
Abstract
The occurrence of abnormally associated movements is inevitable after facial nerve transection. The reason for this post-paralytic syndrome is poor guidance of regrowing axons, whereby a given muscle group is reinnervated by misrouted axonal branches. Olfactory ensheathing glia have been shown to reduce axonal sprouting and stimulate axonal regeneration after transplantation into the spinal cord. In the present study, we asked whether transplantation of olfactory mucosa (OM) would also reduce sprouting of a damaged peripheral pure motor nerve. The adult facial nerve was transected, and the effect of the OM placed at the lesion site was analyzed with regard to the accuracy of target reinnervation, axonal sprouting of motoneurons, and vibrissal motor performance. Accuracy of target reinnervation and axonal sprouting were studied using preoperative/postoperative labeling and triple retrograde labeling of facial motoneurons, respectively. The vibrissal motor performance was monitored using a video-based motion analysis. We show here that implantation of OM, compared with simple facial–facial anastomosis, (1) improved the protraction, amplitude, angular velocity, and acceleration of vibrissal movements up to 80% of the control values, (2) reduced the percentage of branching motoneurons from 76 to 39%, and (3) improved the accuracy of reinnervation from 22 to 49%. Moreover, we present evidence, that transplanted OM but not buccal mucous membrane induced a sustained upregulation of trophic factors at the lesion site. It is concluded that transplantation of OM to the transected facial nerve significantly improves nerve regeneration.
- regeneration
- olfactory ensheathing glia
- axonal pathfinding
- collateral sprouting
- retrograde labeling
- whisking behavior
The peripheral nervous system is generally characterized by its ability to achieve functional recovery after peripheral nerve injury. However, complete recovery of function in the clinical practice is only rarely observed. The occurrence of abnormally associated movements and altered reflexes is practically inevitable, especially after surgical repair of the facial nerve (Kimura et al., 1975; Anonsen et al., 1986; Baker et al., 1994).
The main reason for this “post-paralytic syndrome” is the “misdirected” or “aberrant” reinnervation of muscles (Montserrat and Benito, 1988), which has two components. First, because of malfunctioning axonal guidance, a muscle becomes reinnervated by a “foreign” axon, which has been misrouted along the “wrong” fascicle (Esslen, 1960; Aldskogius and Thomander, 1986; Brushart and Seiler 1987), and second, because of the presence of supernumerary branches from all transected axons (Shawe, 1954; Morris et al., 1972), a muscle is simultaneously reinnervated by several motoneurons (Ito and Kudo, 1994), a state known as “polyneuronal innervation” (Brown et al., 1981; Rich and Lichtman, 1989) or “hyperinnervation” (Angelov et al., 1993, 1996).
Attempts to act on the first component of aberrant reinnervation, achieving a “fascicular” or “specificity” (Mackinnon et al., 1986; Evans et al., 1991) have failed so far. Although claimed to be transient (Hennig and Dietrichs, 1994), the aberrant innervation may persist for extended periods (Mackinnon et al., 1991; Madison et al., 1999), with deleterious effects on synchronized function. Thus, the post-lesional axonal branching, considered to represent the search of axons for local guidance cues (AlMajed et al., 2000), may be the reason for the failure of axons to reinnervate solely their original domain (Fu and Gordon, 1997; Angelov et al., 1999).
Observations suggested that implantations of olfactory ensheathing cells (OECs) minimized axonal sprouting in the spinal cord. We therefore studied the effects of purified cultured neonatal OECs transplanted into the transected facial nerve and were surprised to find that implantation of OECs dramatically increased sprouting of axotomized facial motoneurons (Guntinas-Lichius et al., 2001). This effect may be explained by high levels of trophic factors providedin situ by the transplanted cells (Bunge, 1993; Gallo and Letourneau, 1998; Davies, 2000; Mackay-Sim and Chuah, 2000;Raisman, 2001). Recent evidence indicates that the dissociation and prolonged cultivation of OECs results in upregulation of trophic factors involved in axonal sprouting, e.g., the ciliary neurotrophic factor (CNTF) and its α receptor subunit (Lee et al., 1997; Wewetzer et al., 2001). To prevent excessive trophic factor supply by cultured OECs, we chose to use instead intact autologous olfactory mucosa (OM) for transplantation. OM has been successfully applied recently to the experimentally lesioned spinal cord (Lu et al., 2001, 2002).
Comparing (1) the degree of axonal branching, (2) the accuracy of muscle target reinnervation, and (3) the motor performance of the vibrissal hairs between rats with facial nerve suture only (FFA) and rats with FFA plus small pieces of OM transplanted at the suture site, we found that the addition of OM significantly improved the quality of target innervation and recovery of function.
MATERIALS AND METHODS
Materials
Fluoro-Gold (FG) was purchased from Fluorochrome Inc. (Denver, CO), Fast Blue (FB) was from EMS-Chemie GmbH (Groβ-Umstadt, Germany), and 1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanin perchlorat (DiI) was from Molecular Probes (Leiden, The Netherlands; cat. no. D-282).
