
Abstract
During development, neural networks progress through important functional changes such as the generation of spontaneous activity, the expression of a depolarizing chloride gradient, and the appearance of neuromodulation. Little is known about how these processes are integrated to yield mature behaviors. We showed previously that, during the maturation of the locomotor network of the zebrafish, endogenous serotonin (5HT) increased motor activity by reducing intervals of inactivity, without affecting the active swim periods that are the target of 5HT in other and more mature preparations. Because membrane properties were constant during the rest intervals, we examined here whether 5HT modulates chloride homeostasis. We compared the effects of blocking (inward) chloride cotransport with bumetanide to the effects of 5HT and its antagonists, both behaviorally by video imaging and cellularly by whole-cell and gramicidin-perforated patch recordings. Bumetanide mimicked the effects of 5HT antagonists, by prolonging rest intervals without affecting the properties of swim episodes (duration; frequency; extent of depolarization) either behaviorally or during fictive swimming. Furthermore, bumetanide and 5HT antagonists suppressed the amplitude of depolarizing responses evoked by ionophoresis of glycine onto spinal neurons in the presence of tetrodotoxin and transiently suppressed the amplitude of responses to glycine measured after fictive swimming. The effects of bumetanide contrasted with and occluded the effects of 5HT. We suggest that, during development, endogenous 5HT modulates chloride homeostasis during the quiescent intervals and thereby offsets the long periods of quiescence commonly observed in developing networks to allow expression of sustained and behaviorally relevant activity.
- neuromodulation
- locomotion
- maturation
- neural network
- chloride homeostasis
- zebrafish
Introduction
Many regions of the developing CNS, including the brain (O'Donovan, 1999; Ben-Ari, 2001; Zhang and Poo, 2001) and the spinal cord (Kudo and Nishimaru, 1998; Nakayama et al., 1999; Branchereau et al., 2000; Hanson and Landmesser, 2003; Ren and Greer, 2003; Yvert et al., 2004), exhibit a characteristic recurrence of spontaneous neural network activity. During that period, GABA and glycine can exert excitatory effects (Cherubini et al., 1991; Ben-Ari, 2001; Payne et al., 2003; Rivera et al., 2005), and this excitation has been implicated as a fundamental developmental process (Zhang and Poo, 2001; Payne et al., 2003; Rivera et al., 2005). GABA and glycine are excitatory in the embryo because of the depolarizing chloride gradient (with a high intracellular chloride level), which is thought to result from a developmental gradient of chloride cotransporter expression with an initial dominance of the inwardly directed chloride cotransporter (Delpire, 2000; Payne et al., 2003; Rivera et al., 2005). Recent evidence from work in the embryonic spinal cord of chick (Chub and O'Donovan, 2001; Marchetti et al., 2005) has suggested that the occurrence of spontaneous episodes (for review, see O'Donovan, 1999) is mainly limited by chloride replenishment, which takes many minutes during the interepisode intervals (Chub and O'Donovan, 2001; Marchetti et al., 2005). Little is known, however, about the mechanisms that modulate chloride homeostasis during development (Delpire, 2000; Payne et al., 2003).
We showed previously that endogenous serotonin (5HT) modulates the activity state of the neuronal network underlying locomotion in the developing zebrafish by modulating the duration of quiescent intervals that interrupt activity periods (Brustein et al., 2003b). Surprisingly, this occurs without affecting the properties of active (swim) episodes, which retain a normal presynaptic drive. This excludes the mechanisms described in other and more mature preparations in which presynaptic modulation and postsynaptic changes in conductance occur (for review, see Schmidt and Jordan, 2000; Brustein et al., 2003b). In contrast to the lack of effects of 5HT on the active (swim) period, 5-HT reduced intervals of quiescence, whereas its antagonists prolonged them considerably (Brustein et al., 2003b), a pattern reminiscent of that described in other highly excitable developing neuronal networks (O'Donovan, 1999; Ben-Ari, 2001; Zhang and Poo, 2001). These effects of 5HT and its antagonists on the duration of the quiescent intervals were not accompanied by apparent changes in membrane properties, excluding ionic conductances as the major targets for 5HT modulation during quiescent intervals (Brustein et al., 2003b). We therefore examine here the novel possibility that 5HT modulates chloride homeostasis during the quiescent intervals. The effects of blocking the (inward) chloride cotransport with bumetanide (Russell, 2000) were compared with the effects of 5HT and its antagonists, behaviorally by video analyses and during fictive swimming by whole-cell and gramicidin-perforated patch recordings. We show that bumetanide mimics the effects of serotonergic antagonists and opposes the effects of 5HT, behaviorally by introducing long quiescent intervals and cellularly by suppressing evoked chloride-mediated responses in isolation or during fictive swimming. We suggest that, early during development, 5HT regulates chloride homeostasis during the rest intervals to offset long periods of quiescence, which are commonly observed in other excitable developing networks. Thus, 5HT modulation allows the locomotor neuronal network to express a more mature, sustained activity pattern.
Materials and Methods
Experiments were performed on zebrafish (Danio rerio) embryos and larvae raised at 28.5°C, according to previously established procedures (Westerfield, 1995) and in compliance with the guidelines of the Canadian Council for Animal Care, McGill University.
Behavioral experiments
The procedures of the behavioral experiments were described in detail previously (Brustein et al., 2003b). Briefly, 3-to 4-d-old larvae from 3-10 different batches were embedded in low-melting-point agarose (1%; Invitrogen, Gaithersburg, MD) with the tail free to move. The swimming patterns of the agarose-embedded animals, which were similar to those of free swimmers (Fuiman and Webb, 1988; Budick and O'Malley, 2000; Buss and Drapeau, 2001; Brustein et al., 2003b), were videotaped at 60 or 120 Hz using CCD cameras (wv-bp510 or Pulnix TM-640; Panasonic, Montreal, Canada) mounted on a dissection microscope. Images and time codes [Society of Motion Picture and Television Engineers (SMPTE) time codes engraved onto each video field using a time code generator] (TG-50; Horita, Mission Viejo, CA) were registered using a videotape recorder (AG-1960; Panasonic).
To study the effects of drugs on the swimming behavior, the solutions were pressure ejected with a Picospritzer (General Valve, Fairfield, NJ) into the pericardial sac of the agarose-embedded larvae using a fine glass pipette. The drugs (50 μm to 1 mm) were dissolved in extracellular Evans solution (see below, Cellular recordings) with the vital dye fast-green (1%; Sigma, St. Louis, MO) to monitor the quality of the injections. Based on the injected volume (∼0.2 μl) and the volume of the larva (∼3 μl), we estimated the effective drug concentration to be ∼15 times less than that of the injected solution.
