

Abstract
TRPM8 is a nonselective cation channel activated by cold and the cooling compounds menthol and icilin (Peier et al., 2002). Here, we have used electrophysiology and the calcium-sensitive dye Fura-2 to study the effect of pH and interactions between temperature, pH, and the two chemical agonists menthol and icilin on TRPM8 expressed in Chinese hamster ovary cells. Menthol, icilin, and cold all evoked stimulus-dependent [Ca2+]i responses in standard physiological solutions of pH 7.3. Increasing the extracellular [H+] from pH 7.3 to approximately pH 6 abolished responses to icilin and cold stimulation but did not affect responses to menthol. Icilin concentration-response curves were significantly shifted to the right when pH was lowered from 7.3 to 6.9, whereas those with menthol were unaltered in solutions of pH 6.1. When cells were exposed to solutions in the range of pH 8.1-6.5, the temperature threshold for activation was elevated at higher pH and depressed at lower pH. Superfusing cells with a low subactivating concentration of icilin or menthol elevated the threshold for cold activation at pH 7.4, but cooling failed to evoke [Ca2+]i responses at pH 6 in the presence of either agonist. In voltage-clamp experiments in which the intracellular pH was buffered to different levels, acidification reduced the current amplitude of icilin responses and shifted the threshold for cold activation to lower values with half-maximal inhibition at pH 7.2 and pH 7.6.
The results demonstrate that the activation of TRPM8 by icilin and cold, but not menthol, is modulated by intracellular pH in the physiological range. Furthermore, our data suggest that activation by icilin and cold involve a different mechanism to activation by menthol.
Introduction
The transient receptor potential (TRP) channel TRPM8 is a Ca2+-permeable nonselective cation channel activated by cold and the cooling compounds menthol and icilin (Tsavaler et al., 2001; McKemy et al., 2002; Peier et al., 2002). TRPM8 is expressed in the small DRG neurons and in neurons from trigeminal ganglia (McKemy et al., 2002; Peier et al., 2002). In mouse DRG neurons in vivo, TRPM8 is not colocalized with either TRPV1, IB4 binding, or CGRP (Peier et al., 2002), whereas a proportion of TRPV1-positive DRG neurons in culture express TRPM8 (Reid et al., 2002; Story et al., 2003). This pattern of expression, in small unmyelinated cells distinct from polymodal nociceptors, is consistent with a role in thermosensation and nociception (noxious cold). Furthermore, the activation threshold of TRPM8 is similar to the reported threshold for noxious cold in human tissues (Klement and Arndt, 1992). Although TRPM8 is a prime candidate to detect cold and noxious cold stimulus, not all cold-responsive peripheral neurons express TRPM8 (Nealen et al., 2003; Story et al., 2003). Indeed, a novel TRP channel (ANKTM1 or TRPA1) is expressed in a population of DRG neurons that lack TRPM8 and is activated by cold (Story et al., 2003). Another ion channel that has been suggested to act as a cold receptor is TREK-1, a twin-pore domain potassium channel that is inhibited by cold (Maingret et al., 2000).
Studies on the thermosensitive TRPV1 channel have illustrated the importance of external pH (pHo) on channel function. Changes in the pH in the (patho)physiological range can modulate the ability of capsaicin to activate TRPV1 (Kress et al., 1996; Tominaga et al., 1998; McLatchie and Bevan, 2001). Acidification to pH <6.5 can also activate TRPV1 at room temperature by lowering the thermal threshold for activation (Tominaga et al., 1998).
In this study, we investigated the effects of pH on the activation of TRPM8 induced by two chemical mediators, menthol and icilin, and by cold. Our results demonstrate that pH has a differential effect on TRPM8 activation and that the modulatory effect is determined by intracellular pH (pHi). Acidic pH inhibits responses to icilin and cold, whereas alkaline pH has a potentiating effect. In contrast, menthol responses are not affected by pH, suggesting that activation by icilin and cold involve a different mechanism to activation by menthol.
Materials and Methods
Cell culture. Chinese hamster ovary (CHO) cells expressing mouse TRPM8 (generated as described by Peier et al., 2002) were grown in MEM α medium, without ribonucleosides and deoxyribonucleosides, supplemented with penicillin (100 U/ml), streptomycin (100 μg/ml), l-glutamine (2 mm), FCS (10%), and hygromycin B (200 μg/ml).
