
Abstract
The macaque flexor digitorum profundus (FDP) consists of a muscle belly with four neuromuscular regions and a complex insertion tendon that divides to serve all five digits of the hand. To determine the extent to which compartments within FDP act on single versus multiple digits, we stimulated the primary nerve branch innervating each neuromuscular region while recording the tension in all five distal insertion tendons. Stimulation of each primary nerve branch activated a distinct region of the muscle belly, so that each primary nerve branch and the muscle region innervated can be considered a neuromuscular compartment. Although each neuromuscular compartment provided a distinct distribution of tension across the five distal tendons, none acted on only one digital tendon. Most of the distribution of tension to multiple digits could be attributed to passive biomechanical interactions in the complex insertion tendon, although for the larger compartments a wider distribution resulted from the broad insertion of the muscle belly. Nerve ligations excluded contributions of spinal reflexes or distal axon reflexes to the distribution of tension to multiple digits. We conclude that the macaque FDP consists of four neuromuscular compartments, each of which provides a distinct distribution of tension to multiple digits.
- compartment
- digits
- distribution
- fingers
- force
- hand
- macaque
- multitendoned
- muscle
- neuromuscular
- tension
- thumb
Several muscles in various mammalian species in recent years have been shown to consist of multiple neuromuscular compartments. A compartmentalized muscle consists of multiple anatomic regions of muscle fibers, each region innervated by a separate primary nerve branch (English and Weeks, 1984;Balice-Gordon and Thompson, 1988; English, 1990; Segal et al., 1991;Drake et al., 1993; Sanders et al., 1994; Vanden Noven et al., 1994;Liu et al., 1997). The regional glycogen depletion produced by prolonged stimulation of a given primary nerve branch indicates that each compartment contains all the muscle fibers of motor units whose axons enter the muscle through that nerve branch (English and Letbetter, 1982). The borders between different compartments often are sharply defined by fascicular boundaries within the muscle, although in some cases interdigitation of muscle fibers from adjacent compartments occurs at a less discrete boundary. EMG recordings during active behaviors demonstrate that each compartment can be activated by the nervous system relatively independently of the others (English, 1984;English and Weeks, 1987; Pratt and Loeb, 1991; Pratt et al., 1991). Because compartmentalized muscles often have distributed origins and/or distributed insertions, contraction of the various compartments can provide different mechanical effects for generating movements and stabilizing postures (Chanaud et al., 1991; Fritz et al., 1992).
The multitendoned extrinsic finger muscles of the hand have extremely distributed insertions. Unlike most other muscles, these muscles give off multiple insertion tendons, each attaching to a different digit. In macaque monkeys, each of these muscles consists of multiple regions of muscle fibers innervated by different nerve branches (Serlin and Schieber, 1993). One such muscle is the flexor digitorum profundus (FDP). In both monkeys and humans, various regions of FDP can be activated differentially under voluntary control from the nervous system (Jeneson et al., 1990; Schieber, 1993; Kilbreath and Gandevia, 1994). Because most of the power for flexing and extending the digits is provided by extrinsic muscles (Long et al., 1970; Li et al., 2000), knowing whether the neuromuscular compartments of multitendoned muscles each act on a single digit versus multiple digits is key to understanding neural control of finger movements.
The degree to which compartments of multitendoned finger muscles distribute tension across the digits has yet to be examined, however. In the present study, we therefore determined how the tension generated by each region of the macaque FDP is distributed among the five digits of the hand by directly stimulating each primary nerve branch. Comparing the tension distributions produced by passive loading with those produced by active contraction, we also examined the extent to which distribution of tension to multiple digits resulted from passive mechanical connections among the tendons versus the action of each region of the muscle belly. Finally, we cut first the proximal median and ulnar nerves and second each primary nerve branch to determine whether spinal reflexes or axon reflexes, respectively, contributed to the distribution of tension to multiple tendons.
MATERIALS AND METHODS
All procedures for the care and use of these purpose-bred monkeys complied with the United States Public Health Service Policy on Humane Care and Use of Laboratory Animals, followed the Public Health Service Guide for the Care and Use of Laboratory Animals, and were approved by the University Committee on Animal Resources at the University of Rochester. All procedures were performed under deep surgical anesthesia.
Surgical preparation. Studies were performed on five male pigtail monkeys (Macaca nemestrina, 2–4 kg). Monkeys were premedicated with ketamine (15 mg/kg, i.m.) and atropine (0.04 mg/kg, i.m.) and then anesthetized with thiopental (25 mg/kg, i.v.), using supplemental doses as needed to maintain deep surgical anesthesia judged by the absence of blink and pinna reflexes. Monkeys were intubated for airway control. Heart rate and blood oxygen saturation were monitored continuously with a pulse oximeter. Core body temperature was monitored with a rectal probe and maintained at 34–37°C with a heating blanket.
Once the monkey was surgically anesthetized, the tendons of FDP were dissected free in each digit of the right hand. The skin on the palmar surface of each digit was incised longitudinally from the metacarpophalangeal joint to the distal phalanx. For eventual attachment to tension transducers, a separate 2-0 supramid suture was tied through and around each FDP tendon proximal to the distal interphalangeal joint. The tendons were then cut from their insertion on the distal phalanges, and each finger was amputated at the proximal interphalangeal joint.
Next, the skin on the ventral surface of the right forelimb was opened with a single incision extending from antecubital fossa to the volar aspect of the wrist. Exposed tissues were kept moist with warmed Tyrode's solution (Sigma, St. Louis, MO). The tendons of flexor carpi radialis (FCR), palmaris longus (PL), and flexor digitorum superficialis (FDS) were severed at the wrist; these muscles were dissected proximally, exposing the median nerve just deep to FDS. The median nerve then was followed proximally. Branches of the median nerve innervating FCR, PL, FDS, FDP, and pronator teres (PT), as well as the anterior interosseous nerve (AIN), were dissected and identified with electrical stimulation. For purposes of nerve branch identification, electrical stimuli consisted of biphasic 50–200 μsec pulses of 5–5000 μA at 1 Hz produced by a battery-powered optically isolated stimulator (Bak BSI-1; Bak Electronics, Germantown, MD) and were delivered via a bipolar pair of hook electrodes spaced 2.5–5 mm apart. Nerve branches not contributing innervation to FDP were cut, denervating all other forearm muscles innervated by the median nerve. However, the AIN, which innervates pronator quadratus, was left intact. FCR, PL, and FDS were dissected to their origins and removed. PT was divided, exposing the median nerve trunk beneath. The ulnar nerve was dissected in a similar manner, and branches innervating flexor carpi ulnaris (FCU) and FDP were identified with stimulation. Ulnar nerve branches innervating FCU were cut, although the muscle was leftin situ. Finally, the median and ulnar nerves were cut just proximal to the wrist, denervating the intrinsic hand muscles.
