

Abstract
Visual, vestibular, and auditory neurons rely on ribbon synapses for rapid continuous release and recycling of synaptic vesicles. Molecular mechanisms responsible for the properties of ribbon synapses are mostly unknown. The zebrafish vision mutant nrc has unanchored ribbons and abnormal synaptic transmission at cone photoreceptor synapses. We used positional cloning to identify the nrc mutation as a premature stop codon in the synaptojanin1 (synj1) gene. Synaptojanin 1 (Synj1) is undetectable in nrc extracts, and biochemical activities associated with it are reduced. Furthermore, morpholinos directed against synj1 phenocopy the nrc mutation. Synj1 is a polyphosphoinositide phosphatase important at conventional synapses for clathrin-mediated endocytosis and actin cytoskeletal rearrangement. In the nrc cone photoreceptor pedicle, not only are ribbons unanchored, but synaptic vesicles are reduced in number, abnormally distributed, and interspersed within a dense cytoskeletal matrix. Our findings reveal a new role for Synj1 and link phosphoinositide metabolism to ribbon architecture and function at the cone photoreceptor synapse.
Introduction
Sensory cells of the auditory, vestibular, and visual systems rely on ribbon synapses to transmit sensory information to afferent neurons. The ribbon, the most conspicuous structural specialization of these synapses, appears to be a platform that tethers and coordinates exocytosis of synaptic vesicles. Compared with conventional synaptic terminals, ribbon synaptic terminals can release vesicles continuously at exceptionally rapid rates, up to several thousand per second, and have an inordinately large pool of vesicles (Parsons et al., 1994; Rieke and Schwartz, 1996; von Gersdorff et al., 1996). With such high rates of continuous exocytosis, ribbon synapses also have mechanisms for efficient endocytosis and recycling of vesicles.
Recent studies have suggested that ribbon synapses rely on special adaptations of endocytic mechanisms (Lenzi et al., 2002; Holt et al., 2003, 2004; Paillart et al., 2003; Rea et al., 2004). Furthermore, the predominant mechanisms of endocytic membrane retrieval may vary at different types of ribbon synapses. For example, synaptic terminals of retinal bipolar cells take up large plasma membrane fragments by bulk endocytosis to generate endosome-like structures (Lenzi et al., 2002; Paillart et al., 2003). In contrast, membrane recycling appears mediated primarily by small vesicles at cone pedicles under steady-state conditions (Rea at al., 2004).
Ribbons are unanchored, and synaptic vesicles aggregate in the cone photoreceptor terminals of the zebrafish vision mutant nrc (Allwardt et al., 2001). The arciform density, which is thought to anchor ribbons to the presynaptic membrane (Ladman, 1958), is absent in the majority of nrc photoreceptor terminals (Allwardt et al., 2001). The nrc synaptic malformities are associated with blindness and defective synaptic transmission between cones and bipolar cells characterized by pronounced asynchronous oscillations in the electroretinogram response (Allwardt et al., 2001; Van Epps et al., 2001). Unlike the photoreceptor synapse, the bipolar and pineal ribbon synapses, as well as conventional synapses, seem morphologically normal in nrc (Allwardt et al., 2001). The nrc mutant therefore presents the opportunity to investigate unique properties of ribbon synapses, as well as differences between specific types of ribbon synapses.
In the present study, we used positional cloning to identify the nrc mutation as a premature stop codon in the gene encoding synj1. Synj1 is a polyphosphoinositide phosphatase, the enzymatic activity of which regulates clathrin-mediated endocytosis and the actin cytoskeleton in conventional synapses (Sakisaka et al., 1997; Cremona et al., 1999; Harris et al., 2000; Kim et al., 2002; Stefan et al., 2002; Schuske et al., 2003; Verstreken et al., 2003). Evaluation of Synj defects in mouse, nematode, lamprey, and fly reveal abnormal vesicle maintenance and cytoskeletal matrix (Cremona et al., 1999; Gad et al., 2000; Harris et al., 2000; Kim et al., 2002; Schuske et al., 2003; Verstreken et al., 2003). PI(4,5)P2 acts as a cofactor both in the nucleation of actin and in the recruitment and assembly of clathrin coats and its accessory factors. The role of SynJ1 in ribbon synapses has not been characterized previously. nrc cone synapses reveal a novel role for Synj1 in synapse architecture. Our study shows that Synj1 is essential for proper cone photoreceptor ribbon synapse structure, function, and vesicle maintenance in the cone pedicle.
Materials and Methods
Zebrafish maintenance and behavioral screening. The nrc mutant was originally identified in a behavioral screen for blind zebrafish (Brockerhoff et al., 1995). Zebrafish were maintained as described previously (Westerfield, 1994) on a 10/14 hr light/dark cycle. nrca14 larvae were isolated and screened for the optokinetic response (OKR) as described previously (Brockerhoff et al., 1995). The nrca14 mutation is recessive and segregates as a simple Mendelian allele. nrca14 have no OKR in white light of any intensity (Allwardt et al., 2001). However, they do spontaneously move their eyes, suggesting the mutation does not affect eye movement. Mutant larvae normally die between 10 and 13 d post fertilization (dpf).
Orientation of larvae was scored by first placing five 5-6 dpf larvae in a Petri dish. The dish was then swirled, and the orientation of larvae was scored after the water stopped moving. This was repeated four times with 5 min between trials.
The startle response was scored by first placing 5-6 dpf larvae in a 12-well tissue culture plate. Two to three wild-type (WT) larvae were placed in one well, and three nrc larvae were placed in a second well. The plate was then tapped from a 1.5 cm height with tweezers ∼7 cm from the larvae. Tapping was repeated after 30 sec. Larvae were videotaped, and the distance traveled during 0.33 sec after the stimulus was calculated using the public domain NIH Image program developed at the National Institutes of Health (available at http://rsb.info.nih.gov/nih-image/). The experiment was repeated with 12 sets of larvae.
Positional/candidate cloning. A zebrafish bacterial artificial chromosome (BAC) library (Incyte Genomics, Fremont, CA) was screened with PCR primers designed to expressed sequence tag (EST) fb79h03, simple sequence length polymorphism (SSLP) Z7504, and synj1 EST fb02f11. The EST fb79h03 was mapped to BAC88i17. The SSLP Z7504 was mapped to BAC14p23. The fb02f11 synj1 EST was mapped to BAC56h15. The 1500 kb on each end of BAC56h15 were sequenced, and recombination analysis was conducted via denaturing HPLC.
