

Abstract
The paraventricular hypothalamic nucleus (PVH) is a key site for integrating neuroendocrine, autonomic, and behavioral adjustments to diverse homeostatic challenges, including “physiological” (e.g., infection or hemorrhage) and “emotional” [e.g., restraint (RST) or footshock] stresses. Both types of challenges ultimately converge to activate common response systems represented in PVH, including the hypothalamo–pituitary–adrenal axis and the sympathoadrenal system. Oligonucleotide microarrays (U74A; Affymetrix, Santa Clara, CA) were used to compare and contrast gene expression profiles in the PVH elicited at 1 and 3 hr after acute exposure to representative physiological [intraperitoneal injection of 10 μg lipopolysaccharide (LPS)] and emotional (30 min RST) stressors. In general, the two challenges recruited relatively few genes in common, with the degree of overlap varying across functional classes of genes. The greatest degree of commonality was seen among signaling molecules and neuropeptides, whereas transcription factors upregulated by RST and LPS were largely distinct. Unexpectedly, RST induced a number of immune-related molecules, which were not regulated by LPS. Hybridization histochemical analyses localized a subset of responsive transcripts to the PVH and/or immediately adjoining regions. Immunerelated molecules in particular distributed broadly to vascular and other barrier-associated cell types. These global transcriptional profiles inform the search for early (transcription factors) and late (target genes) mechanisms in the modulation of PVH, and generalized CNS, responses to categorically distinct stressors.
Introduction
It is now common among workers in the field of stress neurobiology to group stressors into two broad categories. These may be termed “emotional” (also known as psychological, processive, or neurogenic) and “physiological” (also referred to as homeostatic or systemic) and are distinguished on the basis of the nature of the sensory input that registers the challenge, the general pattern of activational responses that they induce within the brain, the extent to which they invoke affective responses, and the circuitry that mediates certain adaptive responses to them (Sawchenko et al., 1996, 2000; Watts, 1996; Herman and Cullinan, 1997; Dayas et al., 2001). Examples of emotional stress paradigms include restraint (RST) or footshock, whereas certain immune challenges (cytokine or endotoxin injection) or hemorrhage are widely used physiological insults.
Despite this distinction, both types of challenges ultimately converge to activate common response systems that almost invariably include the hypothalamo–pituitary–adrenal (HPA) axis and the sympathoadrenal system. The paraventricular hypothalamic nucleus (PVH) is a critical structure in the integration of adaptive responses to stress in that it plays prominent roles in governing HPA and sympathoadrenal output and contains ample representations of other hormonal and behavioral functions that may be called into play in a challenge-specific manner (Swanson and Sawchenko, 1983; Sawchenko et al., 1996, 2000).
Representative physiological (systemic cytokine injection) and emotional (electrical footshock) stressors elicit indistinguishable patterns of cellular activation within the PVH (Ericsson et al., 1994; Li and Sawchenko, 1998). Responsive cell types include parvocellular neurosecretory neurons that express corticotropin-releasing factor (CRF) for the initiation of HPA responses, preautonomic neurons that participate in sympathoadrenal control, and magnocellular neurosecretory cells that produce the hormone oxytocin, the functional role of which in the adaptation to either stressor is uncertain. Extrahypothalamic patterns of cellular activation induced by the two models are quite distinct, but they do share one major source of inputs to the PVH in common: catecholamine neurons in the caudal brainstem. Nevertheless, disruption of this projection system interferes with PVH responses elicited by immune challenge but not footshock (Ericsson et al., 1994; Li and Sawchenko, 1998). Molecular factors that may be involved in eliciting common response profiles to insults that are processed by the brain in distinctive manners are not completely understood.
The purpose of the present study was to compare global patterns of gene expression in the PVH after acute exposure to representative physiological and emotional stressors in an effort to obtain an unbiased evaluation of the ways in which the PVH responds to disparate insults. Two time points were examined in an effort to capture early-responding transcription factors and later-responding effector molecules.
Portions of these results have been presented previously in abstract form (Reyes et al., 2002).
Materials and Methods
Animals and challenge procedures. C57BL/6 mice (25–40 gm) were housed in a colony room with a 12 hr light/dark cycle (lights on at 6 A.M.), with ad libitum access to food (rodent chow 8604; Harlan Teklad, Madison, WI) and water. All procedures were approved by the Institutional Animal Care and Use Committee of the Salk Institute. All challenge procedures began at 10 A.M.. Control animals received intraperitoneal saline injections. Lipopolysaccharide (LPS) (Escherichia coli serotype 055:B5; Sigma, St. Louis, MO) was injected intraperitoneally (10 μg/mouse in 100 μl), and animals remained in the home cage until they were killed. For acute RST, mice were placed in 50 ml conical tubes that had multiple (∼12) air holes to allow increased air flow and placed back into their home cages. After 30 min of RST, the mice were released back into the home cage until they were killed. Animals were killed by chloral hydrate overdose and cervical dislocation.
