

Abstract
The proper guidance of migrating growth cones relies on the balance of multiple guidance cues in the embryonic environment. In addition to guidance cues, growth cones are in contact with other substrates that may contribute to the pathfinding of neurons. For example, in the developing insect peripheral nervous system, pioneer neurons migrate on and between layers of the basal lamina. Previous studies have demonstrated that one basal lamina molecule, laminin, promotes outgrowth of many classes of neurons in vitro. In this study, the simple grasshopper nervous system was used to investigate the role of laminin in neuronal pathfinding. Laminin expression precedes axonogenesis of the Tibial (Ti1) pioneer neurons in the developing limb bud, and expression continues during outgrowth and guidance of the pioneer neurons. The role of a nidogen-binding motif on laminin was investigated using subunit-specific antibodies and peptides as blocking reagents in vivo. Antibodies and peptides that block the nidogen-binding site on laminin resulted in stalled Ti1 axon migration, predominantly at the precise location where they normally turn ventrally. After prolonged culturing, Ti1 axons remained stalled at the same location. Therefore, although Ti1 axons were capable of outgrowth in the presence of blocking reagents, they were not able to navigate an essential turn. This study indicates that the interaction of the Ti1 growth cone with the nidogen-binding site on laminin is vital for neuronal pathfinding in vivo and suggests that permissive cues may be essential for growth cone steering.
A growing axon encounters many cues in the embryonic environment that are processed by the growth cone and result in directed outgrowth. These cues can be considered instructive or permissive. Instructive cues generally have a restricted expression pattern and guide neurons by conferring either attractive or inhibitory information on the growth cone, leading to reorganization of the cytoarchitecture that culminates in a steering event. Growth cones may also encounter permissive cues as they steer in response to instructive cues. However, it is unclear whether permissive cues aid in the decision making of growth cones. Recent work has indicated that the stimulus history of a neuron affects responsiveness to upcoming cues (Shirasaki et al., 1998; Matise et al., 1999; Diefenbach et al., 2000). This could be one mechanism by which permissive cues act on migrating neurons; however, this has not been demonstrated in vivo.
A variety of permissive cues can be found in the basal lamina, which is a complex array of many different proteins, including laminin, type IV collagen, nidogen, and perlecan (Timpl and Brown, 1996). Laminin is a major component of the basal lamina and consists of three subunits, α, β, and γ. The three subunits are secreted as a trimer and can self-assemble to form polymeric sheets (Timpl and Brown, 1996). Genetic analysis of laminin function reveals that laminin is required for basal lamina stability (Noakes et al., 1995; Garcia-Alonso et al., 1996; Miner and Li, 2000). Therefore, testing laminin function in axon guidance in a genetic system is problematic, considering its widespread involvement in developmental events. Because of these experimental constraints, the role of laminin in directing neurons during development in vivo has been difficult to address.
Several studies have implicated laminin as a potent promoter of neurite outgrowth in vitro (for review, see Powell and Kleinman, 1997), as a directional cue for migrating axons in vitro (Kuhn et al., 1995; Halfter, 1996; Kuhn et al., 1998), and as an important guidance molecule for developing axons in vivo (Garcia-Alonso et al., 1996; Forrester and Garriga, 1997). One class of laminin receptors, the integrins, are critical in mediating laminin-induced neurite outgrowth (for review, see Powell and Kleinman, 1997) and are modulated depending on laminin availability (Condic and Letourneau, 1997), laminin conformation (Calof et al., 1994; Ivins et al., 1998), and developmental age (Cohen et al., 1986,1989; Hall et al., 1987; Ivins et al., 2000). This suggests dynamic interplay between laminin and integrins on the neuronal cell surface.
In this study, we examine the role of the permissive cue laminin in axon guidance. Sequence analysis of the grasshopper laminin γ-chain demonstrates a single conserved nidogen-binding site that has been shown to be important for epithelial tissue morphogenesis in other systems (Gerl et al., 1991; Mayer et al., 1993a; Ekblom et al., 1994; Poschl et al., 1996; Kadoya et al., 1997). We show that this nidogen-binding site is important for axonal pathfinding and may be required for growth cone turning in vivo.
MATERIALS AND METHODS
Antibody staining. Embryos were dissected out of their egg cases in saline, and the amnion was removed and staged according to the method of Bentley et al. (1979). Embryos were fixed for 1 hr in 3.7% formaldehyde in PIPES, EGTA, and MgSO4. Embryos were blocked for 1 hr at 4°C in either PBT and 5% normal goat serum or PBT and 5% normal donkey serum, depending of the host of the secondary antibody. Primary antibodies (see below) were incubated overnight at 4°C, followed by several washes in PBS supplemented with 0.1% bovine serum albumin and 0.1% Triton X-100, pH 7.2 (PBT) and secondary antibody incubation at 1:250 in PBT for 1 hr at room temperature. Embryos were again washed in PBT and mounted in Slowfade antifade (Molecular Probes, Eugene, OR). Primary antibody concentrations were as follows: goat anti-HRP, 1:500; rabbit anti-HRP, 1:500; rabbit anti-laminin γ-chain, 1:500; rabbit anti-semaphorin 2a, 1:250; mouse anti-semaphorin 1a, 1:1; rabbit anti-β-integrin (against the full-length protein), 1:20; and rabbit anti-β integrin (against the intracellular domain), 1:50. Rabbit anti-β-integrin antibodies were courtesy of Salvatore Carbonetto (McGill University, Montréal, Québec, Canada). Goat anti-HRP and rabbit anti-HRP were from Jackson ImmunoResearch (West Grove, PA), as were the secondary antibodies used in this study (FITC-conjugated donkey anti-goat, FITC-conjugated donkey anti-rabbit, Cy3-conjugated goat anti-mouse, and FITC-conjugated goat anti-rabbit). For double labeling, primary antibodies were incubated together with embryos overnight at 4°C. Secondary antibodies were also incubated together for 1 hr at room temperature. For integrin immunofluorescence, embryos were immobilized on glass coverslips previously coated in 5 mg/ml poly-l-lysine and filleted along the proximal–distal axis of the limb (O'Connor et al., 1990). Filleted limbs were rolled open to expose the ventral epithelium, containing the Tibial (Ti1) neurons. Embryos were fixed and stained with anti-β1-integrin antibodies.
Confocal microscopy. Confocal immunofluorescent images were collected on a Nikon Optiphot-2 microscope using an MRC 600 confocal system (Bio-Rad, Hercules, CA) equipped with a krypton–argon laser. The images collected from the confocal microscope were captured in a 768 × 512 pixel field of view with the optical sections collected at 0.8 mm intervals. The confocal images were composed of 100–150 optical sections for each embryo. Data collected from the confocal microscope were analyzed in NIH Image 1.61, and Adobe (Mountain View, CA) Photoshop 4.0 was used for presentation. Confocal microscopy was conducted at the Electron Microscopy facility at the University of British Columbia.