The WT synj1 sequence was determined from the following sources. The sequence of bp 789-1500 and 2136-2756 was obtained from the synj1 EST fb02f11. The sequence of bp 705-789, 1500-1524, and 1716-1944 was obtained from reverse transcription (RT)-PCR from zebrafish larvae. The sequence of bp 789-948 was obtained by direct sequencing of BAC56h15. The sequence for bp 210-705, 1524-1716, 1944-2136, and 2294-2429 was obtained from the Sanger sequencing center (http://www.sanger.ac.uk/Projects/D_rerio/). The sequence data of the 5′ untranslated region (UTR) through bp 210 were obtained from RT-PCR from a zebrafish brain cDNA library kindly donated by John Ngai (University of California, Berkeley, CA). Finally, bp 2756 through the 3′ UTR were obtained by screening the plated brain cDNA library. All regions of the gene were sequenced at least twice and most several times from multiple sources.
In situ hybridization. An antisense RNA probe was transcribed from EcoR1 linearized fb02f11 EST plasmid using digoxigenin-labeled UTP and T7 RNA polymerase as described by the manufacturer (DIG RNA labeling kit; Roche Applied Sciences, Indianapolis, IN). A sense RNA probe was transcribed from XhoI linearized fb02f11 EST plasmid using digoxigenin-labeled UTP and T7 RNA polymerase. These probes cover a 1.99 kb region of synj1 corresponding approximately to exons 5-20, which includes part of the SacI domain and part of the 5′-phosphatase domain. A third probe to cone transducin α subunit was used as a control (Brockerhoff et al., 2003). Whole-mount in situ hybridization experiments on 5-6 dpf larvae was done as described previously (Brockerhoff et al., 1997). Embryos were developed 1 hr, 20 min at room temperature with 4-nitro blue tetrazolium chloride and 5-bromo-4-chloro-3 inolyl-phosphate (Boehringer Mannheim, Indianapolis, IN).
Real-time PCR. RNA was isolated from pools of 30-60 WT or homozygous mutant 6 dpf larvae using the RNAwiz reagent (Ambion, Austin, TX). RNA preparations were resuspended in 10 μl of DEPC-treated water. RNA was then DNase treated for 30 min at 37°C. Phenol extraction and RNA precipitation with 100% ethanol (EtOH) was performed. RNA was washed with 75% EtOH, dried, and resuspended in 25 μl of DEPC-treated water.
RNA (1.3 μg) was reverse transcribed with Thermoscript Reverse Transcriptase (Invitrogen, Carlsbad, CA) with synj1-specific primer SynjEx20.R3 (CCTCGCCAAAATTAGCAAAC). Conditions for reactions were as described by manufacturer's manual (Thermoscript RT PCR system; Invitrogen) at 50°C overnight. cDNA was RNase H treated for 30 min at 37°C. cDNA was then amplified with Sybr green PCR master mix (Applied Biosystems, Foster City, CA) with either synj1 or EF1α control primers. synj1 primer pair1 was SynjEx9.F1 (ACTGCTTGGACCGAACAAAC) and SynjEx11.R3 (TCGTGCTTTATCTGCCAGGTC); synj1 primer pair 2 was 5′UTR.F1 (GCAAGACAGGCACAACTCTG) and SynjEx3.R3 (GGACTTTTCCCACTGAGCTG); synj1 primer pair 3 was SynjEX3.F1 (CAGCTCAGTGGGAAAAGTCC) and SynjEx4.R4 (CTCAGAAGCCAGTCGTCACA); synj1 primer pair 4 was SynjEx10.F2 (AAGCCATGGGTTTGACAGAG) and SynjEx11.R4 (GCTTGTCGTGAGAAGCGCTCG). The sequence of EF1α control primers was GTGCTGTGCTGATTGTTGCT and TGTATGCGCTGACTTCCTTG. Fluorescence and melting curves of reaction products were monitored with an Opticon (MJ Research, South San Francisco, CA).
Morpholinos. Fish were given injections of 2.5, 5, 10, or 20 mg/ml morpholino diluted in Danieau buffer [in mm: 58 NaCl, 0.7 KCl, 0.4 MgSO4, 0.6 Ca(NO3)2, and 5.0 HEPES, pH7.6] and 0.05% phenol red as a dye marker. A morpholino is a modified antisense oligonucleotide that specifically blocks translation of a targeted gene (Corey and Abrams, 2001). synj1 1 morpholinos were synj1-MO-A with a sequence ACGATAACCTTTACTGAATGCCATG and synj1-MO-B with a sequence TCGACAACCTATTTCAGACCTGT. These are both translation-blocking morpholinos targeting zebrafish synj1. Control morpholinos were either a standard control from Gene Tools (Philomath, OR) that has no target and no significant biological activity with a sequence CCTCTTACCTCAGTTACAATTTATA, or a sense synj1 morpholino with a 3′ carboxyfluorescein tag with a sequence CATGGCATTCAGTAAAGGTTATCGT. Controls gave identical results. The embryos were injected at the one- to four-cell stage to a total volume equaling approximately one cell at the eight-cell stage. Microinjections were done in accordance with Meng et al. (1999). Embryos were then raised to 5 dpf and evaluated for an OKR. All embryos were compared with uninjected siblings or uninjected and control injected siblings. A “robust OKR” was qualified as consistent stripe tracking in both directions. A “reduced OKR” was qualified as inconsistent and infrequent stripe tracking. “No OKR” larvae displayed no stripe tracking under two trials of three rotations of the drum in both directions.
Microscopy. Dark-adapted (at least 2 hr) or light-adapted embryos were fixed, embedded, and sectioned as described previously (Brockerhoff et al., 1995). For electron microscopy (EM), 60-80 nm sections were analyzed. For light microscopy, 10-15 μm sections were analyzed. Alexa Fluor 488 phalloidin staining was conducted as described by the manufacturer (Molecular Probes, Eugene, OR). Immunofluorescence microscopy with frozen sections of mouse retina was performed as described (De Camilli et al., 1983). Briefly, 4% paraformaldehyde-fixed retina was sectioned (7 μm thick). Sections were incubated with rabbit polyclonal anti-rat Synj1 proline-rich domain sera (1:100) (Haffner et al., 2000), followed by incubation with Oregon green-conjugated anti-rabbit IgG secondary antibody (Molecular Probes).
