
Abstract
Autonomic and limbic information is integrated within the lateral hypothalamus (LH), and excitability of LH neurons is important in the control of feeding and behavioral arousal. Despite the prominent expression of P2X-type ATP receptors throughout the hypothalamus, the role of ATP in LH excitability is not known. Perforated-patch-clamp recordings of synaptically coupled neurons from both embryonic chick and postnatal mouse lateral hypothalamus in vitro reveal robust stimulus-evoked purinergic synaptic transmission. Suprathreshold activation elicits reliable and concurrent release of ATP with GABA. Tetrodotoxin-resistant P2X receptor-mediated events are readily observed at LH synapses from the embryonic chick, whereas GABA miniature postsynaptic currents (mPSCs) are recorded in innervated LH neurons from either embryonic chicks or postnatal mice. Two distinct mPSCs are recorded at ATP–GABA cosynapses; one has a monoexponential decay phase and is modulated by flunitrazepam, and the other has a decay phase that is best fit by a sum of two exponential functions (τfast and τslow), and only the τslow component is affected by flunitrazepam. Bicuculline does not completely inhibit all mPSCs. The remaining bicuculline-resistant mPSCs are blocked by suramin, and their decay phase is briefer than that of GABAergic mPSCs. Furthermore, at a holding potential intermediate for the reversal potentials of GABAA and P2X receptors, little or no current is observed, consistent with concomitant release (and detection) of GABA and ATP. Together, our data suggest that a subset of spontaneous and evoked PSCs arise from the concurrent activation of both GABAA and P2X receptors.
- cotransmission
- feeding
- P2X receptor
- plasticity
- lateral hypothalamus
- ATP
- GABA
- GABAA receptor
The lateral hypothalamus (LH) is an important center for the integration of autonomic and limbic information. The LH is implicated in the modulation of visceral motor and sensory pathways, including those underlying feeding (Bernardis and Bellinger, 1996) and behavioral arousal (Chemelli et al., 1999; Hara et al., 2001; Willie et al., 2001). Animal models with lesions of the lateral hypothalamic area (LHA) exhibit hypophagia, increased metabolic rate, and decreased arousal, often resulting in death by starvation (Willie et al., 2001). The LHA has classically been regarded as an important component of the autonomic control of feeding behavior, with extensive projections both within the hypothalamus per se and throughout the neuroaxis.
The hypothalamus is a prominent region of P2X-type ATP receptor expression (Xiang et al., 1998; Kanjhan et al., 1999) and, as such, might constitute a region of robust purinergic synaptic transmission. Several of the ATP receptor subunits cloned (Brake et al., 1994; Valera et al., 1994; Chen et al., 1995; Buell et al., 1996; Collo et al., 1996; Surprenant et al., 1996) are expressed in specific brain regions, including the LH. Mammalian CNS neurons express P2X4, P2X6, and to a lesser degree, P2X2-type purinergic receptors (Buell et al., 1996; Collo et al., 1996; Kanjhan et al., 1999). A novel ATP P2X receptor (P2X8) is expressed in embryonic chick brain (Bo et al., 2000).
Despite the prominent expression of ATP receptor subunits in many regions of the CNS (Collo et al., 1996; Kanjhan et al., 1999; Norenberg and Illes, 2000), relatively little is known about purinergic transmission in the brain. This is primarily because of the difficulties inherent in the reliable identification of small-amplitude synaptic currents and the lack of highly selective ATP P2X receptor antagonists. So far, ATP P2X receptor-mediated transmission has been detected as a minor component of synaptic activity in the mammalian habenula (Edwards et al., 1992) and hippocampus (Pankratov et al., 1998). In contrast, studies of peripheral (autonomic) neurons and of spinal cord dorsal horn neurons suggest a prominent role of ATP as a cotransmitter with norepinephrine, acetylcholine, or GABA (Burnstock, 1999; Jo and Schlichter, 1999). The net effect of ATP at such cotransmitting sites is difficult to evaluate and may also be complicated in view of recent reports demonstrating cross-inhibition of ATP P2X receptors with both nicotinic acetylcholine receptors (Khakh et al., 2000) and GABAA receptors (Sokolova et al., 2001).
Our initial aim was to ascertain whether ATP P2X receptor expression in the hypothalamus underlies a significant contribution of ATP to synaptic transmission. To test this idea, we examined 139 purinergic synapses. We subsequently examined 72 synapses at which we detected purinergic and GABAergic cotransmission. We found that a subset of miniature postsynaptic currents (mPSCs) is composed of both GABAA and P2X receptors and that ATP and GABA may be costored within the same synaptic vesicle. ATP and GABA cotransmission could play an important role in the control of excitability of LH neurons at early stages of development, when GABA exerts a depolarizing effect in hypothalamic neurons.
MATERIALS AND METHODS
Neuronal cultures. Primary cultures of lateral hypothalamic neurons were prepared from embryonic day 11 chicks or from postnatal day 3 mice using the following basic protocols. The region of the hypothalamus was identified on the ventral aspect of the brain with the exterior border delineated by the optic chiasm and the posterior border delineated by the infundibular stalk. The most lateral portions of the demarcated area were excised, microdissected, and incubated at 37°C in divalent cation-free Earle's balanced salt solution (EBSS; Invitrogen, San Diego, CA) containing papain (20 U/ml; Sigma, St. Louis, MO) with l-cysteine (1 mm; Sigma). After 20–30 min of incubation, the tissue was mechanically dispersed by repeated passage through a 10 ml plastic pipette. Papain activity was stopped by adding 3 ml of EBSS containing bovine serum albumin (1 mg/ml; Sigma) and DNase (0.01%; Sigma), and a mechanical dissociation was performed with a 1 ml plastic pipette. The homogenate was deposited on top of 4 ml of a solution with a composition similar to that described above, except that the concentration of bovine serum albumin was increased to 10 mg/ml. After centrifugation (5 min at 1000 rpm), the preparation for plating was collected by removing the supernatant and resuspending the neurons in complete medium composed of DMEM (Invitrogen), heat-inactivated horse serum (10% v/v; Invitrogen), chick extract (10% v/v, house-made), penicillin and streptomycin (50 IU/ml for each; Invitrogen), and nerve growth factor (10 nm). The equivalent “complete medium” used for preparation and plating of LH neurons from postnatal mice was MEM-α (Invitrogen), horse serum (5% v/v), fetal calf serum (5% v/v), putrescine (100 nm), transferrin (10 mg/ml), insulin (5 mg/ml), penicillin and streptomycin (50 IU/ml for each; Invitrogen), and nerve growth factor (10 nm). LH neuron preparations obtained from ∼12 embryonic day 11 chick brains were plated on six 35 mm poly-l-ornithine-coated tissue-culture plastic dishes and maintained until use in a water-saturated atmosphere (95% air and 5% CO2) at 37°C. LH neuron preparations obtained from ∼12 postnatal mouse brains were plated on six modified 35 mm tissue-culture plastic dishes. Each dish has a chamber ∼10 mm in diameter with a plastic ring on the bottom. As with the chick neurons, the preferred substrate was poly-l-ornithine.