Cellular recordings
Four-day-old larvae were dissected under anesthesia, 0.2% MS-222 (Sigma) in Evans solution [containing the following (in mm): 134 NaCl, 2.9 KCl, 2.1 CaCl2, 1.2 MgCl2, 10 HEPES, 10 glucose, pH 7.8, 290 mOsm], and two to three somites were exposed to visualize spinal neurons with Hoffman modulation optics (40× water immersion objective) and to record from them in vivo, as described in detail previously (Drapeau et al., 1999). During the experiments, which were performed at room temperature, the anesthetic was replaced by d-tubocurarine (15 μm; Sigma) to paralyze the preparation.
Whole-cell current recordings. To follow the effects of drugs at the cellular level during fictive spontaneous swimming, whole-cell current recordings were obtained from spinal neurons. The patch electrodes (6-8 MΩ) were filled with the following (in mm): 105 d-gluconic acid, 16 KCl, 2 MgCl2, 10 HEPES, 10 EGTA, 4 Na2ATP, adjusted to pH 7.2, 290 mOsm. Sulforhodamine B (2%) was included in the pipette solution for neuronal identification by epifluorescent illumination at the end of the experiment. Five to 10 min after control spontaneous-swimming recordings, 5HT (7-60 μm; Sigma), quipazine (2-10 μm; RBI, Natick, MA), methysergide (2.5-25 μm; RBI), ketanserin (2.5-25 μm; Sigma), or bumetanide (35-100 μm from stock of 2 mg/ml 10% methanol; Sigma) were bath perfused, followed by a period of washout. To verify that bumetanide did not interfere with glycine receptor affinity, we compared the properties of spontaneous miniature postsynaptic glycinergic currents (mPSCs) before and after bumetanide. The glycinergic mPSCs were isolated using d-tubocurarine (15 μm; Sigma), bicuculline (10 μm; Sigma), kynurenic acid (1 mm; Sigma), and tetrodotoxin (TTX) (1 μm; Tocris, Bristol, UK). Recordings were made using an Axopatch 200B amplifier, and data were digitized on-line at 40 kHz using Clampex 8.0 software (Molecular Devices, Union City, CA).
Gramicidin-perforated patch recordings. To test the effects of serotonergic drugs and bumetanide on chloride homeostasis, we ionophoretically evoked glycine responses (see below, Ionophoresis) using gramicidin-perforated (gramicidin D; Sigma) patch recordings to preserve the intracellular chloride concentration. The tip of the pipette was first dipped into a gramicidin-free solution containing the following (in mm): 132 KCl, 2 MgCl2, 10 HEPES, 10 EGTA, and 4 Na2ATP; 290 mOsm, pH 7.2; and then the pipette was backfilled with the same solution, including 10 μg/ml gramicidin (from a 10 mg/ml stock solution in DMSO). In all experiments, sulforhodamine B (2%) was added to the pipette solution to rule out spontaneous rupturing of the patch membrane, as verified at the end of the experiment using standard epifluorescence. Recordings started when the input resistance stabilized (150-350 MΩ). Glycine-evoked responses were isolated using Evans bath solution containing d-tubocurarine, bicuculline, kynurenic acid, and TTX (for doses, see above, Whole-cell current recordings). At the end of some experiments, the glycine-evoked responses were blocked by strychnine (1 μm; Sigma) to confirm their nature. The pharmacological procedure, including drug concentrations, was the same as described above in Whole-cell current recordings. In an additional set of experiments using gramicidin-perforated patches, glycine was ionophoretically delivered during spontaneous swimming. In this case, the experimental protocols and pharmacology were similar to those described for recording of spontaneous fictive swimming (see above, Whole-cell current recordings).
Ionophoresis. Ionophoresis of glycine onto spinal neurons of intact, paralyzed zebrafish larvae was as described previously (Brustein et al., 2003a). Briefly, glycine (1 m; pH 6; Invitrogen) diluted in tissue culture water (Sigma) was ejected locally through a fine glass pipette (20-30 MΩ) using an ionophoretic unit (MVCS-02; NPI, Tamm, Germany) by applying brief 50-100 ms pulses. Positive ejection currents were in the range of 40-70 nA, whereas negative retaining currents ranged between 2 and 5 nA. The ionophoresis parameters, including frequency (0.2-1 Hz), were set to provide steady responses under control conditions and were kept constant for the rest of the experiment.
Data analysis and statistics
Behavioral experiments. Equal time recording sessions were compared before, immediately after, and 30-60 min (data not shown) (Brustein et al., 2003b) after drug injection. The time code (SMPTE) recorded in each video field permitted reconstruction of the distribution of swim episodes in time and the calculation of swim duration and rest intervals using custom-made software.
Cellular recordings. Patterns of spontaneous swimming (whole-cell patch recordings) were analyzed semiautomatically with Clampex and Clampfit 8.0 software and included spinal neurons with constant input resistance (ranging from 150 to 350 MΩ) and resting potentials (approximately -60 mV). The extent of the depolarization during fictive swimming before and after drug was determined by calculating the area under the curve of each swim episode (Clampfit 8.0), and fictive swimming frequency was evaluated by counting the number of large, rhythmic EPSPs during the synaptic drive for each swim episode, as established previously (Buss and Drapeau, 2001). The properties of isolated glycine mPSCs (half-width, rise-time, amplitude, and frequency) were determined using Axograph 4.4. To compare the amplitude of ionophoretically evoked isolated glycine responses before and after drugs (perforated-patch recordings), we used custom-made software after exporting traces acquired with Clampex as text to Matlab. Equal numbers of consecutive events were included in the averages before and after drug, which are presented with confidence intervals of 99% around the mean. To analyze the changes in the amplitude of glycine-evoked responses during spontaneous swimming (perforated-patch recordings), we reconstructed the temporal distribution of the swim episodes and glycine-evoked responses for each experiment, before and after drug. To quantify the changes in the amplitude of the glycine-evoked responses during the rest intervals of fictively swimming larvae, we calculated the ratio between the amplitude of the first and the last glycine-evoked response for each rest period. This ratio, termed “recovery-ratio,” was averaged across all experiments and compared before and after drugs (Student's t test). The results are illustrated by bar graphs.