Dissections. After the animals were killed, the brains were rapidly removed and immediately placed in ice-cold RNAlater (Ambion, Austin, TX). Four hours later, brains were dissected to isolate a PVH-enriched region and an arcuate nucleus (ARH)-enriched region. A series of six cuts was made using a razor blade. Viewing the ventral side of the brain, two coronal cuts designed to isolate a hypothalamic block were placed at the apex of the optic chiasm and at the rostral margin of the mammillary bodies. This slab was then placed flat (Fig. 1), and cuts one and two were placed on either side of the optic chiasm. Cut three was placed just above the third ventricle. Finally, this last block was bisected horizontally, with the dorsal half representing the PVH-enriched region and the ventral half representing the ARH-enriched region.

Dissection procedure. A photograph of a coronal brain slice to illustrate the dissection procedure. A series of six cuts were performed using a razor blade. Viewing the ventral surface of the brain, two coronal cuts were made to isolate a hypothalamic block using the apex of the optic chiasm and the rostral margin of the mammillary bodies as landmarks. This slab was then placed flat and the first two cuts were placed on either side of the chiasm. The third cut was placed just dorsal to the third ventricle. Finally, this last block was bisected horizontally with the dorsal half representing the PVH-enriched region (a) and the ventral half comprising the ARH-enriched region (b). Magnification, 7×.
Array protocol. The dissected regions from five animals were pooled and total RNA was extracted using Trizol (Invitrogen, Rockville, MD) followed by a subsequent clean-up step using an RNAeasy kit (Qiagen, Valencia, CA). Microarray analysis was performed using a double amplification protocol (Luo et al., 1999) because starting total RNA amounts (7–15 μg per condition) were not adequate for standard Affymetrix protocols. Briefly, first-stranded and second-stranded cDNA were synthesized according to standard Affymetrix protocols. Then, unlabeled cRNA was generated using the Megascript kit (Ambion). cRNA was purified with an Rneasy column (Qiagen) and used as a template for priming with random primers and a T7-oligo-dT primer in a reverse transcriptase reaction. This resultant cDNA was purified with Qiaquick columns (Qiagen) and used as a template in a second round of cRNA amplification. For hybridization, cRNA was fragmented and exposed to Affymetrix MGU74Av2 chips [contains probes for more than 7000 mouse genes and 5000 expressed sequence tags (ESTs)] as described in the standard protocol outlined in the Gene Chip Expression Analysis Technical Manual (Affymetrix). After sample hybridization, microarrays were washed and scanned with a laser scanner (Agilent, Palo Alto, CA), primary image condensation was performed with the Genechip software version 4.0 (Affymetrix), and expression values for all chips were scaled to a target intensity of 200. Samples were evaluated for quality by comparison of percentage present values as well as 5′ to 3′ ratios of glyceraldehyde-3-phosphate dehydrogenase and actin. Each sample was profiled in duplicate, with cRNA prepared separately from total RNA.
Tissue processing for histology. Animals were deeply anesthetized with choral hydrate (35 mg/kg, i.p.) and perfused via the ascending aorta with ice-cold saline followed by 4% paraformaldehyde in 0.1% borate buffer, pH 9.5. Brains were postfixed for 16 hr and then cryoprotected in 10% sucrose in 0.1 m phosphate buffer. Brains were frozen on dry ice and sectioned using a sliding microtome. Five series of 30-μm-thick frozen sections were collected in cold ethylene glycol-based cryoprotectant and stored at –20°C until histochemical processing.
Generation of probes. The following procedure was used for the generation of probes for in situ hybridization. First-strand cDNA was generated from whole-brain total RNA collected from normal, LPS-challenged, or restrained animals. Using Primer 3 software, sets of nested primers were designed to amplify (using Advantage2 polymerase; Clontech, Palo Alto, CA) a unique 600–1000 bp sequence of the target gene. Once a PCR fragment was amplified, it was cloned into the Topo II (Invitrogen, Carlsbad, CA) vector and sequenced. Before use in in situ hybridization experiments, plasmid DNA was linearized. Plasmids for orexin and preproenkephalin (ppENK) were generously provided by M. Yanagisawa (University of Texas Southwestern Medical Center, Dallas, TX) and S. Sobol (National Institutes of Health, Bethesda, MD), respectively.
Hybridization histochemistry. In situ hybridization was performed using 35S-labeled sense (control) and antisense cRNA probes. Slides were digested with 0.1–10 μg/ml proteinase K for 30 min at 37°C. Probes were labeled to specific activities of 1–3 × 10 9 dpm/μg and applied to the slide at concentrations of ∼10 7 cpm/ml, overnight at 56°C in a solution containing 50% formamide, 0.3 m NaCl, 10 mm Tris, 1 mm EDTA, 0.05% tRNA, 10 mm dithiothreitol, 1× Denhardt's solution, and 10% dextran sulfate, after which they were treated with 20 μg/ml of ribonuclease A for 30 min at 37°C and washed in 15 mm NaCl/1.5 mm sodium citrate at 65–68°C. Slides were then dehydrated and exposed to x-ray films (β-Max; Eastman Kodak, Rochester, NY) for 24 hr. They were coated with Eastman Kodak NTB-2 liquid emulsion and exposed at 4°C for 15–30 d, as determined by the strength of signal on film. Slides were developed with Eastman Kodak D-19 and fixed with Eastman Kodak rapid fixer.