Phosphate release assay. Total cytosolic protein preparations were obtained by homogenization of 25 or 55 6 dpf zebrafish embryos in the following (in mm): 25 Tris, pH 8.0, 250 sucrose, 500 KCl, 10 MgCl2, 2 EGTA, and 1 DTT containing Complete Mini, EDTA-free Protease Inhibitor mixture (Roche Applied Sciences). Homogenates were ultra-fuged 45 min at 50,000 rpm at 4°C in a TLA 100.2 rotor (Beckman Instruments, Fullerton, CA). The supernatants containing cytosolic proteins were passed over NAP 5 columns (Amersham Biosciences, Piscataway, NJ) and eluted with (in mm) 30 HEPES, pH7.4, 1 EGTA, 1 MgCl2, and 100 KCl. Six independent WT and six independent mutant pools of larvae were assayed in triplicate with and without lipid substrate added. All embryos of mutant pools were screened for an OKR at least twice. Production of free phosphate was assayed as described (Harder et al., 1994) For three WT and three mutant pools, 10 μg of cytosolic protein was incubated with or without 2 μg of synthetic phosphoinositide 4 phosphate [PI(4)P] for 15 min or 2 μg of cytosolic protein was incubated with or without 1 μg of phosphoinositide 4,5 bisphosphate [PI(4,5)P2] for 15 min in the following (in mm): 30 HEPES, pH7.4, 1 EGTA, 1 MgCl2, and 100 KCl at 37°C. Free Pi generated in the assay was measured by malachite green assay using a microplate reader at 620 nm. Data were corrected for background activity by subtracting-PIP absorbance from +PIP absorbance for each reaction. For a second set of three WT and three mutant pools, 10 μg of cytosolic protein was incubated with 1 μg of synthetic PI(4)P for 45 min or 1 μg of cytosolic protein was incubated with 1 μg of PI(4,5)P2 for 20 min. These conditions gave similar results to the first conditions (data not shown).
Endophilin overlay assay. SH3p8-GST encoding full-length rat endophilin 2 fused to glutathione S-transferase (GST) was kindly provided by Ji-fang Zhang (University of Pennsylvania, Philadelphia, PA). The fusion protein was produced in Escherichia coli and purified as described previously (Chen et al., 2003). WT and nrc larvae, 5-6 dpf, were killed by placing a Petri dish containing the larvae on ice for ∼10 min. Brains were removed using 21.5 gauge needles. Five brains were placed in a Microfuge tube, briefly spun to remove excess liquid, and then homogenized for 10 sec on ice in 1 × SDS sample buffer. Samples were boiled and treated with benzonase (Roche Applied Sciences) before loading onto a 7.5% SDS polyacrylamide gel. The gel was transferred for 4 hr at room temperature to immobilon PVDF membranes, blocked overnight in 5% non fat milk in Tris-buffered saline with Triton, and then probed for 4 hr with 1-10 μg/ml SH3p8-GST in blocking solution. Bound endophilin was detected with HRP-conjugated anti-GST antibody and the ECL detection system (Amersham Biosciences).
Anti-Synj1 Western blot. Western blot was conducted as described for the endophilin overlay assay, with the following exceptions. Instead of endophilin, the blot was incubated with rabbit polyclonal anti-rat Synj1 proline-rich domain sera 4 hr at 1:5000 (Haffner et al., 2000) in blocking solution. Primary antibody was detected with alkaline phosphatase-conjugated anti-rabbit secondary antibody at 1:500 (Sigma-Aldrich, St. Louis, MO).
Results
The nrc gene is synj1
The zebrafish mutant nrc was identified in a screen for mutations that affect a vision-dependent behavior, the OKR (Brockerhoff et al., 1995). The abnormal electroretinogram and ultrastructure of the nrc retina (Allwardt et al., 2001; Van Epps et al., 2001) indicate that the product of the nrc gene plays an important role at the cone photoreceptor ribbon synapse. In addition to vision defects, nrc larvae swim abnormally and do not orient themselves properly. WT larvae at 5-6 dpf orient dorsal side up 98 +/- 3% (SD; n = 55) of the time. In contrast, nrc larvae at 5-6 dpf orient dorsal side up only 28 +/- 13% (SD; n = 55) of the time. nrc larvae display a normal startle response [ratio of nrc/WT distance traveled after stimulus, 1.03 +/- 0.06 (SEM; n = 36 nrc and 30 wild type)]. Gross anatomy and FM1-43 staining of the nrc ear and lateral line look grossly normal (data not shown). The balance defect in nrc may, however, be attributable to abnormal ribbon architecture and synaptic transmission at hair cells, but this was not investigated in this study.
To identify the nrc gene, we first mapped it to a 1.3 cM region on linkage group 10 (Van Epps et al., 2001). We then identified closely linked ESTs and Z-markers (simple sequence length polymorphisms) (Shimoda et al., 1999) in the nrc critical region by recombination analysis of 5206 meioses using single-strand conformation polymorphisms (Fig. 1A). The closest linked markers were EST fb79h03 and Z-marker Z7504. This narrowed the critical region to 0.24 cM (∼156 kb) (Postlethwait et al., 1994), and a chromosomal walk using a zebrafish BAC library (Incyte Genomics) was initiated.

The nrc mutation is a stop codon in synj1, and Synj1 is expressed in brain and retina. A, Positional cloning defines the critical genetic region for the nrc mutation. The number between genetic markers represents the number of recombinants identified. B, Schematic of Synj1 domains, SacI (Sac1), 5′-phosphatase (5′Ptase), and proline-rich protein binding domain (pppp). The site of the nrc mutation is shown as a stop on the domain schematic. DNA sequence wave data of the mutation site reveals C to T nonsense mutation in nrc larvae. The full-length protein sequence of zebrafish Synj1 is shown with the mutation site demarcated in red. The border between phosphatase domains is marked by a bold line. The green rectangle demarcates the putative clathrin-binding domain. The putative amphiphysin-binding domain is outlined by a blue rectangle, and the endophilin-binding domain is demarcated by a red rectangle. C, Antisense, but not sense, RNA in situ hybridization reveals synj1 in WT 6 dpf larval brain. The arrowheads point to the brain region. D, Two-week-old mouse retinal section immunohistochemistry with anti-Synj1 proline-rich domain antibody localizes Synj1 to the OPL and IPL. Scale bar, 50 μm. E, Dendrogram shows the amino acid conservation between zebrafish Synj1 and other previously published Synjs. Zebrafish (Zf) Synj1 is more similar to human and rat Synj1 than to human (Hum) and rat Synj2. Dros, Drosophila. The dendrogram was generated using Vector Nti version 9.0 (Informax, Bethesda, MD). The GenBank accession number for zebrafish synaptojanin is AY736013.