Imaging of intracellular calcium levels. To study intracellular calcium ([Ca2+]i) levels in a population of cells, fluorescence was measured using a Flexstation (Molecular Devices, Sunnyvale, CA). Cells were plated in black 96-well plates (Corning, Corning, NY), at an approximate density of 30,000 cells/well and grown overnight. Cells were loaded for ∼1 hr with 2 μm Fura-2 AM (Molecular Probes, Leiden, The Netherlands) in the presence of 2 mm probenecid and 0.01% pluronic F-127 at 37°C. Loading and experiments were performed in an assay buffer containing (in mm) 140 NaCl, 5 KCl, 10 glucose, 10 HEPES (replaced with 5 mm HEPES and 5 mm MES in experiments in which pH was 6.5-6.9, by 10 mm MES below pH 6.5, and by 10 mm N-(1,1-dimethyl-2-hydroxyethyl)-3-amino-2-hydroxypropanesulfonic acid (AMPSO) above pH 7.7), 2 CaCl2, and 1 MgCl2, buffered to pH 7.4 (NaOH) at 24°C. Emission intensity was measured for 90 sec, every 4 or 5 sec using excitation wavelengths of 340 and 380 nm and an emission of 520 nm. Drugs were added after 17 sec. In other experiments, the [Ca2+]i in groups of individual cells was monitored using a microscope-based system as described previously (Savidge et al., 2001). Cells were grown on glass coverslips covered with poly-d-lysine and loaded with Fura-2 as described above and then placed in a laminar flow perfusion chamber. Temperature and pHo were changed by local microperfusion of solution through a fine pipette positioned very close to the cells being studied. The temperature of the perfusion buffer was controlled by a regulated Peltier device (Marlow Industries, Dallas, TX) and monitored by a thermocouple positioned in the tip of the pipette. Images of a group of cells were captured every 2 sec at 340 and 380 nm excitation wavelengths with emission measured at 520 nm with an imaging system (PTI, South Brunswick, NJ). Analyses of emission intensity ratios at 340 nm/380 nm excitation (R, in individual cells) were performed with the ImageMaster suite of software.
Measurements of pHi. Changes in pHi were measured using a Flexstation (Molecular Devices). Cells were prepared in the same way as described above for measuring [Ca2+]i and then loaded for ∼1 hr with 2 μm 2′,7′-bis-(2-carboxyethyl)-5-(and 6)-carboxyfluorescein (BCECF) acetoxymethyl ester in the presence of 2 mm probenecid and 0.01% pluronic F-127 at 37°C. Emission intensity was measured every 5 sec using excitation wavelengths of 440 and 490 nm and an emission of 535 nm. A standard curve of pHi was constructed in the presence of 20 μm nigericin in a buffer prepared as above, except for an equimolar substitution of 110 mm NaCl by KCl. To change pHo, cells were kept in a weakly buffered solution (1 mm HEPES, pH 7.3), and after 22 sec solution (three times the volume present in the well; buffered with 10 mm HEPES or MES) was added to reach the desired pH. The final pH of the solutions was determined in parallel experiments by direct measurement.
Electrophysiology. CHO cells expressing TRPM8 grown on glass coverslips covered with poly-d-lysine were studied by whole-cell voltage-clamp recordings using an Axopatch 200A amplifier filtered at 5 kHz and pClamp 8.0 software (Axon Instruments, Union City, CA). Experiments were performed at a holding potential of -60 mV, unless stated otherwise. Borosilicate glass pipettes (2-5 MΩ) were filled with (in mm) 140 KCl, 0.05 CaCl2, 1 MgATP, 0.1 EGTA, and 10 HEPES (replaced with 5 mm HEPES and 5 mm MES in experiments in which pH was 6.5-6.9 and by 10 mm AMPSO above pH 7.7), pH 7.4 (KOH). Extracellular solutions contained (in mm) 100 NaCl, 80 sucrose, 5 KCl, 10 HEPES (replaced with 5 mm HEPES and 5 mm MES in experiments in which pH was 6.5-6.9, by 10 mm MES below pH 6.5, and by 10 mm AMPSO above pH 7.7), 10 glucose, and 1 CaCl2. The cells were superfused locally through a thin tube positioned very close to the cells. The temperature was controlled as described above for [Ca2+]i measurements. In experiments in which pHi was studied, the cells were left to equilibrate for ∼3 min before recording.
Drugs. (-) Menthol was from Sigma (St. Louis, MO), and icilin was from Tocris Cookson (Bristol, UK).
Calculation and statistics. Data are presented as means ± SEM for the number of experiments indicated (n). Curve fitting was performed in Origin 7 (Originlab, Northampton, MA). Statistical significance was calculated using an unpaired t test or ANOVA, followed by Bonferroni's post hoc test.