IgG purification. The IgG fraction of immune and preimmune sera was isolated using an Immunopure protein A IgG orientation kit (Pierce, Rockford, IL). Sera were loaded onto columns and the columns were washed with 4 × 5 ml 1 m NaCl. The IgG fraction was eluted with 0.1 m glycine, pH 2, and neutralized with 100 μl of 1 m Tris, pH 7.5. Absorbance at 280 nm was taken, and the concentration was determined using the equation 1 OD = 0.75 mg/ml protein.
Dialysis. Before culturing, preimmune and immune IgG fractions were dialyzed against sterile RPMI medium overnight at 4°C. The IgG fractions were placed in 6–8 kDa dialysis tubing, which was placed in 500 ml of sterile RPMI medium. After overnight incubation, the medium was refreshed once, and dialysis continued for another 5 hr.
Western blot analysis of fusion proteins. Fifty nanograms of purified fusion protein was electrophoresed at 200 V in a 7.5% SDS-PAGE gel (4% stacking gel) and electrotransferred to Hybond enhanced chemiluminescence (ECL) nitrocellulose (Amersham Pharmacia Biotech, Arlington Heights, IL). Transfer buffer consisted of 25 mm Tris, 192 mm glycine, and 20% methanol. The blots were blocked in 5% milk powder, PBS, and 0.1% Tween 20 overnight at 4°C. Primary antibody was used at a dilution of 1:10,000 for β and γ antisera and β and γ preimmune sera in 1% milk powder, PBS, and 0.1% Tween 20. Primary antibody incubations were conducted at room temperature for 2 hr, followed by washes in PBS and 0.1% Tween 20. Secondary antibodies (HRP-conjugated goat anti-rabbit) were diluted to 1:1500 in PBS and 0.1% Tween 20 and incubated for 1 hr at room temperature. After washes in PBS and 0.1% Tween 20, blots were reacted in ECL detection buffer (Amersham Pharmacia Biotech), and exposed to X-OMAT x-ray film (Eastman Kodak Co., Rochester, NY).
Western blot analysis of embryonic lysate. Approximately 50 grasshoppers at 35–40% of development were dissected from their egg cases in saline and resuspended in 1× radioimmunoprecipitation assay buffer (with 85 μg/ml PMSF, 0.5 μg/ml aprotinin, and 1 μg/ml leupeptin). Embryos were centrifuged at 14,000 rpm at 4°C for 20 min, and supernatant was retained. Western blot analysis was performed as with fusion protein with the following exceptions: embryonic lysates were electrophoresed on a 6% SDS-PAGE gel (4% stacking gel) overnight at 20 V at 4°C; and the gel was electrotransferred to nitrocellulose in 50 mm Tris, 380 mm glycine, and 0.1% SDS at 30 V overnight at 4°C. The rest of the Western blot analysis was performed as in the protocol for the fusion protein Western blot analysis.
Culturing. Eggs were sterilized in 70% ethanol and rinsed twice in sterile grasshopper culture media (RPMI medium, 4 μm 20-hydroxyecdysone, 0.4 mmCaCl2, 0.4 mmMgSO4, 2 U/ml insulin, 100 U/ml penicillin, 100 μg/ml streptomycin, 10% heat-inactivated FBS, 2 mml-glutamine, 0.45 mm sodium pyruvate, 1 mm oxaloacetic acid, 0.45% d-glucose, and 0.12m sucrose, pH 6.9). Embryos were dissected as before (see Antibody staining) in grasshopper culture media. Embryos were cultured in the presence of blocking peptides or antibodies. For longer cultures, embryos were prepared as described for culturing but with the following modifications: after 24 hr in culture, the medium was removed, and fresh medium was added, with the appropriate blocking reagents.
Blocking peptides. The DPNAV peptide was designed on the basis of grasshopper laminin deduced amino acid sequences. As a control, randomly scrambled peptides were used (PANDV for DPNAV). All peptides were generated at the Protein Services Unit at the University of British Columbia and dissolved in sterile H2O. DPNAV and PANDV were used at a final concentration of 0.2 mg/ml.
Blocking antibodies. A 561 bp EcoRI fragment of domain III of the γ chain of laminin was subcloned intoEcoRI-cut pGEX 2T (Amersham Pharmacia Biotech). pGEX-specific primers were used to sequence clones to confirm cloning junctions with respect to orientation and frame. GlutathioneS-transferase fusion proteins were purified with glutathione-linked agarose beads (Sigma, St. Louis, MO) according to the method of Smith and Johnson (1988).
Rabbits were initially injected with 0.5 mg of fusion protein in 500 μl, which was emulsified with an equal volume of Freund's complete adjuvant (Sigma). For all subsequent injections, 0.1 mg of protein in 500 μl was emulsified with an equal volume of Freund's incomplete adjuvant (Sigma). Rabbits were housed and cared for at the University of British Columbia Animal Care Center. All bleeds were processed according to the method of Harlow and Lane (1988).
All antibodies, including preimmune controls, were prepared as follows: IgG was isolated from sera and dialyzed against sterile RPMI; γ antisera and preimmune sera were used at 1 μm, as determined by an A280spectrophotometric reading.
Assessment of basal lamina integrity in blocked embryos.Embryos were cultured in the presence of the DPNAV peptide for 24 hr as described. After the culture period, embryos were fixed and stained for HRP and semaphorin 2a immunoreactivity and HRP and laminin γ antisera immunoreactivity. Embryos were processed for confocal microscopy as described above.
