
Abstract
In the presence of descending modulatory inputs, the stomatogastric ganglion (STG) of the lobster Homarus americanusgenerates a triphasic motor pattern, the pyloric rhythm. Red pigment-concentrating hormone (RPCH) and Cancer borealistachykinin-related peptide (CabTRP) are colocalized in a pair of fibers that project into the neuropil of the STG. When the STG was isolated from anterior ganglia modulatory inputs, the lateral pyloric (LP) and pyloric (PY) neurons became silent, whereas the anterior burster (AB) and pyloric dilator (PD) neurons were rhythmically active at a low frequency. Exogenous application of 10−6m RPCH activated the LP neuron but not the PY neurons; 10−6m CabTRP activated the PY neurons but not the LP neuron. The actions of RPCH on the LP neuron and CabTRP on the PY neurons persisted when the rhythmic drive from the PD and AB neurons was removed, suggesting that the LP and PY neurons are direct targets for RPCH and CabTRP respectively. Coapplication of 10−6m RPCH and 10−6m CabTRP elicited triphasic motor patterns with phase relationships resembling those in a preparation with modulatory inputs intact. In summary, cotransmitters acting on different network targets act cooperatively to activate a complete central pattern-generating circuit.
- cotransmission
- pyloric rhythm
- stomatogastric ganglion
- lobsters
- red pigment-concentrating hormone
- Cancer borealis
- tachykinin-related peptide
- neuromodulation
Although it is well established that many neurons contain multiple cotransmitters (Kupfermann, 1991;Hökfelt et al., 2000), relatively little is understood of the role that cotransmission plays in shaping the dynamics of neuronal circuits. Much of what we know about the functional aspects of cotransmission comes from studies on peripheral synapses (Jan et al., 1979; Jan and Jan, 1982; Whim and Lloyd, 1989, 1990; Weiss et al., 1992; Vilim et al., 1996a,b). Although it is known that more than one transmitter can be costored or coreleased at many CNS synapses (Hökfelt et al., 2000), relatively little is known about the effects that such cotransmission causes on postsynaptic targets.
The functions of cotransmission at the network level can be studied using the stomatogastric nervous system (STNS) of decapod crustacea. The 26–30 neurons of the stomatogastric ganglion (STG) generate robust gastric mill and pyloric rhythms (Harris-Warrick et al., 1992; Marder and Calabrese, 1996) when the anterior ganglia that send modulatory projections into the STG are left intact. Many of these modulatory projections contain multiple neurotransmitters (Christie et al., 1997a; Blitz et al., 1999; Fénelon et al., 1999; Kilman et al., 1999; Meyrand et al., 2000), thus providing the opportunity to study the coordinated actions of cotransmitters on a well defined neuronal circuit.
In this paper we explore the actions of two neuropeptides, red pigment-concentrating hormone (RPCH) and Cancer borealistachykinin-related peptide (CabTRP) as functional cotransmitters. RPCH is an octapeptide (pELNFSPGW-NH2) that was isolated and sequenced from the shrimp Pandalus borealis(Fernlund and Josefsson, 1972). RPCH-like immunoreactivity is present in the STNS (Nusbaum and Marder, 1988; Dickinson and Marder, 1989;Fénelon et al., 1999). Apart from modulating the motor patterns produced by the STNS (Nusbaum and Marder, 1988; Dickinson and Marder, 1989; Dickinson et al., 1990, 2001), RPCH also affects other motor patterns such as the crayfish swimmeret rhythm (Sherff and Mulloney, 1991). CabTRP Ia and Ib are two tachykinin-related peptides that were isolated from C. borealis (Christie et al., 1997b) and are responsible for the tachykinin staining reported previously (Goldberg et al., 1988; Blitz et al., 1995). CabTRP Ia is also a strong modulator of the gastric mill and pyloric rhythms (Blitz et al., 1995; Wood et al., 2000).
In Homarus americanus, RPCH and CabTRP are colocalized in projections into the STG neuropil. The presence of authentic RPCH and CabTRP in the STNS of H. americanus has been confirmed using matrix-assisted laser desorption/ionization mass spectroscopy (MALDI-MS; Li, 2000). Here we present evidence that shows that RPCH and CabTRP alter the excitability of different target neurons within the pyloric network of H. americanus. The two cotransmitters are both needed to fully activate the canonical triphasic pyloric rhythm.
Parts of this paper have been presented previously in abstract form (Thirumalai and Marder, 2001).
MATERIALS AND METHODS
Adult H. americanus (n = 34) were obtained from Commercial Lobster (Boston, MA) and maintained in artificial seawater tanks at 11°C. Lobsters were anesthetized on ice for 15 min. The complete STNS (Fig. 1A), consisting of the paired commissural ganglia (CoGs), esophageal ganglion (OG), and the STG and their connecting and motor nerves, was dissected and pinned out in a transparent Sylgard (Dow Corning, Midland, MI) dish containing chilled (9–13°C) saline. Saline composition was (in mm): 479.12 NaCl, 12.74 KCl, 13.67 CaCl2, 20 MgSO4, 3.91 Na2SO4, and 5 HEPES, pH 7.45.