Results
We used the calcium-sensitive dye Fura-2 and whole-cell voltage-clamp recordings to investigate the effects of pH on the activity of TRPM8 heterologously expressed in CHO cells. Both icilin and menthol induced concentration-dependent [Ca2+]i responses in Flexstation assays with estimated EC50 values of 125 ± 30 nm for icilin (n = 7) and 101 ± 13 μm for menthol (n = 11), in solutions of physiological pH 7.3 (Fig. 1A,B). To study temperature-induced activation, we used an imaging system that allowed us to monitor the [Ca2+]i levels in individual cells. Exposing cells to a cooling ramp induced [Ca2+]i responses with an activation threshold of 22.6 ± 0.7°C (an average of four determinations of groups of individual cells; n = 18-31) (Fig. 1C). Menthol, icilin, and cold did not produce [Ca2+]i responses in untransfected CHO cells (data not shown). In voltage-clamp experiments, all three stimuli evoked currents (Fig. 1D-F), but responses to icilin developed after a short lag period and desensitized rapidly and completely (1D). In contrast, menthol responses occurred with no obvious lag and showed a more varying degree of desensitization. In some cells, responses to menthol desensitized very little, but in other cells the currents desensitized almost entirely within 5-10 sec (Fig. 1E). Because icilin (but not menthol) requires the presence of extracellular Ca2+ to activate TRPM8 (McKemy et al., 2002), we could not use Ca2+-free solutions to minimize Ca2+-dependent desensitization.

Icilin, menthol, and cold evoke [Ca2+]i responses and currents at pH 7.3. Concentration-response curves for icilin (A) and menthol (B) in TRPM8-expressing CHO cells. Data points represent mean maximal [Ca2+]i response produced by each drug concentration during a 60 sec application, as illustrated in the insets (baseline ratios before the addition of drug has been subtracted; menthol, n = 11; icilin, n = 7). Drug was added after 17 sec. C, Cells exposed to a temperature ramp from 37 to 17°C were activated ∼22°C (average response in 25 cells). The individual responses in the same 25 cells are shown in the inset. Icilin (1 μm) and menthol (1 mm) evoked large inward currents (D,E). Currents in response to both agonists developed rapidly, but for icilin always after a short initial delay (at a holding potential of -60 mV). Icilin-evoked currents desensitized completely, whereas the desensitization of currents evoked by menthol varied. F, Current-temperature relationship in a cell exposed to a temperature ramp (at a holding potential of +60 mV). This particular cell had a threshold close to 24°C and a Q10 of 35 (in the temperature range from 20 to 15°C).
Effects of pHo on [Ca2+]i responses evoked by icilin, menthol, and cold
Changing the extracellular pH to values in the range 5.4-7.4 did not elicit [Ca2+]i responses in the absence of other stimuli (data not shown). We next tested whether acidification of the external solution could affect [Ca2+]i responses induced by a single concentration of agonist (1 μm icilin or 1 mm menthol) or by a temperature stimulus of 17°C (the temperature of the superfusate was ramped from 35°C to 17°C) (Fig. 2). These stimuli were chosen because they could be expected to evoke large responses. In solutions of pHo 6 or below, icilin and cold no longer induced [Ca2+]i responses (Fig. 2A,B). pHo-response curves showed that protons inhibited both cold and icilin with a pIC50 value of pH 6.3 ± 0.1 (Fig. 2C,D). In contrast, [Ca2+]i responses induced by 1mm menthol were unaffected by exposure to solutions with pHo values as low as 5.4 (Fig. 2C). To ensure that an inhibitory action of protons against menthol was not overlooked when using a single high concentration of agonist, we constructed concentration-response curves with menthol and icilin in solutions of different pH (Fig. 3A,B). Acidification of the extracellular solution to pH 6.9 significantly inhibited [Ca2+]i responses to icilin compared with pH 7.3 and shifted the agonist concentration-response curve from a pEC50 value of 7.2 ± 0.1 at pH 7.3 to a pEC50 value of 6.6 ± 0.1 at pH 6.9 (p < 0.001; n = 10). When pH was decreased further, the icilin concentration-response curves were shifted even further to the right, and the maximal responses were depressed (Fig. 3A). As in the experiments using only a single concentration of menthol (Fig. 2C), neither the EC50 values nor the maximal response induced by menthol were significantly affected by acidification of the extracellular solution down to pH 6.1 (p > 0.05; n = 6) (Fig. 3B).

Low pHo inhibits TRPM8 activation by cold and icilin but not menthol.A, Trace from a single cell, demonstrating that icilin is unable to evoke a response at pHo 6.1 but evokes a large [Ca2+]i response when pHo is restored to pH 7.4 (experiment performed at 27°C). B, Cold stimulation (temperature ramp from 35 to 17°C) evokes a [Ca2+]i response at pHo 7.4 but not in solutions of pHo 6.1; data from single cells. C, Low pHo completely inhibits responses induced by 1 μm icilin, whereas responses to 1 mm menthol are not significantly affected. Each data point is the mean response in six wells; similar results were obtained in three experiments. D, Low pHo completely inhibits [Ca2+]i responses induced by cold (17°C). Data points are mean responses to 17°C in groups of cells monitored individually (n = 13-21).

