

Abstract
Memory, as measured by changes in an animal's behavior some time after learning, is a reflection of many processes. Here, using a trace paradigm, in mice we show that de novo protein synthesis is required for acquisition, consolidation, reconsolidation, and extinction of classically conditioned eyelid responses. Two critical periods of protein synthesis have been found: the first, during training, the blocking of which impaired acquisition; and the second, lasting the first 4 h after training, the blocking of which impaired consolidation. The process of reconsolidation was sensitive to protein synthesis inhibition if anisomycin was injected before or just after the reactivation session. Furthermore, extinction was also dependent on protein synthesis, following the same temporal course as that followed during acquisition and consolidation. This last fact reinforces the idea that extinction is an active learning process rather than a passive event of forgetting. Together, these findings demonstrate that all of the different stages of memory formation involved in the classical conditioning of eyelid responses are dependent on protein synthesis.
- anisomycin
- c-Fos
- classical conditioning
- eyelid responses
- mice
- motor learning
- acquisition
- extinction
- retrieval
- consolidation
- reconsolidation
Introduction
The formation of long-term memory (LTM) seems to involve the conversion of a labile short-term memory (STM) into a lasting stable trace (Hebb, 1949). It has been shown that electroconvulsive shocks or protein synthesis inhibitors administered shortly after learning produce amnesia; however, when applied several hours later, they do not affect the stored memory trace (Duncan, 1949; McGaugh, 1966, 2000). The rationale of how STM evolves into LTM is that the synthesis of new RNA and proteins makes temporary changes in synaptic efficacy become persistent modifications of synaptic functioning and architecture (Davis and Squire, 1984; Goelet et al., 1986). This conceptual framework has been called the consolidation theory (Dudai and Morris, 2000).
The consolidation theory was challenged by early studies demonstrating that amnesia could also occur if a fully consolidated LTM is reactivated before amnesia-evoking treatments (Misanin et al., 1968; Sara, 2000). This evidence suggests that reactivated memories undergo a new round of consolidation, a process designated reconsolidation (Przybyslawski and Sara, 1997; Nader et al., 2000a,b). Consolidation and reconsolidation share some common properties, including (1) the requirement of protein synthesis for the memory to persist and (2) time windows during which protein synthesis blockade is effective, impairing the persistence of memory traces. Thus, rather than a single occurrence, memory storage is thought to be a process that is repeated each time the trace is activated.
Extinction is another process related to learning, during which previously established responses are suppressed. It is not known whether extinction is a passive process of erasure or a new learning task. In fact, although the response to the conditioned stimulus is attenuated during extinction, the original association is surprisingly unaffected (Rescorla, 2001). This suggests that the processes operating during extinction act by suppressing the learned response rather than erasing the original memory trace.
Classical conditioning of eyelid responses is an associative motor learning paradigm, which is acquired progressively. The precise cerebral structures involved in this type of motor learning are still the subject of intense controversy. Some groups think specific neural sites are responsible for the acquisition and consolidation of eyelid conditioned responses (CRs) (Aou et al., 1992; Kim and Thompson, 1997; Medina et al., 2000; Attwell et al., 2002), whereas others propose the participation of several cerebral areas in a determined temporal sequence (Llinás and Welsh, 1993; Bloedel and Bracha, 1995; Steinmetz, 2000) or even the activation of preestablished neural programs distributed through wide cerebral areas (Delgado-García and Gruart, 2002). Clearly, the acquisition of trace eyelid CRs is impaired by lesions in the hippocampus (Solomon et al., 1986; Weiss et al., 1999; Takatsuki et al., 2003) and/or in the cerebellum (Woodruff-Pak et al., 1985), and recent reports suggest that lesions in the hippocampus or in the prefrontal cortex impair consolidation of eyelid CRs (Kim et al., 1995; Takehara et al., 2002, 2003). Here we administered the protein synthesis inhibitor anisomycin at different times during acquisition, extinction, and reactivation to isolate the effects of protein synthesis inhibition on these processes independently.
Materials and Methods
Subjects and surgery
Male Swiss mice (n = 180) supplied by an official dealer (Animal Services, University of Granada, Granada, Spain) were used. At the time of surgery, they weighed 30-40 g. Animals were housed in standard polycarbonate cages in a colony room under a light/dark cycle of 12 h. Water and food were available ad libitum. All experiments were performed in accordance with the guidelines of the European Union (EU) Council (86/609/EU) and following Spanish regulations (Boletín Oficial del Estado 67/8509-12, 1988) for the use of laboratory animals in chronic experiments.
Under general anesthesia (a mixture of ketamine and xylazine, 100 and 10 mg/kg, i.p.), animals were implanted with four Teflon-coated stainless steel wires (7910; A-M Systems, Carlsborg, WA). Two of these (the stimulating electrodes) were implanted in the subcutaneous tissue of the right upper eyelid, near the supraorbitary branch of the trigeminal nerve, whereas the other two (the recording electrodes) were placed ipsilaterally, in the caudal portion of the orbicularis oculi muscle (OOM). The wires were then soldered to a four-pin connector that was secured to the skull with dental acrylic resin and two stainless steel screws.
Drug administration
Anisomycin (Sigma, St. Louis, MO) was dissolved in saline and adjusted to pH 7.4 using NaOH. Anisomycin is a potent inhibitor of mRNA translation via interference with transpeptidation and has been used successfully in a number of behavioral paradigms (Davis and Squire, 1984; Lattal and Abel, 2001). Mice received subcutaneous injections of 100 mg/kg anisomycin or an equivalent volume of saline. Injections were performed 30 min before or 30 min or 4 h after the beginning of each session, depending on the experiment. Kainic acid was injected intraperitoneally at a dose of 15 mg/kg (Sonnenberg et al., 1989).
Locomotor activity box
Locomotion was measured in a locomotor activity box (26 × 39 cm; Cibertec, Madrid, Spain) as indicated by the number of broken light beams during periods of 5 min. For this study, anisomycin was injected daily (100 mg/kg, s.c.) for 8 d. Measurements of locomotor activity were performed 1 h after drug administration on days 1, 3, 6, and 8. Effects of anisomycin on locomotor activity 1 h after its first administration were evaluated with a t test. A two-way ANOVA was applied to determine any possible difference in the effects of anisomycin on locomotor behavior between successive days; t tests with Tukey correction were used for post hoc comparisons.