During the positional cloning, EST fb02f11, encoding 1.99 kb of the synj1 gene, was placed on the zebrafish radiation hybrid panel (Hukriede et al., 1999) (most recent maps available at http://zfin.org/cgi-bin/webdriver?MIval=aa-refcrosslist.apg) on linkage group 10. synj1 was a good candidate gene for the nrc mutation, based on its high level of expression in nerve terminals (McPherson et al., 1996) and its importance in synaptic function (Cremona et al., 1999; Harris et al., 2000; Luthi et al., 2001; Verstreken et al., 2003). Therefore, the genetic linkage of synj1 and nrc was assessed. Scorable polymorphisms for synj1 were obtained by screening a BAC library with primers from fb02f11. The ends of BAC56h15, containing the synj1 gene, were sequenced, and recombination analysis was done using denaturing HPLC. One end contained two recombinants, and the other end contained zero recombinants of 5206 meioses. The BAC containing synj1 was therefore at the nrc locus and at most 0.04 cM (∼26 kb) (Postlethwait et al., 1994) from the nrc mutation. synj1 therefore became a strong candidate for the nrc gene.
The full-length WT synj1 sequence (Fig. 1B) was obtained from a zebrafish brain cDNA library (kindly provided by J. Ngai), from EST fb02f11, from the Sanger sequencing center, and from BAC56h15 (see Materials and Methods). The cDNA encoding zebrafish synj1 is 4.8 kb long. The protein sequence is 60% identical to human Synj1 and 40% identical to human Synj2 (Fig. 1E, dendrogram). Synj1 contains a SacI phosphoinositide phosphatase domain (81% identity with human), a 5′ phosphoinositide phosphatase domain (79% identity with human), and a proline-rich C-terminal domain (PRD). The catalytic sites of the zebrafish SacI and 5′-phosphatase domains are highly conserved. Amphiphysin (Cestra et al., 1999), endophilin (Cestra et al., 1999; Ringstad et al., 2001), and putative clathrin binding domains (Ha et al., 2003) are also highly conserved (domains highlighted in Fig. 1). Similar to the mouse and human synj1 genes, zebrafish synj1 encodes stop codons at aa 1309 and 1565. This creates a transcript encoding two splice variants of Synj1, 145 and 172 kDa in size.
To screen the nrc synj1 gene for mutations, we sequenced RT-PCR products from pools of nrc larvae. The analysis revealed a mutation, C to T, that converts ARG 499 to a stop codon (Fig. 1B). This premature stop codon is at the most C-terminal region of the SacI domain. The mutation was confirmed by sequencing genomic DNA from 10 individual nrc (AB/WIK strain zebrafish) and 5 WT (three AB strain, two WIK strain zebrafish) larvae. The mutation was found in genomic DNA from 10 of 10 nrc larvae and 0 of 5 WT larvae.
Synj1 expression
To determine whether the premature stop codon induces degradation of the mutant RNA, we performed quantitative real-time PCR analyses. Multiple primers spanning much of the cDNA (see Materials and Methods) revealed no difference between WT and nrc levels of synj1 RNA, suggesting there is no nonsense mediated degradation of nrc RNA (data not shown).
To evaluate localization of WT synj1 in zebrafish larvae, we performed RNA whole-mount in situ hybridization. The synj1 antisense probe, but not the sense control, revealed intense staining in the brain (Fig. 1C), consistent with the localization of Synj throughout the nervous system and in the retina of other organisms (McPherson et al., 1996; Harris et al., 2000; Verstreken et al., 2003). No antibody tested cross-reacted with zebrafish Synj1 in immunohistochemistry (Gad et al., 2000; Haffner et al., 2000). However, in another vertebrate retina, the mouse, Synj1 is concentrated in nerve terminals of both the outer plexiform layer (OPL) and inner plexiform layer (IPL) (Fig. 1D). Only the rat PRD domain antibody (Haffner et al., 2000) cross-reacted with zebrafish Synj1 on a Western blot (see below).
Knock-down of Synj1 produces a nrc-like phenotype
To further confirm that synj1 encodes the nrc gene, we reproduced the nrc phenotype by injecting synj1 morpholinos. A morpholino is a modified oligonucleotide that blocks translation of the targeted gene (Corey and Abrams, 2001). Key aspects of the nrc mutant phenotype are the absence of an OKR, abnormal balance, and the presence of unanchored ribbons in the OPL, but not in the IPL (Allwardt et al., 2001). In light microscopy sections, the nrc OPL appears thin and patchy. We evaluated embryos injected with two different synj1 morpholinos for these phenotypes. WT embryos were injected either with a control morpholino or with synj1 antisense morpholinos A and/or B at the oneto four-cell stage. Embryos were then scored for the OKR at 5 dpf (see Materials and Methods).
Figure 2A shows that injection of synj1 morpholinos diminished OKR responses significantly compared with controls. The percentage of affected embryos increased with morpholino concentration. Larvae with no OKR also displayed nrc-like swimming and balance behavior. synj1-MO-A was more potent than synj1-MO-B, but both had similar OKR effects. The effect of injection of both morpholinos together was additive. The injected larvae showed no gross abnormalities (Fig. 2B). Thus, injections of synj1 morpholinos induce the nrc behavioral phenotype.