Effect of pH on concentration-response curves for icilin and menthol. A, Acidification of the extracellular solution caused a rightward shift of the responses to icilin, followed by insurmountable inhibition at lower pHo (experiment shown is representative for n = 4). B, [Ca2+]i responses evoked by menthol were not affected by acidification (p > 0.05 for Emax and pEC50; one-way ANOVA; n = 4). C, Effect of menthol and icilin on [Ca2+]i responses induced by cold stimulation. Cells were superfused at 37°C with a solution of pHo 7.3 containing 25 μm menthol, 100 nm icilin (agonist concentrations that did not produce [Ca2+]i responses on their own at this temperature) or vehicle for 1 min before they were stimulated by a temperature ramp down to 15°C. Responses to cold were significantly shifted to higher temperatures in the presence of either agonist (p < 0.001; n = 4 for menthol and n = 5 for icilin, in which each determination was the mean response of n = 21-31 individual cells). When the same experiments were performed in solutions of pHo 6, no [Ca2+]i response developed in the absence or presence of either agonist (n = 4).
The temperature threshold can be modulated by the presence of menthol (McKemy et al., 2002; Peier et al., 2002; Reid et al., 2002) and by icilin (Fig. 3C). Superfusing cells at 37°C for 1 min with a subactivating concentration (at this temperature) of icilin (100 nm) or menthol (25 μm) before starting a cold ramp caused a leftward shift of the temperature-response curve and elevated the threshold for temperature activation from 22.4 ± 0.6°C in the absence of agonist (n = 4) to 26.3 ± 1.4°C (100 nm icilin; n = 5) and 32 ± 0.9°C (25 μm menthol; n = 4). When the same experiments were performed in an external solution of pH 6, no [Ca2+]i responses developed with either agonist, indicating that low pH abolished the synergy between cold and the two agonists (n = 4) (Fig. 3C).
pH affects TRPM8 at an intracellular site
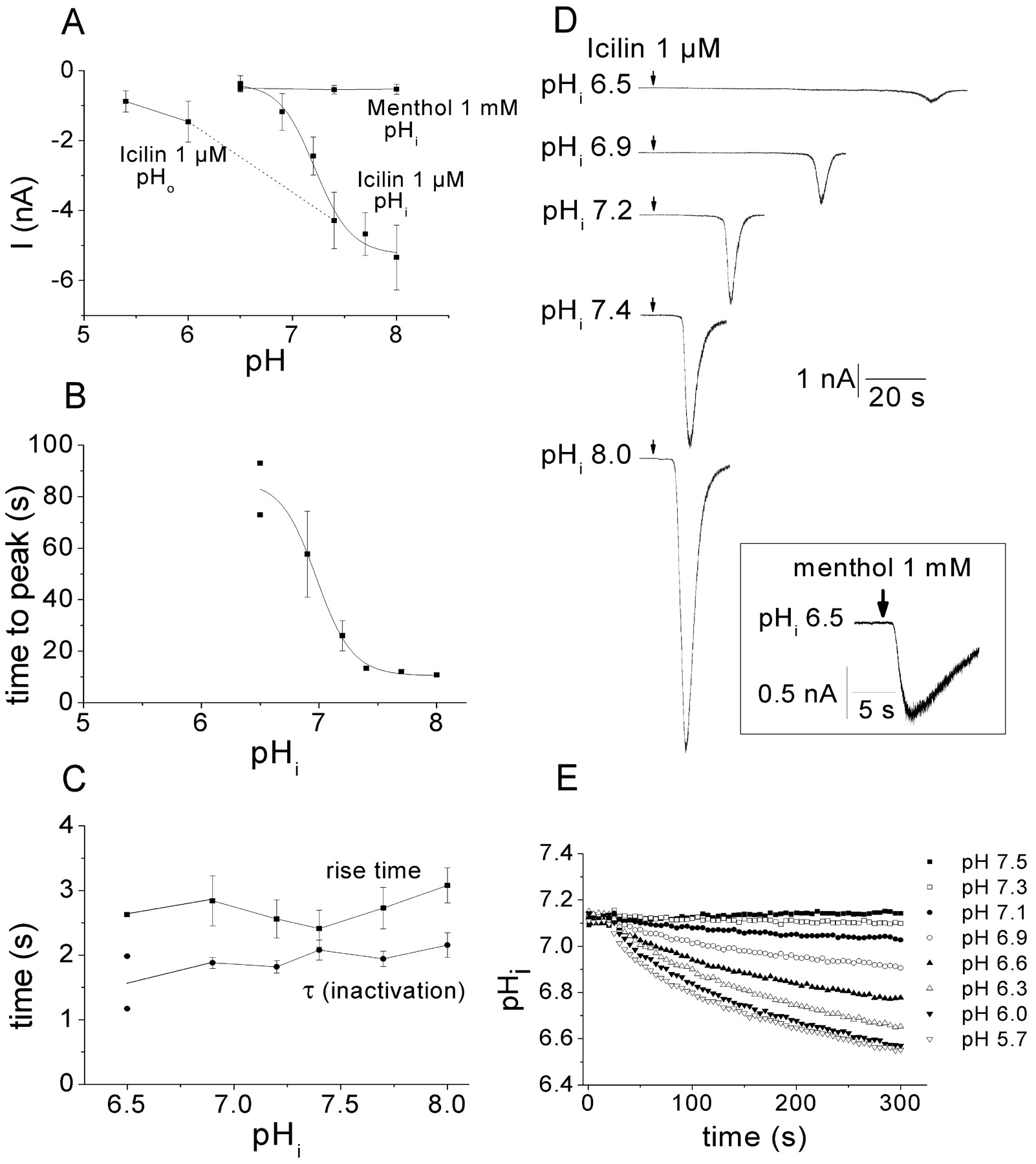
In all the [Ca2+]i measurements described here, the cells studied were equilibrated in test solutions of different pH for 2-5 min before stimulation. To discern whether protons were affecting TRPM8 by acting at a target exposed to the extracellular or intracellular environment, we examined the effect of changing either the external or internal pH on responses to 1 μm icilin in voltage-clamp experiments (Fig. 4). Changing the pH of the extracellular solutions from pH 7.4 to pH 6 or pH 5.4 reduced the response amplitude from -4.3 ± 0.8 nA to -1.5 ± 0.6 nA and -0.9 ± 0.3 nA, respectively (Fig. 4A). When the pHi was altered by using different pH solutions in the recording pipettes, the currents induced by icilin were clearly more sensitive to variations in pHi than extracellular pH (Fig. 4A). Once again, the currents in response to 1 μm icilin were larger at higher pHi values and smaller at lower pHi, with an estimated titration midpoint for current size of pH 