For electrophysiological recordings, the STG was desheathed, and Vaseline wells were made on the motor nerves for extracellular nerve recordings (Fig. 1A). Intracellular recordings from the STG motor neuron somata were made using 20–40 MΩ electrodes filled with 0.6 mK2SO4 and 20 mm KCl and an Axoclamp 2A amplifier (Axon Instruments, Foster City, CA). The lateral pyloric (LP), pyloric (PY), and pyloric dilator (PD) neurons were identified using standard procedures (Selverston and Moulins, 1987). Figure 1A,inset, shows a simplified connectivity diagram for the pyloric rhythm indicating the known synaptic interactions among the neurons relevant for this paper.
The STG was isolated from descending inputs from the CoGs and OG by blocking impulse traffic in the stomatogastric nerve (stn) by filling a Vaseline well made on the stn with 750 mm sucrose and 10 μm TTX. During recordings the preparations were continuously superfused with chilled (11°C) physiological saline, and RPCH (American Peptide Company, Sunnyvale, CA) and CabTRP (Protein Chemistry Laboratory, University of Pennsylvania, Philadelphia, PA) were introduced with a switching manifold into the bath. For some experiments saline containing 10−5mpicrotoxin (PTX; Sigma, St. Louis, MO) was used.
Data acquisition and analysis. Data were acquired using a Digidata 1200 data acquisition board (Axon Instruments) and analyzed using programs written by Dr. Bill Miller (VegaScientific,www.vegasci.com) and Microsoft Excel. Analyzed data were plotted, and statistical tests were performed in SigmaStat (SPSS, San Rafael, CA). In all cases, the pyloric rhythm was characterized using the burst period, the number of spikes in a burst, the duration of the burst, the phase relationship of bursts with respect to the PD neuron, and the duty cycles of each type of pyloric neuron. Data are reported as the mean ± SE. Statistical significance was assessed using a repeated measures one-way ANOVA test followed by the Tukey test when the ANOVA yielded a significant p value.
Immunocytochemistry and confocal microscopy. Whole-mount immunocytochemistry was performed according to published procedures (Kilman et al., 1999). The stomatogastric nervous system was dissected out and pinned on a Sylgard dish. Preparations were fixed with 4% paraformaldehyde in 0.1 m sodium phosphate buffer, pH 7.4, overnight at 4°C and washed several times with 0.1 msodium phosphate buffer containing 0.3% Triton X-100 detergent and 0.1% sodium azide (PTA). Preparations were next incubated overnight at 4°C with rabbit anti-RPCH (a gift from Dr. Hugo Arechiga, Universidad Nacional Autonoma de Mexico, Mexico City, Mexico) and rat anti-substance-P (Accurate Chemical and Scientific Co., Westbury, NY) antibodies at a dilution of 1:100 each in PTA. The rat anti-substance-P antibody specifically labels CabTRP Ia because preincubation of the antiserum with 10−4m CabTRP Ia abolishes all substance-P-like immunoreactivity (Fénelon et al., 1999). After another round of several washes with PTA, preparations were incubated overnight at 4°C with goat anti-rat IgG coupled to Alexa Fluor 488 (Molecular Probes, Eugene, OR) and Goat anti-rabbit IgG coupled to Cy5 (Jackson ImmunoResearch, West Grove, PA) at a dilution of 1:300 each in PTA. Finally, preparations were washed with 0.1 m sodium phosphate buffer, pH 7.4, and mounted on a glass slide with 80% glycerol.
Preparations processed for immunohistochemistry were imaged on a Leica (Nussloch, Germany) TCS NT confocal imaging system with 10 and 20× air interface objective lenses. For all images, a Z series, in which images were captured at depth separations of 5 μm, was acquired. A maximum projection of the Z series images was made. Images were stored both as individual Z series projections and as projections. These images were then processed with standard image processing software (Adobe Photoshop 6.0 and Confocal Assistant). Images were then printed out in false color.
RESULTS
Pyloric rhythm in H. americanus
In decapod crustacea such as the lobster, H. americanus, the STNS produces characteristic motor patterns that move the stomach musculature to allow the animal to ingest and process food (Selverston and Moulins, 1987). The fastest of these rhythms, the pyloric rhythm, is responsible for the constriction and dilation of muscles belonging to a narrow chamber at the posterior end of the stomach known as the pylorus. The pyloric rhythm consists of the rhythmic and repetitive firing of the PD, LP, and PY motor neurons, as shown in the simultaneous intracellular recordings in Figure1B. As in other decapod crustacea, the STG in H. americanus has a single anterior burster (AB) interneuron that is electrically coupled to the PD neurons and with them forms the kernel of the pacemaker ensemble. When the PD and AB neurons depolarize and fire together, they in turn inhibit the LP and PY neurons (Fig. 1A). The only feedback from the follower neurons to the PD and AB neurons comes from the synapse from the LP to the PD neurons (Fig. 1A).

Schematic of the stomatogastric nervous system and the pyloric rhythm it produces. A, The STNS with the anterior ganglia, the paired CoGs, and the single OGs attached to the STG via the stn. The gray circle on the stn indicates the position of the Vaseline well used for blocking action potential conduction along thestn. The pyloric rhythm was recorded through extracellular nerve recordings from the motor nerves low lateral ventricular nerve (llvn), pylonic dilator nerve (pdn), and pyn. The motor neurons PD, LP, and PY within the STG were impaled using sharp glass electrodes and identified by triggering their action potentials and following them on identified motor nerves. Synaptic connections between pyloric neurons are shown in the inset. Connections ending infilled circles represent inhibitory synaptic connections. Resistor symbols represent electrical synapses. B, When the stn is intact, the pyloric neurons AB, PD, LP, and PY burst in a characteristic triphasic pattern with a frequency of ∼0.6 Hz. Horizontal barsto the right of each intracellular tracemark the −60 mV position. C, Intracellular recordings from the pyloric neurons AB, PD, LP, and PY from the same preparation as in B but with the stn blocked. TheLP and PY traces show IPSPs from AB and PD neurons whenever the AB and PD neurons produce bursts.Horizontal bars to the right of eachtrace mark the −60 mV position.
