

Article Figures & Data
Figures
- Fig. 1.
Creation, histology, and Western analyses of the RC3 KO mouse. A, Top, The RC3 KO construct. Exons 1–4 are numbered. The indicated PstI (P), ApaI (A), and XmaI (X) sites within exon 2 were used to create the KO construct (middle). Other restriction sites mentioned in the text are as follows: H, HindIII;S, SalI; N,NotI; B, BamHI;E, EcoRI (not drawn to scale). A neomycin expression cassette (neo) was used to select for recombinants, and a diphtheria toxin expression cassette (DT) was used to select against random insertion events. lacZ was placed in frame with RC3 so that an RC3-β-galactosidase (RC3-β-Gal) fusion protein (bottom) was expressed from the natural RC3 promoter in recombinant mice. B, Western blot showing RC3 and RC3-β-gal expression in WT, heterozygous RC3 KO, and homozygous RC3 KO mice. Twenty, 10, 5.0, and 2.5 μg of total forebrain homogenates were resolved by SDS-PAGE and blotted onto a PVDF membrane. A polyclonal antibody generated against RC3 was used to detect both native RC3 and the RC3-β-gal fusion protein. C, β-Galactosidase activity in a sagittal section from a homozygous RC3 KO mouse brain (top) and, for comparison, a dark-field view of a WT sagittal brain section that was probed in situ with RC3 antisense RNA (bottom). Nissl (D) (magnification, 4×; scale bar, 50 μm) and Timm's stain (E) (magnification, 10×; scale bar, 150 μm) of coronal sections from RC3 KO (top) and WT (bottom) mouse brains.gc, Granule cell layer; Th, thalamus;h, hilus; IP, infrapyramidal mossy fiber layer; SP, suprapyramidal mossy fiber layer.F, Comparison of Ca2+/CaM-independent CaMKII levels in WT and mutant hippocampal slices before and after LTP. CaMKII and phospho-CaMKII immunoreactivities were assayed (left), as was Ca2+/CaM-independent CaMKII activity before and 20 min after inducing LTP. Eachbar is the mean activity in five or six slices (indicated in bar graph) from three mice. G, Synaptophysin density in hippocampal area CA1 of RC3 KO and littermate controls is the same. Synaptophysin binding was determined as a z-stack of 1 μm sections throughout a 15 μm slice, and the integrated intensity of the entire stack was determined.
- Fig. 2.
Comparison of excitatory synaptic strength and PPF in hippocampal slices of RC3 KO and age-matched littermate controls.A, Raw measures. All traces are an average of two successive pEPSPs evoked at 0.02 Hz; truncated stimulus artifacts indicated by arrowhead. Top trace shows the normalization procedure. We measured PVs (y) and pEPSP amplitudes (z) as the y-axis difference between the negative peaks and the baseline values (dotted line) determined preceding the stimulation (x). Bottom traces represent typical I–O curves in littermate control animals and RC3 KO animals.B, I–O curves for mean amplitudes of pEPSPs. Population EPSP amplitudes are similar in slices from age-matched littermate control animals and RC3 KO mice. C, Mean PV amplitudes at stimulation intensities evoking threshold, 30–50% maximal, 50%, and maximal pEPSP amplitude. D, Significantly larger mean values of the PV indicate that more fibers must be stimulated to obtain equivalent pEPSPs in RC3 KO animals. Normalization of the pEPSP amplitudes to PV size shifts the I–O curve down significantly, suggesting that synaptic transmission in RC3 KO animals is chronically depressed (*p < 0.05 indicates significant differences). E, Comparison of the correlation between PV amplitude and stimulation intensity. There was no difference between the linear regression lines (t test;p > 0.05). F, PPF of pEPSPs measured at 20, 50, 100, and 200 msec interstimulus intervals. There is no difference between slices of RC3 KO and littermate control animals. Each point is the mean ± SEM; nindicates number of slices per number of animals.
- Fig. 3.
NMDA-mediated excitability is depressed in RC3 KO mice. NMDA-mediated pEPSPs were isolated as described in Materials and Methods. A, Representative single tracesfrom pharmacologically isolated NMDA pEPSP. B, Mean NMDA–compound–pEPSP ratios in mutant slices are not different from those in WT slices. C, The I–O curve for NMDA-pEPSPs was shifted down in RC3 KO slices to the same degree as were compound-EPSPs (see Fig. 2D). The mean NMDA-pEPSP was normalized to the PV.