RESULTS
Laminin expression is evenly distributed during development of the Ti1 pathway
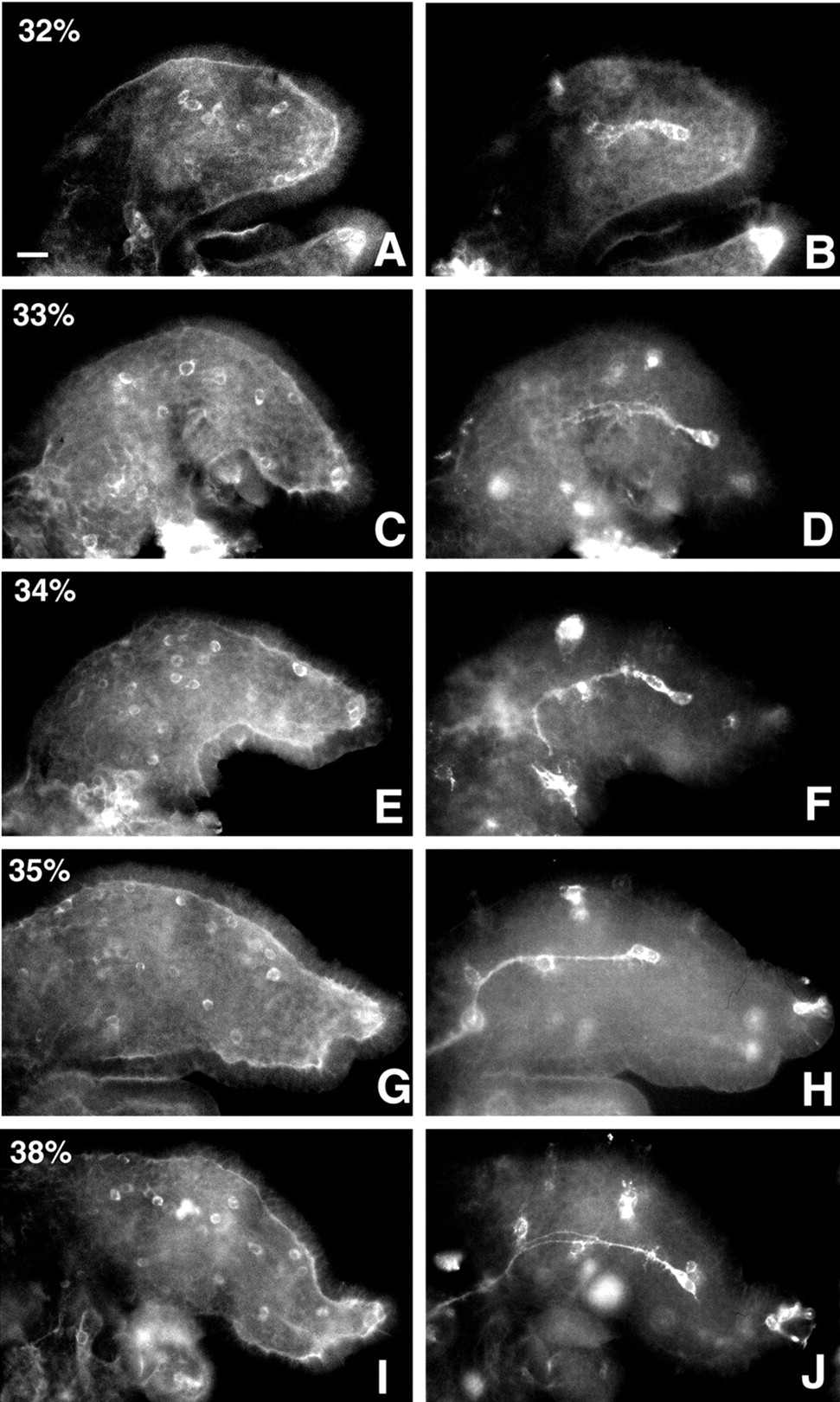
In the developing limb bud, the Ti1 pioneer sensory neuron projection establishes a projection to the CNS by 35% of embryonic development. The pathway is stereotyped and consists of a series of sequential steering decisions (Keshishian and Bentley, 1983). At 30% of embryonic development, the newly formed Ti1 neurons delaminate from the epithelium and extend axons proximally toward the CNS (Fig.1A). After the initial proximal extension of the Ti1 axons, the Ti1 growth cones contact a preaxonogenesis neuron, the Fe1 (femur 1) cell, at ∼32% and continue proximal growth (Fig.2B). Subsequently, at 33% of development, the Ti1 axons contact a second preaxonogenesis cell, the Tr1 (trochanter 1) cell (Fig. 2D). On contact with this cell, the Ti1 axons make a ventral turn along the trochanter epithelium (Fig. 2F). After migrating ventrally in the trochanter epithelium, the growth cones contact another intermediate target, the Cx1 (Coxa-1) neurons, and turn proximally, extending into the CNS (Fig. 2H;Keshishian and Bentley, 1983). Later in development, the Ti1 pathway prefigures the migratory paths of later-arising neurons. As the Ti1 growth cones migrate, they make contact with several substrates, including the laminin-rich basal lamina (Anderson and Tucker, 1989), epithelium, and several intermediate neuronal targets (Keshishian and Bentley, 1983; O'Connor, 1999).

Laminin is expressed by hemocytes at 30 and 35% of embryonic development in the grasshopper limb bud.A, B, Immunofluorescent images of grasshopper limb buds at 30% (A) and 35% (B) of development that are double-labeled with laminin γ antisera (green) and HRP (red).A, Ti1 neurons emerge from the surrounding epithelium at 30% of development (arrow) in a basal lamina (BL) that is rich in laminin. Laminin-expressing hemocytes are visible in the limb (arrowheads).B, Confocal image of a limb bud at 35% of embryonic development. The Ti1 pathway (red) has been established, and arrows point to the Ti1 cell bodies, which have separated slightly in this limb. Laminin immunoreactivity in the basal lamina is even, and laminin-expressing hemocytes (arrowheads) are abundant. C, Western blot analysis of laminin β and γ glutathioneS-transferase fusion proteins with β and γ antibodies. Laminin γ antiserum is specific for the γ fusion protein but does not recognize the β-chain fusion protein, whereas β-chain antibodies cross-react with γ-chain fusion protein (asterisk). D, Western blot analysis of lysate from embryos at 40% of development with β and γ antibodies and preimmune sera (PI). Both β and γ antisera recognize a high molecular weight complex (polymerized laminin) in the stacking gel. The β and γ antibodies recognize a similarly sized band. Preimmune serum reacts with no bands. Scale bar, 10 μm.

Time course of laminin expression in relation to the Ti1 pioneer neuron pathway. Immunofluorescence of laminin (left) and HRP (right) is shown. From 32 to 38% of development, laminin is expressed evenly in the basal lamina during the time that the Ti1 pathway is established. Scale bar, 20 μm.
To determine the specificity of the laminin γ-chain antibodies, Western blots were conducted. Sera raised against epidermal growth factor (EGF) repeats of the γ-chain of laminin reacted with γ fusion protein (Fig. 1C) but not with a fusion protein of EGF repeats from the β-chain (Fig. 1C). On grasshopper embryonic lysates, both the γ antisera and the sera raised against the EGF repeats of the β-chain recognize the same high molecular weight band (Fig. 1D) that represents the laminin heterotrimer. The β and γ antibodies also react with purified laminin (see Materials and Methods) on a Western blot (data not shown), and these bands were of a size comparable with that of the grasshopper laminin bands.