Electrophysiological recording. Electrophysiological recordings from either chick or mouse neurons in vitrowere conducted between 7 and 15 d after plating. Membrane currents were recorded at room temperature (20–22°C) with an Axopatch 200B amplifier (Axon Instruments, Foster City, CA) in the perforated patch-clamp configuration using amphotericin B. The external solution contained (in mm): NaCl 135, KCl 5, CaCl2 2.5, MgCl2 1, HEPES 5, and glucose 10, pH 7.3. In all experiments, 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX) (10 μm),dl-aminophosphonovaleric acid (dl-APV, 50 μm) and strychnine (1 μm) (CAS) were continuously present in the external solution. The amphotericin B (Sigma) stock solution (30 mg/ml) was prepared in dimethylsulfoxide just before the recording session. The pipette was first filled at the tip with internal solution containing (in mm): CsCl (or KCl) 150, HEPES 10, pH 7.3, and subsequently back-filled with the same solution containing amphotericin B (150 μg/ml). Under these conditions, the equilibrium potential for Cl− ions (ECl) was ∼0 mV. To set ECl to −70 mV, we replaced 141 mm CsCl or KCl with 70 mmCs2SO4 or 141 mm K-acetate, respectively, in the pipette solution. Adjustment of ECl to −70 mV and Ecations to 0 mV allowed recording of cation-mediated components of synaptic transmission at a holding potential (VH) of −70 mV and recording of the Cl− ion-mediated component at a VH of 0 mV.
Eliciting monosynaptic evoked PSCs and detection of mPSCs.To evoke PSCs, simultaneous perforated-patch-clamp recordings were obtained from synaptically connected neurons using two independent Axopatch 200B amplifiers. The presynaptic neuron was held in the current-clamp configuration at the resting membrane potential, and depolarizing current pulses were applied at a frequency of 0.1 Hz. For extracellular stimulations, a double-barreled electrode filled with extracellular solution was placed in contact with the cell body of a visually identified neuron. Stimulation was performed with short single stimuli or pairs of stimuli [interval: 300 msec for GABAergic evoked IPSCs (eIPSCs), 150 msec for purinergic evoked EPSCs (eEPSCs); 0.1 msec; −20 and −100 μA] delivered at 0.1 Hz. Our typical protocol for isolation of purinergic eEPSCs and mPSCs involved setting ECl to −70 mV (as described above) and recording at a VH of −70 mV in the continuous presence of CNQX, dl-APV, strychnine, and bicuculline. Exceptions to this standard protocol are noted throughout the text. Likewise, GABAergic eIPSCs were detected by setting ECl to −70 mV and recording at aVH of 0 mV in the continuous presence of CAS. Individual spontaneously occurring mPSCs were analyzed off-line using two commercially available softwares [Mini analysis 5.0 from Synaptosoft Inc. (Decatur, GA) or Axograph 4.0 from Axon Instruments]. For each experiment, the threshold for detection was set to >5 pA, except for the experiments performed at aVH of −35. Under these circumstances, recordings were also visually inspected to minimize data loss despite the very small amplitude of the events. For analysis of the decay phase of mPSCs, the events were selected on the basis of the following criteria: (1) that we were able to obtain a stable baseline recording both before and after the events were detected; (2) a minimum interval of >100 msec between events; and (3) rise times of events that were <3 msec (10–90% of the peak amplitude of the mPSCs). For examination of average mPSC profiles, the events were aligned by their initial rising phase using Mini analysis 5.0. Voltage and current traces were stored on a videotape recorder and/or the hard drive of the analysis computer after being filtered at 5 kHz by Axopatch 200B. Acquisition and analysis were performed using pClamp6, Axograph 4.0 (Axon Instruments) and Mini analysis 5.0. Student's t tests were used to analyze the difference between parameters. The critical value for statistical significance was set at p < 0.05. All statistical results are given as mean ± SEM.
Immunocytochemical staining. LH neurons in vitrofrom postnatal mice were fixed for 30 min at room temperature in 4% paraformaldehyde in 0.1 m phosphate buffer, pH 7.4, and subsequently rinsed three times in PBS. Cultures were subsequently permeabilized with 0.5% Triton X-100 for 5 min and incubated overnight at room temperature with a mouse monoclonal antibody against glutamic acid decarboxylase (GAD)-6 (1:200; a gift from Dr. David I. Gottlieb, Washington University, St. Louis, MO) and a rabbit polyclonal antibody against ATP P2X4 receptor subunit (Alomone Labs, Jerusalem, Israel). Cultures were rinsed three times with PBS and incubated for 1 hr at room temperature with a fluorescein isothiocyanate-conjugated anti-mouse IgG (1:200) to reveal GAD-like immunoreactivity and a rhodamine-conjugated anti-rabbit IgG (1:400) to reveal ATP P2X4 receptor subunit-like immunoreactivity.
Preparation and application of drugs and other reagents. Most reagents were prepared as 1000× concentrated stock solutions. Bicuculline methiodide, strychnine, tetrodotoxin (TTX), (1S,9R)-β-hydrastine (all Sigma), pyridoxalphosphosphate-6-azophenyl-2′,4′-disulfonic acid (PPADS), and ARL67156 (both from Research Biochemicals, Natick, MA) were prepared in distilled water and stored at −20°C.dl-APV (Sigma) was prepared in 100 mm NaOH solution. CNQX (Tocris Cookson, Ballwin, MO) was prepared in dimethylsulfoxide and stored at 4°C. Flunitrazepam (Sigma) was prepared as 10,000× concentrated stock solution in dimethylsulfoxide. The substances to be tested were diluted to the final concentration with extracellular solution just before the recording session. Substances were bath-applied at a flow rate of 2–3 ml/min. For exogenous application of GABA and ATP, a 1.5-mm-diameter U tube controlled by a solenoid valve was used. The threshold for responses was set to >5 pA.
