles). The IL EMG magnitude began to decline in these unsuccessful trials, independently of contact force, which continued to increase (Fig. 9H). Presumably in addition to this decline, the corrective hip flexion was made ineffective by the onset of the relatively powerful hip extensors and increased opposing friction at the obstacle [caused by increased obstacle contact forces (Fig. 9H)]. Once again, the amplitude of successful correction responses was significantly correlated with the peak contact force (Fig. 9F, ●) (p < 0.05,r2 = 0.78). Thus our data suggest that two processes regulate the amplitude of corrective responses. Peripheral feedback signaling the magnitude of contact force appears to scale the amplitude of the corrective response within a well defined time window after movement onset. Second, near the onset of the hip extensor burst, the magnitude of the corrective response is no longer related to contact force. At this point, central processes dominate, causing a rapid decline in amplitude.
In summary, the corrective response is produced by a constant duration (and possibly common) motor drive to multiple motor pools. Both cutaneous feedback and motor phase can regulate the amplitude of the corrective response, but neither affects the duration. The invariance of response duration is explored in more detail in the following section.
Constant timing of corrective response and other EMG elements of the wiping motor pattern
In Figure 9, we showed that corrective responses could be evoked at different phases in the wiping motor program with little variation in the duration of the response. Despite constant duration of the EMG measured at half peak, it remained possible that the shape of EMG waveforms underlying correction responses varied depending on phase and amplitude of response. To examine this, we aligned the peaks of individual corrective EMG responses [shown in Fig.10A (IL EMG)] and normalized their magnitudes at this peak. We then obtained the averaged EMG waveforms and their variances [for example, Fig. 10B (IL waveform)]. We found that when the waveform of activation was scaled in this way in amplitude alone, EMG waveforms of different corrective responses were very similar in shape and displayed low variance. We examined our filter characteristics (phase and frequency response) and averaging procedures, and these cannot account for the low variance of the waveform shape and duration reported here.
We tested whether the conserved shape of the activation waveform was a unique feature of the corrective response. To do this, we applied the same method of time alignment of EMG peaks and amplitude normalization to other muscles' EMGs. We chose muscles that were activated in different phases of the wiping motor pattern and not associated with corrections (for example, RI and ST). We found that these other activation waveforms also exhibited shapes (Fig. 10B,RI and ST) very similar to those observed in correction responses. When the averaged waveforms of different muscles were overlaid (Fig. 10B, last row), it could be seen that they all lay within one SD of the IL corrective waveform. We repeated this analysis procedure in a series of frogs. A second example is shown in Figure10C. Activation waveforms of different muscles in different frogs showed similar shapes and remarkable invariance [Fig. 10D (EMG overlay for frogs bf89 and bf64)]. For certain muscles, [for example in Fig. 10,B (RI) and C (ST)], we were forced to infer inactivation shape because of overlapping phases of activity (for example, ST was commonly activated in both the knee flexor and hip extensor phases of wiping). However, in different frogs, the waveform of these same muscles could be observed directly (Fig. 10C,RI). In general, these data support the idea that different coactivated muscle groups showed very similar patterns of activation through time. We speculate that the conserved activation dynamic described here may be an important feature of the operation of motor primitives recruited in reflex behaviors and pattern generation in the frog.