Immunohistochemistry. Primary antisera included a rabbit polyclonal antiserum directed against a synthetic peptide corresponding to the N-terminal portion (amino acids 5–16) of human Fos protein used at 1:5000 (Santa Cruz Biotechnologies, Santa Cruz, CA), a monoclonal anti-neuronal nuclei (NeuN) (Chemicon, Temecula, CA; 1:500), used to label neurons, and a monoclonal anti-mouse CD31 [also known as platelet–endothelial cell adhesion molecule (PECAM)] (1:500) (PharMingen, San Diego, CA), a marker for endothelial cells. Endogenous peroxidase activity was neutralized by treating tissue for 10 min with 0.3% hydrogen peroxide, followed by 8 min in 1% sodium borohydride to reduce free aldehydes. Tissue was incubated with primary antibody at empirically determined concentrations for 24 hr in PBS–2% blocking serum. Localization was performed using a conventional avidin–biotin immunoperoxidase method. For combined immunohistochemistry and hybridization histochemistry experiments, slight modifications to the protocol were required. Immunostaining was performed first with the following adjustments: nonimmune (blocking) sera, potential sources of RNase contamination, were replaced with 2% BSA and 2% heparin sulfate, and nickel enhancement steps were eliminated from the immunostaining protocol because the nickel-based reaction product does not survive the hybridization steps.
Quantification and imaging. Digital images were captured using a Hammamatsu Orca digital CCD camera affixed to a Leica (Nussloch, Germany) DMR-B microscope. Images were quantified using ImageJ (developed at National Institutes of Health and publicly available at http://rsb.info.nih.gov/ij/). For illustrations, digital images were imported into Adobe Photoshop; only brightness and contrast were adjusted as necessary.
Results
Stress-induced Fos expression
To confirm PVH responsiveness to the stress parameters used, series of sections through the forebrain of mice killed 2 hr after exposure to control procedures, intraperitoneal LPS injection, or 30 min RST were prepared for immunohistochemical detection of Fos immunoreactivity. Whereas control mice displayed at most a few scattered cells in PVH, exhibiting weak nuclear labeling, both acute challenges consistently elicited robust activational responses of similar distribution, with those provoked by LPS appearing somewhat more intense (Fig. 2). The lack of crisp topographic organization of major output neuron classes in mouse PVH (Schonemann et al., 1995) made it difficult to identify affected cell types on the basis of Fos induction patterns alone. Comparisons with known distributions of mRNAs encoding these PVH markers indicated a strong correspondence with presumed parvocellular neurosecretory neurons expressing CRF mRNA and less obvious overlap with oxytocin-expressing or vasopressin-expressing magnocellular neurosecretory cells. The presence of scattered labeled cells in the supraoptic nucleus, a pure magnocellular cell group, indicated at least some involvement of one or both of these cell types in each challenge paradigm.

Induced Fos expression in response to LPS injection or restraint. Expression of the immediate early gene product, Fos, in the PVH of control (saline-injected), LPS-challenged (10 μg, i.p.), and acutely restrained animals (30 min). At 2 hr after stress, both treatments led to comparable patterns of Fos induction in PVH, over and above the low basal levels of expression seen in saline-injected controls, with LPS provoking a somewhat stronger response. Magnification, 130×.
Microarray analysis
The dissection used to generate starting material for microarray analysis encompassed the entire PVH, as defined by Swanson and Kuypers (1980), extended laterally to roughly the level of the descending columns of the fornix, and included aspects of the anterior hypothalamic area (ventrally), small portions of the ventral midline thalamic nuclei and medial zona incerta (dorsally), and the dorsomedial nucleus of the hypothalamus (caudally). Pooled total RNA from five mice per condition (controls and two challenges at 1 and 3 hr after stress) was split, and separate amplifications and hybridizations were conducted on duplicate samples. Pearson correlation coefficients using unfiltered average difference values between duplicates were typically >0.98, and percentage of genes present was in the 30 range. Pairwise comparisons (t test) were conducted to determine whether gene expression levels from LPS or RST animals differed from those of saline-injected controls. A list of genes that were significantly upregulated or downregulated was generated using the following criteria: significant difference between the two groups p < 0.05 (t test), and absolute value of fold change >2.5. The number of genes that displayed increased transcription in the PVH was similar between the two stressors, with the number activated by LPS being slightly higher at both time points (Fig. 3). An examination of the identity of the upregulated genes revealed that the majority of transcriptional activity within the PVH was selective for one or the other stressor. At 1 hr after stress, the genes upregulated in response to either LPS or RST shared only ∼17% overlap, and by 3 hr after stress, this fraction was reduced to ∼7%. A similar pattern was observed for genes that were downregulated, although considerably more genes were shared in common at the 1 hr point (∼42%). Tables 1 and 2 (see supplemental data available at www.jneurosci.org) provide lists of genes that were upregulated and downregulated, respectively, by LPS and RST (RIKEN clones and ESTs have been excluded) grouped by function as signaling-related molecules, nucleic acid binding proteins–transcription factors, neuropeptides–neurotransmitters–receptors, ion channels, and immune-related molecules. This clearly illustrates that the number of genes regulated in common by the two stressors varied across functional categories. On one hand, for example, a number of the signaling molecules that were upregulated during the response to either stressor were for the most part similar, particularly at the 1 hr point. In contrast, although both challenges recruited a similar number of immune-related molecules, these comprised entirely nonoverlapping (stressor-specific) sets of genes.