Animals and surgery
Forty female and 6 male inbred Lewis rats (175–200 gm; strain LEW/SSN; Harlan Winkelmann, Borchen, Germany) were used. Before and after the experiments, rats were kept on standard laboratory food (Ssniff, Soest, Germany) and tap water ad libitum, with an artificial 12 hr light/dark cycle. All surgical procedures were performed under microscopic control after an intraperitoneal injection of ketamin/xylazin as described previously (Dohm et al., 2000; Guntinas-Lichius et al., 2001). All experiments were conducted in accordance with the German Law for Animals' Protection and approved by the local animal care committee (Bezirksregierung Köln, Az. 23.203.2-K35,13/95).
Transection and immediate end-to-end suture of the right facial nerve (facial–facial anastomosis) was performed in female rats only. As described previously, the main trunk of the facial nerve was exposed under deep anesthesia and transected close to its emergence from the foramen stylomastoideum, distal to the posterior auricular branch (see Fig. 1A). The proximal stump was then microsurgically reconnected to the distal stump with two 11-0 atraumatic sutures (Ethicon EH 7438G, Ethicon, Norderstedt, Germany).
In half of the female rats, OM, freshly prepared from deeply anesthetized syngeneic male rats, was cut in small pieces after a short rinse in HBSS (Life Technologies Overseas GmbH; cat. no. 24020083) and then gently laid over the sutured epineurium. Male rats were chosen as donors to facilitate a later identification of the transplant in the female hosts. Finally, the wound was closed by three 4-0 skin sutures (Ethicon).
In four other female rats, buccal mucous membrane (BMM) obtained from the cheeks of deeply anesthetized syngeneic male animals was laid over the sutured epineurium. These control rats were used to prove whether the improved regeneration was really caused by OM and not by the mechanical effect(s) of any transplanted tissue.
Estimation of post-operative axonal branching
Triple retrograde labeling was performed as reported previously (Dohm et al., 2000; Guntinas-Lichius et al., 2001). Briefly, the zygomatic, buccal, and marginal mandibular ramus of intact animals (six rats), of animals that received an OM transplant (six rats), and of animals that underwent FFA-only (six rats) were transected, and tracer crystals (DiI, FG, FB) were applied (see Fig. 1B). Tracing was done 2 months after the operation. Ten days after the post-operative bilateral labeling, rats were transcardially perfused (0.9% NaCl in distilled water) followed by fixation with 4% paraformaldehyde (0.1 m phosphate buffer, pH 7.4) under deep anesthesia. Brainstems were cut coronally (50 μm) using a vibratome.
Sections were evaluated using an epifluorescence microscope (ZeissAxioskop 50) a custom-made bandpass-filter set for Fast Blue, and a HQ-Schmalband-filter set for Fluoro-Gold. The use of an additional filter set (filter set 15, Carl Zeiss; excitation BP 546/12, emission LP 590) revealed the red fluorescence of those motoneurons retrogradely labeled by DiI. The fluorescence cross-talk between the tracers was restricted with this filter combinationad maximum (Dohm et al., 2000; Guntinas-Lichius et al., 2001).
Unfortunately, some FB-labeled motoneurons could be seen even through the HQ-Schmalband-filter set. This imposed serious difficulties when a differentiation between single-labeled (FG or FB) and double-labeled (FG + FB) motoneurons had to be made. Therefore, in the present study we counted as double labeled neurons only those containing DiI + FG or DiI + FB but did not evaluate those labeled by FG + FB.
Image analysis was done using a CCD video camera (Optronics Engineering DEI-470) combined with the image analyzing software Optimas 6.5. (see above). For a detailed description of the procedure, see Dohm et al. (2000). Using the fractionator principle (Gundersen, 1986), all retrogradely labeled motoneurons with a visible cell nucleus were counted in every third section through the facial nucleus on the operated and unoperated sides (Guntinas-Lichius et al., 1993). All counts were performed by two observers who were blind to the surgical procedure used on the rats.
Determining the accuracy of target reinnervation
Intramuscular injection of tracer (FG) was used to reveal the distribution of the original motoneuron pool. Under deep anesthesia, FG dissolved in distilled water (1 mg/100 μl) containing 2% dimethyl sulfoxide (DMSO) was injected into each whisker pad of 18 intact rats. Ten days later, six animals underwent FFA-only, and six underwent FFA + OM. The last six rats remained unoperated and received the second label (FB) 2 months later. Two months after FFA-only or FFA + OM, all rats (including the six intact animals) received bilateral injections of FB (1%) into the whisker pad musculature, at the site of the primary FG injection (Popratiloff et al., 2001, their Fig. 1). Ten days after the post-operative bilateral labeling, all 18 rats were fixed (4% paraformaldehyde/0.1 m phosphate buffer, pH 7.4) under deep anesthesia, and the brainstems were cut coronally (50 μm).
Qualitative fluorescence microscopy was performed with a UV-excitation filter (Carl Zeiss; Filter Set 01, excitation BP 365/12, emission LP 397), which allows recognition of both FG- and FB-labeled neurons. The CCD video camera system, together with the image analyzing software (see above), was used to create separate images of the FG and FB retrogradely labeled facial motoneurons. Custom-made filter sets for FG and FB [(AHF Analysentechnik, Tübingen, Germany) FG-filter: HQ-Schmalband-filter set (no. F36–050; excitation D 369/40; beamsplitter 400DCLP; Barrierfilter HQ 635/30); FB-filter: Bandpass-filter set (no. F31–000; excitation D 436/10; beamsplitter 450 DCLP; Barrierfilter D470/40)] were used. The generated masks of FG-labeled cells were superimposed over the FB image for the unlesioned as well as for the lesioned facial nucleus (Popratiloff et al., 2001). In this way, all motoneurons stained by FG and FB and double labeled by FG + FB could be readily identified and counted.