7.2, which is a value very near the pHi in physiological situations. The Hill slope for the inhibitory action of protons was 2.5, consistent with an action at several sites with a cooperative inhibitory effect. The short lag phase observed previously (Fig. 1D) for icilin-evoked currents in pHo 7.4 solutions was dependent on pHi (Fig. 4B,D). The latency progressively increased with increasing acidification from ∼10 sec in the normal intracellular solution (pH 7.4) until only two of five cells tested at pHi 6.5 responded within 120 sec (Fig. 4B). Unlike the latency, the time course of the currents did not appear to be affected by pHi (Fig. 4C). The current rise time, the time taken for currents to grow from 10 to 90% of the response, was similar at all pHi values (∼2.5-3 sec; p > 0.05; ANOVA), and the current inactivation, which could be fitted by a single exponential function, occurred with a similar time course (τ close to 2 sec; p > 0.05; ANOVA). Consistent with the previous experiments monitoring [Ca2+]i levels, current responses to 1 mm menthol were not significantly inhibited by pHi 6.5 or potentiated by pHi 8 (Fig. 4A) and also developed without the latency observed with icilin.

pH modulates TRPM8 activation at an intracellular site. A, Although icilin could still evoke inward currents after a 2 min exposure to pHo 5.4, intracellular acidification reduced the current amplitude evoked by 1 μm icilin with an estimated half-maximal inhibition of pH 7.2 (n = 5-9). Inward currents evoked by 1 mm menthol were smaller than currents evoked by 1 μm icilin and not affected by intracellular pH (n = 12-16). B, The latency before icilin-evoked currents developed was increased by intracellular acidification. Data points are averages of cells responding within 120 sec (n = 5-9 cells, except for pHi 6.5, where the only two responding cells are included individually); three of five cells recorded at pHi 6.5 and two of seven cells recorded at pHi 6.9 failed to respond during 120 sec. C, Intracellular acidification did not affect the time course of icilin-evoked currents. The rise time (time from 10 to 90% of current developed) and the τ (inactivation) was similar at all pHi tested. Because only two cells responded at pHi 6.5, the individual data points are shown (n = 5-9 for the other pHi values). D, Representative recordings illustrating how pHi affects icilin-evoked currents. As shown in A and B, the current amplitude is depressed and the latency is increased by intracellular acidification. The box demonstrates that inward currents evoked by menthol were unaffected by pHi 6.5 (compare with Fig. 1 E). E, Extracellular acidification reduced the pHi in a time- and pH-dependent manner. Cells were incubated in pHo 7.3 (weakly buffered with 1 mm HEPES). After 22 sec, the pHo was changed from 7.3 to the indicated values by adding a three times larger volume of strongly buffered solution (10 mm HEPES or MES). Error bars have been omitted for clarity.