Conditioning apparatus and procedures
Four cylindrical Plexiglas containers were placed in a sound- and light-attenuated chamber. One mouse was placed in each container. Four low-weight wires were connected to the socket implanted on the animal's head. Conditioned stimuli (CS) and unconditioned stimuli (US) consisted respectively of 50 and 500 μs square cathodic electrical pulses delivered through the stimulation electrodes. Animals were trained using a trace-conditioning paradigm. For this, a stimulus-free interval (250 ms) was interposed between the end of the CS and the onset of the US. Optimal CS and US intensities were determined before the beginning of the habituation sessions. The CS intensity was set at threshold intensity to evoke an eyelid response, as recorded in the electromyographic (EMG) activity of the OOM. The US had the minimal intensity necessary to evoke a complete closure of the eyelid. Across conditioning, eyelid CRs were monitored by the EMG activity of the OOM. Additional details of this classical conditioning preparation in mice have been published previously (Domínguez-del-Toro et al., 2004).
The mean values of CS intensities were 0.67 ± 0.07, 0.57 ± 0.06, 0.68 ± 0.09, and 0.5 ± 0.11 mA for saline, 30 min before (-30 min), and 30 min and 4 h after the start (+30 min and +4 h) of the training session for anisomycin-injected mice, respectively. For the US, the values were 0.5 ± 0.03, 0.52 ± 0.06, 0.56 ± 0.06, and 0.52 ± 0.12 mA for saline, -30 min, +30 min, and +4 h for anisomycin-injected mice, respectively. In both cases, the differences were not significant (F(3,57) = 0.76; p = 0.51 for CS values; F(3,57) = 0.91; p = 0.44 for US values, respectively).
Behavioral procedures
One week after surgery, the EMG activity of the OOM was recorded during three habituation sessions. In each of them, a total of 60 CS-alone trials were performed. Afterward, eight conditioning sessions were performed. Each conditioning session consisted of 60 CS-US paired presentations. Trials were separated by a variable intertrial interval, which was pseudorandomized between 25 and 35 s, with a mean of 30 s. During this period, the animal was assumed to associate the CS with the US and to perform a protective conditioned eyelid response before US presentation. Finally, four extinction sessions were performed, each one consisting of 60 CS-alone trials. Every pseudoconditioned animal underwent 10 recording sessions. These sessions consisted of 60 CS and US unpaired presentations, with a randomly variable interstimuli interval ranging between 1 and 30 s. Every session was delivered with an intersession interval of 24 h. Unless otherwise indicated, eight animals per group were used.
Experiment 1: effects of anisomycin on acquisition and consolidation. Either anisomycin or saline was administered at -30 or +30 min or +4 h for each conditioning session. To test the effect of anisomycin on consolidation, the subjects of each group underwent seven additional conditioning sessions (relearning) 10 d after the end of conditioning.
Experiment 2: effects of anisomycin on reconsolidation. Ten days after the end of conditioning, a dose of 100 mg/kg anisomycin was administered at -30 or +30 min or +4 h with respect to the beginning of a conditioning session (reactivation session). Six and 48 h after the reactivation session, reconsolidation was evaluated in an additional conditioning session.
Experiment 3: effects of anisomycin on acquisition and consolidation of extinction. After the conditioning process, either anisomycin or saline was administered at -30 or +30 min or +4 h with respect to the beginning of the four extinction sessions. Ten days after extinction, the subjects of each group underwent one additional extinction session performed to test extinction consolidation.
EMG analysis
The EMG activity was filtered using a bandwidth ranging from 1 to 10 kHz, digitized on-line at a sampling rate of 10 kHz, and stored on a computer. As criteria, the EMG activity of the OOM recorded in the CS-US interval was considered a “conditioned response” if it lasted >10 ms and was initiated >50 ms after CS onset. Moreover, the rectified and integrated EMG activity recorded during the CS-US interval had to be at least 2.5 times larger than the averaged activity recorded immediately before CS presentation (Domínguez-del-Toro et al., 2004). Those trials presenting EMG activity during the 200 ms before CS presentation or with an evident startle response were rejected from the analysis. The percentage of trials containing a CR in a given session was calculated from the collected EMG records. The basic properties of the reflex blink response were studied in the EMG responses provoked by CS presentations during habituation and conditioning (fourth and eighth) sessions.
Differences in the amplitude and latency of the R1 and R2 components of the EMG response during reflex blinks were evaluated with two-way ANOVA, with drug administration as the between-subject factor (saline or anisomycin) and conditioning session as the within-subject factor (habituation and fourth and eighth conditioning sessions); t tests with Tukey correction were applied for the post hoc comparisons.
Performance during acquisition, consolidation, and reconsolidation of memory was expressed as the mean percentage of eyelid conditioned responses per session. Changes in behavior as a function of drug administration were evaluated by applying a one-way ANOVA with the conditioning session as the within-subject factor (nine sessions) and drug administration as the between-subject factor (saline, -30 and +30 min, and +4 h); t tests with Tukey correction were applied for the post hoc comparisons.
Tissue preparation and immunohistochemistry
To analyze the induction of c-Fos immunoreactivity, several groups, each comprising three untrained animals, were used. Kainic acid (KA; 15 mg/kg) and saline or anisomycin were coadministered at the times indicated in Figure 1 A. Mice from each experimental group were killed by decapitation 30 min after kainic acid administration. Additionally, three sham-paired mice treated with two injections of saline (at the times indicated for kainic acid or anisomycin injections) were included as a control. Their brains were removed and placed on an ice-cold plate. The tissue was fixed by immersion in 4% paraformaldehyde in PBS for 24 h at 4°C and cryoprotected in 30% sucrose in PBS for 2 d at 4°C. Brains were then embedded in 30% sucrose and kept at 4°C until cryotome sectioning. Sagittal sections (50 μm) were obtained from each brain, and processed for free-floating immunohistochemistry (de los Santos-Arteaga et al., 2003). The specific c-Fos antiserum (SC-52; Santa Cruz Biotechnology, Santa Cruz, CA) was used at a 1:4000 dilution. Densitometry and neuron quantification of sections were analyzed with image-J software (Sun Microsystems, Santa Clara, CA).