Analysis of vibrissae motor performance
The key movements of the vibrissae are protraction (see Fig.1B) and retraction (see Fig. 1C). Because all vibrissal piloerector muscles are innervated by the buccal branch (Dörfl, 1985), the whiskers acquire caudal orientation and remain motionless after transection of the facial nerve. Two large vibrissae of the C row on each side of the face were used for biometric analysis, as described previously (Guntinas-Lichius et al., 2001). Under light anesthesia, all other vibrissae were clipped using small fine scissors, and the animals were inserted into a rodent restrainer (Hugo Sachs Electronik–Harvard Apparatus GmbH, AH 52-0292, 79232 March-Hugstetten, Germany) for 30 min to pacify them. Using a digital camcorder (Panasonic NV DX-110 EG), animals were videotaped for 3–5 min during active exploration. After calibration, video images of whisking behavior were sampled at 50 Hz (50 fields per second); the video camera shutter was open for 4 msec. Images were recorded on AY-DVM 60 EK mini-cassettes. Captured video sequences were reviewed, and 1.5 sec sequence fragments from each animal were selected for analysis of whisking biometrics. Thereby the stable position of the animal's head, the frequency of whisking, and the degree of vibrissae protraction were considered as selection criteria.
The tip of the rat's nose and the inner angles of both eyes were defined as reference points. Each vibrissa in the spatial model was represented by two points: its base and a point on the shaft 0.5 cm away from the base. Using this model, the following parameters were evaluated: (1) protraction (i.e., the forward movement of the vibrissae) measured by the rostrally opened angle (in degrees) between the midsagittal plane and the hair shaft (accordingly, maximal protractions were represented by rather low angle values); (2) the whisking frequency as cycles of protraction and retraction (passive backward movement) per second; (3) the amplitude (the difference between maximal retraction and maximal protraction in degrees); (4) the angular velocity during protraction in degrees per second; and (5) the angular acceleration during protraction in degrees per second squared.
Statistics
All data were presented as means ± SD. A one-way ANOVA and a post hoc t test for unpaired data with Bonferroni-Holm correction were applied to detect differences between the different groups. A p value of <0.05 was considered statistically significant.
Morphological analysis of the suture/implantation site
Detection of the transplanted tissue using a Y chromosome-specific DNA probe. To study the definitive localization of the transplanted OM at the end of the experiments and to verify that the transplanted small pieces of olfactory mucosa had not been rejected or degraded, the distribution of the transplanted cells was studied using in situ hybridization and a Y chromosome-specific DNA probe. The Y chromosome is a stable congenic marker that allows the unbiased identification of male cells grafted into female recipients (Irintchev et al., 1997; O′Leary and Blakemore, 1997). All digoxigenin (DIG)-labeled cRNA probes were generated by in vitro transcription, using PCR-generated templates according to the protocols of An et al. (1997).
Hybridization was performed on free-floating sections (40 μm) from the facial nerve at the transplantation site 2 months after suture (Haas et al., 1999). Cryosections proceeded through the following steps. (1) Sections were collected in 2× SSC (1× SSC = 0.15m NaCl, 0.015 m sodium citrate, pH 7.0); (2) sections were pretreated in a 1:2 mixture of 2× SSC/hybridization buffer (50% formamide, 4× SSC, 50 mmNaH2PO4, 250 μg/ml heat-denatured salmon sperm DNA, 100 μg/ml tRNA, 5% dextran sulfate, and 1% Denhardt's solution) for 15 min; (3) prehybridization of sections in hybridization buffer for 60 min at 55°C; (4) probe concentration was 100 ng/ml; hybridization lasted 12 hr at 55°C; (5) sections were washed in 2× SSC at room temperature; (6) sections were washed in 2× SSC plus 50% formamide; (7) sections were washed in 0.1× SSC and 50% formamide; (8) sections were washed in 0.1× SSC (steps 5–8 were performed at a temperature 10°C above the respective hybridization temperatures); (9) immunological detection of DIG-labeled hybrids with anti-DIG-alkaline phosphatase was performed as recommended by the manufacturer (Roche); (10) colorimetric detection was performed using nitroblue tetrazolium and 5-bromo-4-chloro-3-indolylphosphate; (11) development of the color reaction was performed in the dark at room temperature and stopped after 30 min by transfer into 10 mm Tris/HCl, pH 8.0, 1 mm EDTA; (12) sections were embedded in Moviol (Hoechst). Hybridized male cells showed a dark purple spot in the nucleus, whereas female cell nuclei were unstained. Specificity controls included the omission of the labeled probe, the omission of the antibody to DIG, and the use of female instead of male tissue as a negative control.
Detection of trophic factors at the suture/implantation site using immunohistochemistry. To monitor the expression of trophic factors at the lesion site at the end of the experiments, longitudinal cryosections (20 μm) through the facial nerve 2 months after FFA-only, FFA + OM, or FFA + BMM were incubated with (1) mouse monoclonal anti-NGF (1:50; Roche; cat. no. 1087754), (2) mouse monoclonal anti-BDNF (1:1000; R&D Systems; MAB 248), (3) mouse monoclonal anti-basic FGF (bFGF) (1:50; UBI/Biomol; cat. no. 05-117), (4) goat polyclonal anti-rat CNTF (1:100; R&D Systems; AB557-NA), (5) mouse monoclonal anti-IGF-I (1:50; UBI/Biomol; cat. no. 05-172), (6) mouse monoclonal anti-GDNF (1:500; R&D Systems; MAB 212), (7) rabbit polyclonal anti-S-100 protein (1:1000; Dako Diagnostika, Hamburg, Germany).