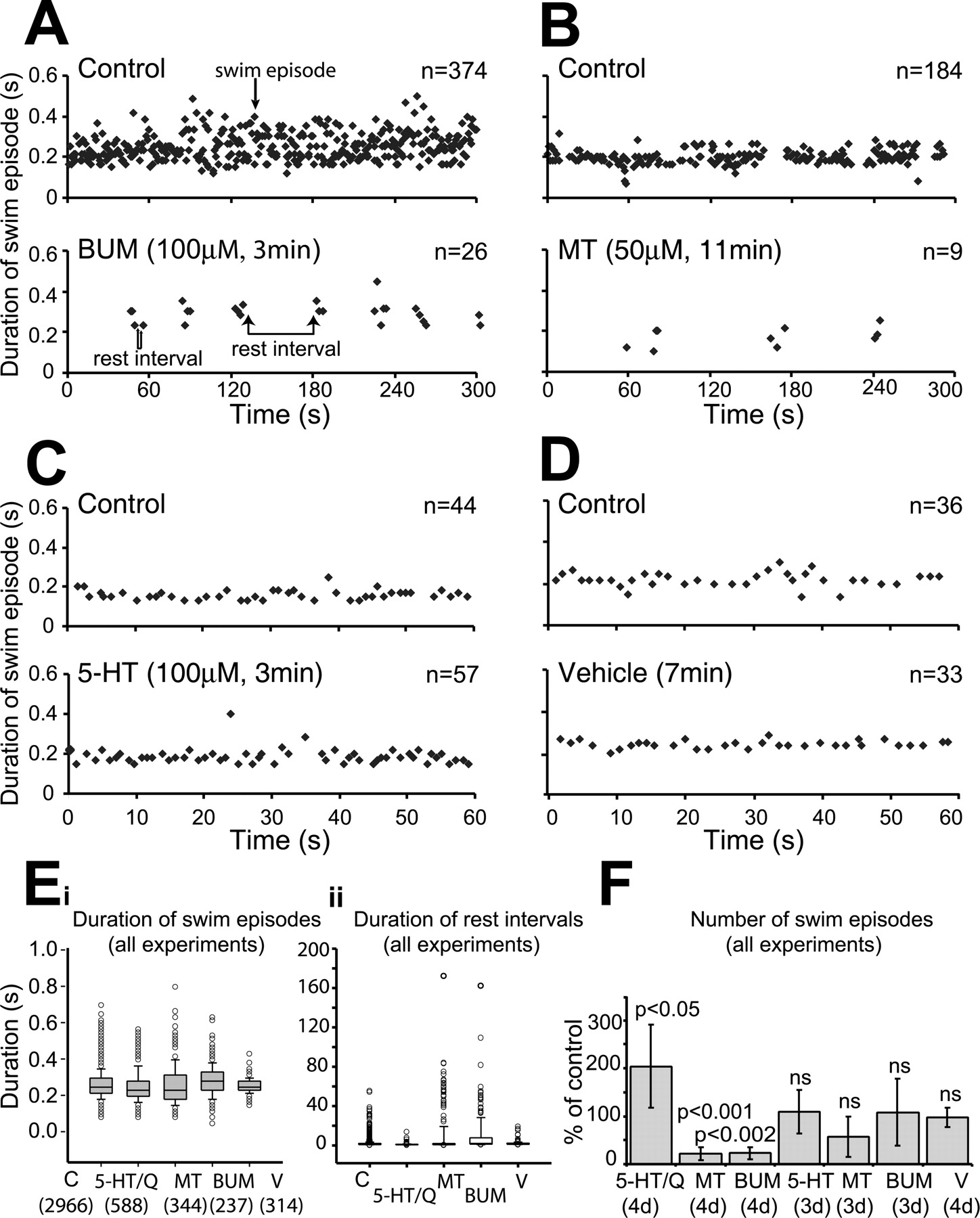
Effects of bumetanide (BUM), serotonergic antagonist methysergide (MT), 5HT, and vehicle on the spontaneous swimming pattern of 4-d-old larvae (behavioral experiments). A, Temporal distribution plots (300-s-long excerpts) of swim episodes (black diamonds) before (control, top) and 3 min after (bottom) bumetanide injection into the pericardial sac. Bumetanide fragmented the robust and consistent swimming pattern into clusters of activity by causing the appearance of long rest intervals lasting over 30 s. B, Temporal distribution plots (300-s-long excerpts) of swim episodes before (control, top) and 11 min after (bottom) methysergide injection. Methysergide, like bumetanide, reduced the number of swim episodes and caused the appearance of long rest intervals. C, Temporal distribution plots (60-s-long excerpts) of swim episodes before (control, top) and 3 min after (bottom) 5HT injection. 5HT, in contrast to bumetanide and methysergide, increased the number of swim episodes by reducing the duration of rest intervals. D, Temporal distribution plots (60-s-long excerpts) of swim episodes before (control, top) and 7 min after vehicle injection (bottom). Vehicle had no effect on the temporal distribution of spontaneous swimming of 4-d-old larvae. E, The box plot on the left (i) summarizes the distribution of swim episode durations across all experiments under different drug conditions: C (control), 5HT/Q (pooled data of 5HT and quipazine), MT (methysergide), BUM (bumetanide), and V (vehicle). All swim episode durations fell into the same range (for statistics, see Results). Event numbers are given in parentheses. The box plot on the right (ii) summarizes the distribution of rest intervals across all experiments. The data show that methysergide and bumetanide caused the appearance of long rest intervals (see outlying symbols), whereas 5HT decreased them significantly (for statistics, see Results). F, Bar graphs summarizing the effects of the different drugs on the number of swim episodes (percentage of control) across all experiments. 5HT and quipazine doubled the number of swim episodes, whereas methysergide and bumetanide decreased it dramatically (5-30% of control) and vehicle had no effect. None of the drugs significantly affected the number of swim episodes of younger 3-d-old larvae (for statistics, see Results). Error bars indicate SD.
Statistics. Most of the experimental results (unless stated otherwise in the text) were not distributed normally and were therefore compared using nonparametric statistical tests (SigmaStat software): the Mann-Whitney rank sum test when comparing two experimental groups and the Kruskal-Wallis ANOVA on ranks when comparing more than two experimental groups. Both in the cellular and behavioral experiments, the effects of the drugs on the different swim pattern parameters, such as number of swim episodes, their durations, swim frequency, and the extent of depolarization, were taken from equal time sessions and compared for each larva before and after drug. Pooled data in each experimental group are usually presented using medians, data ranges, and box plots. Each box plot contains a central line representing the median, the box itself delineating 25-75% of the data, and error bars representing 10-90% of the data range, whereas open circles represent outlying data points. The statistical results were considered different at the level of p < 0.05.