Overlap in the sets of genes regulated by the two stressors. A depiction of the extent of overlap between the genes that met the following criteria: significant change from saline control (p < 0.05) and a fold change of at least 2.5. Numbers of genes that met these criteria are indicated within each box. There was minimal overlap between the sets of genes upregulated in response to either stressor at both time points, with values ranging between 4 and 16% (LPS, white; Shared, gray; RST, black). A similar pattern is observed in the genes that are downregulated in response to either stressor; however, there is substantially greater overlap at the 1 hr time point (24–35%) versus 3 hr (8–9%).
Regulation of immune molecules
In situ hybridization was used to confirm and localize select genes that demonstrated significant upregulation. An average expression level ≥200 was used as a guideline to select candidates that should be detectable by in situ hybridization. Array data had indicated a 54-fold increase in the transcript encoding the chemokine, interferon-induced protein 10 (IP-10; also known as CXCL10), 3 hr after LPS administration. Figure 4 shows the expression pattern of this chemokine. Saline-treated animals exhibited no detectable expression of IP-10 mRNA. However, in response to LPS injection, this transcript was dramatically induced within the PVH and beyond, with the expression of IP-10 mRNA higher within the PVH than in surrounding tissue. Localization of IP-10 mRNA was combined with immunolabeling for neuronal (NeuN) or endothelial cell (CD31) markers to identify the cell type(s) expressing the chemokine. Although scattered NeuNstained cells in the PVH were associated with above-background accumulations of silver grains, IP-10 mRNA expression appeared to be predominantly non-neuronal. The use of the anti-CD31 antiserum suggested extensive association with the vasculature, with expression within either endothelial cells or other vascular-associated cell types, such as perivascular macrophages or pericytes. IP-10 expression was also upregulated in a number of circumventricular organs, including the subfornical organ (SFO) and area postrema (AP), which can be accessed directly by circulating macromolecules (Fig. 4). This expression pattern is consistent with the function of the chemokine of recruiting leukocytes from the circulation into the CNS (Liu et al., 2001). Discrete cells were also apparent throughout the brain parenchyma of LPS-challenged animals.
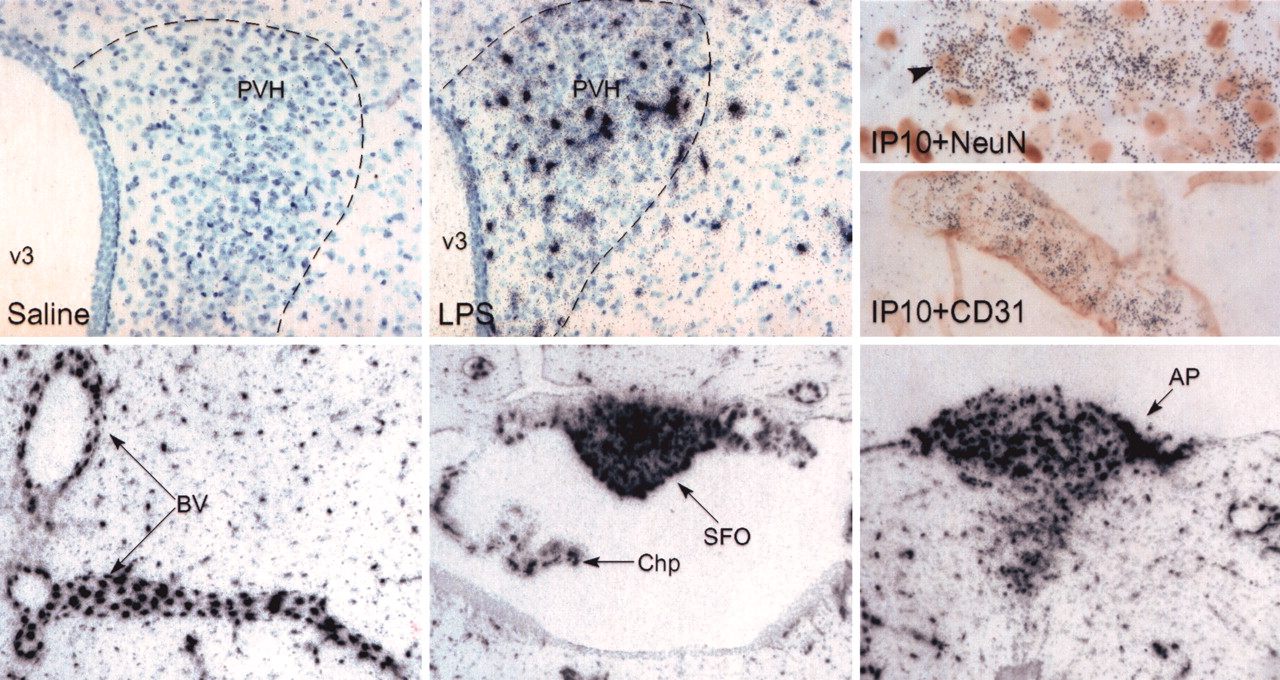
LPS-induced expression of the chemokine IP-10. In situ hybridization was used to confirm the expression of IP-10 in the PVH. Top, Chemokine expression was not detected in saline-treated animals (left) but was rapidly induced in response to LPS (middle; magnification, 70×). Immunolocalization for NeuN to identify neurons (right, top; magnification, 440×) or CD31 to identify blood vessels (BV) (right, bottom; magnification 280×) was combined with in situ hybridization for IP-10 (black grains) in tissue from LPS-treated animals. A NeuN/IP-10 doubly labeled cell (arrowhead) is apparent, but the bulk of IP-10 expression appears to be non-neuronal. Extensive codistribution of CD31 and IP-10 confirms the presence of this transcript on vascular-associated cells. IP-10 was also induced by LPS in other barrier-related areas (bottom), including BV (left), the choroid plexus (Chp) and SFO (middle), and AP (right). Small, discretely labeled cells, possibly glia, are also apparent throughout the brains of LPS-treated animals (magnification, 35×). v3, Third ventricle.