The antibodies generated against the laminin chains were used to determine laminin distribution during Ti1 migration, from 30 to 35% of embryonic development. At 30% of development, Ti1 neurons have delaminated from the epithelium, and an even expression of laminin in the basal lamina is already established (Fig. 1A). In certain preparations, laminin appeared concentrated in the distal tip of the limb, for example, in Figure 2A,G,I. However, Ti1 growth cones appeared to migrate only through regions of uniform laminin distribution as they extended toward the CNS (Fig. 2). Throughout the duration of Ti1 migration, from 30 to 35% of development, laminin is evenly distributed in the basal lamina (Figs.1A,B, 2A–H). At 38% of development, later-arising “follower” neurons begin to migrate toward the Ti1 pathway. At this time in development, laminin is also evenly expressed, (Fig. 2I,J), suggesting a similar role of laminin in guidance of pioneer neurons as well as follower neurons. In addition to being found in the basal lamina, laminin protein (as well as mRNA; data not shown) is also detected in migrating hemocytes (Figs. 1A,B, 2). Laminin deposition in the basal lamina is achieved by high expression of laminin in randomly migrating hemocytes, which secrete laminin as they migrate throughout the limb (our unpublished results).
Localization of laminin receptors on Ti1 growth cones and axons
Laminin is a major component of the basal lamina that is expressed dynamically during development, as early as the 2- to 4-cell stage in mouse, whereas nidogen is not detected until the 8- to 16-cell stage (Dziadek and Timpl, 1985). Laminin has diverse roles in development; for example, it is required for the proper development of many organs, including, muscle, heart, and kidney, as well as the nervous system, such as structuring the synapse (Noakes et al., 1995; Yarnitzky and Volk, 1995; Patton et al., 1998; Sanes et al., 1998; Allamand and Campbell, 2000; Miner and Li, 2000). How can laminin be crucial for so many diverse developmental events? One way is through spatial and temporal regulation of expression of different isoforms of laminin, as observed in muscle (Miner et al., 1997; Patton et al., 1997). Another way is through expression of different receptors at the cell surface. Laminin has been shown to have many receptors, including several classes of integrins, the LAR family of receptor-tyrosine phosphatases, and α-dystroglycan (Powell and Kleinman, 1997; O'Grady et al., 1998;Montanaro et al., 1999). Integrins have been found to mediate laminin-dependent neurite outgrowth (Tomaselli et al., 1990; for review, see Powell and Kleinman, 1997). To investigate potential receptors that may be mediating the effects of laminin at the growth cone, immunolocalization of integrin receptors was conducted. On whole embryos, polyclonal antisera generated from the entire β1 subunit of integrin, as well as another antibody that recognizes the intracellular domain, stained epithelial cell boundaries (data not shown). Because this staining was quite intense, Ti1 neurons could not be visualized. Therefore, limbs were filleted open and immobilized on poly-l-lysine-coated coverslips and double-labeled with HRP and β1-integrin immunofluorescence (Fig.3A,B) and polyclonal antibodies to the intracellular domain of β1-integrin (Fig.3C,D). In this preparation, β1-integrin was found on Ti1 cell bodies, axons, and growth cones at 32–35% of development (Fig.3B,C), during the time when Ti1 axons are navigating to the CNS. Ti1 neurons labeled with anti-β1 antibodies alone showed the same pattern of labeling as neurons that were double-labeled with anti-HRP (Fig. 3, compare A and B withC). In addition to the neuronal labeling, integrin was expressed at epithelial cell boundaries as well as on mesodermal cells (Fig. 3C,D). Expression of β1-integrin on Ti1 growth cones (Fig. 3A,B) suggests that integrins could be transducing laminin signals from the basal lamina.

β1-Integrin is expressed in the developing limb bud. A, B, Ti1 neurons at 32% of development are labeled with anti-HRP antibodies (A) and β1-integrin (B). The arrowindicates a growth cone that is depicted at higher magnification inA′. B, β1-Integrin immunofluorescence of the same neuron in A. β1-Integrin localizes to the cell bodies, axons, and growth cones of the Ti1 neurons. β1-Integrin is also expressed by epithelium and mesoderm, which accounts for the out-of-focus staining. The arrow in Bindicates the growth cone that is magnified in B′.B′, β1-Integrin is expressed on the growth cones of Ti1 sensory axons. C, β1-Integrin staining is confirmed with a different antibody. In an embryo at 34% of development labeled only with β1-integrin antibodies, β1-integrin is expressed in Ti1 cell bodies (asterisks) and axons. Growth cones are out of the field of view. Ti1 axons migrate along an epithelium that expresses β1-integrin at cell junctions.D, Mesodermal cells, possibly hemocytes, express high levels of β1-integrin. Shown is the same embryo depicted inC but at a slightly different focal plane to emphasize mesodermal staining. Ti1 cell bodies (out of focus) are indicated byasterisks. Scale bars, 10 μm.
Blocking a conserved nidogen recognition sequence on laminin disrupts axon guidance
Laminin is found along the entire course of Ti1 axon trajectory (Fig. 2), suggesting that laminin is conferring information to the Ti1 growth cones throughout their migration. Confocal analysis of laminin immunofluorescence in the developing limb bud basal lamina demonstrates that the distribution of laminin is uniform (Fig.1B), suggesting that laminin may play a permissive role in axon guidance. EGF repeat-containing domain III of grasshopper laminin γ contains a completely conserved nidogen-binding site (Gerl et al., 1991; Mayer et al., 1993a; Poschl et al., 1996; our unpublished results). Antibodies generated against this region have shown that this site is important for epithelial morphogenesis (Ekblom et al., 1994;Kadoya et al., 1997), and nidogen has recently been shown to direct migrating neurons (Kim and Wadsworth, 2000). Previous studies have demonstrated that both antibodies and peptides directed at this site interfere with nidogen binding to laminin (Mayer et al., 1993a). With this in mind, antibodies and peptides were designed to target this site, to disrupt laminin–nidogen interactions, and to determine their role in axon guidance. Grasshopper embryos were cultured in the presence of these reagents at 30% of embryonic development, and the effect of Ti1 pathfinding was studied.