Results
To test whether 5HT modulation of the larval zebrafish swim pattern was exerted through control of chloride homeostasis, the effect of blocking inward chloride transport by bumetanide, the specific blocker of the Na+-K+-Cl- cotransporter (Russell, 2000), was tested and compared with the effects of 5HT and its antagonists, both at behavioral and cellular levels.
The effects of bumetanide and of serotonergic drugs on the behavioral swimming pattern
Figure 1 shows the effects of bumetanide, the serotonergic antagonist methysergide, 5HT, and vehicle on the swimming pattern of the 4-d-old zebrafish larvae. The different drug solutions were injected into the pericardial sac of agarose-embedded larvae with the tail free to move (see Material and Methods), after a period of control video recordings. The injections were made over a range of concentrations (from 50 μm to 1 mm) with a final effective drug concentration estimated to be ∼15 times lower (3-50 μm) (see Material and Methods) (Brustein et al., 2003b). Four representative examples (Fig. 1A,B, top temporal distribution plots, each 300 s long; C,D, top temporal distribution plots, each 60 s long) illustrate the normal swimming pattern of 4-d-old larva, which alternates between swim episodes (20-40 per minute, each lasting ∼0.2 s) and longer and more variable rest intervals (0.7-2 s). This swim pattern was similar to that of free-swimming larvae (Fuiman and Webb, 1988; Budick and O'Malley, 2000; Buss and Drapeau, 2001; Brustein et al., 2003b).
Bumetanide (100 μm; estimated <10 μm in vivo; n = 10) (see Materials and Methods) (Fig. 1A, bottom), similar to methysergide (50-100 μm; estimated <10 μm in vivo; n = 12) (Fig. 1B, bottom), reduced considerably the number of swim episodes to 5-30% of control (Fig. 1F, summaries across all experiments) and caused the appearance of unusually long rest intervals lasting over 30 s (Fig. 1, compare A, B, top and bottom plots; note outlying data points in Eii). Thus, bumetanide, like the 5HT antagonist methysergide [and ketanserin (Brustein et al., 2003b)], fragmented the robust and consistent swimming pattern into clusters of activity. The effects of bumetanide were observed ∼5 min after injection, whereas the effects of methysergide were observed ∼15 min after injection. In both cases, the consequences were longlasting (>1 h) [see methysergide effects in the study by Brustein et al. (2003b)]. Despite the pronounced effect of bumetanide and methysergide on the swimming pattern, 90% of swim episode durations fell into the same range (Fig. 1Ei), with a small but statistically significant difference in the median in the presence of bumetanide (0.26 s) compared with control and after methysergide (0.23 and 0.22 s, respectively; p < 0.05). The effects of bumetanide and serotonergic antagonists contrasted with those of 5HT (n = 10) (Fig. 1C) and those of the 5HT receptor agonist quipazine (Brustein et al., 2003b), which decreased rest intervals (medians, 1.3 and 0.8 s, before and after 5HT/quipazine, respectively; p > 0.001) (Fig. 1Eii) and thereby markedly increased the number of swim episodes by twofold (p < 0.05; t test) (for average, see Fig. 1F). However, neither 5HT nor quipazine affected swim episode durations (Fig. 1E) (medians, 0.23 and 0.22 s, before and after pooled data of 5HT and quipazine, respectively), and no changes were observed in the swimming pattern of vehicle-injected larvae (n = 6) (Fig. 1D). Interestingly, bumetanide, much like the serotonergic receptor agonists and antagonists, did not affect the irregular and variable swimming pattern of 3-d-old larvae (data not shown) or the number of swim episodes (bumetanide, n = 5; methysergide, n = 6; 5HT/quipazine, n = 8) (Fig. 1F). Together, these results indicate that bumetanide had similar effects as the 5HT antagonists tested in increasing rest intervals without affecting active periods of swimming in 4-d-old larvae and not earlier.

Effects of bumetanide (BUM), serotonergic antagonist methysergide (MT), and 5HT on the spontaneous neuronal fictive swimming (whole-cell recordings). A, Representative traces of fictive swimming activity recorded in motoneurons before (control, top), after bumetanide (center), and in washout condition (bottom). The peaks of the spikes have been truncated. Bumetanide (3 min) caused clustering of fictive swim episodes that were separated by long quiescent intervals. This effect was reversible (washout). The asterisks point to expanded segments in D. B, 5HT (center) (Fig. 2C, center) reduced the duration of rest intervals compared with control (top). This effect was blocked by bumetanide (2 min), even in the presence of 5HT (bottom). C, Methysergide (bottom), similar to bumetanide, fragmented the robust fictive swimming induced by 5HT (center). D, Two superimposed fictive swim episodes, before (black line) and after bumetanide (gray line), taken from A (indicated by asterisk) to illustrate the similarity in their properties (duration, frequency, and area; illustrated by gray shade). These are summarized across all experiments in E-G. E, Box plots illustrating that the duration of fictive swim episodes across all experiments [before (C) and after 5HT, methysergide, bumetanide, 5HT plus bumetanide, and washout (W)] was conserved. F, The fictive swimming frequency (calculated from the large EPSPs, three of which are indicated by arrows in D; EPSPs could sometimes carry spikes) was similar across all experimental conditions as illustrated by box plots. G, The extent of depolarization during fictive swim episodes, calculated from the area under the curve (D, gray shade), was the same across all experiments, despite the pronounced effects of the drugs on the swim pattern. Error bars indicate SD.
The effects of bumetanide and of serotonergic drugs on the spontaneous fictive swimming pattern: whole-cell recordings
We compared in greater detail the effects of bumetanide, methysergide, and 5HT on spontaneously occurring “fictive” swimming patterns recorded from identified spinal neurons in paralyzed larvae in vivo (see Material and Methods) by whole-cell, current-clamp patch recording. This allowed us to study cellular as well as network properties, because the fictive swimming pattern in the zebrafish larva is synaptically driven, thus reflecting network activity (Buss and Drapeau, 2001). The swim episode properties, such as duration and swim frequency (estimated as the frequency of large rhythmic EPSPs during the synaptic drive for each period of fictive swimming) (Fig. 2D), resembled the properties of locomotor activity, as established previously (Buss and Drapeau, 2001).