In addition to IP-10, other chemokines demonstrated LPS responsiveness, including macrophage chemotactic protein 1 [MCP-1 (also known as CCL2)] and Gro 1 oncogene (also known as CXCL1) (Fig. 5), with values from the array data showing increases in expression ranging from threefold to fourfold at 1 hr to 10- to 20-fold at 3 hr. In situ hybridization studies revealed MCP-1 labeling around blood vessels, as well as labeling of isolated individual cells, potentially representing neurons or glia. In addition, a pronounced upregulation of MCP-1 transcripts was seen in the choroid plexus, circumventricular organs, blood vessels, and meninges. Gro 1 mRNA exhibited upregulation within the PVH proper, which appeared to be representative of a broader expression associated with blood vessels. Gro 1 expression was also detected in meninges and the choroid plexus but not in circumventricular organs.

LPS-induced expression of additional chemokines, MCP-1 and Gro 1. Other chemokines showed induced patterns of expression that were similar, although not as dramatic as that exhibited by CXCL10, including MCP-1 (top) and Gro 1 (bottom). Dark-field images show expression of mRNA for both chemokines within or immediately adjacent to PVH, as well as in barrier-related areas, including SFO and choroid plexus (MCP-1, top right) and blood vessels (Gro 1, bottom right). Magnification: left, 45×; right, 90×.
The immune-related transcription factor, CCAAT/enhancer binding protein δ (C/EBPδ), showed upregulation in similar barrier-related areas of the CNS (Fig. 6) in a pattern strikingly similar to that exhibited by the chemokines. Whereas control animals exhibited little if any C/EBPδ expression in brain, LPS administration induced a rapid and dramatic induction of this mRNA in PVH, the SFO, choroid plexus, blood vessels, and meninges. The array data indicated that C/EBPδ was in fact more responsive than components of the nuclear factor-κB (NF-κB) system, being upregulated 7.5- and 18-fold, at 1 and 3 hr, respectively, whereas IκBα, an inhibitor of NF-κB, the levels of which provide a dynamic index of NF-κB activity (Baeuerle, 1998), increased 1.4- and 2.6-fold at these intervals.

Expression of the LPS-responsive transcription factor C/EBPδ. Dark-field images illustrating the LPS-induced expression pattern of mRNA encoding the transcription factor C/EBPδ. This shows a distribution in barrier-related structures similar to that of the chemokines (CXCL 10, MCP-1, and Gro1). Little if any expression is apparent in control animals (left). After injection of LPS, there is a dramatic upregulation of this transcript in a number of areas, including the SFO, choroid plexus (Chp), PVH, blood vessels (BVs), and meninges (Men). Magnification, 70×.
A number of additional immune-related molecules were identified in the array analysis as being upregulated in response to LPS. This included previously identified inflammatory mediators known to be LPS responsive, including cyclooxygenase-2 (COX-2), NF-κB, the specific, or α, subunit of the interleukin-6 receptor (IL-6R), and other genes related to activation, including IL-2R β, CD2, CD83 (a dendritic cell maturation marker), and components of the complement cascade (c1q α, c3, and CD59). Other molecules involved in cell adhesion were also upregulated, including vascular cell adhesion molecule 1 (VCAM 1) (Wong et al., 1999), syndecan 4 [a transmembrane heparin sulfate proteoglycan (Kaneider et al., 2002)], and ADAM 8 [a protease implicated in neutrophil migration (Yamamoto et al., 1999)]. Two molecules specifically linked to mast cells were responsive, including mast cell protease 4, which is upregulated late in mast cell development (Serafin et al., 1991), and kit ligand, a mast cell proliferation and chemotactic factor (Galli et al., 1995).
Among the most notable and unexpected findings of the present study were the activation of immune-related molecules in response to RST and the fact that none of these were shared in common with LPS. Array data showed increased expression of a number of adhesion molecules in a pattern similar to the response to LPS, including tumor necrosis factor receptor 4, which is expressed on endothelial cells and can mediate endothelial cell– T-cell adhesion leading to CCL5/RANTES production (Kotani et al., 2002); PECAM (CD31), which is critical for leukocyte migration into the CNS (Wong et al., 1999); CXC chemokine receptor 2, which binds interleukin-8 and Gro 1 and directs neutrophil chemotaxis (Goncalves and Appelberg, 2002); CCR6, which recruits antigen-presenting (Varona et al., 2001) and dendritic cells (Dieu et al., 1998) and serves as the single receptor for MIP3α/CCL2O (Ransohoff and Tani, 1998); and CCL27, a chemokine known to attract T cells to skin (Reiss et al., 2001). The cytokine IL-13 and both subunits of the IL-12R, β 1 and β 2, were upregulated, as well as CD80/B7, an induced costimulatory molecule found on B-cells, dendritic cells, and monocytes (Greenfield et al., 1998).