Anisomycin transiently inhibits protein synthesis. A, Representation of KA and anisomycin administration. KA was always injected 30 min before the animal was killed, whereas anisomycin was administered at variable times (indicated by shaded arrows) with respect to KA injection (indicated by the filled arrow). Negative values on the timeline indicate anisomycin administration before KA; 0 indicates simultaneous administration; and positive values represent administration after KA injection (n = 3 per group). B, Representative hippocampal c-Fos protein immunohistochemistry obtained from anisomycin alone-treated (Aniso), KA alone-treated (Sal + KA), and anisomycin- and KA-treated animals. The time between anisomycin and KA injections is indicated in the top right corner. C, Quantitative analysis of c-Fos expression in brain areas related to processes of learning and memory: hippocampus, piriform cortex, and amygdala. Also, the protein inhibition index is shown. CA1, CA1 field; CA3, CA3 field; DG, dentate gyrus; Hp, hippocampus; Pir Cx, piriform cortex; Amyg, amygdala nucleus; Sal, saline. Scale bar, 500 μm.
The percent ratio of c-Fos positive cells in (KA-KA/anisomycin)/(KA-saline) was used as a measure of protein synthesis inhibition. Hence, values of >100% indicate that anisomycin inhibits KA-induced c-Fos expression. It should be noted that these values are not comparable with the levels of protein synthesis inhibition determined by using protein incorporation of radiolabeled methionine. Neurons were considered c-Fos-positive if the densitometric intensity was greater than two thirds the maximum intensity measured by the image-J software.
To study the possible effects of anisomycin administration on neural structures, three mice received eight daily anisomycin injections. As a control, three mice received eight daily saline injections. Brains were extracted 1 h after the last injection and processed as described above. The specific neuronal-specific nuclear protein (NeuN; Chemicon, Temecula, CA), glial fibrillary acidic protein (GFAP; DakoCytomation, Carpinteria, CA), and calbindin (Swant, Bellinzona, Switzerland) antisera were used at 1:1000 dilutions.
Results
Anisomycin transiently inhibits protein synthesis in brain
To investigate whether protein synthesis was necessary for acquisition, consolidation, reconsolidation, and extinction of eyelid CRs, anisomycin was administered systemically during different phases of the learning storage process. Previously, the temporal course of anisomycin-evoked protein synthesis inhibition was determined. For this, the induction of c-Fos expression by a single dose (15 mg/kg) of kainic acid was used as an index of protein synthesis. In the brain, c-Fos expression is detected as soon as 30 min after a single injection of kainic acid (Sonnenberg et al., 1989). Therefore, a 100 mg/kg dose of anisomycin was administered 6, 4, 2, and 1 h and 30 min before, simultaneously with, or 15 min after kainic acid injection (Fig. 1A). Kainate-dependent c-Fos expression was detected only when anisomycin injection was performed 15 min after or 6 h before kainate administration (Fig. 1B). Furthermore, we evaluated (by densitometry or by counting the number of c-Fos-positive neurons) protein synthesis inhibition in different cerebral areas implicated in learning and memory storage: hippocampus (dentate gyrus, CA3, and CA1), amygdala (Lee and Kim, 2004), and piriform cortex (Gruart et al., 2000b). In all of these areas, the percentage of inhibition ranged from 96.69 ± 1.87%, in dentate gyrus to 151 ± 19%, in piriform cortex (Fig. 1C). No c-Fos expression was detected after KA administration in cerebellum or motor cortex. These results indicate that (1) anisomycin required 15-30 min after injection to reach a sufficient concentration to block >95% of the de novo protein synthesis in the brain, and (2) the inhibitory effect of anisomycin on protein synthesis is reversible and lasts ∼6 h.
Anisomycin administration does not alter brain structure
We also explored whether daily anisomycin administration for 8 d would affect the cellular organization of neuronal structures. Coronal sections from brains of saline- and anisomycin-treated mice were compared using different neuronal and glial markers (Fig. 2). NeuN (an antibody widely used as a selective neuronal marker) immunostaining revealed that the cellular composition and orientation of motor, hippocampal, and cerebellar cortices were not different in saline- and anisomycin-treated mice (Fig. 2a-f). Furthermore, GFAP immunostaining revealed a similar glial distribution in the hippocampus (a cerebral area very sensitive to neuronal insults) in saline- and anisomycin-injected animals (Fig. 2g-j). Finally, in both groups of animals, calbindin immunostaining revealed normal mossy fiber projections from the dentate gyrus to the CA3 region (Fig. 2k-m) and Purkinje cell dendritic arborizations in the molecular layer of the cerebellar cortex (Fig. 2l-n). In conclusion, subchronic anisomycin administration did not cause any noticeable alteration in brain morphology with respect to saline-injected animals.

The morphology of motor, hippocampal, and cerebellar cortices was not altered by anisomycin injection. a-f, NeuN immunostaining of the motor, hippocampal, and cerebellar cortices from saline-injected (a-c) and anisomycin-injected (d-f) animals (magnification, 5×). g-j, GFAP immunostaining of the hippocampus obtained from saline-injected (g, h) and anisomycin-injected (i, j) mice. As illustrated at 20× magnification, there was no glial reaction in the CA3 layer in saline-injected (h) or anisomycin-injected (j) animals. k-n, Calbindin immunostaining of the hippocampal and cerebellar cortices of saline-injected (k, l) and anisomycin-injected (m, n) mice. The lack of anisomycin-induced changes in the mossy fiber tract is evidenced by the absence of any noticeable difference in the staining of the stratum lucidum (CA3 field) between saline-treated (k) and anisomycin-treated (m) mice (magnification, 20×). Note the intactness of the Purkinje cell dendritic arborization in the molecular layer of the cerebellum after anisomycin treatment (n) compared with saline-treated animals (l). CA1, CA1 field; CA3, CA3 field; DG, dentate gyrus; PCL, Purkinje cell layer; MF, mossy fiber tract. In all cases, n = 3 per group.