Bath perfusion of bumetanide, after control recordings (n = 3) (Fig. 2A, middle trace) or in the presence of 5HT (n = 7) (Fig. 2B, compare bottom and middle traces), fragmented the fictive swimming pattern into clusters of activity separated by unusually long quiescent periods, similar to our observations in the behavioral experiments described above. This effect was reversible (Fig. 2A, washout) and was equivalent to the effect of methysergide bath application after 5HT (n = 7) (Fig. 2C) or immediately after control recordings (n = 10) (data not shown) (Brustein et al., 2003b). Despite the pronounced effect of bumetanide on the fictive swim pattern, it did not appear to affect the properties of the swimming episodes, such as duration (medians, 0.24, 0.22, and 0.23 s, before and after bumetanide or bumetanide plus 5HT, respectively) (Fig. 2E) or swim frequency (medians, 32, 33, and 30 Hz, before drug and after bumetanide or bumetanide plus 5HT, respectively) (Fig. 2F). However, there was a slight but statistically significant increase in the median of swim frequency in the presence of 5HT and methysergide, 36 and 34 Hz, respectively, compared with 32 Hz in control (p < 0.001). In addition, the extent of depolarization during the swim episode was estimated from the area (milliseconds by millivolts) under the curve (Fig. 2D) and was preserved across all experiments, as summarized in Figure 2G.
The effects of bumetanide on the fictive swim pattern contrasted with the effects of 5HT (Fig. 2B, center trace) and those of the 5HT receptor agonist quipazine (data not shown) (Brustein et al., 2003b). 5HT or quipazine (n = 33) increased the number of fictive swim episodes but did not affect their properties, as summarized in Figure 2E-G. We concluded that bumetanide and the serotonergic antagonists had comparable effects on the behavioral and fictive swimming patterns, because both modulated primarily the duration of the rest intervals without significantly affecting the swim episode properties.
The effects of bumetanide and of serotonergic drugs on evoked glycine responses: perforated-patch recordings
Our observations with whole-cell recordings showed that bumetanide mimicked the effects of serotonergic antagonists, suggesting that 5HT acts by modulating chloride homeostasis. As a direct measure of chloride responsiveness, we studied the effects of 5HT, methysergide, ketanserin, and bumetanide on the amplitude of ionophoretically evoked glycinergic responses in spinal neurons using gramicidin-perforated patches to preserve intracellular chloride (see Materials and Methods). Passive current-clamp recordings were obtained, because zebrafish spinal neurons are small (∼10 μm) and have a high input resistance (150-350 MΩ), which prevented adequate space-clamp with perforated patches. Furthermore, passive current-clamp recordings could give a nonbiased measure of the amplitude of evoked responses and changes in membrane potential. Glycine-evoked responses were preferred over GABAergic ones, as glycine, in contrast to GABA, contributes significantly to the synaptic drive underlying fictive swim episodes in the zebrafish larva (Buss and Drapeau, 2001). In addition, spontaneous glycinergic EPSPs are frequent and can be readily detected, whereas GABAergic EPSPs are rare at this stage of development (Legendre and Korn, 1994; Triller et al., 1997; Ali et al., 2000; Hatta et al., 2001). Therefore, glycinergic inputs to spinal neurons are behaviorally relevant.
Figure 3A shows a representative trace of fictive spontaneous swimming recorded from a gramicidin-perforated patch. Fictive swimming was recorded reliably under these conditions from spinal neurons, with resting potentials between -60 and -75mV. The fictive swim episodes were depolarizing and closely resembled those observed in whole-cell current-clamp recordings (compare Figs. 3A, left; 2B, top trace; with 3A, extended trace; 2D).
Having established that fictive swimming can be monitored with perforated patches, bicuculline (10 μm), kynurenic acid (1 mm), and TTX (1 μm) were added to the bath (in addition to d-tubocurarine; 15 μm) to isolate glycine-evoked responses (see Material and Methods). Ionophoresis of glycine resulted in depolarizing responses that were abolished by the selective glycine antagonist strychnine (Fig. 3B, right trace). The observation that glycinergic (chloride-mediated) responses are depolarizing from rest is consistent with cell-attached recordings in embryos (Saint-Amant and Drapeau, 2000), glycine-evoked calcium transients recorded in larval neurons (Brustein et al., 2003a), and the generally reported elevation of intracellular chloride in embryonic neurons (see Introduction). The glycine-evoked responses were elicited repetitively and consistently (Fig. 3B, left trace).
The effects of 5HT, methysergide, ketanserin, bumetanide, and bumetanide plus 5HT on the amplitude of glycine-evoked responses are summarized in Figure 3C. Each example superimposes a representative set of averaged glycine-evoked responses (15-33 consecutive responses) obtained from recording sessions with equal duration under control, after drug, and washout conditions. In addition, confidence intervals (99%; dashed lines) are illustrated. The narrow range of the confidence interval around the mean further supports the stability of the experimental method. Figure 3Ci shows that 5HT significantly increased the amplitude of the glycine-evoked responses by an average of 141 ± 9% of control across all experiments (p < 0.001; n = 3) (Fig. 3C, bar graph). However, methysergide (Fig. 3Cii) and ketanserin (Fig. 3Ciii) decreased the amplitude of glycine-evoked responses. The total average change across experiments was 38 ± 17% of control for methysergide (n = 3; p = 0.003) and 55 ± 12% for ketanserin (n = 4; p < 0.05) (Fig. 3C, bar graphs). Bumetanide on its own, at a low concentration (35-50 μm), reduced the amplitude of the glycine-evoked responses to 81 ± 5% of control. At these concentrations, bumetanide was not as effective as the 5HT antagonists in reducing the glycinergic-evoked responses, but the effect was nonetheless highly significant (n = 3; p = 0.006) (Fig. 3D). One hundred micromolar bumetanide (Fig. 3Cv), like the 5HT antagonists, reduced the amplitude of the glycine-evoked response, even in the presence of 5HT, to an average of 51 ± 20% across experiments (n = 2; p = 0.017) (Fig. 3D). The effect of the drugs on the amplitude of the glycine-evoked responses was reversible (see washout in each set of experiments) (Fig. 3C,D) (average of 84 ± 14% across experiments; n = 7). It should be noted that bumetanide had no direct, artifactual effect on glycine receptor activity, as verified by analyzing the properties of isolated spontaneous glycinergic miniature EPSCs recorded during whole-cell voltage-clamp experiments. In this experimental paradigm, the chloride in the pipette was fixed to 20 mm, as detailed in Materials and Methods (Fig. 3E) (n = 4; averages expressed as the percentage of control ± SD: amplitude, 90 ± 18, p = 0.65; rise time, 107 ± 17, p = 0.37; half-width, 137 ± 29, p = 0.2; frequency, 107 ± 67, p = 0.82). In summary, the experiments with isolated, glycine-evoked responses showed that bumetanide, methysergide, and ketanserin reduced the amplitude of chloride responses evoked by glycine, whereas 5HT increased them.