Embryos at 30% of development were cultured for 24 hr at 30°C in the presence of 1 μm anti-γ IgG antibodies and the synthetic peptide DPNAV (0.2 mg/ml). As controls, IgG-isolated preimmune sera and the randomly scrambled peptide PANDV at the same concentrations as γ antisera and DPNAV, respectively, were used. Ti1 neurons exhibited three types of pathfinding errors. The most prominent error, stalled axons, is characterized by the failure of both Ti1 growth cones to initiate the ventral turn at the trochanter segment and to extend to the CNS (Fig. 4, compareA and B). One or both of the Ti1 axons extending in the dorsal epithelium of the limb compartment typify the second type of abnormality, a dorsal projection. Finally, the third abnormality is characterized by a distal projection of one or both Ti1 axons from the cell bodies instead of extending proximally. However, a small number of dorsal and distal errors were also found in control embryos. A comparison of the dorsal and distal error rate in control cultures with experimental cultures revealed that there was no statistical difference between control and treated embryos. Importantly, for both blocking reagents, similar abnormalities were observed, suggesting that both reagents were acting by the same mechanism, probably as competitive inhibitors.
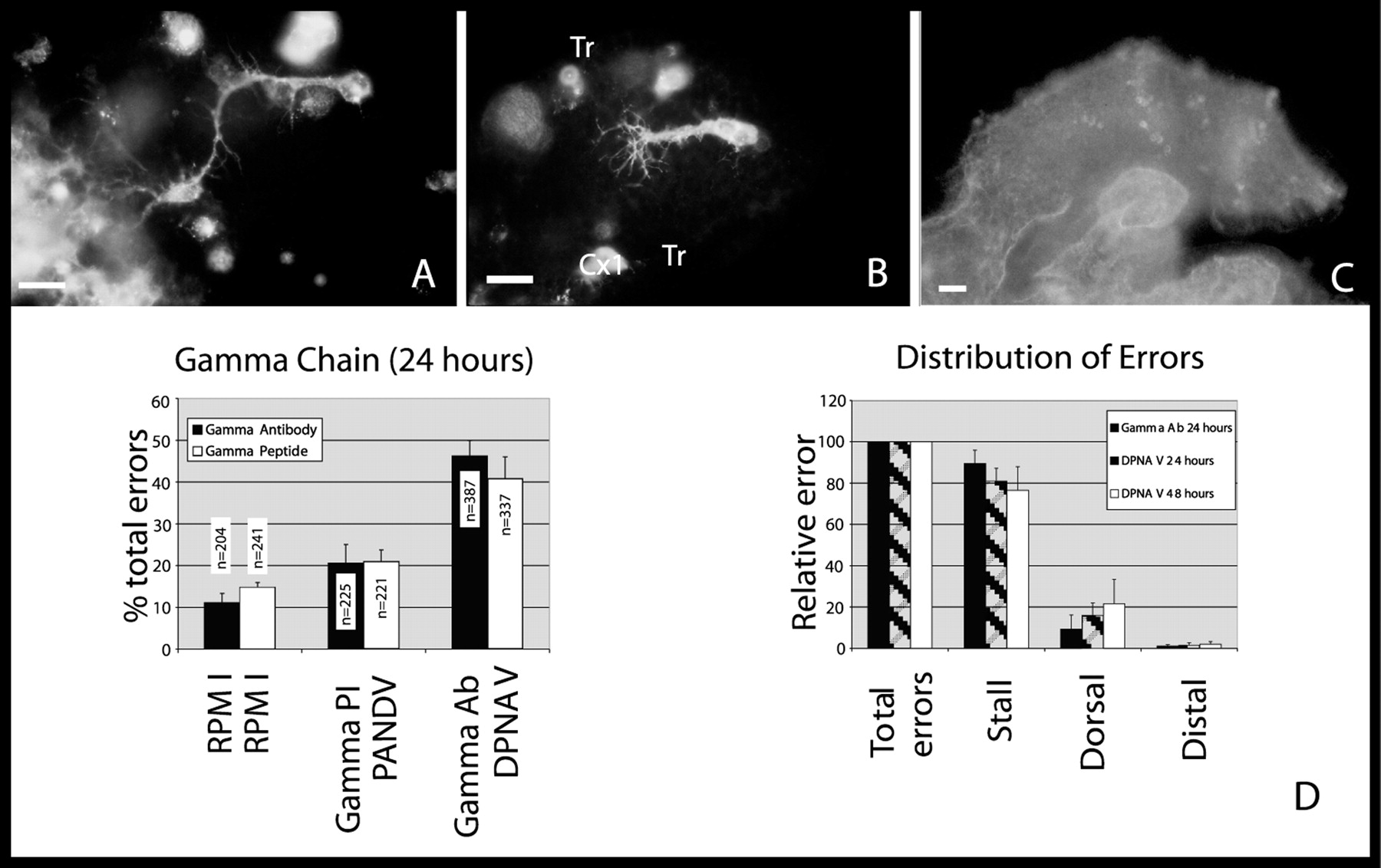
Disruption of laminin–nidogen interactions results in aberrant neuronal pathfinding.A, Control culture demonstrating the Ti1 pathway as visualized with HRP immunofluorescence. Culturing in preimmune sera (PI) or randomly scrambled peptides (see Results) has no effect on Ti1 pathfinding. B, Stalled Ti1 axons when laminin–nidogen is blocked. Ti1 neurons havebrightly labeled cell bodies, axons, and extended growth cones, indicative of healthy neurons. C, Embryo after culturing with laminin γ antisera with the addition of anti-rabbit secondary antibody demonstrating that the antiserum has penetrated the limb bud and is localized, binding to laminin epitopes found in the basal lamina and on hemocytes. Antibodies have access to the lumen of the limb bud during the culture period. D, Antibodies and peptides to the nidogen-binding site on laminin disrupts neuronal pathfinding. In 24 hr cultures, both antibodies to the nidogen-binding site (Gamma Ab) and peptides (DPNAV) result in an increase in total Ti1 pathfinding errors (first graph). The total Ti1 pathfinding error was normalized to 100%, and the total errors were broken down into categories (see Results). In both 24 and 48 hr cultures, most errors were stalled axons (second graph). Scale bars: A, B, 20 μm;C, 30 μm.
The rate of disrupted Ti1 pathways when cultured with 1 μm γ antibody was significantly higher than controls, at rates of 46 ± 3.7% (n = 387, six independent trials; Fig. 4D), compared with 11.1 ± 2.2% in RPMI medium and 20.6 ± 4.5% with preimmune sera;p = 5.2 × 10−6 for RPMI versus γ antibody; p = 0.0007 for preimmune versus γ antibody (unpaired Student's t test). Similarly, 41 ± 5.2% of embryos that were cultured in the presence of 0.2 mg/ml DPNAV (n = 337, six independent trials; Fig.4D) exhibited disrupted Ti1 pathways compared with 14.8 ± 1.1% for RPMI medium and 21 ± 2.8% for control peptide; p = 0.00031 for RPMI versus DPNAV;p = 0.004 for control peptide versus DPNAV. The most frequent error in the Ti1 pathway after laminin–nidogen block was stalled axons, (Fig. 4, compare A and B). The integrity of the growth cones and the Ti1 cell bodies suggested that cell death was not a factor in these experiments (Fig.4B). In antibody- and peptide-blocked embryos, 89 ± 6.4% (n = 181) and 80 ± 6.2% (n = 131) of total errors were of the stalled variety (Fig. 4D). The other errors were dorsal projections (at rates of 9.4 ± 6.7 and 16 ± 5.9% for antibody and peptide, respectively) and distal projections (at rates of 1.1 ± 0.67 and 1.5 ± 1.3% for antibody and peptide, respectively).