Anisomycin administration does not alter locomotor behavior
Because systemic administration of protein synthesis inhibitors can produce general health effects, locomotor responses were studied under nonstressful conditions (locomotor activity box) 1 h after anisomycin administration. Locomotor activity values during the first 5 min of the test were 655 ± 33.5 broken beams for saline-injected mice and 563.6 ± 76.8 for anisomycin-injected mice, values that were not significantly different (t(15) = 1.27; p = 0.22). Because some experiments required anisomycin administration during 8 consecutive days, we also determined the temporal course of its effects on locomotor activity after 1, 3, 6, and 8 d of administration. Although anisomycin-injected mice tended to show less locomotor activity than saline-injected mice, the differences were not significant across the 8 d of observation (F(7,55) = 1.22; p = 0.22). Summarizing, anisomycin did not induce any significant health-deteriorating effect on mice, even when injected for as long as 8 consecutive days.
Anisomycin administration does not affect reflex eyelid responses
The next experiment determined the effect of anisomycin on the activity of the eyelid motor system. For this, the EMG activity of the OOM evoked by CS presentations during habituation and conditioning sessions was recorded, and the latency and amplitude of the R1 and R2 components of the reflex response were measured (Fig. 3A). The R1 component is a reflection of the direct activation of the facial motoneurons by a disynaptic circuit involving the trigeminal nucleus, whereas the R2 component requires previous activation of the reticular formation (Sanes and Ison, 1983; Gruart et al., 1995). R1 and R2 latencies during habituation were 7.13 ± 0.29 and 16.57 ± 0.69 ms, respectively (Fig. 3B). These latencies did not change in the saline-injected mice during the successive conditioning sessions (F(2,20) = 0.21; p = 0.89 for R1; F(2,20) = 0.34; p = 0.71 for R2). In anisomycin-injected animals, R1 and R2 latencies did not change during the conditioning sessions compared with the values observed during habituation (F(2,25) = 1.94; p = 0,166 for R1; F(2,20) = 1.71; p = 0.2 for R2). Furthermore, direct comparison between saline- and anisomycin-injected mice latencies did not show significant differences (F(4,36) = 1.48; p = 0.23 for R1; F(4,32) = 1.69; p = 0.17 for R2). Because amplitude analysis gives an idea of a possible reflex response sensitization, R1 and R2 amplitudes were measured along the training (habituation and fourth and eighth conditioning sessions) in saline- and anisomycin-injected mice (Fig. 3C). R1 and R2 amplitudes did not change in the saline- or anisomycin-injected mice during the successive conditioning sessions (saline-injected mice, F(2,20) = 0.34; p = 0.71 for R1; F(2,20) = 0.43; p = 0.65 for R2; anisomycin-injected mice, F(2,20) = 0.13; p = 0.87 for R1; F(2,20) = 0.63; p = 0.54 for R2). Moreover, the comparison of R1 and R2 amplitudes between saline- and anisomycin-injected mice did not show significant differences (F(4,36) = 0.41; p = 0.79 for R1; F(4,32) = 0.36; p = 0.83 for R2). These results, together, indicate that anisomycin administration did not impair the normal kinematics of the eyelid motor system. Therefore, the possible effects of anisomycin on learning and memory processes were not the consequence of a change in the kinematics of the eyelid motor system. In contrast with previous reports in cats (Gruart et al., 1995) and rabbits (Gruart et al., 2000a), mice did not present any evidence of reflex response sensitization (i.e., α responses) in a similar (shock-shock) trace-conditioning paradigm.

Anisomycin administration does not affect the electrical activity of the OOM. A, Representative EMG records from the OOM showing R1 and R2 components of a reflex blink evoked by the electrical stimulation of the supraorbitary branch of the trigeminal nerve in saline- and anisomycin-injected mice. B, C, Graphs representing the mean ± SEM of the latency (B) and voltage amplitude (C) of R1 and R2 components during habituation and during the fourth and eighth conditioning sessions in saline-injected (Sal; open bars) and anisomycin-injected (Aniso; filled bars) animals (n = 20 per group). H, Habituation; C4, C8, fourth and eighth conditioning sessions, respectively.
Trace weak-strong electrical shock eyeblink classical conditioning is an associative learning paradigm
Before testing the role of protein synthesis inhibition in the different phases of learning and memory, we tested whether weak-strong electrical shock eyeblink classical conditioning was an associative motor learning. To do so, we compared the learning curves of mice submitted to conditioning and pseudoconditioning protocols. For conditioning, we used a trace paradigm. Thus, when a weak electrical stimulation (CS) was consistently paired with a strong electric shock (US), both of them applied to the supraorbitary branch of the trigeminal nerve, the subjects gradually displayed significantly more frequent (F(8,133) = 32.53; p < 0.001) eyelid CRs (Fig. 4A, filled circles). Initially, these responses were infrequent (Fig. 4B); however, as training went on, the conditioned responses acquired an in crescendo profile in the EMG activity of the OOM (Fig. 4B). Although the percentage of eyelid CRs during the first four conditioning sessions was not significantly different between the conditioning and pseudoconditioning groups, after the fifth session, it became significantly greater in the former (p < 0.01). In contrast, during pseudoconditioning, the OOM response to a weak electrical stimulation did not significantly change (F(7,22) = 0.11; p = 0.99) when it was randomly associated with a strong electric shock of the supraorbitary branch of the trigeminal nerve (Fig. 4A, open circles). In this case, recordings did not change along the pseudoconditioning process (Fig. 4C).