The effects of bumetanide and of serotonergic drugs on glycine-evoked responses during spontaneous fictive swimming: perforated-patch recordings
Although the results with isolated glycine responses indicated that the chloride responses were modulated under basal conditions, we sought to determine whether this also occurred during behaviorally relevant activity. Specifically, we wanted to test whether 5HT modulation of chloride homeostasis also had an effect on the activity state of the neural network underlying locomotion. We therefore repetitively evoked glycine responses ionophoretically while recording spontaneous fictive swimming with perforated patches. This methodological approach is based on previous experiments in the chick embryo (Chub and O'Donovan, 2001) in which changes in the amplitude of responses to GABA were used as an indicator for the recovery of the intracellular chloride during the quiescent intervals after spontaneous episodes (Chub and O'Donovan, 2001). A similar approach was used by Wagner et al. (2001) in the suprachiasmatic nucleus to follow recovery of the intracellular chloride after a conditioning train leading to chloride depletion. We adjusted the ionophoresis parameters to avoid initiation of a swim episode and kept them constant for the whole experiment thereafter. The responses were most likely evoked directly in the recorded neuron because of the limited spread of glycine during brief, 50-100 ms ionophoretic pulses (Brustein et al., 2003a).
To illustrate the changes in the amplitude of glycine-evoked responses during rest intervals of spontaneously fictive swimming larvae before and after 5HT, methysergide, and 5HT plus bumetanide (Fig. 4A-F), temporal distribution plots were reconstructed from raw data such as illustrated by Figure 4A, inset. An overall increase (117 ± 19%; p < 0.05; across all experiments) was observed in the amplitude of glycine-evoked responses during rest intervals after 5HT perfusion (Fig. 4, compare A, B, dashed lines), as expected from results using isolated glycine-evoked responses. One can also see that, in the control (Fig. 4A,C), the amplitude of glycine-evoked responses during rest intervals, even when somewhat lengthy, was stable, because there was no difference between the amplitude of the first response in the rest interval versus the last one. This ratio, termed “recovery ratio” (see Materials and Methods), is illustrated in Figure 4G (average of 101 ± 11% across all experiments). A similar recovery ratio was found in the presence of 5HT (Fig. 4B,E,G) (average of 100 ± 7% across all experiments). However, in methysergide (Fig. 4D) and in bumetanide plus 5HT (Fig. 4F), the recovery ratio was reduced to 79 ± 16% (p < 0.001) and 79 ± 18% (p < 0.05), respectively, across all experiments (Fig. 4G). This reduction in the recovery ratio occurred, because the amplitude of the glycine-evoked response (which is indicative of the intracellular chloride concentration) was low immediately after swimming and gradually recovered toward the end of the rest interval, as will be discussed in detail below.

Effects of bumetanide (BUM), serotonergic antagonist methysergide (MT), ketanserin (KE), and 5HT on isolated ionophoretically evoked glycine (Gly) responses (perforated-patch recordings). A, Spontaneous fictive swimming (left trace) was recorded from spinal neurons using gramicidin-perforated patches (the spikes have been truncated). The depolarizing fictive swim episodes (right, expanded traces taken from the boxed segment on the left) resembled those observed with whole-cell recordings (Fig. 2). B, Depolarizing glycine responses, evoked ionophoretically (see Materials and Methods), had constant amplitudes (control, left) and were blocked by strychnine (right). C, Effects of 5HT (i), methysergide (MT) (ii), ketanserin (KE) (iii), bumetanide (BUM) (iv), and 5HT plus bumetanide (5HT plus BUM) (v) on the amplitude of ionophoretically evoked glycine responses. Each example illustrates an averaged glycine response (heavy line) with its interval of confidence (99%, dashed line) in control, after drug, and washout recordings. 5HT (4 min) increased the amplitude of the glycine-evoked responses by 51%, whereas methysergide (2.5 min), ketanserin (4 min), and 5HT plus bumetanide (13 min) decreased it (24-54% of control). Bumetanide (35-50 μm; 3 min) also significantly reduced the amplitude of the glycine-evoked responses (80% of control). These effects were reversible (see washout in each experimental set). D, The effects of the different drugs on the amplitude of glycine-evoked responses across experiments are summarized in the bar graphs. E, Bumetanide (BUM) had no effect on the properties of spontaneous glycinergic mPSCs, as summarized in the bar graphs. The inset superimposes several glycinergic mPSCs (whole-cell voltage-clamp recordings). Error bars indicate SD.
Discussion
Our results show that bumetanide, the specific blocker of the inwardly directed chloride cotransporter (Russell, 2000), and the serotonergic antagonists methysergide and ketanserin (blockers of 5HT1,2 and 5HT2 receptors, respectively), have similar effects on the swimming pattern of the developing zebrafish. Interestingly, 5HT neurons and processes are in close proximity to motoneurons and interneurons of the zebrafish larva at the developmental stages studied here (Brustein et al., 2003b; McLean and Fetcho, 2004). Despite the fact that 5HT immunoreactivity can be detected earlier (Brustein at al., 2003b; McLean and Fetcho, 2004), 5HT affected the swimming pattern only by day 4. This may be because the neural network for swimming is not yet fully established, as discussed in detail by Brustein et al. (2003b). In addition, it is possible that intracellular signaling mechanisms linking 5HT receptor activation and regulation of bumetanide-sensitive chloride cotransport are yet to be established (Marcus and Carew, 1998) (see below).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effects of 5HT methysergide and bumetanide on ionophoretically evoked glycine (Gly) responses during spontaneous fictive swimming (perforated-patch recordings). A-F, Temporal distribution plots of the amplitude of glycine-evoked responses (black symbols) during fictive spontaneous swimming. Swim episodes are illustrated by gray boxes. The plots were reconstructed from raw data as illustrated in the inset in A. During the rest interval, after swimming, the amplitude of the glycine-evoked responses was relatively constant in the control (A, C) and after 5HT (B, E, dashed line). However, after methysergide (D) or 5HT plus bumetanide (F) treatment, the amplitude of the first glycine-evoked response (white symbol) was low after bouts of swimming and gradually recovered during the rest interval (dashed line). G, Bargraphs illustrating the recovery ratio (percentage); the ratio between the amplitude of the first glycine response (first white symbol) in the rest interval and that of the last one in the rest interval (last white symbol) across all experiments under the different experimental conditions, control (C), 5HT, methysergide (MT), and 5HT plus bumetanide (5HT plus BUM). This recovery ratio decreased to ∼80% under methysergide and 5HT plus bumetanide but was constant (∼100%) in the presence of 5HT or in the control (for statistics, see Results). Error bars indicate SD.