- Fig. 4.
Increased LTP in RC3 KO mice.A, Mean data. Two trains of high-frequency stimulation (100 pulses at 100 Hz) induced significantly larger post-tetanic potentiation and LTP in slices from RC3 KO animals than in slices from age-matched littermate controls. B, In slices from RC3 KO mice, LTP saturation was essentially established after the first tetanus. Although additional tetani are able to induce further potentiation, the mean pEPSP slopes were not significantly higher than after the first. In slices from littermate control animals, consecutive tetani progressively potentiated the pEPSP until reaching a similar plateau as the RC3 KO mice. C, Mean pEPSP slopes (after tetanus) and mean normalized pEPSP30–50% amplitudes (before tetanus) are inversely correlated in RC3 KO animals but not in control slices, suggesting that enhanced LTP in slices from RC3 KO mice is caused by a depressed baseline. D, LTP was induced in WT or RC3 KO slices and compared with LTP induced in WT slices after conditioning with either one or three low-frequency stimuli. After a single LFS, WT PTP (gray diamonds) was similar to RC3 KO PPT (black squares), but, by 40 min, LTP in conditioned (gray diamonds) and naı̈ve (white diamonds) WT slices was not significantly different (p > 0.2). However, three consecutive low-frequency stimuli enhanced LTP in WT slices to the point that there was no significant difference between preconditioned WT slices (gray circles) and naı̈ve RC3 KO slices (black squares;p > 0.4). Before LTP induction, a complete I–O curve was constructed, and baseline was adjusted to the new 30–50% of the maximal pEPSP.
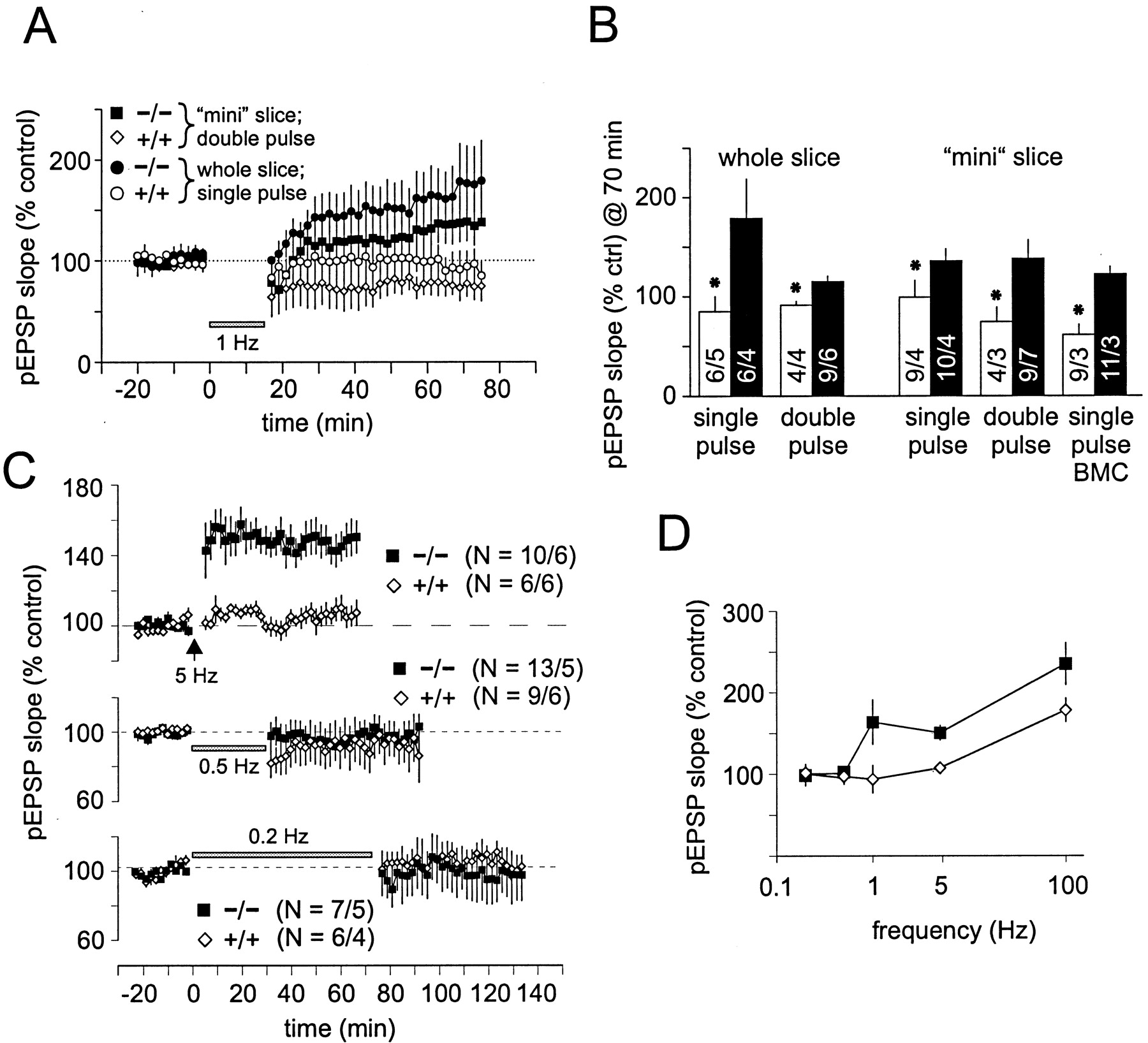
- Fig. 5.