The pyloric neurons in the STG receive modulatory inputs from the paired CoGs and the single OG via the stn (Fig. 1A). Figure1B shows typical pyloric rhythms when the anterior inputs to the STG were left intact. Under these conditions, in H. americanus, the pyloric rhythm had a frequency of 0.64 ± 0.02 Hz (n = 27), somewhat slower than the ∼1 Hz rhythms recorded from other decapod crustacea such as C. borealis or Panulirus interruptus. In the experiment shown in Figure 1, when the anterior inputs were removed by placing a conduction block on the stn (see Materials and Methods), the AB and PD neurons continued to burst slowly (Fig. 1C), and the LP and PY neurons stopped firing but showed IPSPs evoked by the AB and PD neurons (Fig. 1C). The PD burst frequency with anterior inputs removed was 0.17 ± 0.02 Hz (n = 27; p < 0.001 when compared with the intact preparations). The decrease in the frequency of the PD burst after anstn block typically took 10–20 min to take effect. After this transition time, the lower PD burst frequency was stable for many hours.
Colocalization of RPCH and CabTRP in inputs to the STG
Figure 2A shows RPCH- and CabTRP-like immunoreactivity in the neuropil of the STG ofH. americanus. Figure 2A, left, shows RPCH-like immunoreactivity in red. There are thick varicosities and small punctate varicosities within the neuropil of the STG that stain for RPCH. There are also fibers seen entering from thestn into the neuropil. The CabTRP immunoreactivity (green) appears only in the thick varicosities in the neuropil and in fibers entering via the stn. Figure2A, right, shows that the RPCH and CabTRP staining colocalize in the thick varicosities when the two are merged (shown in yellow). However, RPCH-like immunoreactivity in the small punctate varicosities contains no CabTRP-like immunoreactivity.

Colocalization of RPCH and CabTRP in inputs to the STG. A, STG double labeled for RPCH and CabTRP.Left panel, False-color pattern of staining produced by rabbit anti-RPCH antibody. Middle panel, Pattern of staining produced by rat anti-substance P antibody. Right panel, Merge of the two images illustrating the colocalization of RPCH and CabTRP in yellow. Scale bar, 50 μm.B, Schematic of RPCH and CabTRP immunoreactivity in the entire STNS. Redrawn from Fénelon et al. (1999). ion, Inferior esophageal nerve; ivn, inferior ventricular nerve;dvn, dorsal ventricular nerve.
Figure 2B shows a schematic drawing of RPCH- and CabTRP-like immunoreactivities seen in other parts of the STNS. All of the RPCH-like staining in the neuropil of the STG derives from four fibers, two each from each of the CoGs projecting down the superior esophageal nerve (son) and then the stn. One each of these fibers also contains CabTRP. There are several cell bodies in the CoG that stain for RPCH. There are also several CabTRP-positive cell bodies, but it remains to be seen which of those cell bodies stain for both. There is also a cell body in the OG that stains for CabTRP and projects bilaterally to the paired CoGs via the inferior esophageal nerve.
Effects of RPCH application
In H. americanus, RPCH and CabTRP are colocalized in processes that ramify within the neuropil of the STG (Kilman, 1998;Fénelon et al., 1999). In this section, we describe the effects of application of RPCH. In the next section, we describe the effects of CabTRP and then we describe the effects of coapplications of RPCH and CabTRP and compare these with the effects of the singly applied peptides.
Figure 3A shows simultaneous recordings from the PD, LP, and PY neurons in a preparation isolated from anterior inputs (henceforth referred to as the “isolated preparation”) in normal saline. Figure 3B shows the same preparation in the presence of 10−6m RPCH. RPCH strongly activated the LP neuron from its previous silent state, which in this preparation markedly decreased the frequency of the rhythm. Although the PD and LP neurons were strongly activated in RPCH, the PY neuron remained silent (Fig.3B).

Modulation of the pyloric network by 10−6m RPCH. A, Intracellular recordings from PD, LP, and PY neurons in normal saline when the stn was blocked. B, Recordings from the PD, LP, and PY neurons from the same preparation when saline containing 10−6m RPCH was superfused.Horizontal bars to the right of eachtrace indicate the −60 mV position.
Figure 4 summarizes data pooled from 21 experiments to summarize quantitatively the effects of RPCH. The effects of RPCH on the frequency of the PD burst were not statistically significant across the preparations, because in some cases the frequency increased, in others it decreased, and in yet others it was unchanged (Fig. 4A). Figure 4Bshows that in 10−6m RPCH, the number of spikes produced by the PD neuron per burst increased significantly (control, 13 ± 1.5; RPCH, 42 ± 5.9; n = 21; p < 0.001). RPCH also increased the number of spikes per burst produced by the LP neuron (control, 0.09 ± 0.06; RPCH, 27 ± 6;n = 21; p < 0.001; Fig.4C). There was no change in PY neuron activity (Fig.4D). The effects of bath application of 10−6m RPCH were reversed by wash with normal saline.