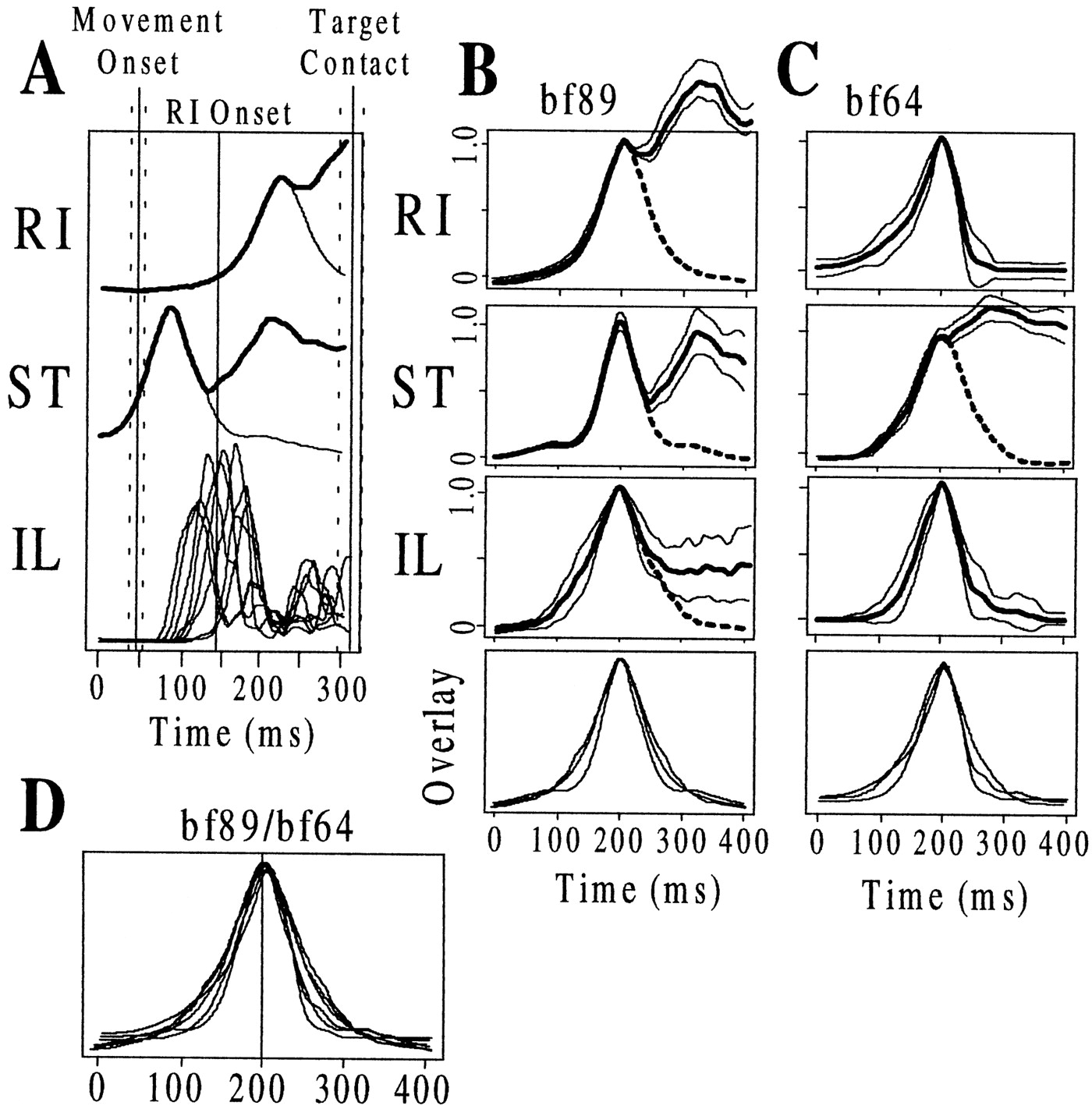
The activation waveform of EMGs comprising the correction response had a shape and duration that were similar to the activation waveforms of other muscles' EMGs in different phases of the wiping motor pattern. A, In the top two rows, the averaged EMG waveforms of ST and RI from movement onset up to target contact are shown. In the bottom row, the individually evoked IL responses are shown (from Fig. 9). IL responses were evoked by obstacle contact that occurred at different times in the interval between movement onset and RI onset. This was a 100 msec time window. The averaged time of movement onset is marked by the first set of vertical lines;dotted lines represent 1 SD about this.B, The averaged EMG waveforms of ST, RI, and the corrective IL burst during trajectory correction trials (n = 11) for the data in A are shown in the top three rows. Averages were constructed by aligning individual EMGs from each trial (n = 11) at its initial peak value (ST and RI exhibit multiple bursts in the wiping motor pattern) and normalizing this peak to a value of 1 (see Results for more detail) before averaging the resultant waveforms. SDs are shown around the averaged EMGs. The dotted linesindicate the inferred relaxation of the activation waveform in the absence of the later, overlapping phases (note however that in frogC, the relaxation of RI can be directly observed). These were obtained by assuming waveform symmetry and reflecting the rising phase of the EMG. In the bottom row, the three averaged EMG waveforms for RI, ST, and IL are overlaid. These waveforms have a similar shape and duration despite occurring at different phases of the motor program, and all lie within 1 SD of the IL waveform.C, The same averaging procedure was performed in a second frog. Individual EMG waveform averages were similar and showed low variance. Overlaid EMG waveforms have a similar shape and duration.D, The ST, RI, SA, and IL EMG waveforms for the two frogs shown in B and C are overlaid. The EMG waveforms in these spinal frogs all exhibit similar shape and duration. The data suggest that different motor primitives may all have common activation dynamics even when recruited in different phases and combinations during reflex behaviors.