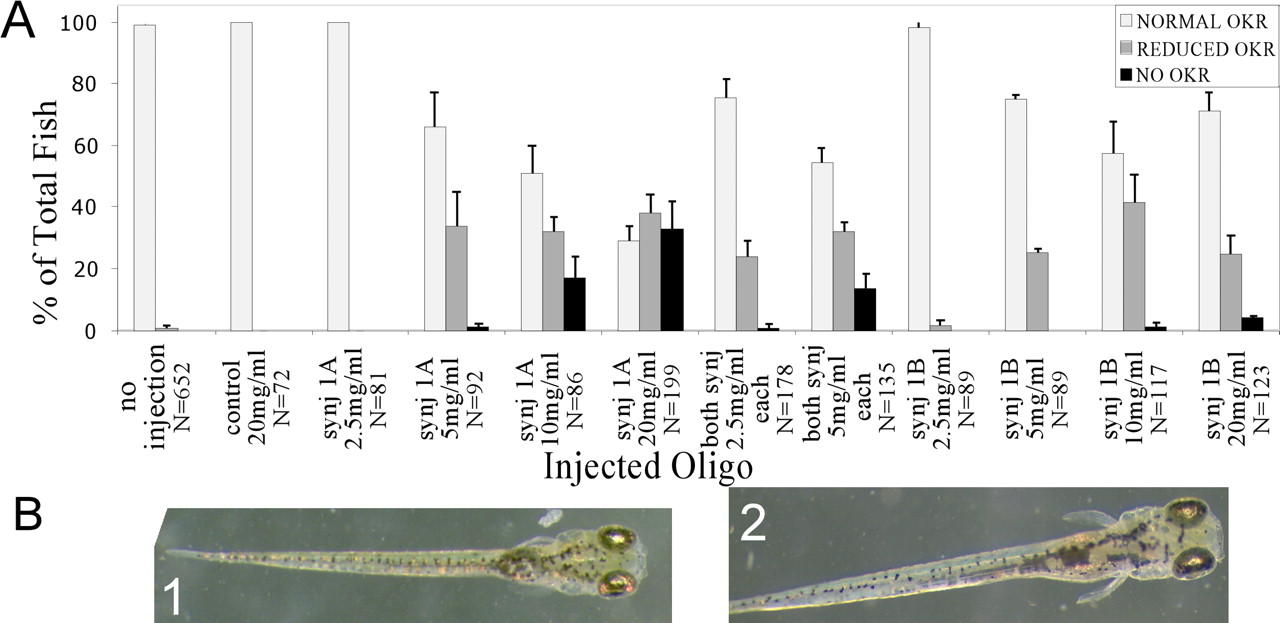
synj1 morpholinos phenocopy the nrc visual behavior defect. A, Percentage of total larvae with no OKR, normal OKR, or reduced OKR plotted for each injection scenario. B, The overall morphology of control (20 mg/ml morpholino, robust OKR) (1) injected and synj1-MO-A (20 mg/ml, no OKR) (2)-injected larvae are grossly indistinguishable. Representative 5 dpf larvae are shown.
To determine whether synj1 morpholino injections cause a morphological defect, we analyzed retinas of OKR-deficient synj1 morpholino-injected larvae by light and electron microscopy. Retinas of larvae injected with synj1 antisense synj1-MO-A had photoreceptors, secondary neurons, and ganglion cells, as did control injected larvae (Fig. 3A). Discs were stacked normally in the photoreceptor outer segments (Fig. 3B). However, the retinas of synj1 morpholino-injected larvae appeared more disordered than in the nrc mutant at a similar stage (Allwardt et al., 2001). The OPL of synj1 morpholino-injected larvae was unusually thin, similar to the nrc mutant (Fig. 3A). Ribbons were predominantly unanchored (85%) in the OPL (Fig. 4) (77 of 92 ribbons unanchored; n = 3 larvae; 64 synapses) and anchored (84%) in the IPL (data not shown; 27 of 32 ribbons anchored; n = 3 larvae; 29 synapses) of synj1 morphant fish similar to nrc larvae (Allwardt et al., 2001). In contrast, retinas of larvae injected with control morpholinos had anchored ribbons in both the OPL and IPL (Fig. 4 and data not shown) similar to WT larvae. Additional changes characteristic of nrc mutant cone pedicles, such as vesicle-free, matrix-rich regions (Fig. 4, bottom, asterisk) (see below), were observed in synj1 morpholino-injected synapses. These results indicate that the synj1 morpholino-injected fish have a retinal morphology phenotype very similar to the nrc mutant and support the conclusion that the nrc mutation inactivates the synj1 gene.

Retina of morpholino-injected larvae. A, Light microscopy of retinas from control (20 mg/ml morpholino, robust OKR) injected and synj1-MO-A (20 mg/ml, no OKR)-injected 5 dpf larvae. The retina is grossly normal. Photoreceptors (PR), secondary neurons (2°), and ganglion cells (GCL) appear properly positioned in the laminated retina. The OPL and IPL are present. However, the OPL in synj1 morpholino-injected larvae appears thinner than in control-injected larvae. B, Electron micrographs of photoreceptor outer segments from control (20 mg/ml morpholino, robust OKR)-injected and synj1-MO-A (20 mg/ml, no OKR)-injected 5 dpf larvae. Both have formed outer segments with nicely stacked discs. Scale bar, 1 μm.

synj1 morpholinos phenocopy the nrc ultrastructural retinal defects of the OPL. EM of the OPL of control (20 mg/ml morpholino, robust OKR)-injected and synj1-MO-A (20 mg/ml, noOKR)-injected 5 dpf larvae. Ribbon structures (R) are unanchored in the retina of MO-A larvae but not in controls. Matrix-rich areas (*) devoid of organelles are also seen in MO-A-injected larvae, consistent with nrc findings in this study (Fig. 6). Representative synapses are shown. Scale bar, 1 μm.
Phosphoinositide phosphatase activity is reduced in nrc mutants
For additional confirmation that the nrc mutation disrupts Synj1, we analyzed phosphoinositide phosphatase activity in extracts from whole WT and nrc larvae. There are two phosphoinositide phosphatase domains in Synj: a SacI homology domain, which can dephosphorylate several positions of the inositol ring, and a central inositol 5′-phosphatase domain. The substrates for the SacI domain are PI(4)P, phosphoinositide 3 phosphate, and phosphoinositide 3,5 phosphate. The substrates for the inositol 5′-phosphatase domain are PI(4,5)P2 and phosphoinositide 3,4,5 trisphosphate [PI(3,4,5)P3] (Guo et al., 1999; Nemoto et al., 2001). To examine the activities of both domains, we assayed release of phosphate when cytosol from either WT or nrc larvae was incubated with PI(4)P or PI(4,5)P2.
Release of free phosphate in nrc extracts was reduced by 23% [PI(4)P substrate] and 21% [PI(4,5)P2 substrate] compared with WT extracts (Fig. 5A,B). This indicates that overall both PI(4)P and PI(4,5)P2 phosphatase activities are compromised in the nrc mutant. We hypothesize that the remaining phosphatase activity is attributable to other phosphoinositide phosphatases in the zebrafish whole larvae extracts. It is particularly notable that PI(4)P phosphatase activity is reduced in the nrc mutant because the nonsense mutation found in nrc is at the most C-terminal amino acid of the SacI domain. The finding that PI(4)P phosphatase activity is reduced in the nrc mutant suggests that a truncated but functional SacI domain is not present at significant levels in the mutant. The reduction in PI(4)P and PI(4,5)P2 phosphoinositide phosphatase activity indicates that the nrc mutation in synj1 causes a loss of function of zebrafish Synj1.