In light of the effects of pHi, we were interested in how incubation of cells in extracellular solutions of different pH affected pHi. We examined this using the pH-sensitive dye BCECF in a 96-well assay format (Fig. 4E). The pHi was monitored for 5 min, and the pHo was changed 20 sec after starting the recording. The resting pHi of CHO cells was 7.12 ± 0.01 in an external solution of pH 7.3 at 24°C. Reducing the pH of the external solution below 7.3 evoked a time- and pHo-dependent intracellular acidification such that pHi was reduced to pH 6.8 or below 2-5 min after changing to external solutions of pH 5.7-6.0 (Fig. 4E).
The threshold for cold activation is modulated by pH
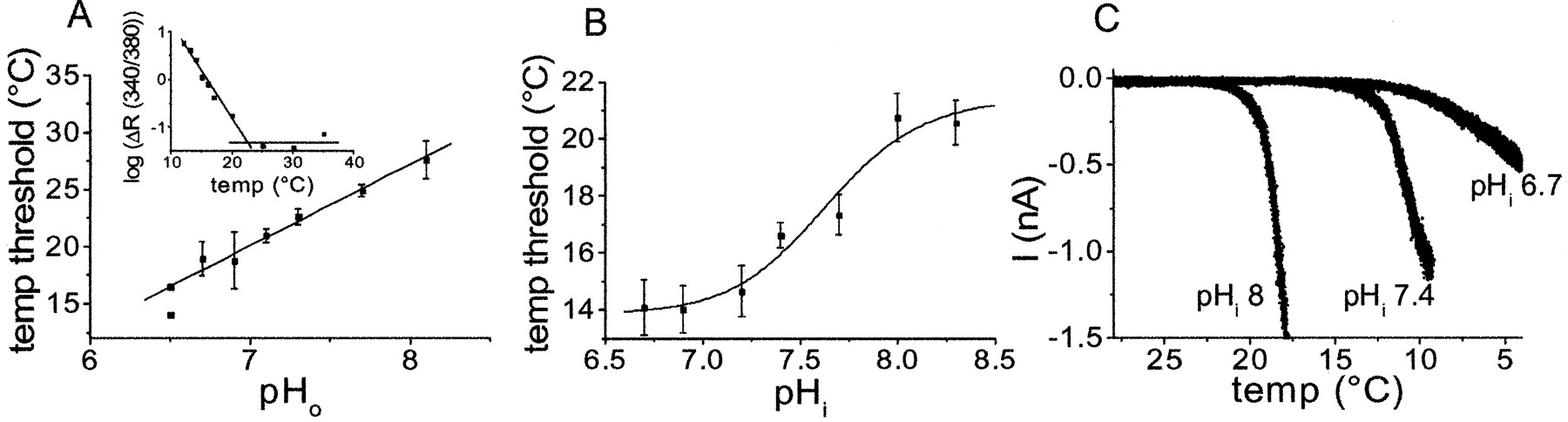
Because responses to cold stimulation (17°C) (Fig. 2D) and the potentiating effect of cooling on responses to low concentrations of menthol and icilin (Fig. 3C) were inhibited by low pH, we investigated whether pH modulated the threshold for temperature activation (Fig. 5). Groups of individual cells were exposed to a temperature ramp (from 37 to ∼12°C) in solutions of different pHo. Thresholds were determined by plotting logarithmic values of the change in emission intensity ratios, log(ΔR), against temperature (Fig. 5A, inset). Two lines were fitted to these plots, one before and one after the cold-evoked [Ca2+]i responses developed, and the intersection of these lines was regarded as the temperature threshold. At physiological pH 7.3, the threshold for cold-induced [Ca2+]i responses determined in this way was 22.6 ± 0.7°C, but in more acidic solution the temperature threshold was shifted to progressively lower values (Fig. 5A). Interestingly, alkaline conditions elevated the activation threshold. In solutions of pH 8.1, [Ca2+]i responses developed with a threshold of 27.6 ± 1.7°C. Alkaline solutions thus potentiated [Ca2+]i responses evoked by cold (Fig. 5A). Given the rather strong difference in sensitivity to pHi and pHo observed with icilin, we next examined the effect of pHi on inward currents induced by cold (Fig. 5B,C). Cells exposed to a cooling ramp from 37 to ∼8°C (holding potential, -60 mV) had an activation threshold of 16.6 ± 0.5°C when the intracellular solution was buffered to pH 7.4 (Fig. 5B). As observed in the [Ca2+]i experiments, the activation threshold was elevated by more alkaline conditions and depressed by intracellular acidification (Fig. 5A). However, unlike the essentially linear relationship between pHo and the cold-evoked [Ca2+]i responses, the pHi current curve determined electrophysiologically had a sigmoidal shape with an estimated half-maximal inhibition at pH 7.6 and a slope factor of 1.9. When pHi was reduced below 6.7, the inhibitory effect was dramatically increased. Only one of seven cells studied at pHi 6.5 responded with a threshold above 8°C. Intracellular pH also appeared to modify the temperature dependence of the currents once they had started to develop, increasing the rate at higher pHi values and slowing the rate at acidic pHi (Fig. 5C).