A weak-strong electrical shock of the supraorbitary branch of the trigeminal nerve, with a trace paradigm, was used for classical conditioning of eyelid responses. A, Differential progress in the percentage of CRs per session across conditioning between conditioned (Cond.; filled circles) and pseudoconditioned (Pseudo.; open circles) groups (n = 8 per group). B, C, Representative single-trial records of the EMG of the OOM during different training sessions obtained from one subject of the conditioned (B) and pseudoconditioned (C) groups. H, Habituation; C1-C8, first through eighth conditioning sessions; PC8, eighth pseudoconditioning session.
In accordance with the above results, the observed changes in the response to the paired CS-US presentation across conditioning should be attributed to a process of associative learning rather than to a simple sensitization of the blink reflex.
Anisomycin administration before classical conditioning training impairs acquisition of eyelid CRs
As described above, anisomycin administration did not deteriorate the animal's general health or eyelid motor performance, either after single administration or after subchronic treatment. Therefore, the capability of anisomycin administration to distort learning processes (i.e., the acquisition, consolidation, and reconsolidation of CRs) in mice was tested. The drug was injected at three different times: -30 or +30 min or +4 h for each conditioning session (Fig. 5A).
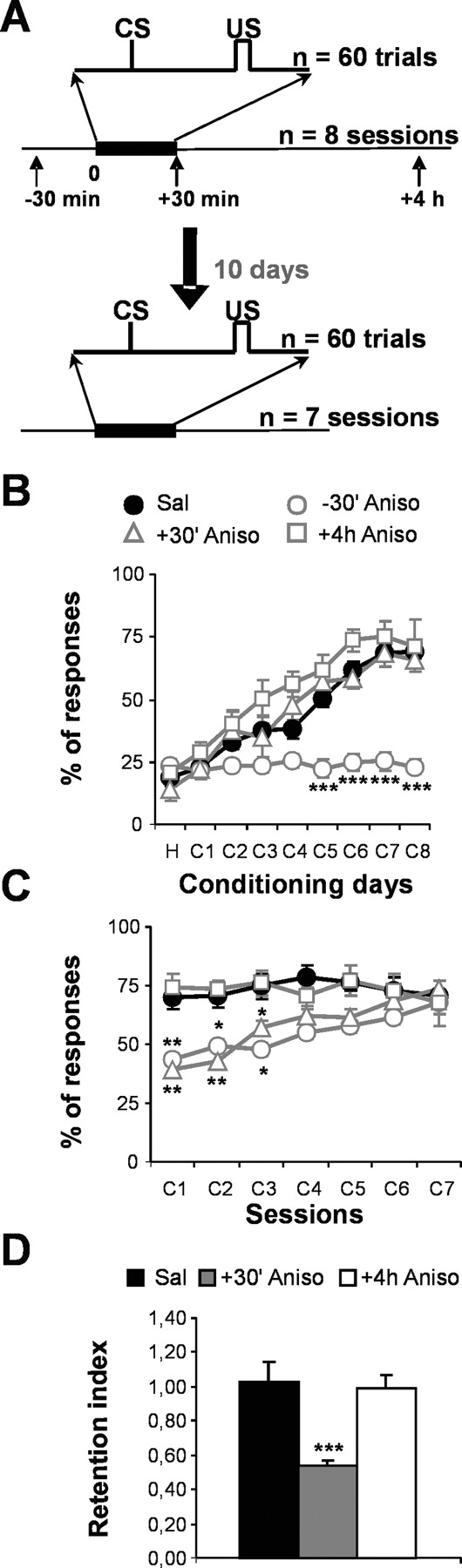
Two critical periods of protein synthesis are required for acquisition and consolidation of eyelid conditioned responses. A, Diagram illustrating the two phases of the experiment. Phase 1 consisted of eight conditioning sessions. During each session, 60 CS-US trials were presented. In this phase, either 100 mg/kg anisomycin (Aniso; experimental subjects) or saline (Sal; control subjects) was injected at variable times from the beginning of each session: -30 and +30 min and +4 h. Because all saline groups were indistinguishable, their results were grouped for the sake of simplicity. For phase 2, the above-mentioned groups received seven additional training sessions 10 d after the end of phase 1; however, neither anisomycin nor saline was administered during these additional sessions. B, C, Acquisition (phase 1) and retention (phase 2) of eyelid CRs, expressed as percentage of response (mean ± SEM). Filled circles, Saline-injected animals; open circles, triangles, squares, mice injected with anisomycin at -30 and +30 min and +4 h, respectively (n = 8 per group). D, The retention index represents the ratio between the percentage of CRs during the first training session of phase 2 and the last training session of phase 1. H, Habituation; C1-C8, first through eighth conditioning sessions; *p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001.
Saline-injected mice exhibited a gradual increase in the percentage of eyelid CRs, which reached a maximum (65-70%) by the seventh conditioning session (F(8,133) = 32.53; p < 0.001) (Fig. 5B). In contrast, mice injected with anisomycin at -30 min showed no significant increase in the percentage of eyelid CRs, which remained at habituation values (20-30%) along the eight conditioning sessions (F(8,32) = 0.07; p = 1). Although the percentage of eyelid CRs during the first 4 conditioning sessions was not significantly different between the saline-injected mice and those injected with anisomycin at -30 min, it became significantly lower after the fifth session in the latter (p < 0.001). The percentages of eyelid CRs for the animals injected with anisomycin at +30 min and +4 h were not significantly different from those of the saline-injected mice. These data indicate that protein synthesis during each training session is required for acquisition of eyelid CRs.
Anisomycin administration just after (+30 min) conditioning sessions blocks consolidation of eyelid CRs
The next experiment examined the effect that anisomycin injection around the conditioning sessions had on memory consolidation. To evaluate consolidation, seven additional conditioning sessions (relearning) were performed 10 d after the conditioning phase (Fig. 5A). Along the whole relearning sessions, saline-injected mice and those injected with anisomycin at +4 h maintained the percentage of eyelid CRs at the level reached by the end of the eight acquisition sessions (Fig. 5C). Because their retention indices (percentage of CRs in session 1 of relearning/percentage of CRs in session 8 of acquisition) were close to unity, clearly, mice of both groups had completely consolidated the newly learned task (Fig. 5D). This result suggests that the molecules involved in long, stable storage of information have already been synthesized 4 h after the end of each conditioning session.