Effect of 10−6mRPCH on firing properties of pyloric neurons. A, PD burst frequency in normal saline (control frequency) plotted against PD burst frequency in 10−6m RPCH. B, The number of spikes produced by the PD neuron in 10−6m RPCH is significantly higher than that in normal saline (Control) or that when the peptide was washed out with saline (Wash).C, RPCH (10−6m) also significantly increases the number of spikes per burst produced by the LP neuron from those in normal saline (Control). LP spikes per burst returned to control values during wash (Wash). D, RPCH (10−6m) did not affect the duration of bursts produced by the PY neurons. Bars indicate mean values from 21 preparations. ***Significance level of p = 0.001. Error bars indicate SE.
Effects of CabTRP application
The effects of 10−6mCabTRP on the pyloric network neurons were quite unlike those of RPCH. Figure 5A shows simultaneous intracellular recordings from the PD, LP, and PY neurons and an extracellular nerve recording from the pyloric nerve (pyn) (see Fig. 1A) in an isolated preparation in normal saline. When saline containing 10−6m CabTRP was applied, the PD neuron bursts increased in frequency, the LP neuron remained silent, and the PY neurons started bursting alternately with the AB/PD group (Fig.5B). Data from 19 experiments were pooled to analyze these results statistically. The PD neuron burst frequency increased significantly in 10−6m CabTRP (control, 0.16 ± 0.02/sec; 10−6m CabTRP, 0.31 ± 0.03/sec; n = 19; p < 0.001; Fig. 6A). The number of PD spikes per burst increased from 14.6 ± 1.7 in normal saline to 36.5 ± 7 in 10−6m CabTRP (n = 19;p < 0.005; Fig. 6B). Although CabTRP significantly increased the duration of the PY neuron burst (Fig.6D) from 0.05 ± 0.05 sec in normal saline to 0.91 ± 0.18 sec in 10−6m CabTRP (n = 19;p < 0.001), the LP neuron remained silent (Fig.6C). The effects of CabTRP were reversible and were washed out by superfusing the preparation with normal saline.

Modulation of the pyloric neurons by 10−6m CabTRP. A, Recordings from the PD, LP, and PY neurons in normal saline when conduction of action potentials along the stn was blocked. Bottom trace, Extracellular nerve recording from the pyn nerve. In normal saline, pynis silent. B, Recordings from the same preparation when 10−6m CabTRP was superfused. Thepyn shows bursts produced by the PY neurons.Horizontal bars next to each intracellular recording indicate the −60 mV position.

Effect of 10−6mCabTRP on firing properties of pyloric neurons. A, CabTRP (10−6m) significantly increases the bursting frequency of the PD neurons from the values found in normal saline (Control). This increase in frequency vanished when bath-applied CabTRP was washed out of the preparation (Wash). B, CabTRP (10−6m) significantly increased the number of spikes produced by the PD neuron when compared with the values obtained in normal saline (Control). This increase disappeared when CabTRP was washed out with normal saline (Wash). C, CabTRP (10−6m) did not alter the number of spikes produced by the LP neuron per burst. D, The durations of bursts produced by the PY neurons in 10−6m CabTRP (CabTRP) were significantly higher than those in normal saline (Control) or those when CabTRP was washed out (Wash). Bars indicate mean values from 19 preparations. **Significance level of p = 0.01; ***p = 0.001. Error bars indicate SE.
Targets of RPCH and CabTRP within the pyloric network
Given that the colocalized peptides RPCH and CabTRP activated different subsets of pyloric neurons, the question remained whether the actions of the two peptides were attributable to direct actions of RPCH and CabTRP on the LP and PY neurons, respectively. In the STNS, glutamatergic synaptic inhibition can be blocked by 10−5m PTX (Bidaut, 1980;Eisen and Marder, 1982; Marder and Eisen, 1984a). This isolates the LP and PY neurons from mutual inhibition and also from inhibition from the AB interneuron (Bidaut, 1980; Eisen and Marder, 1982; Marder and Eisen, 1984a). In Figure 7, we hyperpolarized the AB and PD neurons (traces not shown) while recording intracellularly from the LP and PY neurons in saline containing 10−5m PTX. Under these conditions, the membrane potentials of the LP and PY neurons were flat because the inhibition from the pacemaker kernel was absent [Fig.7A(i),B(i)]. Bath application of 10−6m RPCH in PTX activated slow bursts in the LP neuron, whereas the PY neurons remained silent [Fig. 7A(ii)]. Similar results were obtained in three different preparations. In response to 10−6m CabTRP in PTX, the LP neuron remained silent, but the PY neuron started to burst slowly [Fig. 7B(ii)]. Similar results were obtained when this experiment was repeated in four different preparations.

Targets of RPCH and CabTRP modulation.A(i), Intracellular recordings from the LP and PY neurons in saline containing 10−5mpicrotoxin after the AB and PD neurons were hyperpolarized.A(ii), Intracellular recordings from the same preparation in saline containing 10−5mpicrotoxin and 10−6m RPCH while still hyperpolarizing the AB and PD neurons. B(i), Membrane voltage traces from the LP and PY neurons in saline containing 10−5m picrotoxin after the AB and PD neurons were hyperpolarized. B(ii), Membrane potential traces from the same neurons in the same preparation in saline containing 10−5m picrotoxin and 10−6m CabTRP. Horizontal bars next to the traces indicate the position of the −60 mV mark.