Bone screws then were placed in the distal radius, in the olecranon process of the ulna, and in some experiments in the distal ulna as well. The monkey was placed in a right lateral decubitus position, with the shoulder protracted ∼120° and the elbow extended ∼120°. Via the bone screws, the forearm was then mounted rigidly, ventral side up, on an underlying frame. The skin flaps were used to construct a pool that was filled with warm mineral oil to protect the exposed nerves and muscles. Note that this preparation left the tissues of the carpal tunnel and palm intact (Fig. 1) to preserve any biomechanical effects of these tissues on tension distribution among the FDP tendons.

Experimental setup. Each of the five digital tendons of FDP was tied to a separate, independently positioned tension transducer, served by its own signal conditioner channel. Tissues of the palm and carpal tunnel were not dissected so that biomechanical interconnections here would be as close as possible to those in the intact animal. Bone screws placed in the distal radius, distal ulna, and proximal ulna fixed the forearm to an underlying frame, and a horizontal bar supported the metacarpus against gravity. Superficial forearm flexor muscles were dissected and removed, exposing the belly of FDP and its nerve supply. A protective oil pool was constructed with the skin flaps. Each primary nerve branch in turn was then elevated into the oil pool on a bipolar hook electrode for stimulation.
The suture in each tendon then was tied to a separate tension transducer (Load Cell BG-1000 gm; Kulite Semiconductor Products, Inc., Leonia, NJ) mounted on a custom-made miniature rack and pinion that permitted adjustment of resting tendon length (Fig. 1). All five transducers were positioned such that the resting tension was ∼100 gm in each tendon. The signal from each transducer was passed through a conditioner (model 1902; Cambridge Electronic Design, Cambridge, UK) that permitted gain switching between 10×, 30×, and 300×. Each amplified signal was digitized at 2 kHz (micro1401; Cambridge Electronic Design) and stored to disk using Spike2 software (Cambridge Electronic Design) run on a computer (Gateway, San Diego, CA). Over the 0–1500 gm range typically used, the system provided a resolution of 0.4 gm/bit.
In four experiments, compound muscle action potentials (CMAPs) were recorded via bipolar electrodes with closely spaced (0.15 mm) tips made from twisted pairs of Teflon-insulated, stainless steel wires cut across immediately before implantation in the muscle (Schieber, 1993,1995). The tips of each bipolar pair were inserted together retrograde 2–3 mm into a 22 gauge hypodermic needle, the wires were bent back parallel to the needle, and the needle was thrust into the muscle belly and withdrawn, leaving the bipolar wire pair hooked in the muscle. Such electrodes were implanted into the muscle belly of FDP at seven locations (Fig. 2): proximal, intermediate, and distal along the ulnar side of the muscle; proximal, intermediate, and distal along the radial side of the muscle; and in the superficial accessory head. EMG signals from the electrodes were amplified 1000–5000× (Grass P511; Grass Instruments, West Warwick, RI) and filtered (bandpass 100 Hz to 3 kHz), digitized at 4 kHz, and recorded to disk in parallel with tension records from the tension transducers. Selected channels were monitored on an analog oscilloscope.

Standard map of FDP. This drawing of the superficial aspect of FDP, as if dissected free from the rest of the right forearm and hand, indicates the four neuromuscular regions of the muscle: r, radial region, FDPr; u, ulnar region, FDPu; a, accessory region, FDPa;pu, proximal ulnar region, FDPpu. Whereas the first three regions are innervated by primary nerve branches arising from the median nerve trunk, the latter typically is innervated by a branch from the ulnar nerve. The course of each nerve branch after penetrating the muscle belly is indicated by a string of symbols: ♦, FDPr; ▪, FDPu; ▴, FDPa; and ●, FDPpu. The locations of electrodes inserted to record compound muscle action potentials are indicated by ★: rd, radial distal; ri, radial intermediate; rp, radial proximal; a, accessory; ud, ulnar distal; ui, ulnar intermediate; up, ulnar proximal. The proximal insertion aponeurosis is indicated by stippling, with longitudinal furrows in the aponeurosis shown as horizontal black lines. a–l indicate the locations at which suture loops were tied for passive loading. A bracketindicates the portion of the insertion tendon that lies in the carpal tunnel. The digital insertion tendons on the thumb through the little finger are labeled t1 through t5, respectively.
Data collection. Each primary nerve branch innervating FDP was placed in turn on a bipolar hook electrode, elevated into the mineral oil pool, and stimulated with 50 μsec biphasic, constant current pulses at 1 Hz. Stimulating current was slowly increased from ineffective levels until each pulse produced a just detectable twitch in one or more tension records; a visible twitch of the muscle belly usually became detectable simultaneously. The current required to produce a just detectable twitch was recorded as threshold current. The stimulus current then was increased further until additional increments of current produced no additional increase in tension; this was recorded as the current for maximal contraction. A recording of tension on each tendon and EMG activity from each intramuscular electrode was then made as supramaximal stimuli (2× current for maximal contraction) were delivered for 10 sec at 1 Hz, followed by 1 sec episodes of tetanic stimulation at 5, 10, 20, and 40 Hz, with 10 sec between each stimulus train. Once the tension produced by stimulation of each primary nerve branch had been recorded, the median and ulnar nerves both were cut in the upper arm, eliminating any spinal reflex contribution to tension distribution, and each primary nerve branch was stimulated again in turn. Thereafter, each primary nerve branch was cut proximally from the median or ulnar nerve, eliminating any contribution to tension distribution from axon collaterals, and each primary nerve branch was stimulated again.
To exclude the possibility that the AIN contributed innervation to the midportion of FDP, we stimulated the AIN in each animal with the same protocol used to study the primary nerve branches of FDP. AIN stimulation never produced appreciable tension in the FDP tendons, however. Therefore, we confirmed in each experiment that the AIN does not contribute to innervation of FDP and have not included data from AIN stimulation in our analyses below.
After recording the distribution of active tension to different FDP tendons produced by stimulating each primary nerve branch, we recorded the distribution of tension produced passively by pulling at different radioulnar locations on the FDP tendon just proximal to the wrist. At this level, the muscular portion of the FDP belly is ending, but the FDP insertion tendon still consists of a superficial aponeurosis not yet divided into separate tendons for each digit, which ultimately occurs within the palm (Serlin and Schieber, 1993). Proximal to the wrist, a distinct central furrow typically can be identified separating the radial and ulnar portions of the insertion aponeurosis, and the ulnar portion typically has a second, less distinct furrow separating an intermediate and a more ulnar longitudinal condensation of aponeurotic tissue. We placed four separate figure-of-eight suture loops at different radioulnar locations in the FDP insertion aponeurosis just proximal to the wrist, typically placing two suture loops in the radial portion of the tendon and two in the ulnar portion, with one in the intermediate and another in the most ulnar condensation. Each of the four suture loops in turn was connected to a 1 kg weight suspended proximal to the elbow via a suture run over a pulley, such that the direction of pull exerted by the weight on the tendon was as close as possible to the long axis of the forearm. The tension in all five digital tendons was then recorded as the 1 kg weight was raised and lowered by hand from beneath to examine the distribution of tension produced passively by mechanical interconnections among the more distal tendons and other soft tissues in the palm. At the end of all studies, the monkey was killed with a lethal injection of thiopental.