A salient feature of the stalled axons is the location within the Ti1 pathway in which Ti1 axons stall. Stalled Ti1 axons were never observed proximal to the ventral turn, for example, never after committing to the ventral turn, or in the vicinity of the Cx1 cells (Fig.4B). Therefore, it appeared that Ti1 axons either stalled before the ventral turn or completed the pathway. Several mechanisms could account for the cessation of growth and the inability of Ti1 axons to complete their trajectory. One explanation is the possibility that these neurons are growing slower than in control cultures. Previous work has shown that the growth rate of neuronsin vitro is substrate-dependent. For example, chick dorsal root ganglion neurons extend two times faster on laminin than on fibronectin (Kuhn et al., 1995). Because Ti1 growth cones intimately associate with the basal lamina in the developing limb bud (Anderson and Tucker, 1988), it is possible that Ti1 growth cones exhibit substantially retarded growth as a result of the disruption of laminin–nidogen interactions. This would also account for the consistency of the location of the stall, because embryos are cultured for 22–24 hr.
To address whether Ti1 axons exhibit slower growth rates when laminin–nidogen is blocked, two sets of experiments were conducted. First, the growth rate of laminin-blocked Ti1 axons was examined as they extended toward the trochanter. Embryos were cultured in the presence of blocking peptides at 31% of development for 12 hr, and axons were scored according to their location in the limb bud. In embryos cultured for 12 hr in the presence of 0.2 mg/ml DPNAV, 13.6 ± 7.1% (n = 118) of growing axons reached the femur limb segment, compared with 19.8 ± 1% (n = 81) and 9.7 ± 5.1% (n = 74) for PANDV and RPMI, respectively (Fig.5A); 39.1 ± 4.2% of DPNAV-blocked axons reached the trochanter (but did not turn), compared with 40.2 ± 9.5% and 30.6 ± 3.4 in PANDV and RPMI medium, respectively (Fig. 5A). Importantly, there was no significant difference between controls and DPNAV-blocked embryos in these regions of the limb (Fig. 5A), indicating that laminin–nidogen-blocked axons reach the trochanter at the same time as control cultures and have access to the same developmental cues. However, when axons were examined after the ventral turn [Fig.5A, Tr (turn)], there was a slight, significant increase in numbers of axons that had turned in control versus peptide-blocked cultures (p = 0.04 for RPMI vs DPNAV), which is consistent with the stall phenotype.

Growth rate of Ti1 axons is unaffected in laminin–nidogen-blocked embryos. A, The growth rate of Ti1 axons before the trochanter was determined by culturing embryos for a shortened period of 12 hr and the location of growth cones was determined. At the femur (Fe) and trochanter (Tr) there is no difference between laminin–nidogen-blocked embryos (DPNAV) compared with peptide control (PANDV) and RPMI medium. However, a significantly higher number of control growth cones are found after the ventral turn in the trochanter [Tr (turn)] compared with control peptide and laminin–nidogen-blocked embryos. B, Growth rate of Ti1 axons after the trochanter was assessed by extending the culture period to 48 hr. After 48 hr, the Ti1 axons exhibit the same frequency of pathfinding errors, similar to 24 hr cultures (compare with Fig.4D).
To address whether Ti1 axons required additional time to make the ventral turn and grow down the trochanter, embryos were cultured in the presence of DPNAV blocking peptide for an additional 24 hr (total culture time, 48 hr). When cultured for 48 hr, a similar number of aberrant projections were observed; 36 ± 0.75% of axons were abnormal (n = 141; p = 0.0021 for RPMI vs DPNAV peptide; p = 3.6 × 10−5 for control peptide vs DPNAV; Fig.5B). The distribution of errors was not significantly different: 76.5 ± 11.4% stalled; 21.6 ± 11.7% extended aberrantly dorsally; and 2 ± 1.4% grew distally (n = 51; Fig. 4D). Therefore, despite the extended culture period, the Ti1 growth cones ceased growth at the same location within the limb at the trochanter limb segment epithelium. These results suggest that the stalled phenotype is not attributable to a slower rate of growth along the limb epithelium but is attributable to an inability to turn at the trochanter.
Ti1 axons stall within the filopodial range of guidance cues found within the trochanter
The proximal extension of Ti1 axons is followed by a ventral turn in the trochanter limb segment. The trochanter epithelium contains pertinent guidance information that may be important for directing a correct ventral turn, for example, members of the semaphorin family (Isbister et al., 1999). Although semaphorin 1a is expressed in a circumferential band of epithelium in the trochanter limb segment, semaphorin 2a exhibits a dorsal-to-ventral graded distribution in this region (Kolodkin et al., 1992; Isbister et al., 1999). If Ti1 axons stall before the trochanter epithelium, then they may not be within filopodial range of important guidance information. If the growth cones stall within the trochanter, this would suggest that Ti1 growth cones have access to semaphorins but may not be able to respond to them. To establish whether Ti1 axons are within the range of cues found in the trochanter, the location of stalled axons was assessed using two criteria. First, semaphorin 1a immunofluorescence was used as a marker of the trochanter limb segment. Second, the location of stalled Ti1 axons in relation to a pair of preaxonogenesis neurons, the Cx1 cells, was used. These cells lie just proximal to the trochanter limb segment in the distal compartment of the coxa segment (Fig.6A). The neuronal marker used in all blocking experiments, anti-HRP, also labels these preaxonogenesis neurons. Embryos were cultured overnight in the presence of DPNAV blocking peptide, fixed, and stained for both HRP and semaphorin 1a immunoreactivity. Stalled Ti1 axons were scored in relation to semaphorin 1a-expressing epithelium (Kolodkin et al., 1992) or in relation to the Cx1 cells (Fig. 6A). Ti1 axons were scored as stalling before the trochanter (Fig.6A), within the trochanter (Fig.6B), or after the trochanter but not turning (Fig.6C). Using these criteria, 73% of stalled Ti1 growth cones resided within the trochanter (Fig. 6D), whereas 17 and 8.7% stalled before and after the trochanter, respectively (n = 23; Fig. 6E). Interestingly, the small number of axons that were found after the trochanter had not initiated a ventral turn; instead they had continued extending proximally. These results would suggest that the majority of stalled Ti1 growth cones (81.7%, those that stalled in the trochanter and those that stalled past the trochanter) had access to guidance information within the trochanter, such as semaphorins, but were unable to respond to it.