Coapplication of CabTRP and RPCH
Because CabTRP and RPCH are colocalized, we wished to determine the effects of coapplication of these substances. Figure8 shows the effects of coapplication of 10−6m CabTRP and 10−6m RPCH to an isolated STG. Note that in the presence of the peptides, both the LP and PY neurons were activated, and the full triphasic motor pattern was produced. Figure 9 summarizes the effects on the motor pattern in data pooled from 21 preparations. There was a slight tendency for the frequency of the pyloric rhythm to increase (Fig. 9A) but this did not reach significance. However, the number of PD spikes per burst did increase (isolated STG in normal saline, 13.6 ± 1.3 PD spikes per burst; in 10−6m CabTRP and 10−6m RPCH, 39.9 ± 4.8 PD spikes per burst; n = 21;p < 0.001; Fig. 9B). The number of LP neuron spikes per burst also increased (isolated STG in normal saline, 0 ± 0 LP spikes per burst; in 10−6m CabTRP and 10−6m RPCH, 17.9 ± 2.8 LP spikes per burst;n = 21; p < 0.001; Fig.9C). There was also an increase in the burst duration of the PY neurons (isolated STG in normal saline, 0 ± 0 sec; in 10−6m CabTRP and 10−6m RPCH, 0.94 ± 0.16 sec; n = 21; p < 0.001; Fig. 9D).

Activation of the pyloric rhythm in an isolated STG by 10−6m RPCH and 10−6m CabTRP. A, Intracellular recordings from the PD, LP, and PY neurons in normal saline after the stn has been blocked. B, Recordings from the same neurons as in A but in saline containing 10−6m RPCH and 10−6m CabTRP. The −60 mV position for each intracellular trace is marked by ahorizontal bar next to it.

Modulation of firing properties of pyloric neurons by coapplication of 10−6m RPCH and 10−6m CabTRP. A, Mean PD neuron bursting frequency in an isolated STG when in normal saline (Control), when in 10−6m RPCH and 10−6m CabTRP (R-C), and when the two peptides were washed out with normal saline (Wash). The PD neuron mean bursting frequency is not significantly different from that in control.B, Mean number of spikes produced by the PD neuron in an isolated STG when in normal saline (Control), in 10−6m RPCH and 10−6m CabTRP (R-C), and when the peptides were washed out with normal saline (Wash). C, Mean number of spikes per burst produced by the LP neuron in an isolated STG when in normal saline (Control), when in 10−6m RPCH and 10−6m CabTRP (R-C), and when the two peptides were washed out with normal saline (Wash). D, Mean duration of PY bursts in an isolated STG when in normal saline (Control), when in 10−6m RPCH and 10−6m CabTRP (R-C), and when RPCH and CabTRP were washed out with saline (Wash). Mean values for each parameter came from measurements from 21 preparations. ***Significance level ofp = 0.001. Error bars indicate SE.
Modulation of phase relationships by RPCH and CabTRP
In comparing motor patterns, it is often useful to normalize the motor patterns to ask how relative phase relationships of the constituent neurons change independent of burst frequency. We created phase plots for the motor patterns produced in RPCH, in CabTRP, and in both and compared these with the phase plots generated in the intact preparation and in the stn-blocked case in control saline. In these plots the onset of the PD neuron burst was used as the reference point, and the start and end of the bursts were calculated relative to the PD neuron start. The duty cycle of the activity of each neuron was calculated by dividing the burst duration of the neuron by the cycle period.
Figure 10A shows the phase relationships of the PD, LP, and PY neurons in normal saline when the stn was blocked (white bars), in RPCH (cross-hatched bars), or in normal saline when thestn was intact (black bar). In control saline with the stn blocked, only the PD neuron was active. The phase plots show that the PD duty cycle was significantly smaller (0.08 ± 0.02; n = 10) than seen in the intact preparation (0.35 ± 0.02; n = 10;p < 0.001) or in the presence of 10−6m RPCH (0.22 ± 0.04; n = 10). As described previously, the LP neuron was activated in RPCH. In 10−6m RPCH, the LP neuron had a duty cycle of 0.42 ± 0.06, significantly longer than seen in stn-intact preparations (0.27 ± 0.02;n = 10; p < 0.01).

Modulation of phase relationships between pyloric neurons by RPCH, CabTRP, and both. The phase of the pyloric cycle during which each neuron is spiking is indicated by arectangle. The length of this rectangle represents the duty cycle of the neuron. The start and end positions of the rectangle along the x-axis indicate, respectively, the on-phase and off-phase of that neuron (see Results). In each phase plot, thetop group of rectangles represents the on-phase, off-phase, and duty cycle of PD neurons; the middle group of rectangles indicates the LP neuron on-phase, off-phase, and duty cycle; and the bottom group of rectangles indicates phase data for the PY neuron group. Significant differences are indicated by asterisks placed at the beginning of a bar (to indicate change in on-phase), middle of abar (to indicate change in duty cycle), orend of a bar (to indicate change in off-phase). *Significance level of p = 0.05; **p = 0.01; ***p = 0.001. Error bars indicate SE. A, Modulation of phase relationships between pyloric neurons by 10−6m RPCH.White bars are phase relationships in normal saline when the anterior inputs were removed. Because the LP and PY neurons are silent after the STG is isolated, there are no white bars next to these two neurons in the phase plot. Cross-hatched bars are phase relationships in 10−6m RPCH.Black bars are phase relationships in normal saline in the same preparations when the anterior inputs were attached. These values are plotted to be able to compare phase relationships in RPCH with those in an intact rhythm. Phases plotted are mean values from 10 experiments. B, Modulation of phase relationships between pyloric neurons by 10−6mCabTRP. When the STG is isolated, only the PD neurons are active (white bars). When 10−6m CabTRP is superfused, PD and PY neurons burst (gray bars), whereas in normal saline, when the STG is connected to the anterior inputs, PD, LP, and PY neurons are active (black bars). Phases plotted are mean values from 10 experiments. C, Modulation of phase relationships between pyloric neurons by 10−6m RPCH and 10−6m CabTRP. White bars represent phase data of pyloric neurons in normal saline when the STG is isolated from anterior inputs. Cross-hatched gray bars indicate phase data obtained for pyloric neurons in an isolated STG when saline containing 10−6m RPCH and 10−6m CabTRP was superfused. These data are compared with data obtained from the same preparations when the stn was intact (black bars). Phase data plotted are means of values obtained from 11 preparations.
