

Abstract
Activation of several signaling pathways contributes to long-term synaptic plasticity, but how brief stimuli produce coordinated activation of these pathways is not understood. In Aplysia, the long-term facilitation (LTF) of sensory neuron synapses by 5-hydroxytryptamine (serotonin; 5-HT) requires the activation of several kinases, including mitogen-activated protein kinase (MAPK). The 5-HT-enhanced secretion of the sensory neuron-specific neuropeptide sensorin mediates the activation of MAPK. We find that stimulus-induced activation of two signaling pathways, phosphoinositide 3-kinase (PI3K) and type II protein kinase A (PKA), regulate sensorin secretion and responses. Treatment with 5-HT produces a rapid increase in sensorin synthesis, especially at varicosities, which precedes the secretion of sensorin. PI3K inhibitor and rapamycin block LTF and the rapid synthesis of sensorin at varicosities even in the absence of sensory neuron cell bodies. Secretion of the newly synthesized sensorin from the varicosities and activation of the autocrine responses of sensorin to produce LTF require type II PKA interaction with AKAPs (A-kinase anchoring proteins). Thus, long-term synaptic plasticity is produced when multiple signaling pathways that are important for regulating distinct cellular functions are activated in a specific sequence and recruit the secretion of a neuropeptide to activate additional critical pathways.
Introduction
Activation of several signaling pathways contributes to initiating and maintaining long-term synaptic plasticity. Some forms of long-term potentiation (LTP) are initiated by calcium entry to activate calcium/calmodulin kinase II (CaMKII) (Lynch et al., 1983; Bliss and Collingridge, 1993; Miller et al., 2002). Other signaling pathways also are required: protein kinase A (PKA) (Huang et al., 2000; Schafe and LeDoux, 2000; Nguyen and Woo, 2003), protein kinase C (PKC) (Lovinger and Routtenberg, 1988; Malinow et al., 1988; Sacktor et al., 1993; Ling et al., 2002), mitogen-activated protein kinase (MAPK) (Sweatt, 2004), phosphoinositide 3-kinase (PI3K) (Lin et al., 2001; Man et al., 2003; Opazo et al., 2003), and phospholipase C (Minichiello et al., 2002).
How does the relatively brief stimulus that initiates long-lasting plasticity activate many signaling pathways? Calcium entry or release from internal stores can activate a number of kinases and phosphatases (Frey et al., 1993; Lisman et al., 2002; Malenka and Bear, 2004). Cross talk between signaling pathways also may recruit or inhibit specific signaling pathways (Impey et al., 1998; Roberson et al., 1999; Yamamoto et al., 1999; Waltereit and Weller, 2003). Certain stimuli also lead to the enhanced secretion of neurotrophins, which contribute to some long-lasting forms of hippocampal LTP after binding Trk (tyrosine receptor kinase) receptors, leading to the activation of MAPK and other kinases (Kang et al., 1997; Patterson et al., 2001; Balkowiec and Katz, 2002; Gartner and Staiger, 2002; Minichiello et al., 2002). How is the temporal sequence of kinase activation coordinated, and is that coordinated sequence critical for long-term plasticity?
In Aplysia, several signaling pathways contribute to long-term facilitation (LTF) at sensory neuron synapses produced by 5-hydroxytryptamine (serotonin; 5-HT) or sensitizing stimuli: type I and type II PKA (Greenberg et al., 1987; Liu et al., 2004), p42/44 MAPK (Martin et al.; 1997; Purcell et al., 2003; Hu et al., 2004a), and PI3K (Udo et al., 2005). Some kinases phosphorylate transcription factors and cytoplasmic substrates to regulate macromolecular synthesis (Bartsch et al., 1995, 1998; Chain et al., 1999; Yamamoto et al., 1999; Giustetto et al., 2003; Liu and Schwartz, 2003). Other kinases phosphorylate local substrates that may be critical for regulating synaptic transmission, local protein synthesis, and synaptic growth (Bailey et al., 1992, 1997; Casadio et al., 1999; Angers et al., 2002; Si et al., 2003; Liu et al., 2004; Grabham et al., 2005; Udo et al., 2005). Type II PKA, which is concentrated at synaptic sites, is activated by 5-HT, and its increased expression and its interaction with anchoring proteins that maintain its proximity to substrates near the membrane are required for LTF (Liu et al., 2004). The activation/translocation of p42/44 MAPK in sensory neurons by 5-HT is mediated by the secretion of the sensory neuron-specific neuropeptide sensorin and activation of autoreceptors (Hu et al., 2004a). How does 5-HT affect sensorin secretion and the responses required for LTF?
We report here that the sequential activation of PI3K and type II PKA mediates enhanced sensorin secretion and its activation/translocation of MAPK in sensory neurons. Activation of the PI3K pathway enhances sensorin synthesis, especially at varicosities, whereas activation of type II PKA increases secretion of the newly synthesized sensorin from varicosities and enhances the responses of sensorin that lead to LTF. Thus, the sequential activation of two kinases with relatively brief stimuli not only regulates specific functions but also regulates the timely secretion of a neuropeptide that activates additional kinases required for long-term plasticity.