RESULTS
Evoked ATP P2X receptor-mediated synaptic transmission among the embryonic chick and mouse LH neurons
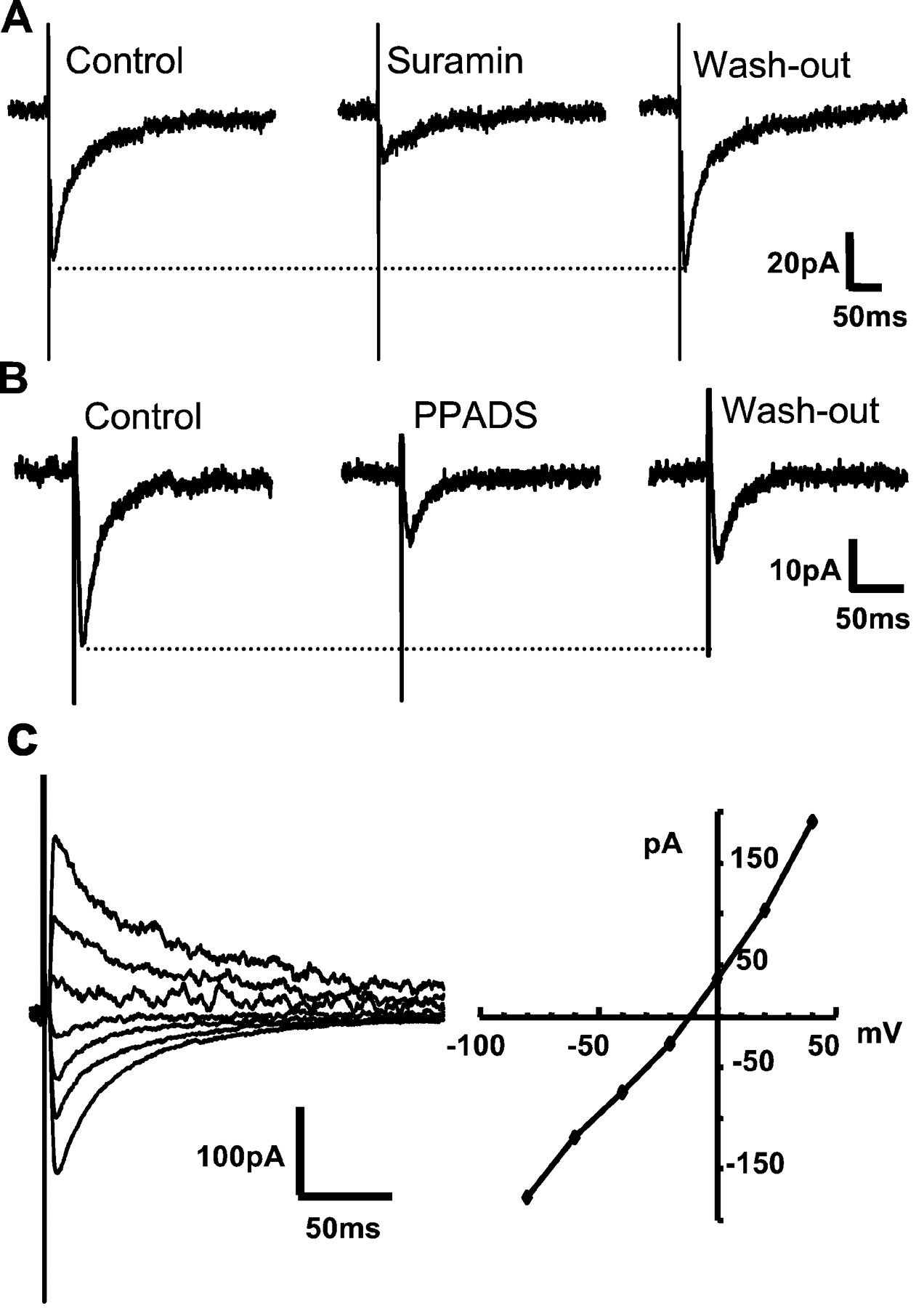
To examine stimulus-evoked release of ATP in isolation from other fast transmitter responses, we first tested for synaptic transmission using focal extracellular stimulation of potential presynaptic partners selected on the basis of the apparent physical contact with the recorded neuron. Such assays were conducted in the continued presence of antagonists of ionotropic glutamate, glycine, and GABAA receptors (in μm: CNQX 10, APV 50, strychnine 1, bicuculline 10; ECl = −70 mV; see Materials and Methods). Stimulation of presynaptic inputs elicited postsynaptic inward currents of approximately −50 pA in amplitude at a VH of −70 mV (Fig.1A–C) (mean amplitude, −51.2 ± 14 pA; range, −6 to −398.4 pA; n = 31 cells). Although the magnitude of such EPSCs is small, evoked responses were reliably detected with stimulation frequencies of 0.1 Hz for up to 30 min. The eEPSCs were reversibly but not completely inhibited by suramin (Fig. 1A) and by PPADS (Fig.1B). The incomplete inhibition of P2X receptors by either suramin or PPADS is consistent with previous studies of native receptors in the CNS (Edwards et al., 1992; Bardoni et al., 1997;Pankratov et al., 1998; Mori et al., 2001). The incomplete blockade could be in part a result of the expression of heteromeric P2X channels, including P2X4 and P2X6 receptor subunits (Le et al., 1998), that are insensitive to suramin and PPADS (Buell et al., 1996; Collo et al., 1996).

Stimulus-evoked transmission mediated by the activation of ATP P2X receptors at embryonic chick LH synapsesin vitro. A, B, Extracellular stimulation of chick LH neurons in the presence of CNQX, APV, strychnine, and bicuculline (10, 50, 1, and 10 μm, respectively; ECl = VH = −70 mV) elicits inward currents that are blocked by suramin (30 μm; mean inhibition, 71.2 ± 8%;n = 10 cells) and by PPADS (50 μm; mean inhibition, 31.4 ± 2%; n = 4 cells). The majority of evoked synaptic currents decay in a manner that is best fit by the sum of two exponential functions (τfast = 9.4 ± 1 msec; τslow = 47.2 ± 3.8 msec;n = 16 of 26 cells); the remainder was best fit by a single exponential function (τ = 19.2 ± 2 msec;n = 10 of 26 cells). The dotted lines indicate the control level of the evoked EPSCs.C, The I–V relationship of the peak amplitude of evoked synaptic currents shows that the mean reversal potential is −14.6 ± 2 mV (inset;n = 3 cells; ECl = −70 mV), consistent with the activation of a relatively nonselective cationic conductance. The current traces shown correspond toVHs between −80 and +40 mV in 20 mV increments.