The subjects of the group injected with anisomycin at +30 min acquired eyelid CRs (Fig. 5B). However, their poor performance during the first relearning session (t(10) = 7.97; p < 0.001) (Fig. 5C) indicates that they consolidated this new task incompletely, as evidenced by their low retention index (Fig. 5D). To eliminate a putative effect of the subchronic administration of anisomycin on the neuronal circuit responsible for the generation of eyelid CRs in this group, the effect of seven relearning sessions on the evolution of the percentage of CRs was evaluated. The subjects of this group maintained their capacity to acquire eyelid CRs (F(6,21) = 15.11; p < 0.001). These results suggest that the synthesis of new protein occurring 30 min-4 h after the beginning of each session was necessary for the long-term consolidation but not for the acquisition of CRs.
Additionally, the reacquisition of eyelid CRs in the group injected with anisomycin at -30 min during seven relearning sessions was evaluated. In this group, the percentage of eyelid CRs increased from the first session (43.57 ± 0.84%), reaching the control level on the seventh day of relearning: 67.63 ± 5.11% in the group injected with anisomycin at -30 min against 71.05 ± 5.79%, in the saline-injected group (F(6,28) = 15.41; p < 0.001) (Fig. 5C). These data ruled out any lasting disruption of the neuronal circuits generating eyelid CRs, produced by the subchronic administration of anisomycin.
Reconsolidation of eyelid CRs is sensitive to the protein synthesis inhibitor
It has been reported that electroconvulsive shock or systemic drug administration given after memory reactivation (retrieval) can cause amnesia for the original learning (for review, see Sara 2000; Nader et al., 2000b; Nader, 2003). This fact indicates that consolidated memories become labile when they are retrieved, thus requiring reconsolidation.
Therefore, it was necessary to determine whether reconsolidation-related protein synthesis was required for retrieved memories to persist in acquired eyelid CRs. To do so, mice were given eight conditioning sessions. Ten days after conditioning, the animals received an additional conditioning session (reactivation) to evaluate the retention index. Afterward, they were injected with either saline or anisomycin (100 mg/kg) 30 min before or 30 min after the beginning of the reactivation session. Because the two groups showed similar results, only results from mice injected 30 min after the beginning of reactivation are represented (Fig. 6). To demonstrate a specific effect of anisomycin on memory reconsolidation, it was necessary to demonstrate an intact performance during a postreactivation STM test (PR-STM), made 6 h after the reactivation session, and an impaired performance in the same subjects during a postreactivation LTM test (PR-LTM), made 2 d after retrieval (Fig. 6A). During reactivation, subjects of the two groups exhibited a comparable percentage of eyelid CRs (74.26 ± 4.28% for the saline-injected group and 75.88 ± 3.15% for the anisomycin-injected group) (Fig. 6B). PR-STM performance was similar in the two groups (F(1,10) = 0.113; p = 0.74) and not statistically different from the performance during the reactivation session of the saline-injected group (F(1,6) = 0.163; p = 0.7) and of the anisomycin-injected group (F(1,14) = 0.07; p = 0.77). In contrast, PR-LTM performance was significantly lower in the anisomycin-injected group than in the saline-injected group (F(1,10) = 10.45; p = 0.009).

Anisomycin administration before (-30 min) or after (+30 min) reactivation led to impaired reconsolidation. A, Diagram illustrating the experimental design. For phase 1, eight training sessions were performed. For phase 2 (i.e., 10 d after phase 1), a single training session [reactivation (RA)] was performed. Either anisomycin (experimental subjects) or saline (control subjects) was injected at -30 or +30 min for the reactivation session (n = 8 per group). Because results were similar in the two anisomycin-treated groups, only data corresponding to the +30 min group are shown. Phase 3 was performed to evaluate reconsolidation. For this, animals underwent training sessions 6 h (to test STM) and 48 h (to test LTM) after the end of the reactivation session. During each session, 60 CS-US trials were presented. B, Percentage of responses (mean ± SEM) during habituation, last conditioning, reactivation, STM, and LTM sessions. Open circles, Group injected with anisomycin at +30 min; filled circles, consolidated saline groups. C, Diagram illustrating an additional experimental design similar to that described in A. In this case, anisomycin or saline was injected at +4 h, and only LTM was tested (n = 8 per group). D, Percentage of responses (mean ± SEM) during habituation, last conditioning, reactivation, and LTM sessions. Open squares, Group injected with anisomycin at +4 h; filled squares, saline group. Cond, Conditioning; Aniso, anisomycin; H, habituation; C8, eighth conditioning session; Sal, saline; **p ≤ 0.01.
Protein synthesis inhibitors typically impair the consolidation of new memories when they are administered during a specific time window (which varies from minutes to hours) after learning. Administration of such drugs after this time window does not affect memory. With this in mind, and to explore the possible existence of such a time window for reconsolidation, anisomycin injection was delayed 4 h after retrieval (Fig. 6C). Anisomycin injection 4 h after retrieval had no effect on memory storage (F(1,6) = 0.04; p = 0.84) (Fig. 6D). Thus, both consolidation and reconsolidation have similar time windows within which protein synthesis is required if a memory is to persist.
Extinction and consolidation of extinction of eyelid CRs have protein synthesis requirements similar to those for CR acquisition
Extinction is the decline of a CR resulting from its repetitive performance in the absence of the US or reinforcer (Pavlov, 1927). Extensive behavioral evidence indicates that extinction reflects a process of relearning rather than one of unlearning or forgetting. Through this relearning process, the new association of the conditioned stimulus with the absence of the original reinforcer comes to control the behavior (Rescorla, 1996). If extinction is, in fact, learning rather than forgetting, will the protein synthesis requirements that subserve learning and extinction be identical? This question was addressed by injecting anisomycin at three different times in four extinction sessions. Injections were performed out 30 min before and 30 min and 4 h after the beginning of each extinction session (Fig. 7A). As during acquisition, extinction was abolished only in the group of mice injected before each extinction session (Fig. 7B). In the groups that received anisomycin injections after each extinction session, the decline in performance was similar to that in the saline-injected group. In all of the extinguished groups, consolidation of this process was evaluated 10 d after the end of extinction. As was found in the consolidation of eyelid CRs, saline- and +4 h anisomycin-injected groups showed a percentage of CRs similar to that displayed during the last extinction session (Fig. 7B). In both cases, the retention index was close to unity (Fig. 7C), confirming that extinction consolidation had occurred. In contrast, although the animals injected with anisomycin 30 min after the beginning of each extinction session learned the new task, they were not able to consolidate it, as shown by an increase in the percentage of eyelid CRs in the retrieval session with respect to the fourth extinction session (74.3 ± 1.58 vs 38.78 ± 2.87%) and by an increase in the retention index (Fig. 7B,C). Together, these results suggest that acquisition and extinction of eyelid CRs and their consolidation have common temporal courses in their protein synthesis requirements.