Data analysis. Data files were reviewed off-line with software that displayed the recorded tension waveforms graphically and permitted time and amplitude measurements to be made either with a mouse-controlled cursor or with a script routine that exported values to text files (Spike2; Cambridge Electronic Design). Additional analyses of exported data were performed on a minicomputer using Excel 97 (Microsoft, Seattle, WA) and SPSS (SPSS, Inc., Chicago, IL) software.
To examine the distribution of tension across the five tendons of FDP, we measured changes in tension on each tendon for single twitches and for 5, 10, 20, and 40 Hz tetani delivered to each primary nerve branch. Average tension was measured over a 1000 msec window beginning at each stimulus train onset, and from these values the average tension in a 500 msec baseline period immediately preceding stimulus onset was subtracted.
These tension measurements were used to calculate two indexes quantifying how the neuromuscular compartment innervated by each primary nerve branch distributed tension among the digital insertion tendons of FDP (Fritz et al., 1992; Schieber et al., 1997). Computation of these indexes is simplified by normalizing the tension in each tendon as a fraction of the total tension exerted on all tendons:
 where τi
is the fraction tension change in the ith tendon of a muscle,Ti
is the absolute tension change, andn is the number of tendons.
where τi
is the fraction tension change in the ith tendon of a muscle,Ti
is the absolute tension change, andn is the number of tendons.
The output index (OPI) quantifies the center of effort of the contraction from −1 to +1, with −1 representing all tension exerted on the most radial tendon, 0 representing tension distributed symmetrically about the center, and +1 representing all tension exerted on the most ulnar tendon. The OPI is calculated as:
 where τi
is the fraction of the total tension exerted on the ith tendon, n is the number of tendons, and wi
is a constant that provides a rank-ordered weighting of the tendons:
where τi
is the fraction of the total tension exerted on the ith tendon, n is the number of tendons, and wi
is a constant that provides a rank-ordered weighting of the tendons:
 The selectivity index (SEL) quantifies the degree to which tension is focused on only one tendon versus being spread across multiple tendons, independent of which particular tendons receive what fractions of the total tension. The fractional tensions of an ideally unselective compartment that exerted equal tension on all tendons of a muscle would be τu
= 1/n, whereas the fractional tensions of an ideally selective compartment that exerted tension on only one tendon, t1 for example, would be τ1
= 1,τ2
= 0, … .τn
= 0. In ann-dimensional fractional tension space, the linear distance between these two points would be:
The selectivity index (SEL) quantifies the degree to which tension is focused on only one tendon versus being spread across multiple tendons, independent of which particular tendons receive what fractions of the total tension. The fractional tensions of an ideally unselective compartment that exerted equal tension on all tendons of a muscle would be τu
= 1/n, whereas the fractional tensions of an ideally selective compartment that exerted tension on only one tendon, t1 for example, would be τ1
= 1,τ2
= 0, … .τn
= 0. In ann-dimensional fractional tension space, the linear distance between these two points would be:
 The linear distance, d, between the point representing any other compartment and the ideally unselective compartment would be:
The linear distance, d, between the point representing any other compartment and the ideally unselective compartment would be:
 The SEL is then calculated as
The SEL is then calculated as
 SEL will vary from 0 for an ideally unselective compartment to 1 for an ideally selective compartment.
SEL will vary from 0 for an ideally unselective compartment to 1 for an ideally selective compartment.
RESULTS
Innervation of the four regions of FDP
Previous anatomical studies have shown that the macaque FDP receives four primary nerve branches, each innervating a separate region of the muscle belly (Serlin and Schieber, 1993; Maurer et al., 1995). As illustrated schematically in Figure 2, the large radial (FDPr) and ulnar (FDPu) regions each receive a primary branch from the median nerve. A smaller accessory region (FDPa), located proximal and superficial to the radial region, is also innervated by a median nerve branch. The most proximal portion of the ulnar head (FDPpu), however, is usually innervated by a branch from the ulnar nerve.
In the present experiments, two of the five animals showed atypical innervation of region FDPpu. In one, the proximal ulnar region was innervated by two nerve branches: a typical nerve branch arising from the ulnar nerve was unusually small, and an atypical branch from the median nerve also innervated the proximal ulnar region of FDP in this animal. Stimulation of this atypical median nerve branch produced a pattern of EMG activation and a distribution of tension characteristic of region FDPpu in the three animals with typical innervation (see below), whereas stimulation of the typical but small branch from the ulnar nerve did not produce appreciable tension. Therefore, we included the data from stimulation of the atypical median nerve branch in our analyses of region FDPpu below. In the second animal with atypical innervation, we could find no primary nerve branch innervating the proximal ulnar region, and no data for FDPpu were collected in this experiment. Therefore, our analyses of FDPpu below were based on data from only four animals (three in which a typical branch from the ulnar nerve innervated region FDPpu and one in which this region received an atypical primary nerve branch from the median nerve).
In our previous anatomical studies of the macaque FDP, each primary nerve branch appeared to enter a separate region of muscle fibers, with little evidence that nerve fibers crossed into another region after entering the muscle. To examine physiologically whether excitation produced by stimulation of each nerve branch was confined to the expected region of FDP, in four of the present experiments we recorded CMAPs from bipolar intramuscular electrodes inserted at seven sites in the FDP muscle belly (Fig. 2, ★). Figure3A shows examples of the CMAPs recorded at all seven sites after stimulation of each primary nerve branch in one experiment. For analysis, the average level of rectified EMG activity from 6 to 25 msec after stimulus onset was normalized for maximum activity at each electrode location. Figure 3B shows normalized EMG activity from each electrode averaged across these four experiments. After stimulation of the nerve branch to FDPr, electrodes inserted at proximal (rp), intermediate (ri), and distal (rd) locations on the radial side of FDP recorded activity, whereas little or no activity was recorded by other electrodes. Stimulation of the FDPa nerve branch resulted in EMG activity confined to the FDPa electrode (a). Stimulation of the FDPu nerve branch produced EMG activity chiefly in the distal (ud) and intermediate (ui) electrodes on the ulnar side of the muscle, with somewhat less activity recorded in the proximal ulnar electrode (up). Conversely, stimulation of the FDPpu nerve branch evoked EMG activity chiefly in the proximal ulnar electrode with less in the intermediate ulnar electrode. Although the boundary between the FDPu and FDPpu regions thus may be indistinct, stimulation of each primary nerve branch produced electrical activation of the appropriate region of the FDP muscle belly, with little evidence of activity in other regions.