Neuropeptides
In contrast to the stressor-specific regulation of immune-related molecules, mRNAs encoding a number of neuropeptides and transmitter-related molecules responded in a generally similar manner to acute LPS and RST. Among the more interesting findings in this regard was that RST markedly upregulated orexin/hypocretin mRNA by 11-fold at 3 hr after stress; LPS induced a sixfold increment at this time point. Hybridization histochemistry revealed that although some positively labeled neurons were detected in close proximity to the PVH, none were within it, and expression was centered in the lateral hypothalamic area (LHA). This highlights the fact that the PVH dissection was imprecise and encompassed additional areas (Fig. 7). Quantification of the orexin/hypocretin mRNA signal by densitometry at the single-cell level confirmed a significant upregulation (1.4-fold) in response to RST (p < 0.003 vs controls). No alteration in the number of positively hybridized cells was apparent.

Orexin induction in response to RST. The left panel shows the distribution of orexin mRNA (black grains) within the LHA. The boxed area indicates the approximate region that was quantified. Orexin mRNA is significantly upregulated in response to 30 min RST. Representative images from the brains of control and acutely restrained animals are shown in dark field in the middle and right panels. The upregulation of orexin mRNA is statistically significant (p < 0.003). Magnification, 70×.
Transcripts encoding three other neuropeptides, neuropeptide Y (NPY), enkephalin (ENK), and cholecystokinin (CCK), were very similarly affected by the two acute stressors, with each being downregulated at 1 hr after acute RST or LPS injection. CCK mRNA continued to be downregulated at 3 hr, whereas NPY and ppENK were upregulated, all in response to both stressors. In addition, the fold change levels for each peptide at each time point were also similar. ppENK expression was examined by in situ hybridization (Fig. 8) at 2 hr and was valuable for understanding the site and nature of the upregulation. Whereas increased signal was apparent within the PVH, more robust increments were seen in a laterally adjacent population situated just medial to the fornix.

Neuropeptides that change similarly in response to both stressors. NPY, ppENK, and CCK are similarly affected by acute exposure to systemic LPS or restraint. The bar graphs show the fold change for each neuropeptide at 1 hr (left) and 3 hr (right). In situ hybridization was used to confirm the changes in ppENK mRNA at 2 hr after LPS administration or 30 min RST. Whereas increased signal is apparent within the PVH proper, the upregulation is primarily localized to the region just lateral to the PVH and medial to the fornix. Magnification, 75×.
Several molecules associated with neuronal inhibition also demonstrated similarly altered transcriptional responses in response to either stressor. The GABAA receptor (β 1 subunit) was upregulated at 1 hr in response to both stressors, with RST being somewhat more potent in this regard (2.6 vs 1.6-fold change). Also demonstrating upregulation at 3 hr were two mRNAs related to inhibitory amino acid transmission, glutamic acid decarboxylase 1 (GAD 67), levels of which were increased twofold by LPS and 1.8-fold by RST, and the vesicular inhibitory amino acid transporter [VIAAT (also known as VGAT)], which was increased 2.6-fold in response to both stressors. In addition, somatostatin receptor 4 message was also upregulated at 1 hr in response to both stressors, more so for RST (4.1- vs 2.8-fold change), and this upregulation persisted in the RST condition (elevated 1.6-fold at 3 hr).
Comparison sampling of the arcuate nucleus
In general, profiling results from the arcuate samples were similar to those obtained from PVH dissections (data not shown). For example, of the previously discussed molecules, IP-10, Gro 1, C/EBPδ, VCAM 1, and COX-2 were all upregulated in response to LPS, and in most cases, to a similar degree. Similarly, broadly distributed molecules such as the GABAA receptor (β 1 subunit) were comparably upregulated in both areas by RST. This is in line with the global pattern of expression for these molecules, as seen in hybridization histochemical material. Orexin was also included in the ARH block, and the pattern of upregulation was identical to that seen in the PVH sample (upregulated by both stressors, with the fold change nearly two times greater after RST). However, there were also molecules that were uniquely upregulated in the ARH block, including the chemokine MCP-3, suppressor of cytokine signaling 3, a melanocortin receptor subtype, and IL-1 receptor-associated kinase, all in response to LPS. There were also genes upregulated in response to RST that were unique to the ARH, including prostaglandin D synthase and GABA-A transporter 2. The well known arcuate neuropeptides proopiomelanocortin and NPY were both upregulated (approximately twofold) by both stressors, whereas agouti-related protein was increased slightly in response to LPS alone.