The synaptic currents evoked in the continued presence of antagonists of glutamate, glycine, and GABAA receptors have nearly linear current–voltage (I–V) curves, with clear outward currents recorded at the most positive potentials (Fig.1C). These data are consistent with the previous reports of stimulus-evoked purinergic transmission in the rat medial habenula (Edwards et al., 1997) and spinal cord (Jo and Schlichter, 1999).
Evoked purinergic transmission is readily detected in neuronal preparations from postnatal mouse and embryonic chick (compare Figs. 1and 2). Under the same experimental conditions as outlined above, extracellular stimulation of LH neurons from postnatal mouse in vitro also elicited eEPSCs that are reversibly but not completely blocked by suramin and PPADS (Fig.2A,B). Mecamylamine (1 μm), a nicotinic acetylcholine receptor antagonist, had no effect on the amplitude of eEPSCs (data not shown). The mean amplitude, decay time constants, and pharmacology of purinergic EPSCs at mouse LH synapses are equivalent to those observed in in vitro preparations of embryonic chick (Table 1). TheI–V relationship for P2X receptor-mediated currents in mice is similar to chicks (data not shown). Together, these experiments demonstrate robust P2X receptor-mediated purinergic transmission atin vitro synapses from embryonic chick or neonatal mouse LH.

Stimulus-evoked transmission mediated by the activation of ATP P2X receptors at postnatal mouse LH synapsesin vitro. A, B, top panels, Recordings from postnatal mouse LH neurons under the same conditions as described in Figure 1. The eEPSCs are inhibited by suramin (30 μm) and by PPADS (50 μm), consistent with the eEPSCs being mediated by P2X receptor activation. The dotted lines indicate the control level of evoked EPSCs.A, B, bottom panels, The normalized amplitudes of the eEPSC peaks from neurons shown in Aand B (top) are plotted as a function of time. The solid line marks the control (baseline) level of eEPSCs. Solid bar, Time of suramin or PPADS application, as noted. C, Summary of inhibition of eEPSCs by P2X receptor antagonists in chicks (14 neurons) and mice (5 neurons).
Comparison of the amplitudes, decay time constants, and antagonist profile of P2X receptor-mediated evoked synaptic currents recorded in embryonic chick versus neonatal mouse LH neuronsin vitro
Spontaneous ATP P2X receptor-mediated synaptic transmission within embryonic chick LH neurons
Recordings from embryonic chick LH neurons in vitro in the presence of TTX (1 μm) revealed that ∼70% of neurons received spontaneous excitatory synaptic input that was resistant to a mixture of glutamate receptor antagonists (in μm: CNQX 10, APV 50, TTX 1;VH = ECl = −70 mV; n = 16) (Fig. 3). The amplitude of these TTX-resistant miniature EPSCs (mEPSCs) ranged from −5.6 to −15.8 pA, with a mean of −9.3 ± 0.9 pA (n = 11 neurons) (Fig. 3A). The decay time course of mEPSCs can be fit by a sum of two exponential functions (τfast, 4.8 ± 0.9 msec; τslow, 24.3 ± 3.3 msec; n= 8 cells). Both suramin and PPADS reversibly inhibited the majority of glutamate receptor-independent mEPSCs (Fig. 3B,C). We propose that this population of fast mEPSCs is attributable to the spontaneous release of ATP and consequent activation of postsynaptic ATP P2X receptors. Unlike embryonic chick LH cultures, recordings in postnatal mouse LH were devoid of spontaneous EPSCs, despite reliable detection of evoked purinergic eEPSCs. The latter results are consistent with other studies of postnatal purinergic synapses (Jo and Schlichter, 1999).

TTX-resistant spontaneous purinergic transmission at embryonic chick LH neuron synapses in vitro.A, Left, Current tracesshowing mEPSCs recorded in the continuous presence of TTX, CNQX, and APV (1, 10, and 50 μm, respectively;VH = ECl = −70 mV).Right, Averaged trace of P2X receptor-mediated events. Bottom, Histograms show the distribution of mEPSC amplitudes and decay time constants from a representative recording. B1, C1, The TTX-, CNQX-, and APV-resistant mEPSCs were reversibly blocked by suramin and PPADS (30 μm, n = 10; 50 μm, n = 4, respectively). B2, C2, The frequency (number of events per 10 and 30 sec, respectively) of mEPSCs shown in B1 andC1 is plotted versus time. Solid barscorrespond to the time of suramin or PPADS application, as indicated.
Corelease of ATP and GABA from individual presynaptic neurons
Although ATP has been shown to act as a cotransmitter in the PNS and in spinal cord (Burnstock, 1999; Jo and Schlichter, 1999), ATP cotransmission has been difficult to study at supraspinal synapses. Reports to date suggest that ATP is released alone, independent of other synaptic transmitters in the medial habenula (Robertson and Edwards, 1998).
Rigorous proof of ATP and GABA corelease is provided by concurrent recordings from identified presynaptic and postsynaptic partners. Such dual-perforated patch-clamp recordings from embryonic chick LH neuronsin vitro are shown in Figure4. In the presence of antagonists of ionotropic glutamate and glycine receptors, single presynaptic action potentials evoke rapidly rising unitary postsynaptic currents. The brief and constant latency (2.6 ± 0.4 msec; n = 8) as well as the absence of failures indicates that evoked PSCs are monosynaptic in origin (Fig. 4A1). We assessed the relative contributions of ATP P2X and GABAAreceptors to the ePSCs by dual patch-clamp recording from synaptic pairs under control conditions and with the sequential addition of bicuculline and suramin. In this series of experiments, we set the ECl of the postsynaptic neuron to 0 mV and recorded at a VH of −70 mV. Under these conditions, the evoked currents were largely blocked by bicuculline, and the remaining currents were significantly inhibited by suramin (Fig. 4A2,B) (n = 5 of 5 pairs tested).