Acquisition and consolidation of extinction require protein synthesis. A, Diagram illustrating the experimental design. Phase 1 was performed as illustrated in Figure 5A. For phase 2 (i.e., extinction), four CS-alone sessions were performed. During each extinction session, 60 CS-alone trials were presented. Either 100 mg/kg anisomycin (experimental subjects) or saline (control subjects) was injected at variable times from the beginning of each session: -30 and +30 min and +4 h (n = 8 per group). Because all saline groups were indistinguishable, their results were grouped for the sake of simplicity. For phase 3, the above-described groups received an additional CS-alone session [i.e., reactivation (RA)] 10 d after the end of phase 2. B, Percentage of responses (mean ± SEM) during habituation, eight conditioning, extinction, and reactivation sessions. Filled circles, Saline-injected animals; open circles, triangles, squares, mice injected with anisomycin at -30 and +30 min and +4 h, respectively. C, Retention index represents the ratio between the first training session in phase 3 and the last training session in phase 2. Cond, Conditioning; Aniso, anisomycin; H, habituation; C8, eighth conditioning session; Sal, saline; E1-E4, four extinction sessions; *p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001.
Discussion
Early in the study of memory processes, it was demonstrated for different species that transcription and translation are required for LTM formation (Abel and Kandel, 1998; Milner et al., 1998). The time-dependent molecular processes underlying LTM take place at vastly different scales and include biochemical changes occurring within seconds to hours and interactions of neuronal ensembles throughout the brain taking milliseconds to years (Dudai, 1997, 2004; McGaugh, 2000). Here, we focused on the role of protein synthesis in the acquisition, consolidation, reconsolidation, and extinction of eyelid CRs in mice assessed by the administration of anisomycin. At low concentrations, anisomycin is a potent MAP kinase activator (Kyriakis et al., 1994), whereas at high doses, it acts as a protein synthesis inhibitor (Rosenblum et al., 1993). Because both events are ubiquitous, anisomycin can elicit anxiety-like states or alterations in exploratory behavior (Igaz et al., 2002). Nevertheless, at 100 mg/kg, anisomycin abolishes (>95% inhibition) protein synthesis in the brain starting 15 min after and lasting up to 6 h without causing any evident neuropathological change and/or any modification in locomotion or blink reflex parameters. Therefore, the effects of anisomycin on learning and memory processes described here were not the result of unspecific alterations of the skeletal and/or eyelid motor systems.
The acquisition of eyelid CRs was impaired when anisomycin was administered before training sessions but not when administered after their end. These data contrast with those obtained by Bourtchouladze et al. (1998) and Igaz et al. (2002), who reported that the administration of a protein synthesis inhibitor immediately after the training session impaired STM and LTM in singletrial learning paradigms. Together, however, these results indicate that protein expression during conditioning sessions is crucial for the acquisition of CRs. Administration of anisomycin in the rabbit cerebellum produces a similar acquisition deficit; however, in this case, the percentage of CRs increases marginally along conditioning sessions (Bracha et al., 1998). This might indicate that cerebral structures other than the cerebellum are involved in the acquisition of CRs or, alternatively, that anisomycin does not affect cerebellar areas relevant to learning. Furthermore, we found that protein synthesis inhibition, in a temporal window well after (4 h) the beginning of conditioning sessions, has no effect on the acquisition of eyelid CRs.
Consolidation in anisomycin-treated animals that have acquired eyelid CRs was tested using an additional session 10 d after the last conditioning session. In this case, animals injected +4 h for each training session remembered the learned task, whereas those injected with anisomycin just after (+30 min) each training session did not. This result suggests that the molecules implicated in the long storage of information have already been synthesized 4 h after the end of each conditioning session. These results are not in agreement with those from passive avoidance training and contextual fear conditioning (Freeman et al., 1995; Bourtchouladze et al., 1998; Igaz et al., 2002). In the latter learning tasks, two protein synthesis temporal windows have been described: immediately after and 3-6 h after the end of each training session. This disagreement may be attributable to the different learning paradigms used in each experiment: one-step learning in contextual fear conditioning versus several-step learning in classical conditioning of eyelid responses. In several-step learning protocols, short-term or synaptic consolidation, which spans from seconds to 24 h (in the period between two consecutive sessions), can be separated from long-term or system consolidation, spanning from days to years (Dudai, 2004). Cooke et al. (2004) and Attwell et al. (2002) have shown that muscimol infusion into the cerebellum just after each training session abrogated the acquisition of eyelid CRs. These results are in apparent contradiction with those reported here. In our view, both the experimental manipulation of Cooke et al. (2004) and Attwell et al. (2002) and anisomycin injection at -30 min for each training session disrupted the synaptic consolidation taking place between training sessions. In contrast, anisomycin administration at +30 min for each training session affected system consolidation. Thus, although synaptic consolidation would require effector proteins encoded by genes expressed during training and the correct activation of neurotransmitter systems, system consolidation would require proteins encoded by genes expressed after each training session. It is not established whether synaptic and systemic consolidation happen simultaneously or whether they represent consecutive process (first synaptic and then system consolidation). Thus, it cannot be discarded that anisomycin administration only after the last (sixth to eighth) training sessions would be enough to disturb system consolidation.