Secondary antibodies (all diluted 1:400) were (1) biotinylated goat anti-mouse IgG (Fc specific, Sigma, no. B-9904), (2) biotinylated rabbit anti-goat IgG (Dako, no. E0466), and (4) biotinylated goat anti-rabbit IgG (Dako, no. E0432), according to standard protocols (Streppel et al., 2002).
RESULTS
Transplantation of olfactory mucosa reduces the collateral axonal branching
Transplantation of OM to the site of FFA significantly (p < 0.05) reduced axonal collateral branching, as determined by counting the motoneurons that sent an axon through the zygomatic nerve branch and twin branches in two different nerves (Table1). After transplantation of OM, 312 ± 88 motoneurons sent an axon or an axonal branch through the zygomatic ramus compared with 482 ± 50 after FFA-only and 302 ± 25 in the intact animal. OM also significantly (p < 0.05) reduced the number of double-labeled motoneurons to ∼39% compared with 76% after FFA. This was accompanied by a significantly (p < 0.05) increased portion of motoneurons (61%) that sent a single unbranched axon through the zygomatic ramus (Table 1).
Numbers of retrogradely labeled facial perikarya, the axons of which project through the zygomatic, buccal, and marginal mandibular branches in intact rats and in rats that underwent unilateral FFA-only or unilateral FFA + OM
Triple retrograde labeling of motoneurons in the intact animals revealed the myotopic organization of the facial nucleus. No double-labeled perikarya were observed; i.e., under normal physiological conditions, motoneurons do not send twin branches to different nerve rami (see Fig. 2A). No fluorescent perikarya were ever found in the medial or ventromedial facial subnucleus, the motoneurons of which project through the posterior auricular and cervical branch, respectively; neither nerve was an object of transection and labeling. This is in accordance with our previous studies (Dohm et al., 2000; Guntinas-Lichius et al., 2001).
Transection and suture of the facial nerve completely abolished the myotopic organization (see Fig. 2B). Furthermore, ∼75% of the motoneurons sending an axon to the zygomatic branch gave rise to a twin branch and got double labeled (see Fig.2B, Table 1). The total number of retrogradely labeled motoneurons in all operated animals was higher than that in the control intact rats.
This post-transectional hyperinnervation of targets (Angelov et al., 1996) was caused by a three- to fourfold increase in the number of motoneurons single labeled by FB (Table 1, last column). The majority of axons that took up the retrograde tracer originated from motoneurons projecting under control conditions to the cervical branch of the facial nerve (Fig. 1A). Because this branch was not traced in intact animals (Fig.1B), the corresponding motoneurons were not visible until the post-transectional misguidance “lead” their axons into the FB-traced marginal mandibular branch. Although we could detect and report similar changes after insertion of the transected facial nerve in a conduit containing cultured OECs (Guntinas-Lichius et al., 2001) or neutralizing antibodies to trophic factors (Streppel et al., 2002), we still cannot explain why this was not the case in animals that were subjected to FFA-only. Transplantation of OM did not restore the myotopic organization.
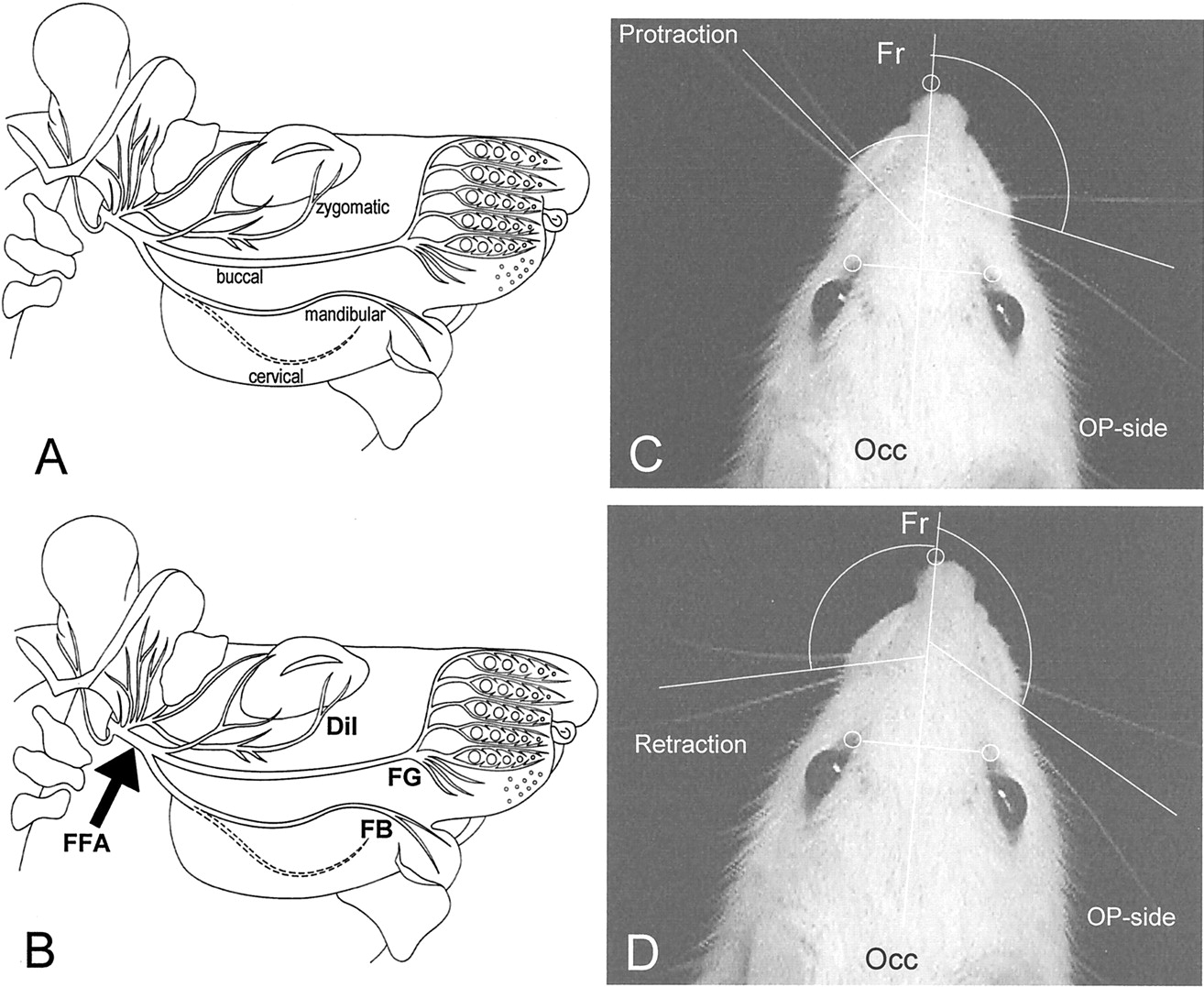
A, B, Schematic drawings of the infratemporal portion of the rat facial nerve. The site of transection and end-to-end suture of the facial nerve trunk, i.e., facial-facial anastomosis (FFA), FFA + buccal mucous membrane, or FFA + olfactory mucosa, is indicated by anarrow. The tracer application sites in the distal parts of the transected zygomatic, buccal, and mandibular nerves are indicated by abbreviations of the three different labels applied, i.e.,DiI, FG, and FB, respectively (B). C,D, The developed spatial model allows precise measurement of angles, angular velocity, and angular acceleration on the intact (left) and operated side (right) during protraction (C) and retraction (D) of the vibrissae. Note the significant change in angle between the fronto-occipital (Fr-Occ) sagittal line during protraction and retraction on the intact side. The vibrissae on the operated (OP) side remain spastic.