Concurrent perforated patch-clamp recordings from connected pairs of presynaptic and postsynaptic LH neurons demonstrate purinergic and GABAergic cotransmission. A1, Schematic diagram of recording configuration and representative recordings.A–C, Dual perforated-patch-clamp recordings from synaptically coupled pairs of neurons show that single presynaptic action potentials induced by injecting positive currents (87.5 msec; 100, 150, and 300 pA, respectively) evoke postsynaptic currents in the presence of CAS. The resting membrane potential of the presynaptic neuron was −50 mV. A2, A3, Postsynaptic currents (i) evoked by single presynaptic action potentials induced by injecting positive currents (5 msec; 300 pA) in the presence of CAS were partially inhibited by bicuculline (ii; Bicu; 10 μm). These bicuculline-resistant, stimulus-evoked synaptic currents were inhibited by the addition of the P2X receptor blocker, suramin (30 μm). Superimposition of i andii shows the difference in the synaptic current decay time course of the GABAergic and/or purinergic components (A3). B, Left, Representative recordings of GABAergic and purinergic eEPSCs.Right, The normalized eEPSC amplitude is plotted as a function of time.Solid bars, Time of application of bicuculline (10 μm) with or without suramin (30 μm), as noted. a–c (left) are sample traces from the experiment shown in B (right). Solid lines indicate the duration of application of the antagonists.C, Stimulation of the presynaptic neuron of another synaptically coupled pair of LH neurons in vitro reveals both the inward purinergic currents and outward GABAAreceptor-mediated responses, depending on theVH of the postsynaptic neuron (VH = −70 mV andVH = 0 mV, respectively).
Additional examination of postsynaptic currents evoked by single action potentials in individual presynaptic neurons revealed both inward and outward currents, depending on the VHof the postsynaptic neuron relative to ECl (Fig.4C). Thus, with the postsynaptic neuron membrane potential clamped at −70 mV (close to ECl), we recorded inward P2X receptor-mediated currents. Clamping the same postsynaptic neuron of the synaptic pair to VH of 0 mV elicited outward GABAA receptor-mediated currents (the reversal potential, VREV, for ATP P2X receptor-mediated currents is approximately Ecations = 0 mV; n = 3 of 3 pairs tested in this manner). These data indicate that the recorded currents are attributable to action potential-dependent, synaptic release of both ATP and GABA from the same presynaptic neuron. The paired-recording experiments also revealed that a key feature of GABA and ATP cotransmitting neurons was their strong accommodation to direct depolarization (i.e., a single action potential rather than a burst of action potentials with a prolonged depolarizing pulse; data not shown).
Similar assays of ATP and GABA cotransmission in mouse LH neuronsin vitro confirmed and extended the above studies (Fig.5). When postsynaptic neurons were held at a VH of −70 mV (close to ECl), stimulation of the presynaptic neuron elicited bicuculline- and (+)-β-hydrastine-resistant, inwardly directed eEPSCs in the presence of ionotropic glutamate, glycine, and GABAA receptor antagonists (Fig. 5A). Stimulation of the presynaptic input to the same cell, held at 0 mV, elicited no current. Subsequent removal of the GABAA receptor antagonists led to recovery of the outward eIPSCs mediated by GABAA receptors. The double immunocytochemical staining with antibodies against P2X4 receptor subunit and GAD-6 also supports the potential overlap of a subset of GABAergic synapses with regions of P2X receptor expression in vitro (Fig. 5B). As shown in Figure 5, mouse LH neurons in vitro were stained with anti-P2X4 receptor subunit. P2X4 receptor subunit-like immunoreactivity was readily observed on a subset of neuronal somata and occasionally on neurites. GAD-like immunoreactivity was observed in many neurites, indicating regions of GABA-containing synapses. GAD-positive neurites made contacts with a P2X4receptor subunit-positive somatic area and neurites, consistent with P2X receptor-positive neurons receiving GABAergic inputs. These immunocytochemical data are consistent with observed monosynaptic PSCs proposed to include both ATP P2X and GABAAreceptor-mediated components.

Corelease of ATP and GABA at postnatal mouse LH synapses in vitro. A, Schematic diagram of a recording configuration and representative recordings of synaptic currents evoked by extracellular stimulation of a presynaptic input to a mouse LH neuron in vitro. At aVH of −70 mV, close to ECl, in the presence of CAS and (+)-β-hydrastine (10 μm) (a GABAA receptor antagonist), extracellular stimulation elicits eEPSCs in contacted mouse LH neurons. While the same cell was held at Ecations (close to 0 mV), we observed no current. After washout of (+)-β-hydrastine from the recording solution, the eIPSCs return, consistent with their being mediated by GABAA receptors. B, Double immunostaining with antibodies against GAD-6 and P2X4receptor subunit. GAD-like immunoreactivity is observed on neurites (green). P2X4 receptor subunit-like immunoreactivity is observed on soma and occasionally on neurites (red). Note GAD-positive neurites contacting P2X4 receptor subunit-positive somata and neurites (yellow arrow). The green arrowindicates GAD-positive neurons that appear to contact regions that lack P2X4-like immunoreactivity. Scale bar, 20 μm.
A minority of synaptic pairs appeared to be solely GABAergic in the embryonic chick preparation (30%; n = 16 of 52 pairs). In contrast, we were unable to elicit purinergic currents without concomitant GABAergic responses at any of the 52 synaptic pairs examined (i.e., we found no “ATP-only” synapses). In parallel studies assaying the effects of exogenous application of GABA and ATP (50 and 30 μm, respectively) (Fig.6), we found that applied GABA elicited robust responses, whereas ATP was without effect [approximately one-third of the neurons tested were “GABA only” (5 of 13 neurons)]. Most neurons (∼60%) assayed by exogenous application responded to both GABA and ATP (Fig. 6) in the embryonic chick preparation. The average amplitudes of the responses to GABA and ATP were −904 ± 312 pA (n = 13) and −164 ± 60 pA (n = 8) at a VH of −60 mV, respectively. The mean percentage inhibition of the response to ATP by suramin was 70 ± 3%. Bicuculline completely blocked the response to GABA (98.6 ± 1% inhibition; n = 5). Similar results were obtained for mice.