Comparison of long-term plasticity in RC3 KO mice.A, Two different low-frequency stimulation protocols (either 900 single pulses or 900 double pulses at 50 msec interval, both at 1 Hz) induced LTD in slices from littermate control animals (mean data). However, the same stimulation protocols induced slow-onset potentiation in slices from RC3 KO animals. In mini slices, CA3 was removed to reduce feedback excitation, and, in some experiments, bicuculline (BMC) was added to the bath to reduce GABAergic inhibition.B, Pooled data from all 1 Hz LTD experiments. Under all conditions tested, LFS consistently induced depression in slices from control mice but potentiation in slices from RC3 KO animals (*p ≤ 0.04 indicates statistical significance).C, Nine hundred pulses at 5 Hz induced a significant potentiation in slices from RC3 KO animals but not in slices from littermate control animals. Lower frequencies (0.5 and 0.2 Hz, 900 pulses) induced neither short-term nor long-term changes of pEPSP slopes in RC3 KO or control slices. D, Consistent with our model, the frequency–response curve of RC3 KO is shifted. However, at 0.2 and 0.5 Hz, there were no apparent changes, suggesting that RC3 KO slices were maximally depressed.
- Fig. 6.
Depotentiation and saturation of pEPSP slopes after repeated LFS with and without priming. A, Depotentiation is much larger in RC3 KO animals than in control animals. Twenty-two minutes after a priming stimulus (2 trains of 100 pulses at 100 Hz), we induced depotentiation with 900 stimuli at 1 Hz. In WT mice, pEPSP slope values return to control levels (before LTP), whereas RC3 KO animals were depotentiated much more so (50%) and stayed significantly depressed for over 60 min. Parallel analysis of the PVs (bottom) did not show any changes.B, After high-frequency priming, five consecutive low-frequency tetani (900 pulses at 1 Hz) were needed to saturate depotentiation in slices from WT animals (50% of baseline, similar to depotentiation after 4 sets of LFS without priming) (see Fig. 5D).C, Even after two low-frequency tetani, depotentiated slices from RC3 KO slices still expressed a higher LTP magnitude. Whereas a second LFS induced a slight potentiation in slices from RC3 KO animals, mean pEPSPs in WT slices were further depotentiated. A consecutive HFS induced pronounced LTP in both. After 40 min, LTP magnitude was significantly higher in slices from RC3 KO compared with WT (p = 0.006). D, In slices from RC3 KO animals, consecutive tetani of LFS caused increasing degrees of slow-onset LTP (plateaus at 140–150% of control). The same LFS induced increasingly robust LTD in slices from littermate control animals. E, Superfusion of the NMDA receptor antagonist APV during the priming stimulus inhibited initial potentiation. However, in slices from both RC3 KO and control animals, a subsequent LFS (without APV) depotentiated the slices to the same extent as inA. There were no changes in the PV (bottom).
- Fig. 7.
Slices from RC3 KO mice are less sensitive to group 1 mGluR agonists. A, Although 100 μmACPD for 20 min initially reduced the mean pEPSP in slices from RC3 KO mice less than in slices from WTs, after 20 min, the differences were not statistically significant. A full recovery was established after 40 min of washout. B, DHPG, the group 1-specific mGluR agonist, induced a significantly larger and nonreversible reduction of mean pEPSPs in slices from WT animals compared with those from RC3 KOs. The slight but insignificant reduction compared with baseline suggests that mGluR5 maybe involved in mediating the phosphorylation of RC3.