Materials and Methods
Cell culture and electrophysiology. Sensory neurons were isolated from pleural ganglia dissected from adult animals (60–80 g), and L7s were isolated from juvenile abdominal ganglia (2 g) and maintained in coculture for 5 d (Montarolo et al., 1986). Standard electrophysiological techniques were used to record EPSP amplitudes evoked in L7 (Hu et al., 2004). L7s were held at –85 mV. EPSP amplitudes were recorded before and 24 h after various treatments. Changes in EPSP amplitudes were measured by dividing the post-treatment EPSP amplitude by the pretreatment EPSP amplitude times 100%. No change in amplitude is represented as 100%. LTF was produced by 5× 5-HT (5 min each at a final concentration of 5 μm) applied at 20 min intervals. Cultures were rinsed after each application with 10 ml of a 1:1 mixture of L15 medium and seawater. The entire protocol for producing LTF lasted ∼90 min. LTF also was produced as described previously (Hu et al., 2004a) with a single application of 5-HT (5 min), a rapid rinse with L15–sea water medium (10 ml in 90 s), followed immediately by a 2 h incubation with sensorin B neuropeptide (100 ng/ml).
Drug treatments. Cultures were incubated for 2 h with anti-sensorin antibody (Ab) or with protein A-purified preimmune serum (400 ng/ml) (Hu et al., 2004a), starting either immediately after the last application of 5-HT or 3 h later. Protein synthesis inhibitor rapamycin (50 nm; Sigma, St. Louis, MO), PI3K inhibitor 2-(4-morpholinyl)-8-phenyl-4H-1-benzopyran-4-one (LY294002; 10 μm; Calbiochem, La Jolla, CA), or PKA inhibitor (9R,10S,12S)-2,3,9,10,11,12-hexahydro-10-hydroxy-9-methyl-1-oxo-9,12-epoxy-1H-diindolo[1,2,3-fg-3′,2′,1′-K1]pyrrolo [3,4-1][1,6]benzodiazocine-10-carboxylic acid (KT5720; 10 μm; Calbiochem) was added to cultures 15 min before 5×5-HT until immediately after the last application. In some cultures, LY294002 was applied for 2 h immediately after the last application of 5-HT. The permeable peptide inhibitor of type II PKA interaction with A-kinase anchoring proteins (AKAPs) S-Ht31 (10 μm; Promega, Madison, WI) or the mutated control peptide S-Ht31P (10 μm; Promega) was added for 2 h immediately after the last application of 5-HT. For experiments examining the distribution of phosphorylated MAPK, the inhibitor or control peptide was added for only 1 h immediately after 5-HT.
Immunocytochemistry. Immunocytochemistry was used to monitor expression or changes in the distribution of sensorin or phosphorylated p42/44 MAPK (Hu et al., 2004a). Cultures at various times after 5-HT or after the application of control solutions were rinsed briefly in artificial seawater and fixed in 4% paraformaldehyde and processed as described previously (Liu et al., 2003; Hu et al., 2004a). Cells were exposed to rabbit polyclonal antibody specific for sensorin (1:1000) or phospho-p42/44 MAPK (1:200; Cell Signaling Technology, Beverly, MA) diluted in 2% normal goat serum in 0.01 m PBS with 0.3% Triton X-100 at 4°C for 24 h. The incubated cultures were washed in 0.01 m PBS and incubated in FITC-conjugated goat anti-rabbit IgG (1:200; Sigma) at 4°C for 4 h. After being washed in 0.01 m PBS, the cultures were imaged directly with a filter set used for detecting fluorescent signal. The cultures were viewed with a Nikon (Tokyo, Japan) Diaphot microscope attached to a silicon-intensified target (SIT; Dage 68; Dage-MTI, Michigan City, MI) video camera, the images were processed by a Dell computer, and the images were captured and processed by the Microcomputer-Controlled Imaging Device (MCID) software package (Imaging Research, St. Catharines, Ontario, Canada). Illumination for detecting fluorescent signals was maintained at a constant setting for all experiments.
In situ hybridization. The specific antisense oligonucleotide probe for sensorin was designed for the specific coding portions of the target sequence used successfully in reverse transcription-PCR analyses of the mRNAs (Schacher et al., 1999) [for probe sequence, see Hu et al. (2002, 2003, 2004b)], synthesized (Genset, La Jolla, CA), lyophilized, and dissolved in sterile distilled water. The sense probe also was designed for use as nonspecific controls. These probes were labeled at the 3′ end with biotin-ddUTP according to the instructions of the manufacturer (Roche Diagnostics, Indianapolis, IN). In situ hybridization was performed as described previously (Hu et al., 2002, 2003). Fixed cultures were hybridized overnight at 42°C in hybridization buffer containing 1.5 μg/ml of the biotin-labeled oligonucleotide probes. Unbound probe was washed out. Cultures then were incubated in streptavidin–FITC (1:200; Invitrogen, Gaithersburg, MD) for 4 h at 4°C. After unbound streptavidin–FITC was washed out, the hybridization signals were visualized directly with fluorescent microscopy with the imaging system described above. The specificity of biotin-labeled antisense oligonucleotide probe was examined by hybridizing with labeled sense probe or excess unlabeled probe or by omitting probe in hybridization solution as well as RNase pretreatment. All controls were negative. Cells were hybridized under the same conditions to reduce variability.