GABA- and ATP-induced responses in embryonic chick LH neurons. A, Approximately 60% of neurons tested responded to both GABA (50 μm) and ATP (30 μm). B, Approximately 30% of neurons tested were responsive to GABA but not to ATP (n = 13).
GABAA and P2X receptor-mediated components in spontaneous mPSCs
We subsequently examined whether individual mPSCs are composed of GABAA and/or P2X receptor-mediated components. We approached this question in two series of experiments. First, we analyzed individual mPSCs at synapses receiving both purinergic and GABAergic mPSCs (i.e., synapses at which mPSCs were blocked only by combined GABAA and P2X receptor antagonists.). Under these conditions, we observed two distinct types of mPSCs: one has a decay time course that is well fit by the single exponential function (τ = 35 ± 1 msec) and the other has a decay phase that is best fit by the sum of two exponential functions (i.e., “mixed” mPSCs; τfast = 7 ± 0.5 msec; τslow = 61.6 ± 3 msec) (Fig.7A). We speculated that mixed mPSCs may be composed of both P2X and GABAAreceptor-mediated components, because activation of two kinetically distinct ligand-gated ion channels could yield a composite current with a decay time course best fit by (at least) two exponential functions. Indeed, mixed mPSCs have significantly greater peak amplitudes than those with a monoexponential decay phase (−34.5 ± 1.3 vs −14.7 ± 0.8 pA) (Fig. 7A2). Additional evidence for the mixed nature of these mPSCs is the incomplete block by bicuculline. The remaining bicuculline-resistant mPSCs were inhibited by suramin (30 μm), and the decay phase of the currents was well fit by a single exponential function. The decay phase of these suramin-sensitive mPSCs is significantly briefer than those with a monoexponential decay phase, observed in the absence of GABAA receptor antagonist (τ = 24 ± 3 msec vs 35 ± 1 msec) (Fig. 7B). Such observations suggest that the mixed mPSCs would be a result of the concurrent activation of both P2X and GABAA receptors.

GABAA and P2X receptor-mediated components in spontaneous miniature PSCs. A1, Two distinct populations of mPSCs are detected in recordings at the ATP–GABA cosynapse. Left, Superimposition of average oftraces (>200 events) and individual events, showing two kinetically distinct events. Right, Average amplitude of events classified according to the decay rate(s) of mPSCs.A2, Mixed mPSCs have a significantly greater mean peak amplitude than those with a monoexponential (Monoexp.) decay phase (−34.5 ± 1 vs −14.7 ± 1 pA;p < 0.05). A3, Plot of the percentage contribution of monoexponential decay mPSCs versus mixed-decay mPSCs to recorded spontaneous activity (mixed component, 72%; monoexponential component, 28%).B1, Top, Averaged tracesof bicuculline (Bicu)-sensitive (a) and bicuculline-resistant (b) events. Bottom, Superimposition of the normalized traces ofa and b, showing that the decay phase of bicuculline-resistant events is briefer than that of bicuculline-sensitive events. B2, Comparison of the decay time constant of ATP and GABA mPSCs (∗p < 0.05). C1, C2, Normalized tracesof average amplitude of mPSCs at GABAergic and purinergic synapses tested in the absence or presence of flunitrazepam (Flu) (in the presence of CAS; ECl = 0 mV; VH = −70 mV). In mixed mPSCs, flunitrazepam has no effect on the τfast component, whereas the τslow component is significantly affected (∗p < 0.05; left). Flunitrazepam (100 nm) significantly prolongs the mean decay time constant of mPSCs with a monoexponential decay phase (right).
To examine the possibility that mixed mPSCs reflect the concomitant release of GABA–ATP and the coordinate activation of both postsynaptic receptors, we examined the effect of flunitrazepam, a selective positive modulator of GABAA receptors, on individual mPSCs (Fig. 7C). We reasoned that if both GABAA and P2X receptors are present at the postsynaptic site opposing ATP–GABA input, then flunitrazepam should increase the duration of the time course of all GABAergic mPSCs. Indeed, flunitrazepam (100 nm) significantly prolonged the decay time course of mPSCs with a monoexponential decay phase (from 35 ± 1 to 53.3 ± 3 msec) (Fig.7C1,C2). The effect of flunitrazepam on the population of mixed mPSCs was particularly striking. Thus, the τslow component of mixed mPSCs was significantly prolonged by a low concentration of flunitrazepam, whereas the τfast component of this mPSC population was unaffected (Fig. 7C1,C2). Together, these data indicate that the τslow component of mixed PSCs is sensitive to bicuculline and flunitrazepam, whereas the τfast component of mixed PSCs is insensitive to GABAA receptor modulators. The bicuculline-resistant minicomponents are blocked by P2X receptor antagonists. Thus, the subpopulation of mPSCs identified as mixed mPSCs arises from the coordinate activation of postsynaptic receptors that include both GABAA and P2X receptors.
As suggested by the above experiments, spontaneous release (i.e., mPSCs) appears to involve concurrent activation of both ATP P2X and GABAA receptors at a subset of synapses. Under such circumstances, coreleased ATP and GABA may be costored within the same synaptic vesicle, rather than in separate vesicles. We attempted to test the hypothesis of costorage by further examination of the mPSCs. mPSCs were recorded over a range ofVHs, including theVREV for GABAA(−70 mV), the VREV for P2X receptor (0 mV), and at VHs intermediate to the reversal potentials (i.e., approximately −35 mV). If ATP and GABA are stored within different synaptic vesicles, the spontaneous release of each transmitter should be an independent event. Thus, under these circumstances, we should detect mPSCs of both inward (ATP) and outward (GABA) currents at a VH intermediate to the reversal potentials for both receptors. In contrast, if ATP and GABA are costored in the same vesicle, voltage clamp of the postsynaptic neuron at a potential intermediate toVREV for both receptors should reveal mPSCs that are mixed (i.e., fast inward and then outward currents). Alternatively, if the transmitters are costored, then the costored and coreleased ATP plus GABA may elicit few detectable mPSCs, because the concomitant activation of these two oppositely directed conductances may sum to little net current.