- Fig. 8.
A model of the postulated mechanism governing metaplasticity in the CA1 region of the mouse hippocampus.Top, A sliding threshold model of metaplasticity in the hippocampus. The phosphorylation states of RC3 and CaMKII determine the LTD threshold (θ−), the LTP threshold (θ+), and the ceiling at which LTP is saturated. NMDA receptor-mediated events predominate during HFS, causing dephosphorylation of RC3 and phosphorylation of CaMKII, thereby shifting θ− and θ+ to the right. Type I mGluR-mediated events predominate during LFS, leading to phosphorylation of RC3 and dephosphorylation CaMKII, shifting θ− and θ+ to the left. LTP is saturated when RC3 is minimally and CaMKII maximally phosphorylated, placing θ+ at its rightmost limit. When RC3 is maximally phosphorylated and CaMKII minimally so, θ− and θ+ are situated to the far left, and LTD is induced by small levels of basal activity. Bottom, Circuit diagram of pathways governing CaM availability, the frequency–response threshold, and metaplasticity. Blue arrows denote pathways involved in the induction of LTP (high Ca2+). Red arrows denote pathways involved in LTD (medium Ca2+), and green arrows signify steady-state conditions in the resting synapse. Based on mechanistic insights gleaned from in vitroobservations concerning the dissociation kinetics of RC3 and CaM, we mapped a hypothetical network of commonly accepted biochemical pathways that could explain various metaplastic phenomena. Unphosphorylated RC3 acts as a CaM sink when Ca2+ levels are low (green arrows), releases CaM slowly during medium Ca2+ fluxes (red arrows), and releases Ca2+ rapidly during large NMDA-dependent Ca2+ fluxes (blue arrows). In the latter instance, both high- and low-affinity CaM-binding enzymes are activated. High-affinity enzymes include the Ca2+/CaM-dependent phosphatase calcineurin (PP2B) and the CaM-dependent phosphodiesterase (PDE). Low-affinity enzymes comprise CaMKII and adenylyl cyclase (AC). When high levels of Ca2+ are present and CaM is freely available, cAMP is synthesized by adenylyl cyclase faster than it can be hydrolyzed by PDE, resulting in the activation of PKA. Inhibitor 1 (I1) is phosphorylated by PKA faster than it isdephosphorylated by PP2B, resulting in the inhibition of protein phosphatase 1 (PP1). Thus, the accumulation of phosphorylated CaMKII and the activation of PKA are favored, resulting in LTP. As CaM is sequestered by phospho-CaMKII, activation of the high-affinity binders becomes favored over the low-affinity binders, placing an upper limit on LTP. The activities of high-affinity binders are also favored during smaller Ca2+ fluxes (red arrows). In this case, PDE hydrolyzes cAMP faster than adenylyl cyclase creates it, decreasing the rate of inhibitor 1 phosphorylation by PKA. This, along with increased PP2B activity, decreases the phosphorylation state of inhibitor 1, which disinhibits protein phosphatase 1, decreases the phosphorylation state of CaMKII, and results in LTD. When Ca2+ levels are very low, neither low-affinity nor high-affinity binders are favored, and the final equilibrium concentrations of phosphorylated and dephosphorylated CaMKII at resting Ca2+ levels will depend on the CaM-buffering capacity of RC3. Thus, the phosphorylation state of RC3 would determine the degree of potentiation or depotentiation in the resting synapse by setting baseline levels of phospho-CaMKII and PKA activities. The amount of available CaM, which is governed by the phosphorylation states of both RC3 and CaMKII, would determine the kinetics, direction, and magnitude of synaptic responses to subsequent Ca2+ fluxes. The dotted green linesemphasize the notion that significant CaMKII and PKA activities exist in the resting synapse. Phosphorylation of RC3 increases sensitivity to Ca2+ such that the red pathwaysbecome more dominant at basal Ca2+ concentrations and the blue pathways become favored at medium Ca2+ concentrations. Thus, ablation or phosphorylation of RC3 would shift the resting synapse into a state of LTD and displace θ+ to the left. At modest levels of Ca2+, RC3 and CaM dissociate rather slowly, so we would expect RC3 to constrain the speed and magnitude of CaM-dependent reactions. Thus, ablation or phosphorylation of RC3 would tend to amplify Ca2+/CaM-induced shifts in θ+.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}