Phosphatase function and Synj1 protein in nrc larvae. A, B, Free phosphate release from PI(4)P and PI(4,5)P2 substrates is reduced in extracts from nrc mutants. Free phosphate-generated per mg of cytosolic protein from 6 dpf zebrafish larval extracts is plotted for WT and nrc mutants. A, PI(4)P was used as substrate to test the activity of the SacI phosphoinositide phosphatase domain. B, PI(4,5)P2 was used as substrate to test the activity of the inositol 5′-phosphatase domain. Error bars represent SDs. Standard two-tailed t tests show that the differences between mutant and wild type are statistically significant [PI(4)P, p = 0.03; PI(4,5)P2, p = 0.005]. C, Immunoblotting of 6 dpf zebrafish brain extracts reveals reduced levels of ∼150 kDa Synj1 (S) in nrc brain extracts. Five brains are loaded per lane. The ∼65 and ∼30 kDa proteins do not correspond to expected sizes of isoforms or SacI truncated products of synaptojanin and appear to be nonspecific. D, Endophilin overlay of larval zebrafish brain extracts. The 150 kDa protein (Synj1) that binds to GST-endophilin is not present in nrc 6 dpf larval brains. However, the GST-endophilin binding to the ∼95 kDa protein (predicted size of dynamin) is observed in both WT and nrc brains from 6 dpf larvae. Lanes 1 and 2 are independent brain samples. Five brains are loaded per lane. S, Zebrafish Synj1; D, putative zebrafish dynamin with the expected electrophoretic mobility.
The Synj1 protein is absent in nrc mutants
To determine whether Synj protein was indeed absent in nrc mutants, two independent approaches were used. First, we searched for a Synj1-immunoreactive band by Western blotting with antibodies raised against the proline-rich region of rat Synj1. A 150 kDa protein (the predicted size of zebrafish Synj1) was recognized in WT, but not nrc, extracts (Fig. 5C).
Second, we performed overlay assays with a major Synj-binding partner. Synj normally interacts with other proteins involved in clathrin-mediated endocytosis. One of these, endophilin, helps localize and stabilize Synj (Gad et al., 2000; Schuske et al., 2003; Verstreken et al., 2003). The binding site for the SH3 domain of endophilin is highly conserved in zebrafish Synj1 (Fig. 1). Zebrafish brain extracts were separated by SDS-PAGE, transferred, and probed in an overlay assay either with GST-rat endophilin or with GST-zebrafish transducin as a control. In WT brain extracts, GST-endophilin, but not GST-transducin, binds a ∼150 kDa protein (Fig. 5D) that has the same electrophoretic mobility as the Synjimmunoreactive band (Fig. 5C). The 150 kDa protein was missing from nrc extracts, consistent with the absence of full-length Synj1 in nrc (Fig. 5D). This result demonstrates that the interaction of endophilin with Synj is conserved in zebrafish and further confirms the absence of Synj in nrc mutants. An ∼95 kDa protein, corresponding to the predicted size of dynamin, binds to GST-endophilin (Ringstad et al., 1997) in both nrc and WT larval brain extracts (Fig. 5D). Dynamin is the other major binding partner of endophilin besides Synj (Ringstad et al., 1999). Binding of GST-endophilin to dynamin serves as a positive control for protein loading and efficient endophilin binding in nrc extracts. The endophilin overlay and Synj1 immunoblot together show full-length Synj1 is absent in nrc extracts.
nrc cone photoreceptor ribbon synapses: actin staining and vesicle traffic
Synj1, via its ability to degrade PI(4,5)P2, participates in clathrin-mediated endocytosis and actin cytoskeleton rearrangement in conventional synapses (Sakisaka et al., 1997; Cremona et al., 1999; Gad et al., 2000; Harris et al., 2000; Raucher et al., 2000; Stefan et al., 2002; Verstreken et al., 2003). We analyzed the distribution of polymerized actin in WT and nrc retinas by staining with phalloidin-conjugated to Alexa Fluor 488 (Fig. 6). The distribution of polymerized actin in the nrc OPL (n = 8) appears patchy and irregular in comparison with the WT OPL. There are also regions that appear to have increased polymerized actin within the nrc synapse in comparison with wild type. This is consistent with the positive role of PI(4,5)P2 on actin nucleation (for example, see Takenawa and Itoh, 2001). Increased polymerized actin is consistent with a null Synj1 phenotype.

Actin localization. Fluorescence micrographs showing polymerized actin in the photoreceptor layer of 6 dpf light-adapted nrc retinas, as revealed by staining with phalloidin. The phalloidin staining is more uneven in nrc than in wild type. Punctate regions of phalloidin staining are seen throughout the nrc OPL. No obvious differences in phalloidin staining were seen in the IPL (data not shown). All nrc larvae examined showed similar staining.
We also revisited the previously described ultrastructure of cone photoreceptor ribbon synapses in nrc to further assess changes that could reflect defects in membrane traffic and actin cytoskeleton. EM was performed on cone pedicles of nrc and WT 6 dpf retinas in both light-adapted (Fig. 7A-E) and dark-adapted states (data not shown). At this age of development, cone pedicles, but not rod spherules, have formed completely. Several changes were observed in nrc cone nerve terminals (Fig. 7). We confirmed the presence of a disrupted synaptic architecture (Fig. 7, compare A, B) and numerous unanchored ribbons (Fig. 7B,C), although some anchored ribbons were also present. Free ribbons often aggregated into clusters separated by one row of vesicles (Fig. 7C). In addition, there were ∼50% fewer synaptic vesicles in nrc versus WT cone synapses (Fig. 7E). These vesicles were distributed unevenly in the terminal, with vesicle-rich areas interspersed with regions devoid of vesicles and rich in a dense cytomatrix (Fig. 7B, asterisk). Most likely, this is the actin cytoskeleton. Clathrin coats were difficult to detect, possibly because of the density of the cytomatrix surrounding vesicles. This prevented a morphometric analysis of clathrin-coated vesicles. However, several vesicles with what appeared to be a partial or complete coat were seen (Fig. 7B, insets). Clusters of vesicles away from the active zone and peculiar rows of vesicles, like “beads on a string,” were often observed, as seen at synapses of Drosophila and Caenorhabditis elegans Synj and endophilin mutants (Schuske et al., 2003; Verstreken et al., 2003). We also found a 10 times increase in endosomal area and 5 times increase in endosome membrane perimeter in nrc synapses [area: light-adapted wild type, 0.02 +/-0.006 μm2 (SEM; n = 18); light-adapted nrc, 0.23 +/- 0.03 μm2 (SEM; n = 20); perimeter: light-adapted wild type, 0.51 +/- 0.13 μm (SEM; n = 18), light-adapted nrc, 5.76 +/- 0.74 μm (SEM; n = 20)]. (Similar results were found for dark-adapted retinas.)