Taken together, our data clearly demonstrate that transplantation of OM significantly reduced collateral branching compared with FFA-only but did not completely restore the original innervation pattern as observed in the intact animal. Interestingly, transplantation of OM significantly increased the number of single-labeled motoneurons projecting in the marginal mandibular branch.
Transplantation of olfactory mucosa does not increase the accuracy of reinnervation
After transplantation of OM to the suture site, a mean of 1625 ± 553 motoneurons was found to reinnervate the whisker pad compared with 2024 ± 103 after FFA and 1540 ± 94 in the intact animal (Table 2). However, the reduction of 33% compared with FFA was statistically not significant. After transplantation of OM, an increased portion of motoneurons was observed that succeeded in reinnervating their original target: 597 ± 283 motoneurons correlated to 49% compared with 320 ± 50 (22%) after FFA and 1367 ± 96 (93%) in the intact animal. This increase was also not statistically significant compared with FFA.
Mean number and SD of retrogradely labeled facial perikarya after injection of 100 μl 1% FG as a preoperative label and 100 μl 1% FB as a postoperative label in intact rats and in rats that underwent unilateral (on the right side) FFA-only or unilateral (on the right side) FFA + OM
Injection of FG into the whisker pad of intact rats was found to label 1472 ± 71 facial motoneurons. Two months later, injection of FB as close as possible to the primary injection site labeled 1515 ± 78 motoneurons. This demonstrates that there is no difference in the labeling efficiency for FG and FB (Table 2). Moreover, both tracers labeled only motoneurons that were localized in the lateral facial subnucleus, which is in agreement with the myotopic organization of the facial nucleus in intact rats (Fig.2C). Control experiments, including the injection of tracers into the intact facial nucleus of the side contralateral to FFA (left side), again revealed no differences in labeling efficiency between both tracers. The data were comparable to those of the intact animal (see above).

Estimation of postoperative axonal branching (A, B) and accuracy of reinnervation (C, D). A, Triple labeling in the facial nucleus after application of crystalline DiI to the zygomatic, FG to the buccal, and FB to the marginal mandibular nerves of an unoperated control rat. Note the myotopic organization of the nucleus. The DiI-labeled motoneurons (red) are localized mostly in the dorsal, FG-labeled (white-yellow) motoneurons are localized mainly in the lateral, and FB-labeled (blue) motoneurons are localized mainly in the intermediate facial subnucleus. B, Eight weeks after FFA-only or FFA + OM, the myotopic organization was completely eliminated, and various double-labeled motoneurons sending branches in different nerve rami were observed. C, Unoperated control rat. The preoperative and postoperative neuronal labeling with FG (yellow) and FB (blue) support our counts showing that in the intact facial nucleus, the portion of double-labeled (FG + FB) motoneurons is >95% (Table 2). D, Eight weeks after FFA-only or FFA + OM. Although all FG-labeled motoneurons are localized in the lateral facial subnucleus, those labeled with FB are observed dispersed throughout the whole facial nucleus. Our quantitative estimates show that after FFA-only, ∼20% of these FB-labeled motoneurons are double labeled and belong to the original motoneuronal pool of the whisker pad (Table 2).