Discussion
Gene expression profiling was used to provide an unbiased global assessment of transcriptional activity within the PVH in response to distinct physiological and emotional stressors. In view of the similarity in the pattern of PVH activation elicited by these challenges, the limited overlap in responsive molecules was unexpected. The fact that the degree of similarity varied with functional class may illuminate differences in the ways in which the hypothalamus, and the brain in general, responds to different stressors. Recruited transcription factors showed little overlap, with only two relatively obscure molecules being responsive to both challenges. By contrast, downstream target genes and signaling molecules showed greater similarity (∼25 and ∼37%, respectively). In particular, a set of neuropeptides exhibited responses comparable in direction, time course, and extent to either challenge.
Methodological considerations
An arbitrary set of criteria was used to identify responsive genes. The 2.5-fold cutoff is relatively conservative because it substantially exceeds the magnitude of stress effects commonly reported for neuropeptide transcripts in the PVH and gives preference for inclusion of factors expressed near the limit of detection under basal conditions. Additional evidence of the conservatism of this criterion lies in the fact that only 12% of all upregulated genes, and 25% of genes that were significantly upregulated, displayed a >2.5-fold change. Among genes known to be responsive in the two challenge paradigms, oxytocin exceeded the criterion (2.7- and 2.8-fold upregulation at 3 hr after LPS and RST, respectively) despite high basal expression, whereas an inducible factor, c-fos, which showed clear induction in immunohistochemical preparations, did not (1.8- and 1.3-fold upregulation). The latter result may be attributable to the coarseness of the dissection and/or tissue inhomogeneity. Other inducible factors known to be responsive and less focally expressed in the LPS model (COX-2, NF-κB, and inducible nitric oxide synthase) all comfortably exceeded the standard. Unfortunately, CRF was not represented on the chip that we used.
The nondiscrete nature of the dissection and the possibility that cells within the circulation might contribute to the prevalence of immune molecules identified in both stress paradigms necessitated using in situ assays to evaluate localizations suggested by the array data. Each factor selected for follow-up did localize to the PVH and/or immediately adjoining regions, although this was often representative of a broader distribution in barrier-related areas. Fold changes in gene expression indicated by the array analysis had to be relatively large (as with IP-10) or localized to a population of substantial size (orexin) to be readily demonstrable. Thus, an 11-fold change (orexin) was readily detected with in situ hybridization, whereas a 2.6-fold change (ppENK) was more difficult to document. An average expression level of 200 (corresponding to ∼4–5 copies per cell) proved a useful guideline for selecting genes to follow up in situ; lower levels may be more easily confirmed by quantitative PCR assays.
In considering below the mass of data provided by the array analysis, we highlight differences in the manner in which various classes of molecules responded to the two stress paradigms, attaching particular importance to instances in which clusters of functionally related genes responded in tandem to a given insult.
Response to LPS
LPS treatment recruited more transcription factors than did RST, including several known to regulate inflammatory mediators such as NF-κB and one not previously implicated in the CNS response to a peripheral immune challenge, C/EBPδ. In the periphery, C/EBPδ is induced by proinflammatory cytokines and activates transcription of many inflammatory mediators, including COX-2, IL-6, MCP-1, and iNOS (Hu et al., 1998; Kim and Fischer, 1998; Kolyada and Madias, 2001). Its distribution in barrier-related structures mimicked that of many of the genes that it regulates, suggesting a functional role for this factor in vivo. These data suggest that along with NF-κB, C/EBPδ is positioned to play a significant role in CNS responses to inflammation and infection.
LPS provoked a dramatic upregulation of chemokine transcripts, with threefold to fivefold changes seen at 1 hr and increases to 20- to 40-fold at 3 hr. More than 50 ligands and 18 receptors (all G-protein-coupled) are currently known to comprise the chemokine family, the members of which display promiscuous interrelationships in which ligands can bind multiple receptors and vice versa (Bajetto et al., 2001). Both IP-10 and MCP-1 recruit primarily lymphocytes and monocytes, with little activity for neutrophils, and in the current study, both chemokines demonstrated increased expression in all barrier-related areas examined. The chemokine Gro 1 demonstrates an opposite recruitment pattern, selective for neutrophils, with little activity for monocytes, and this chemokine showed a more restricted expression pattern, being absent in circumventricular organs. This profound induction of functionally distinct chemokines at barrier-related brain areas suggests that recruitment of peripheral immune cells into the CNS is an important aspect of the brain's response to systemic immune challenge. The relative importance of individual chemokines, as well as the impact of immune cell recruitment into the CNS, will be informed by a better understanding of the details of the timing of expression and cellular localization of each.
RST-induced activation of immune molecules
Perhaps the most unexpected finding of the present study was that RST induced a similar number of immune-related genes, completely distinct from the set that exhibited LPS responsiveness. Bacterial translocation from the gut to peripheral circulation has been reported after RST, but the nature and time course of this phenomenon (Ando et al., 2000) make it unlikely to be involved in acute RST-induced immune activation. The RST-induced transcriptional profile included upregulation of the cytokine, IL-13, which has both proinflammatory (Wills-Karp and Chiaramonte, 2003) and anti-inflammatory (Minty et al., 1993; Di Santo et al., 1997) activities. Interest in IL-13 is augmented by the fact that two molecules active upstream of IL-13 transcription were also upregulated, tumor necrosis factor receptor 8 (TNFR8 or CD30) and TNF receptor-associated factor TRAF-interacting protein (Harlin et al., 2002). Although the role of IL-13 in the CNS response to RST remains obscure, it is of interest that IL-13 can act centrally to potentiate acute phase behavioral effects of systemic LPS (Bluthe et al., 2001). Additionally, other cytokine and chemokine receptors, as well as adhesion molecules, exhibited increased expression in response to RST. This profile suggests that immune cell migration into the CNS is also characteristic of the response to the emotional stressor. Altered leukocyte trafficking in response to emotional stress and the importance of glucocorticoids in that response have been described for peripheral tissues (Dhabhar et al., 1996). Both stressors used here elicit glucocorticoid release and increased expression of signals for leukocyte recruitment, yet they appear to do so using distinct molecular cues.