DISCUSSION
Overview
Construction of limb movements by summation of force-field primitives was suggested by previous work. This paper examines this mechanism of movement generation and provides the first analysis of trajectory correction responses during hindlimb wiping in spinal frogs.
We showed that cutaneous feedback critical to obstacle detection acted on-line to trigger activation of a correction response during free limb wiping. We exploited this property to test in the immobilized limb whether correction responses were produced by on-line summation of force-field primitives. A force-field primitive was observed here as an isometric force field that exhibited a conserved structure over time (Giszter et al., 1993). We found that in the absence of a correction response, two force-field primitives were sequentially activated during the wiping reflex. When a correction response was triggered during wiping, the pattern of forces produced by the limb could be described most simply by summation of a third, corrective force field with the underlying sequence of wiping force fields. We showed that both impact force and time of impact within the wiping motor program acted to regulate the amplitude of the evoked correction response without affecting response duration. Finally, we found that the activation waveform of the correction response was similar to the activation waveforms of other muscles recruited in different phases of the wiping motor pattern.
Thus, our data are consistent with the view that lower vertebrates construct and adjust certain hindlimb movements by sequencing and summing force-field primitives that each exhibits a common activation waveform. In addition, our data indicate that vector summation, which was demonstrated in dual microstimulation experiments, occurs naturally and describes the effect of combining force-field primitives during real behavior. We discuss these findings in greater detail below.
Spinally organized correction responses
The spinal cord of lower vertebrates can generate targeted scratching or wiping movements to different skin areas (Stein, 1983;Berkinblit et al., 1986; Giszter et al., 1989; Schotland and Rymer, 1993; Sergio and Ostry, 1993). Here we observed correction responses that occurred during wiping in the spinal frog and appeared modular in terms of both muscle activation and force production. The muscle activation pattern was quite simple when compared with corrective patterns observed in the swing phase during mammalian locomotion (Forssberg, 1979; Wand et al., 1980; Drew and Rossignol, 1987;Rossignol, 1996). For instance, in wiping corrections, there was only a single period of muscle activation that summed with and did not disrupt the underlying motor pattern, and there were no noticeable inhibitory or contralateral effects. In the frog, the limbs are mechanically uncoupled from the environment during wiping, there are reduced postural requirements, and the relatively small mass of the hindlimbs may minimize mechanical interactions between the limbs relative to muscle force and stiffness. Response simplicity might also result from phylogenetic differences (Simpson, 1969). Nonetheless, decomposition of correction responses in other animals into a summation of force-field patterns has not been attempted. In the cat, Buford and Smith (1993)suggested that two sequential force patterns first withdraw the hindlimb from the obstacle and secondarily lift it over the obstacle. These force patterns must act in concert with intersegmental and contact forces to produce the kinematics associated with corrections. Finally, prolonged obstacle contact during forelimb correction responses in the cat may be an active information-gathering strategy (Drew and Rossignol, 1987).
Value and limitations of force-field descriptions
Primitives were observed as isometric force fields that exhibited a conserved structure over time. Force fields map the limb's configuration into isometric forces at the ankle. Force fields were measured by holding the limb immobile in some location and recording the isometric forces produced after evoking wiping. This measurement was repeated at each of an array of positions, for the same stimulation. In this way a sampled map of force-position pairs was constructed. This map is a summary of the endpoint effects of the mechanical properties of the activated muscles in isometric conditions, limb configuration, and the effects of neural feedback pathways operating under isometric conditions. The isometric description summarizes the output of the primitive under defined conditions. In part this may help to predict the motion of the leg and its interactions with the environment. However, in free-limb conditions, limb velocity and motion-related feedback must also be considered, position feedback is altered, muscles operate in different ranges of their force-velocity curves, and inertial interactions among limb segments must be accounted for. In the frog, data support an important role of proprioceptive feedback in the organization and control of wiping trajectories (Kargo and Giszter, 2000).
We focused on an isometric description of primitives to test the hypothesis that force-field summation underlies trajectory adjustments. As outlined in the preceding paragraph, the isometric description provides only part of what is needed to fully understand the structure and regulation of field primitives and their superposition in trajectory generation. However, in the behavior observed here, the isometric description was directly relevant to the function of the correction response. During free limb trials, collision with the path obstacle brings the limb to rest and initially prevents further limb movement. Thus, as a result of the collision, the limb enters isometric conditions. The isometric measurement therefore matches the initial mechanical effects of the obstacle and directly represents the forces generated by the frog that serve to accelerate the arrested limb in a new direction to clear the obstacle.