Electromyographic localization.A, Compound muscle action potentials recorded simultaneously by intramuscular electrodes at each of the seven locations (rd through up), as indicated in Figure 2, are shown after a single supramaximal stimulation pulse in a 1 Hz train delivered to each primary nerve branch in turn (FDPr, FDPu, FDPa, or FDPpu) in a single experiment. The amplification of each channel was constant throughout, and the vertical scale represents ±5 V for each channel. The stimulation pulse was delivered to each nerve branch at the time indicated by the arrows at thebottom. B, The amplitude of the compound muscle action potentials recorded by a given electrode during stimulation of each primary nerve branch was normalized from 0 to 1 in each experiment, and these normalized values were averaged across experiments. Lines connect the pointsrepresenting the mean ± SD normalized EMG responses recorded from each of the seven intramuscular electrodes during stimulation of a given nerve branch. Abbreviations for electrode locations are as described in the legend to Figure 2. Stimulation of each primary nerve branch evoked compound muscle action potentials localized to the appropriate region of the muscle. For most points, the SD error bars fell inside the symbol representing the mean. Normalized EMG amplitude was most variable near the ulnar proximal and ulnar intermediate electrodes, consistent with the variable innervation of region FDPpu found in these monkeys.
Effects of stimulation frequency
Stimulation of a given primary nerve branch produced different amounts of tension on the five digital tendons of FDP. Figure4A shows, for example, the raw data record from stimulation of the FDPr nerve branch in one experiment. Stimulation at 1, 5, and 10 Hz produced unfused twitches, whereas progressive temporal summation produced successively greater tensions at 20 and 40 Hz. For all frequencies of stimulation, the greatest amount of tension was exerted on tendon 3 and the least amount on tendon 5. Figure 4B shows the 1 sec mean tensions exerted on each FDP tendon measured from these data as a function of stimulation frequency. Because twitches did not summate at frequencies up to 10 Hz, mean tension increased linearly in each tendon from 1 to 10 Hz. Tension increased more rapidly from 10 to 20 Hz, as twitches temporally summated, with a relatively slower additional increase from 20 to 40 Hz. At all frequencies of stimulation, however, the rank order from the tendon receiving the most tension to the tendon receiving the least tension remained constant (t3 > t2 > t4 > t1 > t5). Furthermore, the ratios of tension exerted on different tendons remained relatively constant as well.
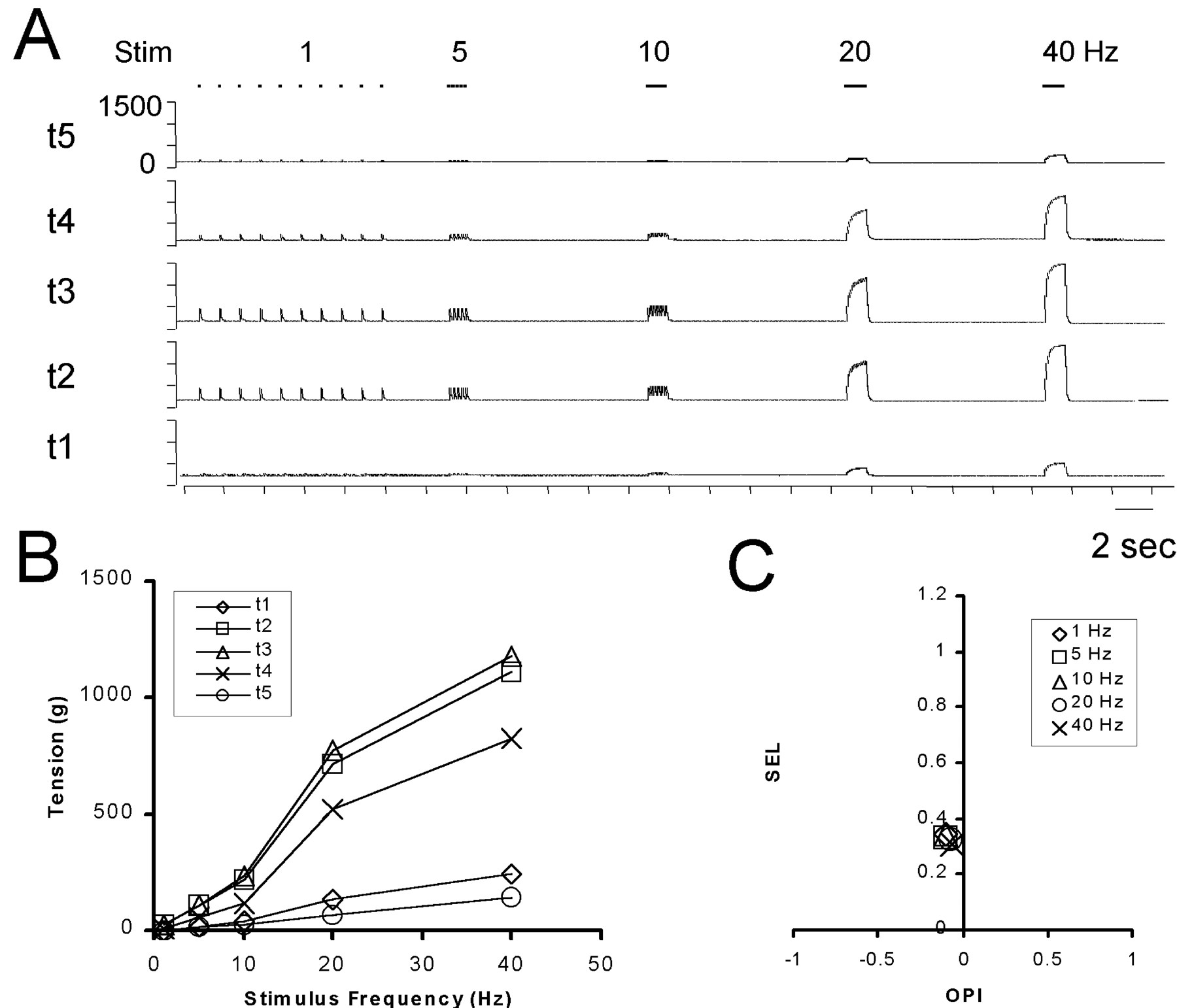
Effects of stimulation frequency.A, An original recording shows tension in each FDP tendon during stimulation of an FDPr primary nerve branch at 1, 5, 10, 20, and 40 Hz. The vertical scale representing 0–1500 gm is the same for each digital tendon, t1 through t5.B, Mean tension (over 1 sec) in each tendon has been plotted as a function of stimulation frequency. Data are from the record shown in A. C, SEL and OPI indexes calculated from the same data are plotted against one another for each stimulus frequency to demonstrate that the distribution of tension among the five tendons of FDP did not vary with stimulation frequency.