Neuropeptides
The most substantial overlap in the transcriptional profiles elicited by the two stressors was seen among a group of neuropeptides. Orexin/hypocretin was markedly upregulated in response to RST (11-fold at 3 hr) and to a lesser extent after LPS (5-fold). This peptide system is best known for its activity in arousal and behavioral state (Sutcliffe and de Lecea, 2002), although it is also linked to neural systems controlling feeding–energy balance (Broberger et al., 1998; Elias et al., 1998) and a range of neuroendocrine (Russell et al., 2001) and autonomic responses (Shirasaka et al., 1999) as well. It should be noted that orexin neurons adjoin, but are not intrinsic to, the PVH (de Lecea et al., 1998; Date et al., 1999). Whereas the prominence of their response to both stressors is of interest with respect to potential roles in effecting changes in behavioral state and endocrine/autonomic outflow, whether and how the orexin system may be linked to PVH effector neuron output under either challenge condition remains to be determined.
Transcripts encoding three other peptides, NPY, ENK, and CCK, were all modulated in tandem at both time points in response to each stressor. These neuropeptides are all relatively abundant in CNS, are involved in major behavioral processes such as food intake and energy regulation, anxiety, and pain perception, and have been shown to be regulated by different stressors (Larsen and Mau, 1994; Giardino et al., 1999; Juaneda et al., 2001; Sweerts et al., 2001; Watts and Sanchez-Watts, 2002). Cellular NPY expression has not been localized to the PVH, and the response of this transcript is likely attributable to an adjoining population in the anterior hypothalamic area, which has been shown to exhibit responsiveness to a systemic cytokine challenge (Reyes and Sawchenko, 2002). In contrast, both ENK and CCK are expressed by intrinsic PVH neurons, including parvocellular neurosecretory CRF-expressing cells that govern HPA output (Sawchenko and Swanson, 1985; Mezey et al., 1986; Ceccatelli et al., 1989). Expression of both peptides can be enhanced in this latter cell type by exposure to emotional and/or immune challenges similar to those used here (Van Koughnet et al., 1999; Juaneda et al., 2001), and the capacity of each to serve as corticotropin cosecretagogues, albeit weak ones (Mezey et al., 1986; Ceccatelli et al., 1989), defines potential roles in sculpting the neuroendocrine response in the two distinct stress paradigms.
In terms of informing the goal of identifying factors that might be involved in shaping similar PVH response profiles to disparate challenges, the present analysis identified just a few transcription factors worthy of consideration. In contrast, neuropeptides expressed within (CCK, ENK) and immediately beyond (ENK, NPY, orexin) the PVH were found to respond similarly to the two challenges. With regard to the extrinsic populations, questions remain about the extent to which they may be involved in the PVH response, and if so, whether as cause or consequence. The equally prominent modulation of immune genes by both stressors would suggest that both are perceived by the brain as immune events. In the case of the LPS, the list of responsive factors includes many known mediators, as well as novel ones such as C/EBPδ, that clearly warrant additional attention and is consistent with reports of immune cell migration into the brain under similar challenge conditions (Proescholdt et al., 2002). The unexpected propensity for RST to recruit a comparably sized yet distinct set of chemokines, adhesion molecules, and other immune mediators suggests that such traffic is also characteristic of the CNS response to acute emotional stressors. The relatively slow time course of leukocyte infiltration makes it an unlikely contributor to acute responses (such as HPA activation) in either stress paradigm. Single exposures to immune or emotional stresses are known to be capable of effecting lasting changes in HPA (Johnson et al., 2002a) and other CNS responses (Johnson et al., 2002b) to subsequent insults of various kinds. Whether and how leukocyte infiltration may participate in such phenomenology remains to be evaluated.
Footnotes
This work was supported by National Institutes of Health Grant NS-21182 and was conducted in part by the Foundation for Medical Research. P.E.S. is an investigator of the Foundation for Medical Research. T.M.R. is the recipient of National Research Service Award support (DK-10135). We thank Genelyn Aquino and Mimi Hayakawa for technical expertise, and Kris Trulock for excellent photographic assistance.
Microarray data have been deposited in the Gene Expression Omnibus at www.ncbi.nlm.nih.gov/geo/, series entity number: GSE367.
Correspondence should be addressed to Dr. Paul E. Sawchenko, Laboratory of Neuronal Structure and Function, The Salk Institute, 10010 North Torrey Pines Road, La Jolla, CA 92037. E-mail: sawchenko{at}salk.edu.
Copyright © 2003 Society for Neuroscience 0270-6474/03/235607-10$15.00/0





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}