After FFA, no myotopic organization into subnuclei was evident, and the bulk of motoneurons (∼80%) that were retrogradely labeled by the postoperative tracer FB were scattered throughout the other facial subnuclei (Fig. 2D, Table 2). Because of the post-transectional axonal branching (see above), the number of retrogradely labeled motoneurons in all operated animals was higher than that in the control intact rats.
Transplantation of olfactory mucosa promotes functional recovery of vibrissal motor performance
The detailed biometric analysis of whisking behavior was performed to test whether the implantation of OM, besides its effect on the axonal collateral formation, also improved motor performance as measured by a functional test. All animals that underwent FFA and transplantation of OM showed a significantly (p< 0.05) better recovery of the biometrical parameters than the rats with FFA-only (Table 3). This is well demonstrated by the curves representing the angle at maximal protraction and amplitude in Figure 3. Furthermore, the values for the angular velocity and angular acceleration parameters did not differ significantly from those of the intact animals. After surgery and implantation of OM, the vibrissae dropped motionless and “rose” at 10–14 d post-operation (DPO). Initial signs of restoration of rhythmical whisking occurred at 21–28 DPO. An almost complete recovery of function was detected 2 months after surgery.
Biometrics of recovering whisking behavior in rats after FFA-only and FFA + OM

Biometric analysis of vibrissae motor performance.A, Graph of the changes in angles of two, large, C-row vibrissae during explorative cyclic whisking in intact rats. The parallel course of both curves indicates the synchronous movements of the two vibrissae. In this case, the frequency was 6 Hz, the protraction was ∼55°, the retraction was ∼105°, and the amplitude was ∼50°. B, Faint and desynchronized whisker movements (protraction and retraction with very small amplitude) during an active exploration of a representative animal 2 months after FFA-only. C, Synchronous movement of two large vibrissae with a frequency of 5 Hz, a protraction of ∼50°, and an amplitude of ∼40° in a representative animal 2 months after FFA.
After FFA-only or FFA + BMM, the vibrissae dropped and acquired a caudal orientation. At 10–14 DPO, the vibrissae rose again to the level of the mouth and acquired a posterior orientation. No signs of restoration of rhythmical whisking were observed.
This is the first demonstration indicating that transplanted tissue exerts beneficial effects on facial nerve regeneration. Recent experiments have shown that neither entubulation in chambers containing agents known to foster neurite regrowth (Dohm et al., 2000) nor implantations of cultured OECs (Guntinas-Lichius et al., 2001) or Schwann cells (our unpublished observations) are able to promote similar recovery. Because there was no evident functional improvement after transplantation of BMM, we feel confident that this beneficial effect is not caused by a purely mechanical role of any transplanted tissue but solely by the transplanted OM. At the same time, this is why our present results bear particular relevance not only for the basic processes underlying axonal regeneration and pathfinding, which are important for developing novel cell-based therapeutic strategies of nerve injury, but also for the general use of olfactory ensheathing glia in restoration studies.
Identification of the transplanted olfactory mucosa at the injury site using congenic markers
Hybridization of sections from the transplantation site with a Y chromosome-specific DNA probe labeled a number of cell nuclei directly adjacent to the sutured facial nerve, indicating that the localization of the implant had not changed during the postoperative period. The reaction product was confined exclusively to portions of the cell nucleus. The nuclei of the host fibroblasts and Schwann cells remained blank (Fig. 4). Sometimes fragments of cross-sectioned conchae could be found. Our results, therefore, confirm previous reports on successful transplantation of olfactory epithelium to the anterior chamber of the eye (Barber et al., 1982) and to the brain (Holbrook et al., 2001).

Detection of transplanted olfactory mucosa byin situ hybridization of a Y chromosome-specific cDNA probe on 40-μm-thick longitudinal sections. A, Overview of a section through the facial nerve and transplanted olfactory mucosa. B, In some cases, portions of preserved nasal conchae, derived from a male donor, could be observed adjacent to the parotid gland and the facial nerve.C, High-power magnification of a portion fromA showing numerous donor-derived positive nuclei (arrows) incorporated into the tissue close to the facial nerve. No nuclear staining is present in the fibers of the host levator labii muscle.
Transplanted olfactory mucosa induce a sustained expression of trophic factors at the lesion site
It is well known that with the exception of CNTF, a number of trophic factors (e.g., NGF, BDNF, bFGF, IGF-I, and GDNF) are expressed by the denervated Schwann cells after buccal facial branch transection. In general, the immunoreactivity is detected at 1 d post-axotomy (DPA), reaches a maximum at 5–6 DPA, and after a gradual decline is no longer evident at 21–28 DPA (Heumann et al., 1987; Meyer et al., 1992;Funakoshi et al., 1993).