Initial recordings under control conditions revealed both P2X and GABAA receptor-mediated mPSCs atVHs of −70 and 0 mV, corresponding to the reversal potentials for GABAA and P2X receptors, respectively. Subsequent recording from the same neuron while changing the VH to −35 mV revealed very few mPSCs in 75% of the neuron pairs tested in this manner (Fig. 8A1). Application of a low concentration of β-hydrastine (1 μm) blocked the few outward mPSCs that were detected, as expected for this GABAA receptor antagonist (Fig. 8A2) (n = 4). In addition, at later times it appears that the blockade of GABAA receptors subsequently unmasked a population of inward mPSCs. The mean amplitude of inward mPSCs observed at a VH of −35 mV in the presence of β-hydrastine is significantly smaller than that observed at aVH of −70 mV (5 ± 0.1 vs 6.7 ± 0.5 pA; p < 0.01) (Fig.8A3). The decay time constants of mPSCs observed atVHs of both −70 and −35 mV are similar (14 ± 0.8 vs 15 ± 0.7 msec; p > 0.05) (Fig. 8A3), whereas those observed at aVH of 0 mV are significantly different (28.4 ± 1 vs 14 ± 0.8 and 15 ± 0.7 msec;p < 0.01) (Fig. 8A3). Thus, the inward mPSCs at a VH of −35 mV plus β-hydrastine are likely to be ATP P2X receptor-mediated mPSCs. Together, these data suggest that cotransmission of GABA and ATP may be a result of the costorage and corelease of ATP and GABA and the consequent coactivation of postsynaptic ATP P2X and GABAA receptors.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Analysis of GABAA and P2X receptor-mediated mPSCs over a range of VHs.A1, Representative recordings of GABAergic and purinergic mPSCs. Top, Sample traces at aVH of 0 mV (VREVfor P2X receptors); only outward currents were seen.Middle, Sample traces at aVH of −70 mV (VREV for GABAA receptors); inward currents were observed. Bottom, Sampletraces at a VH of −35 mV (intermediate to the reversal potentials for both receptors) with or without β-hydrastine. A2, The amplitudes of mPSCs are plotted as a function of time. At a VH of −35 mV, few events are detected. In the continued presence of β-hydrastine (1 μm), there was an increase in the number of inward current events detected. The solid lines indicate noted changes in conditions (i.e., top line, δVH from 0 to −35; bottom line, addition of β-hydrastine). A3, Histograms of the cumulative probability of mPSC amplitudes and decay time constants recorded at VHs of −70, −35, and 0 mV as indicated. The mean amplitude of mPSCs detected at aVH of −35 mV in the presence of β-hydrastine is significantly smaller than that observed at aVH of −70 mV, but there is no difference between the decay time constants for mPSCs at −70 versus −35 mV. The latter observation is consistent with the idea that all mPSCs recorded at −70 mV and at −35 mV plus β-hydrastine are ATP P2X receptor-mediated. In contrast, both mean amplitude and the decay time constants of mPSCs detected at aVH of 0 mV are significantly different from those observed at VHs of −70 and −35 mV, consistent with the idea that all mPSCs detected at aVH of 0 mV are GABAergic.
DISCUSSION
The major findings of the present study are that (1) ATP is released in an action potential-dependent and in an action potential-independent (TTX-resistant) manner at LH synapses in vitro and (2) recordings from identified pairs of presynaptic and postsynaptic neurons reveal that ATP and GABAAreceptor-mediated postsynaptic currents arise from the coordinate release of both ATP and GABA from individual neurons. We also propose, based on our findings, that ATP and GABA may be costored within the same synaptic vesicle.
Properties of P2X receptors expressed in LH neurons
The existence of fast ATP-mediated excitatory synaptic transmission is well documented in the peripheral nervous system (Evans et al., 1992; Galligan and Bertrand, 1994; Burnstock, 1999). Relatively little is known about purinergic transmission in the CNS, despite the large distribution of ATP receptor subunits in many regions of the CNS. The probability of detection of purinergic synapses in the CNS appears to be quite low, thereby limiting detailed analysis of CNS purinergic synapses. Bardoni et al. (1997) were able to detect an ATP P2X receptor-mediated component of synaptic transmission at only ∼5% of neurons tested in a rat spinal cord slice preparation. Likewise, we noted a higher incidence (i.e., more readily detectable) of ATP P2X receptor-mediated synaptic currents in dispersed cell cultures than in slice recordings from postnatal mice (data not shown). The relatively low incidence of purinergic synapses in the slice and the lack of purinergic mPSCs in recordings from dispersed LH neurons from postnatal mice compared with the other preparations may be attributable to several factors. One possible explanation for the differences in purinergic mPSCs in postnatal mouse versus prenatal chick preparations is the difference in developmental stage. Several studies show decreases in the probability of neurotransmitter release with the development of central synapses (Bolshakov and Siegelbaum, 1995; Choi and Lovinger, 1997; Brenowitz and Trussell, 2001; Iwasaki and Takahashi, 2001). Other confounders may include a higher local activity of ectonucleotidase and soluble ectonucleotidase in slice preparations (Zimmermann, 1996), the presence of more “silent” purinergic synapses in vitro (Khakh et al., 2001), and/or a higher local concentration of metabolites that interfere with ATP P2X receptor activation. Optimal detection of purinergic transmission in slice preparations may require higher frequencies, different patterns of presynaptic activation, or more effective perfusion of the slice.
Our present study provides evidence for robust and reliable action potential-dependent ATP P2X receptor-mediated purinergic transmission in two different in vitro preparations of LH, including neurons from embryonic chicks and neurons from postnatal mice. The characteristics of the evoked purinergic transmission obtained from the present and previous studies in culture and/or in slice preparations (Bardoni et al., 1997; Edwards et al., 1997; Jo and Schlichter, 1999;Mori et al., 2001) are strikingly similar (pharmacology, time course,I vs V characteristics) despite the differences in species, age, etc. Thus, the in vitro culture preparation may provide a better system for the study of ATP P2X receptor-mediated transmission than the in vitro slice preparation (until the highly selective inhibitors for ectonucleotidase and/or P2X receptors are available).