Ti1 axons stall in a discrete location in the limb bud when laminin–nidogen is blocked. A–C, Schematic depicting the three possible locations of stalled Ti1 axons, as assessed with HRP and semaphorin immunofluorescence. D, Laminin-and nidogen-blocked embryo that has been labeled with HRP to visualize neurons; arrowheads indicate trochanter.E, The location of 23 stalled Ti1 growth cones was determined; 73% of stalled Ti1 axons resided within the trochanter (Tr), site of the ventral turn. Scale bar, 30 μm.
Basal lamina integrity
The complex arrangement of the basal lamina results from self-assembly of laminin and type IV collagen as well as from interactions between these networks and other basal lamina constituents (Yurchenco and Furthmayr, 1984; Yurchenco et al., 1985; Timpl and Brown, 1996; Cheng et al., 1997). Genetic knock-out of basal lamina molecules often results in a disrupted basal lamina, for example, in the case of laminin, type IV collagen, and perlecan but not in the case of nidogen (Garcia-Alonso et al., 1996; Costell et al., 1999; Kim and Wadsworth, 2000; Miner and Li, 2000; Murshed et al., 2000; Norman and Moerman, 2000). Although genetic disruption of nidogen does not affect basal lamina integrity, antibodies to the nidogen binding site on laminin disrupt the basal lamina in vitro (Kadoya et al., 1997). Therefore, interference of laminin–nidogen interactions with antibodies and peptides could result in a disruption of the basal lamina. Additionally, basal lamina disruptions could alter the localization of secreted guidance molecules. For example, secreted members of the semaphorin family may require laminin or another component of the basal lamina for proper localization.
To assess whether neurons ceased growth because of an alteration of the basal lamina or distribution of secreted semaphorins, laminin and semaphorin 2a distributions were examined in embryos cultured in the presence of blocking peptide. If the basal lamina had suffered a structural defect, then laminin immunoreactivity may appear disrupted. In laminin–nidogen peptide-blocked embryos, immunofluorescence of laminin was similar to that of control embryos (Fig.7A,B). Laminin protein was evenly distributed similar to control cultures, indicating that the laminin network was not severely disrupted. Figure 7Bdepicts stalled Ti1 growth cones in the trochanter, surrounded by a normal laminin distribution. To determine whether secreted guidance molecules are disrupted in blocked embryos, semaphorin 2a immunofluorescence was used. Semaphorin 2a is secreted and is normally localized in a dorsal-to-ventral and distal-to-proximal gradient (Fig.7C; Isbister et al., 1999; Bonner and O'Connor, 2000). In peptide-blocked embryos, semaphorin 2a distribution is similar to that of control cultures, suggesting that at least in the case of semaphorin 2a, secreted guidance cue localization was intact. Stalled Ti1 axons therefore have access to appropriately localized semaphorin 2a (Fig.7D, arrow), which repels Ti1 axons from the dorsal limb compartment. Furthermore, from these experiments, it appears that the extracellular graded distribution of semaphorin 2a does not require laminin–nidogen interactions.

Basal lamina and secreted guidance cue localization are intact in laminin- and nidogen-blocked embryos.A, Immunofluorescence microscopy of a control embryo that has been double-labeled with laminin antisera (red) and HRP (green). B, DPNAV-blocked embryo that has been double-labeled with laminin (red) and HRP (green). Ti1 axons stall in the trochanter (arrow), and basal lamina integrity is normal. C, Semaphorin 2a (Sema 2a), a secreted guidance cue, is distributed in a dorsal-to-ventral and distal-to-proximal gradient (Isbister et al., 1999) in control cultures. D, Semaphorin 2a distribution is preserved in DPNAV-blocked embryos. In this embryo, Ti1 axons (green) have stalled in the trochanter (arrow), and semaphorin 2a (red) is expressed in a dorsal-to-ventral and distal-to-proximal gradient. Scale bar, 20 μm.
DISCUSSION
During the establishment of the Ti1 pathway, laminin immunoreactivity is evenly distributed in the basal lamina. This suggests that laminin is a permissive cue for migrating Ti1 axons. β1-integrin was found on the cell bodies, axons, and importantly, on the growth cones of Ti1 neurons, implicating these receptors as potential mediators of laminin signals. Disruption of the laminin–nidogen interaction during development of the Ti1 pathway disrupts pioneer neuron pathfinding. Typically, the Ti1 growth cones cease proximal extension and cannot navigate an essential ventral turn, resulting in stalled axons. This does not appear to be attributable to a slower growth rate, because blocked Ti1 axons arrive at the trochanter at the same time. In addition, extending the length of the culture period does not overcome the effect of peptides and antibodies. Localization of stalled growth cones indicates that they stall within a region of semaphorin expression in the trochanter limb segment, and in the case of secreted semaphorins, these cues are properly localized. In addition, the basal lamina is not disrupted in antibody- and peptide-blocked embryos.
The basal lamina is intact in laminin–nidogen-blocked embryos
Because the antibodies and peptides used in this study were designed to disrupt basal lamina interactions, and were found to disrupt the integrity of the basal lamina in vitro (Kadoya et al., 1997) it was essential to determine the structural integrity of the basal lamina in laminin- and nidogen-blocked embryos. We demonstrated that laminin and semaphorin 2a distribution was unaffected in laminin- and nidogen-blocked embryos at the light microscopic level. Thus from these observations we conclude that blocking laminin and nidogen using antibodies and peptides causes a discrete and specific molecular lesion that does not compromise the integrity of the basal lamina or the localization of other secreted molecules. Furthermore, blocking a cell adhesion site on the β-chain of laminin that is involved in modulating netrin activity (Hopker et al., 1999) has no effect on Ti1 pathfinding (data not shown). Therefore, the defects observed with laminin and nidogen blocking are a direct result of blocking this site and not a general effect of blocking laminin or a secondary effect of disrupting basal lamina interactions.