We also evaluated the degree to which tension distribution varied with stimulus frequency by calculating the SEL and OPI at each stimulation frequency. The SEL will be 1 if all tension is exerted on a single tendon and 0 if tension is evenly distributed across all tendons. The OPI describes the center of effort of the tension from −1 for the most radial tendon to +1 for the most ulnar tendon. The SEL and OPI indexes for the data shown in Figure 4, A andB, are plotted against one another in Figure 4C. At all stimulation frequencies, SEL clustered at low values from 0.30 to 0.35, indicating that the tension was distributed across multiple tendons. The OPI also remained relatively constant at approximately −0.1 for each frequency, demonstrating that the tension maintained the same center of effort, slightly radial to t3. In general, the SEL and OPI remained relatively constant for stimulation of a given nerve branch at different frequencies. Because the distribution of tension thus remained relatively constant at all stimulus frequencies, because 20 Hz represents a relatively high physiological discharge frequency and because 40 Hz stimulation occasionally produced tensions large enough to saturate our transducers, we chose tensions exerted during 20 Hz stimulation to characterize the distribution of tension across the five tendons in the following analyses.
Tension distribution by the different regions of FDP
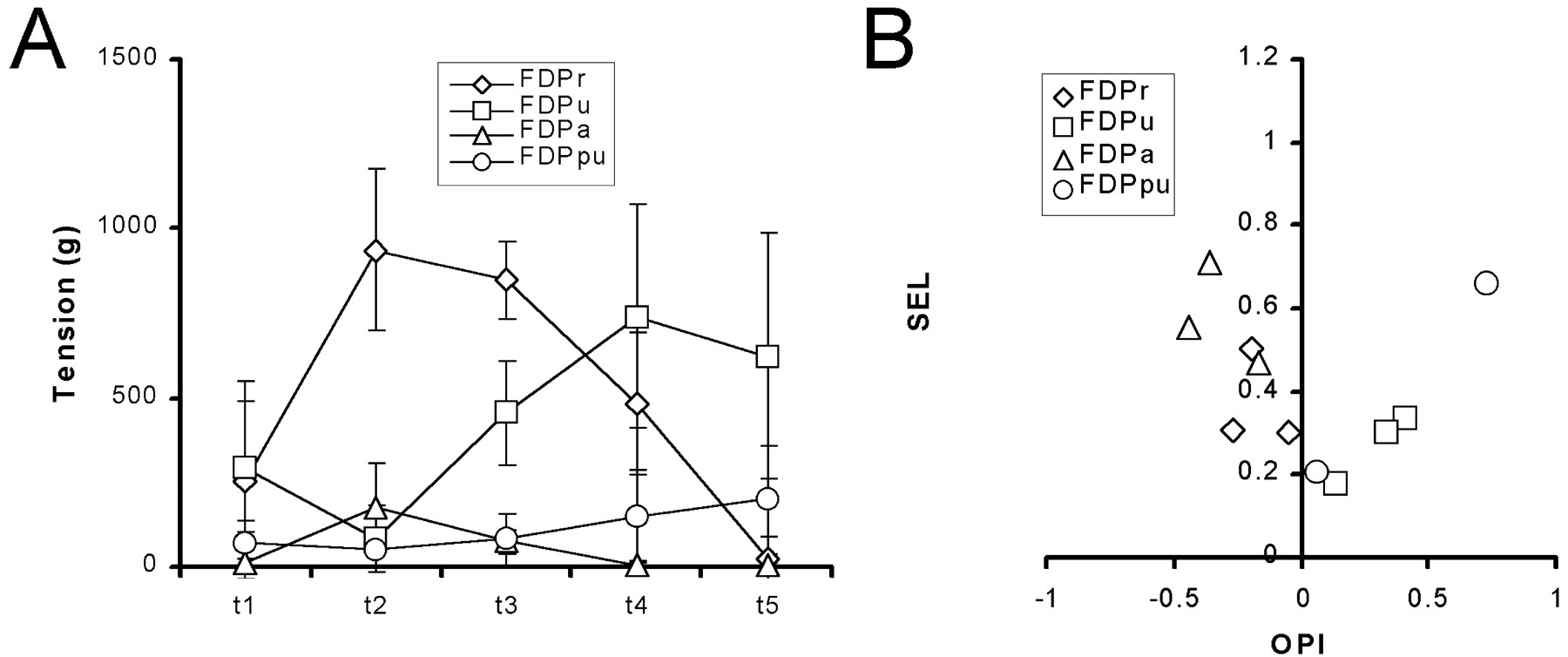
Contraction of each region of FDP produced a different distribution of tension among the tendons. Although the absolute tension exerted on a given tendon varied between animals in relation to animal size, the relative distribution of tension among the tendons produced by a given region tended to be similar from monkey to monkey. Figure 5A shows the distribution of 1 sec mean tension during 20 Hz stimulation of each region (FDPr, FDPu, FDPa, and FDPpu) averaged across monkeys. Contraction of FDPr resulted in the greatest amount of tension on tendons 2 and 3, with less tension on tendons 1 and 4 and little if any tension on tendon 5. FDPu produced the greatest tension on tendons 4 and 5, less on tendons 1 and 3, and very little on tendon 2. FDPa produced tension on tendon 2, less on tendon 3, and very little on tendon 1; the tension produced by FDPa on tendons 2 and 3 was always less than that produced by FDPr in the same animal. FDPpu produced more tension on tendon 5 than tendon 4, although in all cases the tension was less than that produced by FDPu. Each region of FDP thus produced a distinct and characteristic distribution of tension across the tendons. The two larger regions of FDP (FDPr and FDPu) produced quantitatively more tension than the two smaller regions (FDPa and FDPpu). The tension produced by the two smaller regions, however, was somewhat more selective than that produced by the two larger regions.

Tension distribution by the different regions of FDP. A, Plots of the tension exerted on each digital tendon during 20 Hz stimulation of each primary nerve branch. Values are the average across subjects with SD error bars. B, The SEL and OPI indexes calculated for the tension distribution produced by 20 Hz stimulation of each primary nerve branch in each experiment have been plotted against one another. (Note that, because of atypical innervation in one animal, only 4 points were available for FDPpu. Only 4 points were available for FDPu because stimulation of this nerve branch inadvertently was omitted at this stage in one experiment.) The same plot shows SEL versus OPI for passive loading at each location tested on the proximal insertion aponeurosis (Fig.7).