Vesicles at nrc photoreceptor synapses vesicles. A, WT cone synapse in the OPL, showing the even distribution of vesicles and typical ribbon (R) organization. The ribbon is anchored to the plasma membrane via the arciform density (A). Secondary neurons properly invaginate and juxtapose against the anchored ribbon. B, nrc photoreceptor synapse, demonstrating that vesicles are fewer and unevenly distributed. Organelle-free areas (asterisk) occupied by a dense cytomatrix are present among vesicle clusters. The ribbon (R) is unanchored, and increased endosomes (E) are present throughout the cytoplasm. Insets, Vesicles surrounded by a coat. nrc photoreceptor synapses showing a cluster of floating ribbons (C) and peculiar rows of vesicles (D), like beads on a string. Scale bars: A, B, 1 μm; C, D, 200 nm. E, Quantification of vesicles in 18 WT and 20 nrc light-adapted cone photoreceptor synapses is shown. The difference in overall vesicle number per synapse section is statistically significant (p = 6 × 10-9). Similarly, the difference in vesicle number per micrometer of plasma membrane is statistically significant (p = 6 × 10-9). Error bars report the SEM. Vesicles numbers were reduced to nearly the same extent in nrc dark-adapted photoreceptor synapses (24 wild type and 25 nrc; data not shown) as in light-adapted synapses. The difference in vesicle number in nrc versus WT dark-adapted synapses is also statistically significant (p = 2 × 10 vesicle number per micrometer of plasma membrane).
Discussion
In this study, we used the zebrafish nrc mutant to show that Synj1, a polyphosphoinositide phosphatase, is required for anchoring of ribbons, for vesicle maintenance, and for synaptic transmission at cone photoreceptor ribbon synapses. We showed that the nrc mutation introduces a stop codon that truncates Synj1 just C terminal to its SacI phosphoinositide phosphatase domain. The zebrafish nrc mutant makes it possible to evaluate the role of Synj1 in the vertebrate retina. Previously, mice deficient in Synj1 have been characterized (Cremona et al., 1999; Di Paolo et al., 2002; Kim et al., 2002). However, the mutant mice die at postnatal day 1, more than 1 week before ribbon synapses form and retinal function can be assessed. The abnormal swimming behavior of the zebrafish nrc mutant larvae suggests that the mutation causes multiple neurological defects, possibly including vestibular or lateral line defects. However, the larvae survive long enough for cone photoreceptor ribbon synapses to form and for synaptic transmission and vision to be evaluated by biochemical, morphological, and electrophysiological methods.
The protein encoded by the nrc gene appears to be the zebrafish ortholog of mammalian Synj1. Zebrafish Synj1 has greater primary sequence similarity to mammalian Synj1 than to Synj2 (Fig. 1), and it cross-reacts with antibodies raised against rat Synj1 that do not recognize rat Synj2 (data not shown). In addition, conservation of critical protein-protein interaction sites between zebrafish Synj1 and mammalian Synj1, as well as the binding of zebrafish Synj1 to rat endophilin, suggests functional conservation. The absence of a full-length Synj1 protein in nrc was demonstrated by immunoblotting and GST-endophilin overlays. Furthermore, the decreased ability of nrc whole larvae cytosol to dephosphorylate not only PI(4,5)P2 but also PI(4)P suggests that a truncated protein comprising the SacI domain alone is not present at a significant level and that the nrc mutation likely causes complete loss of Synj1 protein. This conclusion is further corroborated by the demonstration that a phenotype similar to that produced by the nrc mutation can be induced by morpholino-induced suppression of the expression of Synj1.
The main in vivo substrate of Synj at synapses is PI(4,5) P2, a plasma membrane-enriched phosphoinositide (Cremona et al., 1999). PI(4,5)P2 functions as a precursor of other signaling molecules such as DAG, IP3, and PI(3,4,5)P3. In addition, PI(4,5)P2 acts as a cofactor both in the nucleation of actin (Sakisaka et al., 1997; Cremona et al., 1999; Takenawa et al., 1999; Gad et al., 2000; Raucher et al., 2000; Takenawa and Itoh, 2001; Kim et al., 2002; Verstreken et al., 2003) and in the recruitment and assembly of clathrin coats and accessory factors (Cremona et al., 1999; Gad et al., 2000; Harris et al., 2000; Kim et al., 2002; Schuske et al., 2003). Abnormalities that occur in Synj-deficient synapses most likely reflect processes that are affected by impaired PI(4,5)P2 hydrolysis leading to enhanced actin polymerization at endocytic/periactive zones and defects in shedding of clathrin coat components after endocytosis.
The predominant characteristics of Synj deficiency vary depending on the organism, the synapse, and the method (genetic disruption or antibody injection) used to disrupt Synj function. However, several structural characteristics of nrc mutant cone photoreceptor synapses resemble abnormalities that have been observed previously at conventional synapses of Synj-deficient nerve terminals in other organisms. These include peculiar rows of vesicles, like beads on a string (Harris et al., 2000; Schuske et al., 2003; Verstreken et al., 2003), the presence of a more pronounced cytoskeletal matrix than at normal nerve terminals (Cremona et al., 1999; Gad et al., 2000; Kim et al., 2002; Verstreken et al., 2003), and clusters of vesicles that often seem trapped in this matrix, isolated away from zones of secretion (Cremona et al., 1999; Gad et al., 2000; Allwardt et al., 2001; Kim et al., 2002; Verstreken et al., 2003). In other organisms, the membranes of these clustered vesicles are either more dense than the membranes of normal synaptic vesicles (Verstreken et al., 2003) or they are clathrin coated (Cremona et al., 1999; Gad et al., 2000; Harris et al., 2000; Kim et al., 2002; Schuske et al., 2003). The high density of the matrix in zebrafish synapses did not allow a clear visualization of clathrin coats, so a reliable morphometric analysis of coated structures could not be performed.
An additional defect shown by the present study is a decrease in the number of synaptic vesicles and an increase in endosomes within cone photoreceptor pedicles. This may reflect a disruption of the normal balance between exocytosis and endocytosis. Surprisingly, there were no obvious differences in the number of vesicles in cone synaptic terminals in dark- versus light-adapted retinas (data not shown). Photoreceptors continuously release neurotransmitter in the dark and decrease continuous release in the light. The similar reduction in the number of vesicles under both conditions suggests effects of Synj1 deficiency on exocytosis and endocytosis that are similar at both low and high levels of activity.