Figure 10B illustrates the changes that occur in 10−6m CabTRP. CabTRP activates the PD and PY neurons such that their activity patterns resemble those seen in normal saline when the stnis intact (Fig. 10B). There were no significant differences between the stn-intact and the CabTRP conditions in the phases of PD off and PY on. The PY-off phase in 10−6m CabTRP was significantly different from the PY-off phase in normal saline when thestn was intact (stn-intact, 1.07 ± 0.02; CabTRP, 0.98 ± 0.02; n = 10; p < 0.001).
In 10−6m RPCH and 10−6m CabTRP, a full triphasic motor pattern is produced. Figure 10C shows that this pattern closely resembles the triphasic motor pattern produced with the anterior inputs intact. Only the PY-off phase in 10−6m CabTRP and 10−6m RPCH (0.96 ± 0.03) was significantly different from the corresponding values in the stn-intact condition in normal saline (1.05 ± 0.02; n = 11; p < 0.005).
DISCUSSION
Despite the ever-growing literature establishing the existence of multiple neurotransmitters and neuromodulators in neurons involved in all aspects of behavior (Kupfermann, 1991; Hökfelt et al., 2000;Nusbaum et al., 2001), the importance of cotransmission in shaping the dynamics of neural systems has been mostly neglected. This is partially attributable to the fact that many of the synapses or targets amenable to cellular analyses of cotransmission are found in the periphery (Jan et al., 1979; Jan and Jan, 1982; Adams and O'Shea, 1983; Bishop et al., 1987; Brezina and Weiss, 1997; Thorne and Horn, 1997). The studies at the periphery established that peptide cotransmitters could be released at a distance from their targets (Jan et al., 1979; Jan and Jan, 1982), that different patterns of presynaptic stimulation might result in qualitative changes in the nature of the synaptic potential evoked by the presynaptic neuron (Adams and O'Shea, 1983; Marder, 1998), and that given neural targets could display receptors to several cotransmitters (Adams and O'Shea, 1983; Vilim et al., 1996a,2000).
Presumably, similar mechanisms of cotransmission are present at synapses between neurons in the CNS and could play vital roles in modulating the networks in which these neurons are present. Because of the structural complexity and high degree of connectivity in CNS networks, studying cotransmission at these synapses has proved difficult if not impossible. The crustacean STNS is advantageous for asking questions pertaining to cotransmission at central synapses. The STNS has several advantages, not the least of which are the small number of identifiable neurons in the STG and knowledge of all synaptic connectivity within the STG neurons. Added to this, using immunohistochemistry and MALDI-MS, it has been possible to study the patterns of colocalization of neuroactive substances within the STNS. In the past, the STNS of crustacea has been successfully used to study cotransmission in functional networks. For example, the modulatory proctolin neuron (MPN) present in the OG, projects to targets present in three ganglia: the two CoGs and the single STG. Although MPN contains the small molecule GABA and the neuropeptide proctolin, it appears to liberate only GABA on its postsynaptic targets within the CoG; although these targets have receptors for proctolin, they receive only GABAergic inputs from MPN (Blitz and Nusbaum, 1999). MPN actions in the CoG modulate the gastric mill rhythm, whereas its actions in the STG modulate the pyloric rhythm (Blitz and Nusbaum, 1999). MPN actions on the pyloric circuit are mimicked by bath application of proctolin (Nusbaum and Marder, 1989). This is an example in which cotransmitters modulate different circuits.