We examined these differences among the regions of FDP further using the OPI and SEL. These indexes for each region in each experiment are plotted against one another in Figure 5B. Each region tended to have its center of effort at a different radioulnar location. FDPa had the most radial OPI (mean of −0.36). FDPr had a less radial OPI (mean of −0.21). FDPpu had the most ulnar OPI (mean of +0.4), whereas the OPI of FDPu was less ulnar (mean of +0.26). The regions of FDP also differed in their selectivity, with the smaller regions exerting tension more selectively on particular tendons (mean SEL for FDPa and FDPpu, 0.56 and 0.45, respectively) than the larger regions (mean SEL for FDPr and FDPu, 0.35 and 0.26, respectively). The smaller regions of FDP thus exerted tension somewhat selectively on the radial (FDPa) or ulnar (FDPpu) tendons, whereas the larger regions, although producing quantitatively more tension, distributed their tension rather broadly over the more radial (FDPr) or ulnar (FDPu) tendons.
Although our sample size was small (n = 5 for FDPr and FDPa; n = 4 for FDPu and FDPpu), we performed multivariate ANOVA on the OPI and SEL values, confirming significant variation in these indexes by neuromuscular region (p < 0.01). Contrasts performed for each region against the others showed that, whereas the OPIs of the two more radial regions (FDPr and FDPa) were not significantly different and the OPIs of the two more ulnar regions (FDPu and FDPpu) were not significantly different, the OPIs of FDPr and FDPa each were significantly different from the OPIs of FDPu and FDPpu (p < 0.008; Bonferroni corrected value). Contrasts also demonstrated that, whereas SEL values for FDPpu did not differ significantly from other regions, SEL values for FDPa were significantly higher than SEL values for the two larger regions, FDPr and FDPu (p < 0.008).
Contributions of passive biomechanical interconnections to tension distribution
The FDP tendons to different digits are heavily interconnected from the muscle belly distally to the base of the digits. Most muscle fibers of the macaque FDP insert from below into a sheet-like aponeurosis on the superficial aspect of the muscle belly; only fibers of region FDPa insert from above (Serlin and Schieber, 1993). By the level of the carpal tunnel, this aponeurosis has developed longitudinal furrows (Fig. 2), but clearly separate tendons to the different digits do not arise until the level of the palm. Within the palm, the tendon to digit 2 comes from the radial aspect of the tendon sheet, the tendons to digits 5 and 4 come from the ulnar aspect, the tendon to digit 3 arises from both radial and ulnar bands on the deep surface, and the tendon to digit 1 arises from the central portion of the superficial aspect. These biomechanical interconnections among the FDP tendons can cause tension exerted at one point on the proximal insertion aponeurosis to be distributed among multiple distal finger tendons.
In three monkeys, we examined the extent to which these interconnections contribute to the tension distribution by applying a 1 kg load to the proximal insertion aponeurosis at four different radioulnar locations just proximal to the carpal tunnel. Each point to which the load was attached was marked on our standard drawing of FDP (Fig. 2, a–l). Figure6A shows the tension in each distal tendon recorded simultaneously as a 1 kg weight raised and lowered by hand was used to gradually load and unload one point in the proximal insertion aponeurosis (Fig. 2, point l). At any time in this recording, the greatest tension was transmitted to tendon 2 and the least to tendons 4 and 5. In Figure6B, these same data have been used to plot tension in a given tendon as a function of the total tension in all five tendons. As illustrated by this example, such plots tended to be quite linear, with little hysteresis during loading versus unloading. Therefore, we used the slope of these linear relationships to quantify the proportion of total tension passively distributed by biomechanical interconnections from proximal points in the insertion aponeurosis to each digital tendon.

Passive loading of the proximal insertion aponeurosis. A, An original record shows the simultaneous tension in all five distal tendons as a 1 kg weight was lowered (↓) and raised (↑) by hand, alternately applying and removing a proximally directed load to the proximal insertion aponeurosis of FDP at point l in Figure 2. The vertical scale represents 0–400 gm for each tendon. B, The data from A have been plotted as a function of the instantaneous total tension in all five tendons. The linear relationships indicate that a constant fraction of the total passive load on the proximal insertion aponeurosis was distributed to each distal insertion tendon.
Figure 7A shows the normalized distribution of tension produced by passive loading at each of the 12 points marked by letters a–l in Figure 2. Here each value is the slope of the single tendon versus the total tension relationship described above, equivalent to the fraction of the total tension summed across the five tendons. At only one point from one experiment was virtually all of the tension exerted at a proximal point near the radial edge of the proximal insertion aponeurosis (k) transmitted to a single distal tendon (t2). In all other cases, tension was distributed passively to multiple digital tendons. Nevertheless, each distribution had a relatively sharp peak, indicating that tension was distributed somewhat selectively to a particular digital tendon. As the point of loading moved progressively from radial to ulnar, the digital tendon receiving the greatest tension shifted stepwise from t2 to t5, and the overall OPI shifted progressively from more negative to more positive values (Fig. 7B). No broad peaks spanning two or more digits were observed with passive loading, however, indicating that despite their biomechanical interconnections, different radioulnar points on the proximal insertion aponeurosis transmit approximately half of the tension exerted at that point to a particular digital tendon.
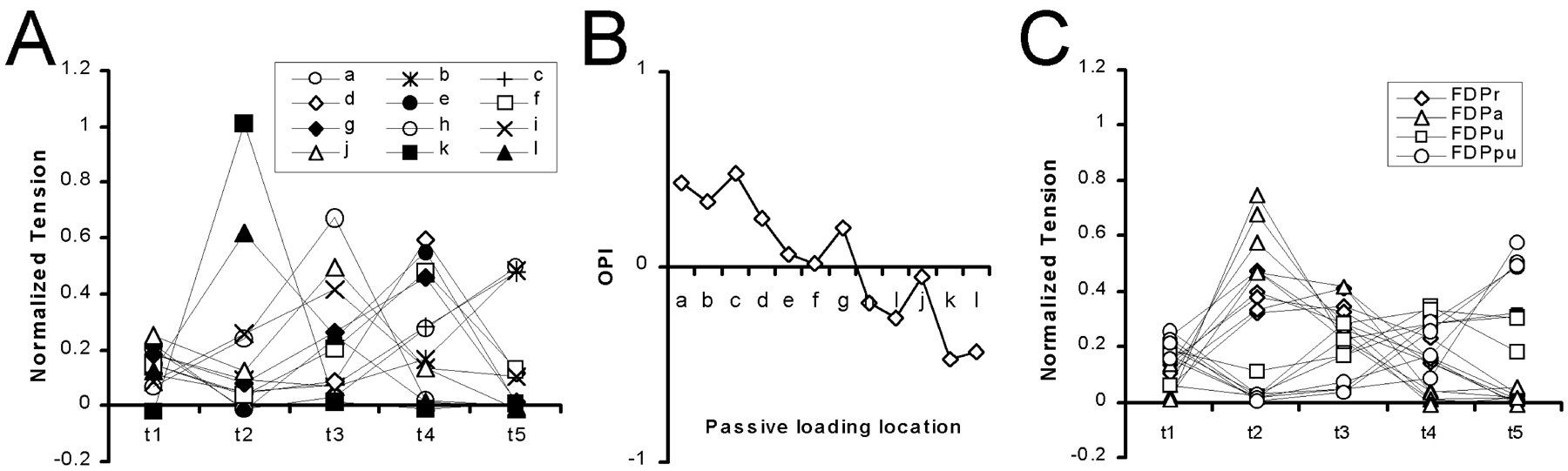
Tension distributions from passive loading versus active contraction. A, The distribution of tension, normalized such that the total on all five digits equals 1, has been plotted for each passive loading site. The SEL and OPI indexes for each of these distributions are shown in Figure 5B (+).B, As the site of passive loading gradually shifted from more ulnar (a) to more radial (l), the OPI appropriately shifted from more ulnar (positive) to more radial (negative) values. C, The normalized tension distributions from stimulation of each primary nerve branch in each experiment have been plotted for comparison with the normalized tension distributions from passive loading inA. The underlying data are the same as for Figure 5. Note that these distributions for active contraction of regions FDPr (⋄) and FDPu (■) are broader (less selective) than those for any point under passive load, as also indicated by the low SEL values in Figure 5B.