Native ATP P2X receptors are thought to be heteromeric combinations of the various P2X receptor subunits (Lewis et al., 1995) and seem to be assembled as trimers (Nicke et al., 1998). This complexity of P2X receptor structure is thought to underlie the difficulties in reconciling the well characterized pharmacological effects of antagonists on homomeric P2X receptors with those observed in studies with native receptors. The ATP P2X receptors found in the LH are likely to be heteromeric ATP receptors composed of P2X2, P2X4, and P2X6 subunits, which are often coexpressed in the same cells (Collo et al., 1996; Le et al., 1998; Kanjhan et al., 1999). In our experiments and in those of others (Edwards et al., 1992; Bardoni et al., 1997; Pankratov et al., 1998; Mori et al., 2001), the incomplete blockade by ATP P2X receptor antagonists is likely to reflect the heteromeric nature of the receptors involved. Of the subunits expressed in the LH, only the P2X2 subunits show strong sensitivity to suramin and PPADS (North and Surprenant, 2000). In addition, our immunocytochemical data show that LH neurons from postnatal mice express P2X4 subunits, one of several known to be relatively insensitive to PPADS and suramin. It is likely that LH neurons express heteromeric P2X receptors composed of suramin- and PPADS-sensitive P2X2 as well as antagonist-insensitive P2X4 (and/or P2X6) subunits.
Coordinate release of ATP and GABA
GABA is copackaged with glycine within the same synaptic vesicle by vesicular inhibitory amino acid transporter (Gasnier, 2000). Two fast-acting inhibitory neurotransmitters, GABA and glycine, are coreleased and subsequently activate their respective receptors, GABAA and glycine receptors, in the spinal cord (Jonas et al., 1998; Keller et al., 2001). Thus, GABA acts as a cotransmitter in the developing CNS. GABA has been also shown to be coreleased with an excitatory neurotransmitter, ATP, in the spinal cord (Jo and Schlichter, 1999). Our present study is consistent with previous findings of coordinate release of ATP and GABA from the individual presynaptic neurons and provides the first demonstration of ATP–GABA cotransmission in the brain.
Several lines of evidence support our proposal that ATP and GABA are coreleased. Using the dual whole-cell patch-clamp technique, we demonstrate that individual events are stimulus-locked (i.e., individual presynaptic action potentials elicit the corresponding postsynaptic events with the same time of onset and without failure). Second, the latencies for ATP and GABA components are identical. Third, individual events are blocked by a mixture of suramin and bicuculline, but not by bicuculline alone. Finally, intracellular stimulation of the same presynaptic neuron while recording from the same postsynaptic neuron at different membrane potentials elicits both cation-mediated inward currents and Cl−-mediated outward currents that are sensitive to suramin and bicuculline, respectively. Thus, our detection of cotransmission of ATP and GABA is attributable to monosynaptic (rather than polysynaptic) mechanisms.
It is not known how ATP enters the synaptic vesicles or whether ATP is copackaged with GABA within the individual synaptic vesicles. The vesicular transporter for nucleotides has yet to be identified (Gasnier, 2000). The current electrophysiological studies are consistent with (but do not prove) the thesis that ATP and GABA are costored. In experiments examining synaptic transmission at an intermediate VH, we detected few inward or outward current events and could not record mixed inward/outward currents. In the same experiments, outward and inward PSCs were readily detected at a VH of either 0 or −70. Comparison of the time course of mPSCs indicates that the events recorded at VH = −35 after the inhibition of GABAA receptors are equivalent to those detected at VH = −70 (i.e., both are ATP P2X receptor-mediated). The lack of mixed inward/outward currents is also consistent with recent studies of cross-inhibition of ATP P2X with GABAA receptors in the dorsal root ganglions (Sokolova et al., 2001). Sokolova et al. (2001) show that P2X and GABAA receptors can interact or “cross-talk” at the level of the membrane, thereby resulting in mutual inhibition of receptor activation. Inhibition by GABA is strongly dependent on the extent of GABA-gated Cl− efflux. Previous work on NG108-15 cells also indicated that extracellular Cl− may depress P2X receptor function, probably via the inhibition of agonist binding to the receptor site (Kaiho et al., 1997). We cannot exclude the possibility that our inability to detect mixed mPSCs is at least in part a result of the confounding effects of GABAA–ATP P2X receptor interactions and/or of the small mPSC amplitude versus the relatively high baseline noise at VH = −35.
Possible physiological significance
GABA is the primary inhibitory transmitter in the hypothalamus, and GAD mRNA is widely expressed with the LH (Elias et al., 2001). Recent studies demonstrate the existence of GABAergic interneuronal synapses in in vitro preparations of rat LH (Gao and van den Pol, 2001b). These and other studies support a potentially important role of LH inhibitory circuits in the central control of feeding (for review, see Bernardis and Bellinger, 1996). In mature hypothalamic neurons, GABA elicits hyperpolarizing responses and, as such, is inhibitory (Randle et al., 1986; Tasker and Dudek, 1993; Strecker et al., 1997). In contrast, GABA can exert depolarizing actions in developing hypothalamic neurons (Chen et al., 1996). Indeed, in early development, the synaptic release and depolarizing effect of GABA may contribute to suprathreshold activity even more than glutamate (Gao and van den Pol, 2001a). In view of the depolarizing effects of GABA at early developmental stages versus GABA-mediated hyperpolarization at mature LH synapses, the coordinate release of ATP with GABA may support distinct mechanisms for synaptic tuning in the embryonic versus postnatal animal. Thus, the existence of ATP–GABA cotransmission at early stages of development may provide a synergistic mechanism for excitatory influences on postsynaptic neurons. In mature hypothalamic synapses, the activation of GABAA receptors may provide an enhanced driving force for Ca2+entry, thereby increasing the net Ca2+influx via P2X receptor activation. The latter mechanism for GABA and ATP interactions could provide significant synergistic enhancement of ATP receptor-mediated events, as proposed by Robertson et al. (2001). The net physiological effects of ATP and GABA cotransmission will depend on the regulated changes in reversal potential of GABA-evoked current and the localized expression of ATP P2X receptor during development. Our present study supports novel forms of synaptic flexibility via ATP–GABA cotransmission during the development of avian and mammalian hypothalamic circuits.
Footnotes
- Received December 4, 2001.
- Revision received February 15, 2002.
- Accepted February 22, 2002.
This work was supported by National Institutes of Health Grants NS22061 and DA09366 to L.W.R. We thank Drs. Steven A. Siegelbaum, Amy B. MacDermott, and Ron Yu for helpful comments on a previous version of this manuscript and Tom Davis for technical assistance.
Correspondence should be addressed to Dr. Lorna W. Role, Columbia University College of Physicians and Surgeons, Center for Neurobiology, 1051 Riverside Drive, P. I. Annex, Room 807, New York, NY 10032. E-mail: lwr1{at}columbia.edu or Lrole{at}aol.com.
- Copyright © 2002 Society for Neuroscience