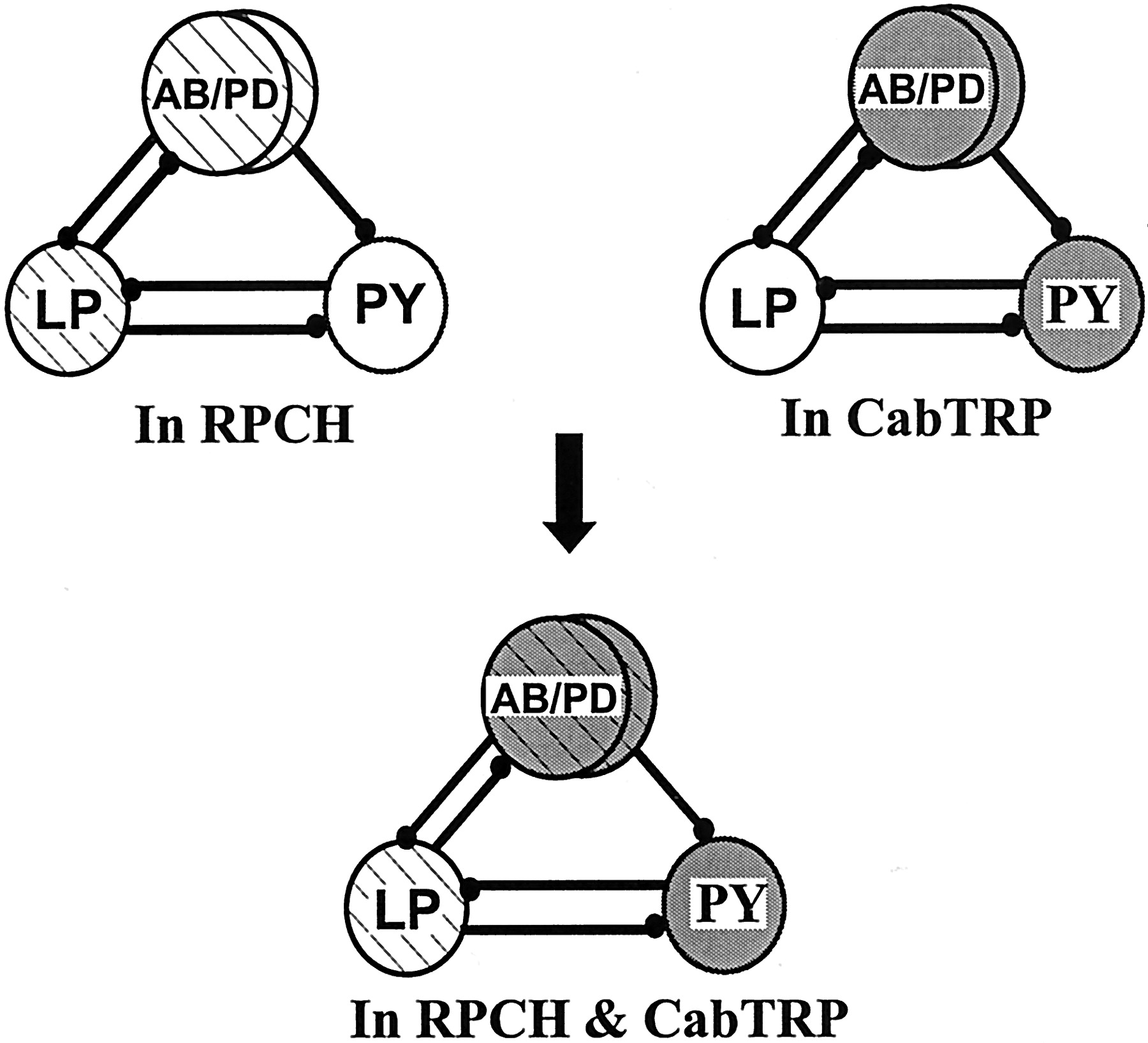
In this paper, we show that a modulatory neuron can use its cotransmitters to activate different targets within the same circuit. In Figure 11, we show a schematic of the pyloric circuit, the model network that we used for our studies. The schematic shows that in RPCH, the activity patterns of the AB/PD and LP neurons are modulated (circles withcross-bars); in CabTRP, the AB/PD and PY neurons are modulated (gray circles); but when both peptides are present, the AB/PD, LP, and PY neurons are activated (gray circles with cross-bars), and a triphasic PD, LP, and PY rhythm results. Thus we demonstrate a mechanism of divergence of cotransmitter actions, which results in the activation of all members of a network.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Activation of neurons belonging to the pyloric network by RPCH and CabTRP. The pacemaker kernel AB/PD and the LP neuron are targets of RPCH modulation (cross-hatched circles). CabTRP targets the AB/PD group and the PY neuron group (gray circles). When these two peptides are coapplied, all three groups of neurons are activated, resulting in a triphasic pyloric rhythm (cross-hatched gray circles).
Individual actions of RPCH and CabTRP in the STNS of other decapod crustacea
RPCH and CabTRP are colocalized in the STNS of H. americanus but not in the STNS of C. borealis (Christie et al., 1997a). What are the effects produced by these peptides on the pyloric rhythm in species where they are not found colocalized? In bothP. interruptus and C. borealis, as in H. americanus, RPCH activates the LP and PD neurons (Nusbaum and Marder, 1988; Dickinson et al., 2001). Additionally, in P. interruptus, RPCH strongly activates the cardiac sac rhythm (Dickinson and Marder, 1989; Dickinson et al., 1990, 1993), resulting in interruptions in the LP bursting pattern that are phase-locked to the cardiac sac bursts (Dickinson et al., 2001). Similar interruptions occur in the H. americanus LP motor pattern in RPCH, most pronounced when the anterior ganglia are left connected to the STG (data not shown). In contrast, the effects of CabTRP in H. americanus are different from those in C. borealis, where it strongly activates the LP neuron (Christie et al., 1997b; Wood et al., 2000).