Role of basal lamina in neuronal pathfinding
We have shown a surprising role of laminin in neuronal pathfinding. Previous work by Condic and Bentley (1989a) suggests an adhesive role for the basal lamina, because they found that enzymatic removal of the basal lamina resulted in decreased neuronal adhesion to the substratum and axon retraction to the cell bodies. Surprisingly, Ti1 axons could extend and pathfind in the absence of a basal lamina (Condic and Bentley, 1989c). One possible explanation for this is that enzymatic removal of the basal lamina may result in a compensatory mechanism within Ti1 growth cones, such that they no longer depend on the basal lamina for adhesive contacts. Consistent with this idea, they found that in the absence of the basal lamina, Ti1 axons could adhere to the epithelium in regions where they normally relied on basal lamina adhesion as well (Condic and Bentley, 1989c). This indicates that the neurons are capable of compensating for the lack of basal lamina interactions, perhaps by downregulating basal lamina receptors such as integrins and upregulating cell adhesion molecules. It is possible that in our study, disrupting the laminin–nidogen interactions may be insufficient to effect the expression of cell adhesion molecules of the Ti1 growth cones. Therefore, if a compensatory mechanism within Ti1 growth cones accounts for proper pathfinding in the absence of basal lamina, as in the studies of Condic and Bentley (1989a,c), this mechanism may not have been activated in laminin- and nidogen-blocked embryos.
Growth cone responsiveness may underlie permissive cue signaling
The pathfinding of neuronal growth cones relies on several factors, which can be classified as instructive or permissive. A balance of growth cone adhesion to the substratum and growth cone motility is essential to guarantee growth cone responsiveness. If this balance is disrupted, the growth cone may not be able to respond to instructive cues, resulting in pathfinding errors. Hopker et al. (1999)have demonstrated that laminin can alter the guidance activity of the laminin-related netrin family of molecules, suggesting that the simultaneous presentation of instructive and permissive cues is pertinent to the effect of guidance molecules. In the developing limb bud, laminin has the potential to modulate the activity of several guidance molecules because of its ubiquitous and even distribution in the basal lamina during axon outgrowth. In this capacity, laminin could be an important cofactor to guidance molecules that guide Ti1 neurons.
Because the growth cone is in constant contact with the environment, sampling and responding to many molecular cues, disruption of laminin–nidogen interactions in the immediate environment of the Ti1 growth cones could have severe effects on the molecular constituents found on the growth cone. It has recently been demonstrated that integrin levels in neuronal growth cones can be rapidly modulated based on availability of both laminin and aggrecan in vitro(Condic and Letourneau, 1997; Condic et al., 1999), implicating integrins as mediators of the balance between adhesion to the substratum and growth cone motility. Condic and Letourneau (1997) have established that when neurons are plated on low laminin concentrations, integrin receptors increase on the growth cones, resulting in increased adhesion to various substrates, suggesting that neurons can rapidly and dynamically respond to their environment and can maintain a balance between adhesion and motility. When applied to our system, blocking laminin in the developing embryo could increase integrin receptors on the growth cone, thereby increasing adhesion to the substratum, resulting in decreased motility at the trochanter.
Why is motility inhibited at the ventral turn and not before? One possibility is that a combination of permissive and instructive cues is important for generating the abrupt turn that growth cones make in the trochanter. For example, although an instructive guidance cue may be signaling the growth cone with respect to the directionality of a turn, a concomitant permissive signal may also be required. This scenario would suggest that the pathfinding in the femur does not require laminin signaling. In this region, the graded expression of semaphorin 2a may be sufficient for guiding Ti1 axons proximally (Isbister et al., 1999). In contrast, possibly a number of cues, including laminin, are required to generate the abrupt ventral turn in the trochanter.
Although Ti1 axons may be incapable of turning, why do the growth cones stall in the trochanter and not continue proximal growth? Previous work has demonstrated that the trochanter epithelium is substantially more adhesive for Ti1 growth cones than epithelium in other areas of the limb bud (Condic and Bentley, 1989b; Isbister and O'Connor, 1999). The simplest explanation is that in the absence of a turning signal, the Ti1 growth cones prefer to remain on the highly adhesive trochanter rather that extend proximally into the coxa. This could be mediated in part by upregulation of receptors on the growth cones, which could increase growth cone adhesion.
Potential roles of nidogen
When the nidogen-binding site on laminin is blocked, Ti1 axons stall at the location of a stereotyped ventral turn. Because laminin is evenly expressed in this region, it is not likely that laminin is a chemoattractant that directs axons ventrally, nor is it likely to be a chemorepulsive agent that repels axons from the dorsal epithelium. Therefore, why do disruptions in laminin–nidogen interactions perturb axon pathfinding? One explanation is that the defects observed could be attributable to disruption of nidogen function rather than laminin function. Once nidogen is removed from the laminin heterotrimer, it is susceptible to degradation by proteases (Mayer et al., 1993b), however, it is unclear whether this is the case in laminin–nidogen-blocked embryos. The expression pattern of nidogen in the developing embryo is unknown, and the possibility that nidogen is discretely expressed in the developing limb bud cannot be discounted. As a result, discrete localization of nidogen could guide Ti1 neurons directly. Alternatively, nidogen could act in concert with other guidance cues to facilitate the steering event. Kim and Wadsworth (2000) have demonstrated that nidogen is essential for pathfinding of neurons inCaenorhabditis elegans and may modulate the effect of other guidance cues such as netrins. Thus nidogen has important guidance activity and could be the sole mediator of the stalled Ti1 axon effect.
As they migrate, growth cones encounter different cues that are spatially and temporally regulated. The exact sequence of encountered cues may be critical for the growth cone to appropriately respond to future guidance information, suggesting that the growth cone retains information, perhaps in the form of modulated receptor subtypes (Dodd et al., 1988; Shirasaki et al., 1998; Brose and Tessier-Lavigne, 2000;Diefenbach et al., 2000). Because the ventral turn of the Ti1 neurons involves multiple cues, we speculate that permissive cues are responsible for guaranteeing that the growth cone expresses the appropriate complement of receptors for permissive cues, such as integrins, to facilitate axon pathfinding.
Footnotes
This work was funded by a Rick Hansen Institute Neurotrauma Initiative studentship to J.B., by Rick Hansen Institute Neurotrauma Initiative Grant 99019 to T.P.O., and by Canadian Institutes of Health Research Grant 13246 to T.P.O. T.P.O. is an EJLB Research Scholar. We gratefully acknowledge Sal Carbonetto for β1-integrin antibodies, Kim Gerrow for providing limb fillets, and Ken Norman for confocal expertise. In addition, we thank Katharine Sepp and Joost Schulte for critical reading of this manuscript.
Correspondence should be addressed to Timothy P. O'Connor, Department of Anatomy, Program in Neuroscience, University of British Columbia, 2177 Wesbrook Mall, Vancouver, British Columbia, Canada V6T 1Z3. E-mail: jimo{at}interchange.ubc.ca.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}