The relatively sharply peaked tension distributions produced by passive loading of the proximal insertion aponeurosis contrasted with the relatively broader tension distributions produced by active contraction of different regions of FDP, particularly FDPr and FDPu (Fig. 5). To compare these passive versus active tension distributions more directly, we normalized the tension produced by stimulation of each primary nerve branch in each experiment as a fraction of the total tension (first equation in Materials and Methods) and plotted these normalized active tension distributions in Figure 7C. The normalized active tension distributions of FDPa and FDPpu were rather selective for t2 and t5, respectively, and closely resembled the tension distributions produced by passive loading at the far radial and far ulnar aspects of the proximal insertion aponeurosis, respectively. In contrast, the normalized tension distributions produced by active contraction of FDPr or FDPu had less selective, broader peaks that spanned two or three adjacent digital tendons, unlike any seen with passive loading of a single point on the proximal insertion aponeurosis.
To examine these comparisons quantitatively, we computed the SEL and OPI for each tension distribution produced by passive loading (Fig.7A) and plotted them against one another along with those for active contraction of each region (Fig. 7C) in Figure5B. The SEL and OPI values produced by active contraction of FDPa were comparable with those produced by passive loading at the most radial aspect of the proximal insertion aponeurosis (except for passive loading at point k that produced tension only on t2, with OPI equaling −0.48 and SEL equaling 1.01), suggesting that FDPa exerted active force at this location, which then was passively distributed chiefly to t2. The SEL and OPI of FDPpu were comparable with those produced by passive loading at the most ulnar aspect of the proximal insertion aponeurosis, suggesting that FDPpu exerted active force at this location, which then was passively distributed chiefly to t5. In contrast, the SEL of FDPr tended to be slightly lower than that produced by passive loading with similar OPIs, and the SEL of FDPu tended to be considerably lower. These observations indicate that FDPr and FDPu each exert active tension at multiple radioulnar locations on the proximal insertion aponeurosis, consistent with the anatomical arrangement of their muscle fibers. The broad distribution of tension produced by contraction of FDPr or FDPu thus resulted both from active distribution of tension to multiple radioulnar locations across the proximal insertion aponeurosis and from passive distribution of this active tension through biomechanical interconnections between the tendons.
Excluding effects of spinal reflexes and axon reflexes
Electrical stimulation of a given primary nerve branch will excite both motor and sensory axons. Although discharge of the motor axons will cause the stimulated region to contract directly, centrally propagating discharge of sensory axons might elicit additional contraction through spinal reflexes. If sensory axons from one region of FDP have reflex effects on motoneurons innervating another region, then the distribution of active tension measured during stimulation of one primary nerve branch might represent both direct contraction of one region and reflex-induced contraction of another. Such spinal reflexes might play a role in tension distribution in the normal, behaving animal. To evaluate any contribution of spinal reflexes, we therefore repeated stimulation of each primary nerve branch in three monkeys after cutting the median and ulnar nerves in the upper arm, eliminating any possibility of spinal reflexes. Figure8A shows that, after proximal ligation of the median and ulnar nerves, stimulation of each nerve branch produced a tension distribution that was quite similar to that observed before ligation (Fig. 5A). The greatest amount of tension from stimulation of the FDPr nerve branch was on tendons 2 and 3, with less tension on tendons 1 and 4 and little or no tension on tendon 5. FDPu stimulation produced most tension on tendons 4 and 5, less tension on tendons 1 and 3, and the least tension on tendon 2. FDPa stimulation produced tension on tendon 2 and less on tendon 3. FDPpu produced tension on tendons 4 and 5. SEL and OPI values for each region were also similar to those observed before proximal nerve ligation (Fig. 8B). The similarity of tension distributions before and after proximal ligation of the median and ulnar nerves indicates that spinal reflexes did not cause appreciable contraction in additional neuromuscular regions that might have broadened the observed tension distributions.

Tension distributions after eliminating spinal reflexes. A, Plots of the tension exerted on each tendon during 20 Hz stimulation of each primary nerve branch after cutting the median and ulnar nerve trunks in the upper arm. Values are the average across subjects (n = 3) with SD error bars.B, The SEL and OPI indexes calculated for the tension distribution produced by 20 Hz stimulation of each primary nerve branch in each experiment have been plotted against one another. (Note that, because FDPpu received atypical innervation in 1 of the 3 monkeys studied here, only 2 points were available for this region.)
Motor axons commonly are assumed to ramify to innervate multiple muscle fibers only after the axon has entered the muscle. However, we wanted to exclude the possibility that axons innervating FDP give off collaterals in the parent median nerve, such that collaterals of the same axon might enter more than one primary nerve branch. Stimulating one primary nerve branch then would cause, via axon reflexes, contraction in more than one region, thereby producing a broader distribution of tension. Therefore, we stimulated each primary nerve branch in three monkeys a third time, after the branch had been cut away from the median or ulnar nerve trunk, eliminating any possible contribution to tension distribution of extramuscular axonal branching. The tension distribution observed for each region of FDP again was similar to that observed before cutting each primary nerve branch off of the parent nerve trunks, as well as before cutting the median and ulnar nerves in the upper arm (Fig.9A). SEL and OPI values for each region also were similar (Fig. 9B). Axon reflexes thus did not contribute appreciably to the distribution of tension produced by contraction of the regions of FDP.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Tension distributions after eliminating axon reflexes. A, Plots of the tension exerted on each tendon during 20 Hz stimulation of each primary nerve branch after cutting the branch off its nerve trunk. Values are the average across subjects (n = 3) with SD error bars. B, The SEL and OPI indexes calculated for the tension distribution produced by 20 Hz stimulation of each primary nerve branch in each experiment have been plotted against one another. (Note that, because FDPpu received atypical innervation in 1 of the 3 monkeys studied here, and in another the FDPpu nerve branch failed to conduct after being cutoff the ulnar nerve trunk, only 1 point was available for this region.)