The similarities found by us in protein synthesis requirements for acquisition and extinction of eyelid CRs and for their consolidation suggest that these two processes share common molecular mechanisms. Our results coincide with those obtained with taste aversion conditioning (Bahar et al., 2003). However, although the molecular mechanisms for acquisition and extinction in taste aversion conditioning are akin, the structures involved in both processes are different. It has been reported recently that, although the consolidation of acquisition and extinction of fear conditioning require de novo protein synthesis, the signal transduction pathways involved in the two processes overlap only partly (Lin et al., 2003; Suzuki et al., 2004).
There is some controversy regarding the characteristics, and even the existence, of the reconsolidation process. Thus, reconsolidation impairment of contextual fear conditioning after local or systemic anisomycin administration has been described in rodents (Nader, 2003), but other groups did not get similar results (Lattal and Abel, 2001; Taubenfeld et al., 2001; Tronel and Sara, 2002). This inconsistency can be ascribed to differences in the conditioning paradigm, the retrieval protocol, or the consolidation time. A recent report found that the probability of disturbing reconsolidation decreases after 14 d of memory consolidation (Milekic and Alberini, 2002). Reconsolidation may not be a global phenomenon, as gathered from the use of taste aversion conditioning (Berman and Dudai, 2001) and passive avoidance (Vianna et al., 2001) procedures. Our results imply the existence of reconsolidation after retrieval of eyelid CRs. Furthermore, here we have shown similar, although not identical, de novo protein synthesis requirements for reconsolidation and consolidation, as has been found in other studies of the reconsolidation process (Dudai, 2004). Recently, Lee et al. (2004) reported a double dissociation in the molecular processes in the hippocampus that are necessary for the consolidation and reconsolidation of long-term contextual fear conditioning. Thus, BDNF is required for initial consolidation but not for reconsolidation, whereas Zif-268 is necessary for reconsolidation but not for consolidation of hippocampus-dependent fear memory. Furthermore, reconsolidation is established more quickly than consolidation. These data suggest that reconsolidation is a partial recapitulation of the consolidation process, and reconsolidation requires the expression of both early and delayed effector genes. The latter proposal explains the reconsolidation sensitivity to anisomycin reported here. Consolidation and reconsolidation could happen in different structures: synaptic consolidation in cerebellum (Attwell et al., 2002; Cooke et al., 2004) and hippocampus and system consolidation in prefrontal medial cortex (Takehara et al., 2002, 2003; Dudai, 2004). Thus, the retrieval of a learned task might require its recovery from neocortex as well as from hippocampus and cerebellum. Moreover, these structures could have different sensitivity to anisomycin.
Using protein synthesis inhibitors, it has been shown that protein synthesis is required in critical time windows for the formation of LTM (Davis and Squire, 1984; Dudai, 2004). The transcription and translation of specific genes are dynamically regulated by neural activity, and this regulation involves a number of signal transduction pathways (Shaywitz and Greenberg, 1999; Sweatt, 2001). Synaptic activity driven by experience leads to alterations in intracellular messenger (cAMP and calcium) levels, which in turn activate protein kinases and phosphatases (Abel and Lattal, 2001; Guzowsky, 2001). These enzymes modulate the activity of a wide range of preexisting cellular proteins, including synaptic components involved in STM, and nuclear-localized transcription factors (mainly cAMP response element-binding protein). In the nucleus, activation of these transcription factors initiates a cascade of gene expression required for the formation of LTM. The immediate-early genes (IEGs) form the first group of genes expressed after synaptic activation. IEGs encode diverse proteins, including regulatory transcription factors, structural and signal transduction proteins, growth factors, and enzymes. Synaptic activity also induces the expression of late effector proteins, which in turn require the expression of IEG transcription factors. Both late effector and IEG-encoded proteins are involved in long-term plasticity that leads to LTM (Fig. 8).

Proposed model of experience-dependent gene expression in synaptic plasticity and memory consolidation processes. Synaptic activity driven by experience leads to alterations in intracellular second messenger levels, which in turn activate cellular kinases and phosphatases. These enzymes modulate the activity of a wide range of preexisting cellular proteins, including synaptic components and nuclear transcription factors. In the nucleus, the activation of transcription factors initiates a cascade of gene expression required for the formation of the long-term memory.
In conclusion, de novo protein synthesis is needed for all processes related to learning and memory. Two waves of protein synthesis appear to be involved in the establishment of LTM: a first wave consisting of IEG-encoded effector proteins that, together with changes in neurotransmission, would be required for the acquisition, extinction, and reconsolidation of eyelid CRs and a second wave in which late effector proteins are involved in (general) consolidation processes. Nevertheless, a role for IEG-encoded effector proteins in consolidation cannot be ruled out. This interpretation coincides with recent reports describing two temporal windows of protein synthesis (Freeman et al., 1995; Quevedo et al., 1999; Igaz et al., 2002). From our experimental design, however, we cannot rule out a single wave of protein synthesis responsible for acquisition and consolidation of eyelid CRs. The determination of the cerebral areas involved in classical conditioning of eyelid responses and their precise temporal activation will help identify the genes that encode the proteins participating in learning and memory processes.
Footnotes
This work was supported by the Agencia Española de Cooperación Iberoaméricana, Junta de Andalucía Grant CVI-122, and Spanish Ministry of Education and Science Grant BFI2002-00936. We thank Drs. A. Múnera, L. Jiménez-Díaz, and M. Atienza for critical reading of this manuscript, M. C. Sutil, M. D. Marín, and M. Cadena for technical assistance with animal housing, and R. Churchill for editorial help.
Correspondence should be addressed to Dr. Ángel M. Carrión, División de Neurociencias, Universidad Pablo de Olavide, Carretera de Utrera, Km. 1, 41013 Seville, Spain. E-mail: amancar{at}dex.upo.es.
M. C. Inda's present address: Instituto de Neurobiología Ramón y Cajal, Avenida Dr. Arce 37, 28002 Madrid, Spain.
Copyright © 2005 Society for Neuroscience 0270-6474/05/252070-11$15.00/0





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}