This is confirmed by the results of the present study, demonstrating no expression of CNTF, IGF-I, and GDNF and only a faint expression of BDNF and bFGF at the suture site 2 months after FFA (Fig.5A,B) or FFA + BMM (Fig. 5C,D).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Expression of trophic factors at the suture/implantation site 2 months after FFA-only, FFA + BMM, or FFA + OM. Longitudinal sections through the facial nerve show a very faint BDNF (A, C) and bFGF (B, D) immunofluorescence after FFA-only and FFA + BMM, respectively. By contrast, 2 months after FFA + OM, the immunofluorescence for BDNF (E, G,H) and bFGF (F) is intensive.
Contrary to this, an intense immunostaining for BDNF (Fig.5E) and bFGF (Fig. 5F) was expressed in OM-transplanted animals. The higher magnification indicated immunoreactivity mostly in the Schwann cells, which were identified by their long and slender cell processes running parallel to the long axis of the nerve (Fig. 5G,H). A parallel immunostaining for S-100 protein, considered as a general marker of Schwann cells, showed that most of the cells that expressed NGF were Schwann cells (data not shown).
This may indicate that the transplanted OM directly or indirectly maintained the expression of trophic factors that otherwise would have been downregulated. Although these observations are in line with some earlier results (Buckland and Cunningham, 1999; Boruch et al., 2001), we cannot confine the immunoreactivity exclusively to Schwann cells, OECs, or basal cells of the olfactory epithelium.
DISCUSSION
The major finding of the present study is that transplantation of olfactory mucosa to the transected facial nerve reduces axonal branching and improves vibrissal motor performance. Implying a direct link between diminished axonal branching and recovery of motor function, our results also show that reinnervation of muscle targets by “foreign” motoneurons can yield good functional parameters, thus revealing the vast possibilities of neuronal plasticity.
Neurobiological importance of post-transectional axonal branching
When an axon is severed, the proximal cut end rapidly closes with a membrane seal (Fishman et al., 1990; Spira et al., 1993) and soon afterward forms a terminal swelling or “end-bulb” (Friede and Bischhausen, 1980; Fried et al., 1991). Within the following 3 hr, numerous axonal branches begin to emerge from the end-bulb and start to elongate (Bisby and Pollock, 1983) at a rate of 1–4 mm/d (Tetzlaff and Bisby, 1989; Fawcett and Keynes, 1990). Under ideal conditions, only one of these branches actually reaches its original target. The navigation of these immature axons is under the control of short-range acting guidance cues, most of them deriving from the distal nerve stump. Thus, post-lesional axonal branching is considered to represent the search of axons for local guidance cues necessary for their navigation (AlMajed et al., 2000). In general, however, these “actively searching” axons not only choose the wrong way, but also branch and project simultaneously along several different fascicles (subdivisions, rami) of a transected nerve trunk (Angelov et al., 1999). Thus, the process of branching, originally aimed at both neuronal and target survival, turns out to be responsible for the failure of axons to reinnervate solely their original domain (Fu and Gordon, 1997).
The combined approach to evaluate the quality of peripheral nerve regeneration
Recently we established simultaneous multiple neuronal labeling to study axonal branching (Angelov et al., 1999). In several earlier experimental sets, we conceptually divided the process of axonal regrowth into “axonal elongation” and “axonal branching.” We hypothesized that an axonal elongation, which had been fostered by local application of extracellular matrix proteins or OECs, might reduce axonal branching and thus improve recovery of function. The subsequent neuron counts showed that despite their known effect to support neurite elongation, these procedures failed to suppress axonal branching in the facial nerve plexus (Dohm et al., 2000;Guntinas-Lichius et al., 2001).
Preoperative and postoperative neuronal labeling as a tool to study the accuracy of target reinnervation
Quantitative estimation after single tracing with HRP provides information solely on the reinnervation status of a muscle target, because the one-tracer approach cannot elucidate the relationship between preoperative and postoperative innervation of target muscles (Angelov et al., 1996; Streppel et al., 1998; Guntinas-Lichius et al., 2000). The sequential application of double retrograde labeling should allow an optimal evaluation of preoperative and postoperative distribution of motoneurons in the same animal, avoiding counting errors caused by inter-individual variability. Our experience shows that the best combination of fluorescent retrograde tracers to study the accuracy of post-transectional muscle reinnervation is a preoperative labeling of the original motoneuronal pool by an injection of 1% FG into the muscle target, followed by a post-operative labeling of all motoneurons innervating the same target after surgery by an injection of 1% FB (Popratiloff et al., 2001).
The biometrics of whisking behavior provides a very sensitive analysis that we recently introduced as a tool to study facial nerve regeneration (Guntinas-Lichius et al., 2001). Under normal physiological conditions, the mystacial vibrissae of the rat are erect with anterior orientation. Their simultaneous sweeps known as “whisking” or “sniffing” (Welker, 1964; Semba et al., 1980) occur 5–11 times per second (Komisaruk, 1970; Carvell and Simons, 1990; Bermejo et al., 1996). The striated muscle fibers mediating protraction form a sling around the rostral aspect of each hair follicle; contraction of these muscles via branches of the facial nerve pulls the base of the follicle caudally, moving the distal aspects of the whisker hair forward. By contrast, retraction of the vibrissae depends primarily on passive elastic properties of deep connective tissue (Dörfl, 1985; Wineski, 1985).