DISCUSSION
Factors distributing tension to multiple digits
Our results show that none of the four neuromuscular regions of the macaque FDP exerts tension on only one digit. Each region does distribute tension across the digits in a distinct pattern, however. The two largest regions (FDPr and FDPu) show the broadest tension distributions. FDPr exerts most of its tension on digits 2 and 3, but also exerts considerable tension on digits 1 and 4; FDPu exerts most of its tension on digits 4 and 5, but also exerts considerable tension on digits 1 and 3. The action of FDPr on digit 4 and the action of FDPu on digit 1, although consistent with the anatomic structure of the FDP tendon sheet (Serlin and Schieber, 1993), could not have been predicted from our previous EMG and modeling studies (Schieber, 1993, 1995). The two smaller regions (FDPa and FDPpu) each distribute their tension more selectively. FDPa exerts most of its tension on digit 2, but also exerts some tension on digits 1 and 3; FDPu exerts most of its tension on digit 5, but also exerts some tension on digits 1 and 4.
In large part, the distribution of tension to multiple digits by each region of the macaque FDP results from the complex, heavily interconnected structure of its tendons of insertion. Passive loading at different radioulnar points on the proximal portion of this tendon sheet always produced maximal tension on one distal finger tendon, with lesser degrees of tension on other, adjacent tendons. In most cases, some tension also was transmitted to the thumb. Indeed, the patterns of tension distribution produced by passive loading at the radial or ulnar edge of the proximal insertion aponeurosis closely resembled those produced by active contraction of FDPa or FDPpu, respectively, suggesting that these two regions exert their respective active tensions primarily at the radial or ulnar edges of the proximal insertion aponeurosis, and that tension then is distributed to multiple digits passively through the biomechanical interconnections of the tendons.
The two larger regions (FDPr and FDPu) each distributed their tension more broadly than could be attributed to passive mechanical distribution of tension from a single point on the proximal insertion aponeurosis. Although these broader tension distributions most likely reflect the insertion of FDPr and FDPu muscle fibers on more extensive radioulnar portions of the proximal insertion aponeurosis, we considered two alternative explanations. First, although the reflex connections of afferents typically are partitioned to affect motoneurons of the compartment from which the afferent originated (Windhorst et al., 1989), Iaafferents from a given region of the feline biceps femoris or FDP have been shown to make additional connections to motoneurons of other regions of the same muscle (Botterman et al., 1983; Fritz et al., 1989). Therefore, we considered the possibility that, if afferent axons were stimulated along with motor axons in a given primary nerve branch, spinal reflexes might result in contraction of other regions of FDP, broadening the distribution of recorded tension. We consistently found, however, that the tension distribution remained unchanged after the median and ulnar nerves were cut proximally to eliminate any effects of spinal reflexes. Second, we considered the possibility that motor axons might bifurcate proximal to formation of the primary nerve branches from the median nerve, such that branches of the same motor axons might enter more than one primary nerve branch. Then axon reflexes might result in contraction in more than one region after stimulation of a single primary nerve branch. We found, however, that tension distributions did not change when each primary nerve branch was cut off the median or ulnar nerve trunk to eliminate any axon reflexes. Therefore, we conclude that the broad tension distributions produced by contraction of FDPr or FDPu result in part from the fact that each of these regions exerts active tension on a broad portion of the proximal insertion aponeurosis.
Compartmentalization of FDP
The macaque FDP shows many characteristics of a compartmentalized muscle. Each region of the macaque FDP is innervated by a primary nerve branch. Although glycogen depletion studies have yet to be performed on this muscle, the present EMG recordings indicate that the vast majority of the muscle fibers innervated by axons in each primary nerve branch are localized within the appropriate region. In microdissection, a clear border can be defined between FDPr and FDPu, with fascicles of FDPr arising radial to the anterior interosseous nerve and fascicles of FDPu arising ulnar to the nerve (Serlin and Schieber, 1993). The superficial insertion aponeurosis separates FDPa from the rest of the muscle. A clear anatomical border between regions FDPpu and FDPu is not evident on microdissection, and in the present study the compound muscle action potentials evoked by stimulation of the FDPpu and FDPu primary nerve branches were not sharply localized. Therefore, this border might be represented by interdigitated fascicles of the two adjacent regions. Our previous EMG recordings from FDPr and FDPu in awake behaving monkeys indicate that these two regions each can be activated independently of one another, whereas EMG recordings from FDPa and FDPpu have yet to be obtained (Schieber, 1993, 1995). Many features of the macaque FDP thus indicate that its four regions are in fact separate neuromuscular compartments.
Compartmentalization often is most apparent in muscles with spatially distributed origins and/or insertions. Although the different compartments of such a muscle typically have similar histochemical fiber-type composition, their different origins and/or insertions give contraction of each compartment a different biomechanical effect (Chanaud et al., 1991). Similarly, the macaque FDP has a distributed origin (from the internal aspects of both the radius and the ulna, from the interosseous membrane, and from the medial epicondyle of the humerus), as well as a spatially distributed insertion, with distal tendons inserting separately on each digit. We have shown previously that all four compartments of the macaque FDP have similar fiber-type composition (Maurer et al., 1995). The present results demonstrate that contraction of each compartment also has a distinct mechanical effect, producing a unique distribution of tension across the five tendons of the muscle. None of these compartments exerts tension on only a single tendon, so none can produce motion of only a single digit. Nevertheless, various levels of contraction in the different compartments of FDP could be combined with the action of additional finger muscles to produce more movement of one digit than the others, as observed during individuated finger movements performed by awake monkeys (Schieber, 1991, 1995).
Footnotes
- Received October 20, 2000.
- Revision received December 27, 2000.
- Accepted January 4, 2000.
This work was supported by Grant R01-NS36341 from the National Institute of Neurological Disorders and Stroke. We thank Lee Anne Schery for technical assistance and Marsha Hayles for editorial comments.
Correspondence should be addressed to Dr. Marc H. Schieber, University of Rochester Medical Center, Department of Neurology, 601 Elmwood Avenue, Box 673, Rochester, NY 14642. E-mail: mhs{at}cvs.rochester.edu.
- Copyright © 2001 Society for Neuroscience