Identity of the cotransmitting modulatory neuron
Although we know that the cotransmitters RPCH and CabTRP are present in a single fiber projecting down each son and then the stn, we do not yet know the location of the cell body that gives rise to these fibers. It is possible that this cell is one of the many cell bodies staining for RPCH and CabTRP in the CoGs. Also not known is whether this neuron also contains a classic small-molecule neurotransmitter such as glutamate, ACh, or GABA. Of the substances known to be present in the projections within the neuropil of the STG, it is clear that proctolin, allatostatin, and serotonin (Fénelon et al., 1999; Kilman et al., 1999) are not present within this neuron. The possibility that this projection neuron is purely peptidergic exists, because previous ultrastructural studies have revealed the existence of neurohemal profiles with only dense core vesicles and no small clear vesicles (Kilman and Marder, 1996). Regardless, it would be interesting to determine whether CabTRP, RPCH, and any other possible cotransmitters are released stoichiometrically at all stimulation frequencies, as is the case for peptide cotransmitters of theAplysia B15 neuron (Vilim et al., 1996a,b).
Cellular mechanisms of RPCH and CabTRP
When peptides activate a target neuron, they modulate its intrinsic and synaptic properties. Although we have not determined the biophysical mechanisms responsible for RPCH and CabTRP actions inH. americanus, in C. borealis RPCH and CabTRP converge to activate the same voltage-dependent inward current (Swensen and Marder, 2000). Nonetheless, RPCH and CabTRP produce different circuit outputs by acting on a different subset of neural network targets (Swensen, 2000; Swensen and Marder, 2001). In our work, it is clear that the receptors for RPCH and CabTRP are differentially distributed on the different pyloric neurons, because even bath applying the modulators to the whole preparation activated only a specific subset of neurons.
If RPCH and CabTRP converge on the same current in H. americanus, then the two cotransmitters, if released at high concentrations, might saturate the actions of each other. Although both RPCH and CabTRP act on the pacemaker group, their actions on the AB/PD complex are different: CabTRP, but not RPCH, increases the frequency of the AB/PD bursts. It is possible that RPCH and CabTRP activate the same current, but within the pacemaker group the neuronal targets for the two peptides are different, as has been seen previously for other modulators (Marder and Eisen, 1984b; Ayali and Harris-Warrick, 1999). Alternatively, either RPCH or CabTRP may act on still another membrane current.
Summation of individual effects in mixtures
Here we have described data that show that the effects of RPCH and CabTRP are equal to the sum of their individual effects. This result is not necessarily an obvious one, because there are many reasons why a modulator mixture would not produce the “sum” of its parts. In a number of systems, the effect of one of the cotransmitters is to modulate the response produced by the other. In such a case, the modulating transmitter may not have a response on its own, but its physiological actions are seen only in conjunction with the other transmitter. For example, dopaminergic neurons of the ventral tegmental area also contain glutamate. When placed in microcultures, these neurons make glutamatergic autapses that are modulated by dopamine released by these neurons. The dopamine is released from nonsynaptic release sites and decreases glutamate response via a presynaptic D2 receptor-mediated mechanism (Sulzer et al., 1998). Whether this occurs in vivo depends on whether the presynaptic terminals are close to the dopamine release sites. Spinal dorsal horn neurons use ATP and GABA as cotransmitters. GABA acts via GABAA receptors to cause fast IPSCs. ATP, apart from acting via P2X receptors to produce a fast EPSC, gets degraded to adenosine in the extracellular space that inhibits GABAA currents (Jo and Schlichter, 1999). In both of these cases, the actions of the cotransmitters will not simply sum.
Developmental acquisition of RPCH and CabTRP
During development, RPCH is acquired first, as early as 50% of embryonic development. CabTRP immunoreactivity is first seen during the second larval stage (Kilman, 1998; Fénelon et al., 1999). Early in development, the STG generates a single “embryonic” rhythm (Casasnovas and Meyrand, 1995; Le Feuvre et al., 1999), possibly because of increased electrical coupling. It is not clear whether neurons change their complement of receptors to neuropeptides over development, but if the LP and PD neurons respond to RPCH early in development, then the entire embryonic rhythm might be modulated by RPCH. The acquisition of CabTRP later in development could be timed to occur when separate activation of the functional antagonists in the pyloric rhythm is needed as the stomach matures.
Why cotransmission?
Why do synapses and neurons express more than one transmitter? Many modulatory and command neurons project to a large number of target neurons within a circuit, by which means an entire movement or behavior may be activated. Modulatory neurons can activate an entire circuit simultaneously by using distinct transmitters on each of those circuit elements. In case of cotransmission, the network elements could be differentially modulated in a number of different ways by a single projection neuron: (1) the target neurons could vary in their affinity for the various transmitters; and (2) the different transmitters in the projection neuron could be preferentially released at different firing rates. The modulatory neuron would then be able to bias the network into different configurations by altering its firing rate, regulating the synthesis, storage, or release of each of its cotransmitters separately, yet the projection neuron would not lose the ability to coordinately regulate different neural network elements, as could occur if each neurotransmitter were found in independent presynaptic neurons.
Footnotes
- Received October 29, 2001.
- Revision received December 4, 2001.
- Accepted December 10, 2001.
This research was supported by National Institutes of Health Grant NS17813. We thank Dr. Barbara Beltz for use of the Wellesley College confocal facility and Stefan R. Pulver for help with the double-labeled image of RPCH and CabTRP. We thank Dr. Hugo Arechiga for gifts of the rabbit anti-RPCH antibody.
Correspondence should be addressed to Dr. Eve Marder, Volen Center, MS 013, Brandeis University, 415 South Street, Waltham, MA 02454-9110. E-mail: marder{at}brandeis.edu.
- Copyright © 2002 Society for Neuroscience