Force-field summation during real behavior
In past work, microstimulation of premotor regions of the spinal cord elicited only a few types of force fields (Bizzi et al., 1991;Giszter et al., 1993; Saltiel et al., 1998). A limited number of primitives reduces the degrees of freedom of the musculoskeletal system. However, this modular framework retains flexibility because the individual force fields can be scaled and coupled in different combinations. Mussa-Ivaldi et al. (1994) showed that force fields could be combined, by dual spinal cord stimulation, to produce a new field that represented the vector sum of the component fields. Simulations showed that this combinatorial mechanism could in principle form a powerful framework that allowed the construction of any smooth force pattern (Mussa-Ivaldi 1992, 1997; Mussa-Ivaldi and Giszter, 1992). Thus, this framework offered a powerful account of movement construction and has even been implemented in the design of robot limb control (Williamson 1996; Mataric et al., 1998). However, up to this time there has been no clear demonstration that force-field combination occurs in behaviors produced by spinal pattern generators, reflexes, or descending systems. This paper provides this demonstration and gives new insight into how primitives may be used to generate and control movement. It must be noted that our demonstration of force-field summation is based in part on a decomposition of the corrective response. However, the amplitude of the corrective field extracted in this way could be modulated independently by regulating impact strength. This supports interpretation of the corrective field as an independent component of behavior.
Combination of primitives does not alter the normal motor program
It was unexpected that despite the addition of the corrective response, the timing of phases comprising the wiping motor pattern was not affected in obstacle collision trials when compared with unperturbed trials. This was true when the motor program was examined at the level of free limb EMGs, isometric EMGs, or force production. Despite the lack of adjustment in the timing of the wiping motor pattern, the time to target limb contact was preserved and similar to the times to contact seen in unimpeded trials. Kargo and Giszter (2000)observed that wiping limb trajectories from many initial positions to the target limb occurred with a constant duration. Martin et al. (1995)also observed in cat reaching that many trajectories occurred with a constant duration. Here, duration was conserved even with a correction. Conserving the duration of targeted multi-limb behaviors might represent a strategy to simplify the task of interlimb coordination. The conserved timing of target limb contact could minimize the need for continuous exchange of coordinating information bilaterally and for modification of contralateral limb kinematics as a result of the collision. Indeed, modifications of responses as a result of obstacle collision were primarily localized to the wiping limb. Modularity and encapsulation of controls at several levels thus appear to be supported by our data.
A unit of fixed temporal dynamics
How force-field primitives are recruited, modulated, and combined to construct movements could occur in several ways. First, a primitive could represent a simple feedforward synergy that passively follows arbitrary activation patterns. The tasks of planning and execution systems then involve determining the activation waveforms and feedback controls (Mussa-Ivaldi and Giszter 1992) impinging on the synergy. Alternatively, a primitive could have an intrinsic dynamics or timing when activated. The tasks of planning and execution systems then involve determining appropriate phasing and amplitude settings for each primitive, which resemble wavelets. The finding that different muscles, recruited in different phases of wiping and in the correction response, exhibited similar activation waveforms supports the second alternative. Wavelet and similar approaches have proven powerful ways to control robots, deal with time delays, and teleoperate robots (Cannon and Slotine, 1995; Sanner and Slotine, 1995; Massaquoi and Slotine, 1996).
Voluntary human movement exhibits a wide temporal range, which may vary depending on task. However, several investigators have provided support for schemes of voluntary trajectory generation involving superposition of pulsed or fixed timing elements (Vicario and Ghez, 1984; Burdet and Milner, 1998; Doeringer and Hogan, 1998; Gottlieb, 1998; Krebs et al., 1999). Pattern-generating systems that recruit motor elements in chains and in combination contingent on feedback and probabilistic or fuzzy logic have also been proposed by Prochazka and Gillard (1997). It remains to be seen whether the timing properties of primitives remarked on here, in wiping and its corrections, are preserved across many behaviors and experimental conditions in spinal frogs.
Footnotes
This work was supported by National Institutes of Health (NIH) Grant NS43460 to S.F.G., and NIH Grant 5T32 HD07467. We thank Drs. Donald Faber, Marion Murray, Alberto Pereda, Richard Poppele, Terence Sanger, and John Soechting for comments on earlier versions of this work. Nicholas Auyong designed the driver circuit for the stepper motor system used in part of this work.
Correspondence should be addressed to Simon Giszter, Department of Neurobiology and Anatomy, Medical College of Pennsylvania/Hahnemann Medical School, 3200 Henry Avenue, Philadelphia, PA 19129. E-mail:simon{at}swampthing.auhs.edu.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}