The bumetanide-sensitive chloride cotransporter, which is developmentally regulated (Delpire, 2000; Payne et al., 2003), is implicated in maintaining high chloride levels in neurons. This results in depolarizing responses to GABA or glycine (Delpire, 2000; Payne et al., 2003). Furthermore, a positive correlation between mRNA expression levels of the bumetanide-sensitive chloride cotransporter and intracellular chloride levels was found in single immature cortical neurons (Yamada et al., 2004). In our study, depolarizing responses to glycine were observed directly using perforated-patch recordings and were also detected in previous cell-attached recordings (Saint-Amant and Drapeau, 2000) and by calcium imaging (Brustein et al., 2003a). The low effective concentrations of bumetanide (estimated as <10 μm) (see Materials and Methods) suggest that it had a specific effect on the inward chloride cotransporter (Delpire, 2000; Payne et al., 2003). Similar bumetanide concentrations were applied to study GABA responses of interneurons in Xenopus larva in vivo (Rohrbough and Spitzer, 1996). The lack of effect of bumetanide on the swimming pattern before day 4 (despite the high chloride concentration in neurons) was most probably caused by the immature network organization, because 3-d-old zebrafish larvae swim rarely and irregularly (Buss and Drapeau, 2001), as suggested previously for 5HT modulation (Brustein et al., 2003b).
Modulation of swimming
Both bumetanide and the serotonergic antagonists fragmented the robust swimming into clusters of activity by increasing periods of inactivity and reducing the number of swim episodes. However, the properties of the swim episodes (duration, frequency, and extent of depolarization) (for discussion, see Brustein et al., 2003b) were unaffected by these drugs. The effects of bumetanide were in contrast to the effects of 5HT, which reduced periods of inactivity and increased the number of swim episodes. The result that bumetanide, on its own or even in the presence of 5HT, causes an increase in the duration of the quiescent intervals supports the hypothesis that chloride modulation may set the duration of the quiescent intervals, as suggested by data from the embryonic chick spinal cord (Chub and O'Donovan, 2001). There, changes in chloride levels affected the duration of quiescent periods without affecting the duration of activity periods (Marchetti et al., 2005). In addition, a persistent GABAergic conductance determines, in part, the duration of the interepisode interval, and low concentrations of the GABAA antagonist bicuculline could also prolong their duration (Chub and O'Donovan, 2001). GABAergic transmission is rare in the early zebrafish (Legendre and Korn, 1994; Triller et al., 1997; Hatta et al., 2001), and although glycine contributes to the transient synaptic drive during swimming (Buss and Drapeau, 2001), it does not underlie a persistent conductance. In the zebrafish larva, we did not find changes in input resistance during the rest intervals, excluding an important role for changes in basal ionic conductance (Brustein et al., 2003b).
In the chick embryo, application of bumetanide abolished altogether the recurrent activity recorded in the presence of glutamatergic and cholinergic blockers (Chub et al., 1998; Chub and O'Donovan, 2001; Marchetti et al., 2005). In our study, bumetanide application resulted in clusters of swim episodes, and these episodes retained their normal features such as duration and extent of synaptic drive. It is possible that bumetanide did not abolish swimming in the zebrafish larva because it did not affect all of the neurons in the network, as shown in Xenopus spinal neurons (Rohrbough and Spitzer, 1996) and in hippocampal neurons (Misgeld et al., 1986). Moreover, spontaneous activities in the chick spinal cord and other developing networks (O'Donovan, 1999) are not directly comparable with the patterned locomotor activity in the zebrafish, but as discussed below, they may share common mechanisms that shape the activity state of the neuronal network during early development.
Modulation of chloride-mediated responses
Passive current-clamp recording allowed us to monitor natural variations in the membrane potential and, therefore, gave a more reliable estimate of the chloride response. Consequently, taking into account the lack of changes in input resistance (Brustein et al., 2003b), the extent of depolarization in response to glycine before and after drug application was indicative of the naturally occurring changes in the levels of intracellular chloride (Marchetti et al., 2005). Bumetanide (100 μm in the presence of 5HT) caused a decrease (∼50%) in the amplitude of evoked chloride responses. The possibility that bumetanide acts directly on the properties of glycine receptors was excluded, because no changes were observed in the properties of glycinergic mPSCs. The effect of serotonergic antagonists on the evoked-chloride response was similar to the effects of bumetanide. In contrast, 5HT increased the amplitude of the chloride responses by 40%. Therefore, an increase in 5HT levels can apparently increase intracellular chloride levels, as reflected by an increased depolarization when glycine was applied, whereas under low endogenous 5HT levels (in the presence of 5HT antagonists) less chloride accumulated intracellularly, as reflected by the reduced amplitude of the evoked-chloride depolarization.