The activation threshold for temperature is modulated by pH. A, Cells were superfused with solutions of the pH indicated for at least 1 min before they were exposed to a temperature ramp from 37 to 12°C (each point is the average of 2-4 determinations of groups of individual cells; n = 14-31). No [Ca2+]i responses were produced by superfusion alone at 37°C. Thresholds were determined as the intersection between two lines fitted to the log(Δratio) before and after [Ca2+]i responses developed, as illustrated in the inset for an experiment performed at pH 7.1. B, The temperature threshold for cold activation of inward currents (-60 mV) is inhibited by intracellular acidification. The temperature at which a discernable current first developed was set as threshold (each point is the average of n = 5-7). C, Representative recordings demonstrating that the temperature threshold for cold activation is sensitive to the pHi. Intracellular acidification also affects the slope of the temperature dependence.
Discussion
TRPM8 has been characterized as a nonselective cation channel activated by low temperature and the two exogenous ligands menthol and icilin (McKemy et al., 2002; Peier et al., 2002). Here, we demonstrate that TRPM8 activity is also strongly modulated by pH. Low pHo (<6.0) abolished responses to a strong temperature stimuli completely, and varying the pHo from 6.5 to 8.1 shifted the temperature threshold for cold responses from ∼15 to ∼27°C. Icilin and cold responses were inhibited by intracellular acidification and potentiated by intracellular alkalinization. In voltage-clamp recordings in which the pHi can be readily controlled, protons reduced the current size in response to icilin with a half-maximal effect at pHi 7.2, the textbook value for pHi under physiological conditions (Fig. 4A). The threshold for cold activation was shifted to lower temperatures with a half-maximal inhibition at pHi 7.6 (Fig. 5B). This modulating effect of pHi on responses to icilin and cold exhibited cooperativity, underlining the observation that small changes in pHi produce a considerable increase or decrease in the activity of TRPM8. For example, icilin responses were almost fourfold larger at pH 7.4 than at pH 6.9 (Fig. 4A,D). When pHi was reduced below pH 6.5, cold no longer evoked currents in most cells, even in cells that were cooled to 8°C. Similarly, icilin often failed to produce a current during a 2 min application in cells tested at pHi 6.5. Thus, the pH inhibition seems to increase sharply at approximately pHi 6.5. Because it is unlikely that pHi changes will significantly affect even the local pHo around cells subjected to superfusion, the observed pH modulation must be mostly explained by an intracellular action. This is unlike the effect of pH on TRPV1 that has been shown to involve negatively charged residues in or near the pore-forming loop exposed to the extracellular environment (Garcia-Martinez et al., 2000; Jordt et al., 2000). Experiments in which we monitored the pHi using the pH-sensitive dye BCECF illustrate that extracellular acidification decreases pHi. However, this change is time dependent, and the pHi does not equilibrate with pHo in CHO cells within the time scale of our experiments. This finding explains the difference in pH sensitivity between the experiments with Fura-2, in which pHo was altered and pHi was not controlled (pIC50 values of 6.3 for cold and icilin), and the voltage-clamp experiments, in which pHi was buffered directly (pIC50 values of 7.6 and 7.2 for cold and icilin). It is also a probable explanation for the apparently linear rather than sigmoidal dependence of temperature threshold on pHo observed in Figure 5A.