Our study shows that ribbons in cone pedicles are unanchored at 6 dpf in the absence of Synj. Rod spherules are not fully formed at this stage of development and could not be evaluated. In normal 6 dpf retinas, 71% of cone photoreceptor ribbons are anchored, whereas in nrc retinas only 14% are anchored (Allwardt et al., 2001). Several mechanisms could contribute to this cone defect. As described in detail below, Synj1 may directly influence ribbon anchoring via phosphoinositide metabolism, or the role of Synj1 in endocytosis and actin rearrangement may indirectly influence ribbon anchoring.
The nrc photoreceptor synapse may be arrested or slowed in development (Allwardt et al., 2001). In teleost photoreceptor synapses, ribbons are first formed in the cytoplasm, then secondary neurons invaginate, and finally ribbons anchor to the plasma membrane. The development of the nrc terminal may arrest before full invagination of secondary neurons and anchoring of ribbons can occur (Allwardt et al., 2001).
Critical rearrangements of the cytoskeleton may be compromised. Synj functions as a negative regulator of actin nucleation by dephosphorylating PI(4,5)P2 (Singer-Kruger et al., 1998; Cremona et al., 1999; Takenawa et al., 1999; Gad et al., 2000; Harris et al., 2000; Stefan et al., 2002). Excess actin nucleation caused by the absence of Synj may cause ribbons to become abnormally tethered in the cytoplasm rather than at the plasma membrane. Excess PI(4,5)P2 may alternatively cause misregulation of factors essential for the proper anchoring of the ribbon. Notably, phosphoinositides have been shown to influence ribbon anchoring at cone, but not rod, photoreceptors of carp retinas (Schmitz and Drenckhahn, 1993). Recent studies have demonstrated the presence of focal adhesion molecules at the ribbon synapse (Balkema et al., 2003). These proteins are regulated by PI(4,5)P2 (Di Paolo et al., 2002; Ling et al., 2002; Calderwood and Ginsberg, 2003) and may be important in ribbon anchoring.
Synaptic function can influence ribbon anchoring. Precedence for this comes from other organisms. In mice lacking the β2 subunit of the voltage-dependent L-type Ca+2 channel, photoreceptors can no longer signal properly, and ribbons become unanchored (Ball et al., 2002). When squirrels hibernate, ribbons detach from the membrane and then reattach after hibernation ends (Reme and Young, 1977). Finally, if an afferent nerve is cut, ribbons become unanchored. This has been observed in the ampulla of Lorenzini of the electrosensory organ of the fish Torpedo marmorata (Derbin, 1970).
Other synapses including bipolar cell ribbon synapses appear normal in nrc mutants. This is surprising because Synj1 is widely expressed in neurons. RNA whole-mount in situ hybridization revealed synj1 mRNA throughout the larval zebrafish brain. Furthermore, the nrc mutant displays abnormal balance and swimming, suggesting CNS neurological defects beyond blindness. It is unclear why cone photoreceptor pedicles are the most severely affected nerve terminals and why unanchored ribbons in other ribbon synapses are not increased in number. Unanchored ribbons occur in retinal photoreceptors in mice deficient in laminin β2, the β2 subunit of the L-type Ca+2 channel, and bassoon (Libby et al., 1999; Ball et al., 2002; Dick et al., 2003). However, a significant difference between these proteins and Synj1 is that laminin β2 and bassoon are not present in bipolar ribbon synapses in the IPL. In contrast, Synj1 is likely expressed in both photoreceptor and bipolar synapses.
It is possible that a second synj gene, not yet identified in the currently incomplete zebrafish genome database, may compensate for the loss of Synj1 at other synapses but not in photoreceptors. Alternatively, photoreceptor cells may have unique properties that make them inherently more susceptible to loss of Synj1 activity. This could include photoreceptor-specific regulation of other enzymes that control PI(4,5)P2 metabolism or differences in the predominant pathways of synaptic vesicle recycling. For example, a recent study suggested that cone photoreceptors rely entirely on endocytosis of small vesicles (Rea et al., 2004), whereas endocytosis in bipolar cells is qualitatively different (Holt et al., 2003, 2004; Paillart et al., 2003). Interestingly, we found increased endosomes in nrc mutants. Furthermore, photoreceptors are more sensitive to defects in Ca2+ homeostasis, and the delay before the onset of endocytosis after the initial rise in Ca2+ is unusually short in photoreceptors (Rieke and Schwartz, 1996). Because the PI(4,5)P2 pools controlled by Synj are likely to be involved, at least indirectly, in Ca2+ signaling, Synj deficiency may have a more disruptive effect in photoreceptors because of a special mechanism of Ca2+ homeostasis in these cells. We note that mutations in synj were isolated in forward genetic screens for blindness both in Drosophila and in zebrafish. Phosphoinositide metabolism is critically implicated in the invertebrate phototransduction cascade but not in vertebrates (Yarfitz and Hurley, 1994). Studies of Drosophila (Verstreken et al., 2003) and zebrafish (Van Epps et al., 2001) photoreceptors have demonstrated a selective effect of Synj mutations on synaptic transmission and not on the phototransduction cascade. However, it is possible that important signaling functions of PI(4,5)P2 at photoreceptor synapses have been conserved in evolution.
In conclusion, our analysis of the zebrafish nrc mutant has demonstrated a special dependence of the cone photoreceptor ribbon synapse on the function of Synj. The nrc mutant may now be used to further elucidate the general role of Synj in synaptic transmission and its special function at photoreceptors. In addition, it may help to reveal new aspects of the still elusive cell biology of ribbons.
Footnotes
This work was supported by the Howard Hughes Medical Institute; National Institutes of Health Grants NS36251 (P.D.), EY012373 (S.E.B.), and EY06641 (J.B.H.); a Human Frontiers long-term fellowship to M.H.; and Molecular and Cellular Biology Training Grant 5T326M07270 to H.A.V. We thank John Ngai (University of California, Berkeley, CA) for kindly sending the zebrafish brain cDNA library and Ji-fang Zhang (University of Pennsylvania, Philadelphia, PA) for kindly sending us the SH3p8-GST clone. We thank Irina Ankoudinova and Greg Niemi for technical assistance during the course of this study. We also thank John Saari and Visvanathan Ramamurthy for their critical reading of this manuscript.
Correspondence should be addressed to Dr. Susan E. Brockerhoff, Department of Biochemistry, University of Washington School of Medicine, Box 357350, Seattle, WA 98195. E-mail: sbrocker{at}u.washington.edu.
Copyright © 2004 Society for Neuroscience 0270-6474/04/248641-10$15.00/0





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}