The nature of the beneficial effect of OM transplantation: moderate but long-lasting secretion of trophic molecules at the lesion site
In this paper, we show that transplanted olfactory mucosa improves peripheral nerve regeneration after facial nerve injury. This effect as measured by functional and morphological analysis appears to be based mainly on the reduction of axonal collateral formation. Whether the observed effect was caused directly by the olfactory ensheathing glia present in the lamina propria or by other cells of the transplanted olfactory tissue cannot be determined by the present data. What can be said, however, is that the transplant was still in place at the lesion site at the end of the experiments, as shown by in situhybridization using a Y chromosome-specific DNA probe. The second important point is whether the transplant itself directly exerted the regeneration-promoting effects or whether it may have stimulated beneficial secondary processes. Again, the presented data cannot answer this question unequivocally, mainly because only the outcome of the transplantation was monitored. Immunostaining for trophic factors demonstrated that the lesion site of OM-transplanted animals but not of the operated controls (FFA) was characterized by an increased expression of NGF, BDNF, and FGF-2. Thus, it is conceivable that the transplant prevented the downregulation of trophic factors and that the trophic factors may be responsible for the observed reduction in collateral sprouting and the good functional recovery. It remains to be demonstrated to what extent the OECs of the transplant contributed to the trophic factor expression.
OECs have been shown to promote axon regeneration and remyelination in various model systems (Li et al., 1997, 1998; Ramon-Cuéto et al., 1998). It was found that OECs, contrary to Schwann cells, reduced sprouting of central neurons and stimulated the growth into the distal part of the experimentally transected spinal cord. Contrary to this, transplantation of pure cultures of neonatal OECs to the transected facial nerve, using the same model system as in the present study, resulted in a dramatic increase in axonal sprouting (Guntinas-Lichius et al., 2001).
How is the apparent discrepancy with the present data explained? In our previous study, we used cultured cells of the neonatal rat instead of adult olfactory tissue, which was used in the present study. The expression profile both between neonatal and adult tissue and between cultured cells and intact tissue may account for the differing effects. Recent evidence implies that trophic factors and their receptors become upregulated on dissociation and cultivation of OECs. The expression of the CNTF receptor α subunit, for example, cannot be found in the developing olfactory bulb (Lee et al., 1997) but displays a robust expression in cultured OECs (Wewetzer et al., 2001). The differences in collateral sprouting after transplantation of cultured OECs and olfactory tissue therefore may reflect differing levels of trophic factors. This in turn would suggest that the amount of trophic factor is the critical factor that determines the degree of sprouting at the lesion site. This speculation is underscored by recent findings demonstrating that application of antibodies against trophic factors reduces collateral sprouting in the same model (Streppel et al., 2002).
The application of olfactory tissue that very likely contained fewer OECs than the implants may have induced more moderate effects. Because expression of trophic factors was noted in structures outside the implant itself, it is likely that the transplant induced and maintained trophic factor expression outside the implant.
After axotomy, trophic factors are taken up by regenerating axons to keep the neurons trophically satisfied (Di Stefano et al., 1992;Unsicker et al., 1992; Yan et al., 1992; Friedman et al., 1995;Sendtner et al., 1997). Accordingly, the synthesis of receptors for NGF, BDNF, bFGF, GDNF, CNTF, and IGF-I is upregulated at the lesion site after axotomy (Meyer et al., 1992; Raivich and Kreutzberg, 1993;McMahon and Priestley, 1995; MacLennan et al., 1999). Apart from this “central effect” on neuronal cell bodies (promoting survival), numerous studies report on other “peripheral effects” of the trophic factors. For example, neurotrophins and bFGF have been shown to stimulate and hypermodulate neurite outgrowth, enlarge axon caliber, and induce sprouting of neurons in vitro and in vivo (Aebischer et al., 1989; Unsicker et al., 1993; Laquerriere et al., 1994; Fujumoto et al., 1997; Gallo and Letourneau, 1998; Batchelor et al., 2000; Davies, 2000; Deng et al., 2000; Mamounas et al., 2000).
Thus, we share the opinion that this unnecessarily forced axonal branching was most likely caused by excessive amounts of trophic molecules provided by the cultured OECs. In contrast, our present results show that the transplanted olfactory mucosa may provide a weaker but long-lasting secretion of neurotrophins and bFGF at the lesion site (quantitative evaluations are under way).
In conclusion, we showed that transplanted autologous olfactory mucosa to the sutured perineurium of a peripheral motor nerve significantly improved the quality of target innervation. The reduced axonal branching promoted a better axonal pathfinding, which in turn provided an excellent recovery of function.
Footnotes
- Received March 20, 2002.
- Revision received May 13, 2002.
- Accepted June 6, 2002.
This study was supported by the Deutsche Forschungsgemeinschaft (AN-331/2-1), the Köln Fortune Program of the Faculty of Medicine, University of Cologne, the Jean-Uhrmacher-Foundation, and the Megapharm GmbH, Germany. The skillful technical assistance of I. Rohrmann and D. Felder and the fine photographical work of I. Koch and the precise drawings of J. Ribbers are highly appreciated. We thank N. v. Dornick for editorial help.
Correspondence should be addressed to Dr. Doychin N. Angelov, Anatomical Institute University of Cologne, Joseph-Stelzmann-Strasse 9, 50931 Cologne, Germany. E-mail:angelov.anatomie{at}uni-koeln.de.
- Copyright © 2002 Society for Neuroscience