During spontaneous fictive swimming (perforated patches after 5HT perfusion), an overall increase of 17% was observed in the amplitude of glycine-evoked responses during rest intervals, as expected from results using isolated glycine-evoked responses. In addition, both in control and when 5HT is applied in the bath, the amplitude of the glycine-evoked responses was constant during the rest intervals, as expressed by an average recovery ratio of ∼100%. Although gradual replenishment of chloride was observed in the presence of methysergide or bumetanide (see below), the constant amplitude of the glycine-evoked responses under control conditions is, at first glance, surprising, because we expected chloride depletion after repetitive swimming because of the strong glycinergic synaptic input to motoneurons and interneurons during bouts of swimming (Buss and Drapeau, 2001). However, because the zebrafish larva expresses 5HT endogenously (Brustein et al., 2003b, McLean and Fetcho, 2004), we presume that the rate of inward chloride transport is constitutively modulated and sufficiently high that the intracellular chloride level is reconstituted rapidly (in <2 s, the limit of our glycine application method); thus, changes in the chloride-mediated response were not detected. In the presence of additional exogenous 5HT, the intracellular chloride gradient can be even further emphasized (more depolarization), as seen by an overall increase of 17% in the amplitude of glycine-evoked responses during rest intervals after 5HT perfusion, assuring expression of swimming. Although the time resolution of our experimental method (2 s) (Fig. 4) is presumably not sufficient to resolve the acceleration in chloride replenishment when 5HT is added exogenously, the pronounced decrease in the duration of the rest intervals (from 1.3 to 0.8 s) (Fig. 1) and a twofold increase in the number of swim episodes, observed behaviorally (Fig. 1) and during fictive swimming (Fig. 2), suggests indirectly that exogenous 5HT can further accelerate chloride replenishment. The time course of chloride replenishment could be followed when endogenous 5HT was blocked by the serotonergic antagonist methysergide or when the activity of the inward chloride cotransporter was blocked by bumetanide applied in conjunction with 5HT. Thus, the amplitude of the chloride-mediated responses (Fig. 4) was low after a bout of swim episodes and gradually recovered toward the end of the rest interval, as expressed in a recovery ratio of ∼80%. These results suggest that chloride replenishment is far less efficient when 5HT or the inwardly directed chloride cotransporter are blocked, because it takes minutes (rather than <2 s) for the chloride-mediated responses to recover.
This delay in reconstitution of the chloride gradient may account for the appearance of long quiescent intervals observed in behavioral and during fictive swimming experiments and the reduction in the number of swim episodes (Figs. 1, 2). Similarly, in the embryonic chick spinal cord, chloride currents evoked by a GABAA agonist were suppressed after an episode of activity and gradually recovered during the interepisode interval lasting minutes (Chub and O'Donovan, 2001). There, the changes in the amplitude of chloride currents after activity were attributed to chloride depletion (estimated as a decrease of 15 mm) followed by a gradual replenishment of the intracellular chloride levels during the interepisode interval. In the zebrafish, the constant glycine responses during the intervals between episodes in the control (Fig. 4) suggest that the inward and outward chloride fluxes are balanced. Chloride homeostasis is also maintained when exogenous 5HT is added, but it is set to a different level, as evident from the increase in the average chloride-mediated responses (117%). Little is known, however, about the origin of outward chloride flux at early developmental stages (Payne et al., 2003). The increase in the amplitude of chloride-mediated responses [isolated (Fig. 3) and during fictive swimming (Fig. 4)] and the opposing effect of bumetanide, which is similar to the effects of 5HT antagonists, imply that it is the chloride replenishment mechanism that is affected by endogenous 5HT. Supporting that is evidence that 5HT can modulate the activity of the inward bumetanide-sensitive chloride cotransporter (Mayer and Sanders-Bush, 1994) (see below).
Recently, Marchetti et al. (2005) proposed a model based on the findings in the chick embryo, by which they could vary the duration of the interepisode interval by manipulating solely the rate of inward chloride transport. Accordingly, in the zebrafish, blocking endogenous 5HT or in the presence of bumetanide, the rate of chloride replenishment by the inwardly directed chloride cotransporter is likely reduced, as illustrated by the gradual recovery of the amplitude of the glycine-evoked responses, and may explain the prolonged rest intervals under these conditions. The time scale of recovery under blockade of endogenous 5HT or of the inwardly directed cotransporter (minutes), the saturating levels of transmitter used in our experiment to evoke chloride-mediated responses, and the lack of effects of either of the tested drugs on the active (swim) episode further exclude the implication of a presynaptic mechanism (for detailed discussion and related literature, see Brustein et al., 2003b). Therefore, in the developing zebrafish, much like in the chick embryo (Marchetti et al., 2005), the time taken for chloride replenishment could determine the activity state of the neuronal network.
5HT and chloride homeostasis
One of the most significant findings of our study is that bumetanide occluded the 5HT effects, both during fictive swimming and when tested on isolated chloride-mediated responses, suggesting that 5HT and bumetanide effects converge. It is possible, therefore, that endogenous 5HT modulates the activity of the bumetanide-sensitive (inward) chloride cotransporter. Modulation of this cotransporter by neurotransmitters such as GABA (Sun and Murali, 1999; Schomberg et al., 2003) and glutamate (Sun and Murali, 1998; Sun and Murali, 1999; Schomberg et al., 2001) was demonstrated in cultured immature cortical neurons. Interestingly, activation of 5HT2 receptors expressed in fibroblasts increased the activity rate of the bumetanide-sensitive (inward) chloride cotransporter (Mayer and Sanders-Bush, 1994).
Modulation of the chloride gradient is not necessarily limited to developing networks and may occur in the adult. GABAA responses in neurons of the suprachiasmatic nucleus switch between excitatory and inhibitory because of day and night variations in intracellular chloride levels (Wagner et al., 1997, 2001), because the bumetanide-sensitive inward chloride cotransport is activated during the day and is downregulated during the night (Wagner et al., 2001). The mechanisms underlying this modulation are unknown. Under pathological conditions (Payne et al., 2003, Rivera et al., 2005), a depolarizing chloride gradient may reappear in the adult as a recapitulation of a developmental program necessary for repair of lesions. Then, chloride homeostasis might be regulated through mechanisms observed during development (Marchetti et al., 2005).
In summary, our results from behavioral and fictive experiments suggested that endogenous 5HT levels modulate chloride homeostasis to determine the activity state of the developing neuronal network underlying locomotion in the zebrafish larva, with important behavioral consequences. We speculate that endogenous 5HT at this early stage of development, before the expression of specific conductances normally modulated by 5HT in other preparations (Schmidt and Jordan, 2000; Brustein et al., 2003b), can promote chloride replenishment between periods of activity, thus offsetting long periods of quiescence to sustain activity.
Footnotes
- Received May 19, 2005.
- Revision received September 26, 2005.
- Accepted September 28, 2005.
This work was supported by a Canadian Institute of Health Research fellowship to E.B. and grant to P.D. We are indebted to Dr. Serge Rossignol and Philippe Drapeau for video and analysis software. G. Laliberté is thanked for excellent animal care. Dr. J. McDearmid is appreciated for his helpful comments on this manuscript.
Correspondence should be addressed to Dr. Pierre Drapeau, McGill Center for Research in Neuroscience, 1650 Cedar Avenue, Montréal, Quebec, Canada H3G 1A4. E-mail: pierre.drapeau{at}mcgill.ca.
Copyright © 2005 Society for Neuroscience 0270-6474/05/2510607-10$15.00/0