In contrast to our findings with icilin and cold, menthol was entirely resistant to the effects of pHo as well as pHi. Thus, the concentration-response relationship for menthol was unaffected by conditions under which icilin could no longer activate TRPM8 (Fig. 3A,B). In comparison, Behrendt et al. (2004) recently showed that [Ca2+]i responses to single concentrations of menthol and icilin were reduced by incubation in solutions of pH 6.8 and pH 6.3 (although responses to menthol were less sensitive than were those to icilin). Another difference in the responses to the two agonists was that although the responses to menthol developed rapidly without any obvious latency, responses to icilin occurred after a delay. This initial delay or lag phase was strongly increased by low pHi (Fig. 4B,D). With menthol, we never observed a lag phase, even at the lowest pH value tested (pHi 6.5). Curiously, after the lag phase, the response to icilin itself developed rapidly. A striking finding was that the time course of the current responses to icilin were not affected by pHi, although the latency and amplitude were markedly altered. This suggest that the icilin-evoked responses are triggered in an all-or-none-type manner. However, the observation that low pHi extends the time required to reach such a trigger or threshold does not help to clarify why the current size is simultaneously reduced and suggests that these two phenomena may be separate effects of acidification. Together, the pronounced differences in the properties of the responses evoked by menthol and icilin are hard to reconcile with the notion of them acting as agonists through a similar mechanism. Both compounds are stable in the pH range used in this study, so our data cannot be explained by a direct effect of pH on either of the compounds. However, the results would be consistent with icilin responses being dependent on a pH-sensitive intracellular metabolic event or pathway that menthol is not. Because cold responses are also sensitive to pHi,it can further be speculated that differences in this putative mechanism can explain why the threshold for temperature activation observed here in CHO cells was considerably lower than in cultured neurons from trigeminal (27°C) (McKemy et al., 2002) and DRGs (∼28°C) (Reid et al., 2002). The temperature- current relationship for cold-activated currents was very steep. Furthermore, the temperature dependence was reduced by intracellular acidification (Fig. 5C). Analysis of the recording performed at pHi 6.7 (Fig. 5C) produced an apparent Q10 of 40, but much greater values were consistently seen for recordings performed at higher pHi. Such high temperature dependence is hard to reconcile with a direct effect on the channel but would be consistent with an indirect effect acting via a cascade system or another form of amplification.
The temperature threshold for cold activation was strongly elevated by the presence of a subactivating concentration of menthol or icilin (concentrations that did not evoke responses on their own at 37°C). At low pH these synergistic effects on activation were not seen because cooling failed to elicit responses in the presence of either agonist (Fig. 3C). Because menthol responses are pH insensitive (Fig. 3B), this effect must be on cold activation, which fails to operate under acidic conditions even in the presence of menthol. The threshold for cold activation was considerably lower in the voltage-clamp experiments than in the [Ca2+]i measurements (Fig. 5A,B). Although the reason for this is not known to us, it seems likely that this lower sensitivity to cold can be explained by the assay conditions in voltage-clamp measurements, such as the membrane potential or dialysis of the cellular contents by the pipette solution.
Alkalosis (metabolic alkalosis and hypocapnic alkalosis) are common clinical manifestations of many different etiologies (Galla, 2000; Laffey and Kavanagh, 2002). It has long been known that alkalosis increases the excitability of central and DRG neurons (that may lead to cramps and seizures among other symptoms) and increases the activity of voltage-operated calcium channels (Somjen and Tombaugh, 1998; Tombaugh and Somjen, 1998). Intracellular alkalinization strongly increases the affinity of Ins(1,4,5,)P3 for Ins(1,4,5,)P3 receptors (Worley et al., 1987) and increases the release of Ca2+ through Ins(1,4,5,)P3 receptors as well as ryanodine receptors (Joseph et al., 1989; Heppner et al., 2002). Recently, experiments with vascular smooth muscle cells have shown that raising pHi activates a nonselective calcium-permeable cation channel with properties resembling those of many TRP channels, but of unknown identity (Poteser et al., 2003). Thus, different pathways that can activate or potentiate TRP or TRP-like channels and lead to increased [Ca2+]i share a common potentiation by elevated pHi.
We found that not only menthol, but also icilin and intracellular alkalinization, elevate the temperature threshold for cold activation of TRPM8. TRPV1 is often referred to as a polymodal molecular sensor because its activity is regulated by stimuli of different modalities. Heat, low pH, and chemical ligands can all gate TRPV1 and also potentiate the effect of each other (Tominaga et al., 1998). By similar criteria, TRPM8 would qualify as a polymodal molecular sensor, sensing temperature, pH, and “cooling compounds.”
Although it remains to be shown whether the pH effect observed here affects TRPM8 in vivo, the pIC50 values of 7.2 and 7.6 against icilin and cold are close to the pHi under physiological conditions, indicating that small changes in pHi can affect cold perception by regulating the excitability of TRPM8-expressing cold fibers. The inhibitory effect of protons applied intracellularly showed a steep concentration dependence, consistent with an action at more than one site. As yet, it is unclear to us whether these sites are intrinsic to the channel protein or not, but the fact that menthol is resistant to the modulating effects of pH implies that TRPM8 can be activated by at least two distinct mechanisms.
Footnotes
We thank Ilaria Dragoni and Peter McIntyre for the TRPM8 cell line and for valuable discussions.
Correspondence should be addressed to Dr. David Andersson, Novartis Institute for Medical Sciences, 5 Gower Place, London WC1E 6BS, UK. E-mail: David.Andersson{at}pharma.novartis.com.
Copyright © 2004 Society for Neuroscience 0270-6474/04/245364-06$15.00/0





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}