Quantification and data analysis. All data are expressed as the mean change (percentage) ± SEM produced by the indicated treatments. The intensity of sensorin immunostaining was tested by measuring average fluorescent intensity in the sensory neuron cell body, the entire main axon, and varicosities contacting the major processes of L7 with the MCID (7.0) software package from Imaging Research. The hybridization signal of sensorin mRNA was measured by averaging fluorescent intensity over the entire sensory cell body. Staining intensity for sensorin mRNA or protein for the various experimental treatments was compared with control treatments that were normalized to 100%. The overall staining intensity of phosphorylated MAPK immunofluorescence was determined by averaging intensity for the cell body (cytoplasm plus nucleus). Nuclear staining was determined by measuring average intensity over that area. ANOVA and Scheffé's F test were used to gauge significant differences between treatments.
Results
5-HT induces the rapid increase in sensorin protein expression that is critical for LTF
New macromolecular synthesis is required for LTF (Montarolo et al., 1986). Because sensorin secretion is enhanced significantly immediately after five applications of 5-HT (5×5-HT) (Hu et al., 2004a), we tested whether 5× 5-HT produces a rapid change in the expression of sensorin protein and mRNA that contributes to the sensorin that is secreted after 5-HT.
Expression of sensorin throughout the sensory neuron was enhanced significantly immediately after the last application of 5-HT (0 h) (Fig. 1A,B). Increased staining in distal processes was concentrated primarily in punctate granules along the axon or in varicosities. Compared with controls (n = 5 cultures; staining intensity in each compartment normalized to 100%), 5-HT increased sensorin staining at 0 h (n = 7 cultures) in the cell body by nearly twofold (195 ± 8%), in the axon by nearly fivefold (483 ± 25%), and in the distal varicosities by more than threefold (307 ± 9%). Then, 2 h later (n = 6 cultures) sensorin expression in axons and varicosities returned to or below baseline. Staining in the axons was only 112 ± 10%, and staining in varicosities was reduced below control levels to 64 ± 5%. This rapid decrease in staining was consistent with previous results [Hu et al. (2004a), their Fig. 2] and indicated that newly expressed sensorin was secreted from distal processes during the 2 h period after 5-HT.

5-HT produces a rapid increase in sensorin, the release of which is critical for LTF. A, Nomarski contrast (top) and sensorin immunofluorescent (bottom) images of cocultures after mock applications (Cont) or at various times after 5-HT (0 h is immediately after the last 5-HT application). Staining in the sensory neuron axon is confined to large granules after treatment and in the distal processes to varicosities. Scale bar, 50μm. B, Summary of sensorin immunostaining in sensory neurons. Staining for each compartment at 0,2, and 12 h after 5-HT was normalized to the average staining for that compartment in control (dashed line at 100%). ANOVA indicated a significant effect of treatment (df = 6, 38; F = 48.276; p<0.001). At each time point (0, 2, and 12 h) there was an approximately twofold increase in staining in the cell body (F = 19.538 and p<0.01; F = 11.077 and p<0.01; F = 11.169 and p<0.01, respectively). Staining in the axon and at distal varicosities increased significantly at 0 h (F = 81.211 and p<0.01; F = 92.067 and p<0.01, respectively) and then reversed to control levels or below by 2 h (both were not significantly different from control). In contrast, staining in axon and varicosities at 12 h was greater than control (F = 18.054 and p<0.01; F = 8.908 and p<0.01, respectively). C, Sensorin released between 0 and 2 h is critical for LTF. EPSPs were recorded before and 24 h after 5-HT. Cultures were incubated with anti-sensorin Ab (SEN Ab) for 2 h either immediately after 5-HT (0–2) or 3 h after 5-HT (3–5). ANOVA indicated a significant effect of treatment (df = 2, 30; F = 17.208; p<0.001). Anti-sensorin Ab at 0–2 h blocked LTF but failed to block LTF when applied between 3 and 5 h after 5-HT (F = 19.393 and p<0.01 compared with 5-HT alone; F = 16.57 and p<0.01 when Anti-sensorin Ab was applied between 3 and 5 h). EPSPs that are unchanged (100%) are indicated by the dashed lines (see Figs. 2C, 3C, 5C, and 6).Calibration: vertical, 20 mV; horizontal, 25 ms. D, The first wave of increased sensorin expression was independent of transcription, whereas the second was preceded by an increase in sensorin mRNA. Each figure is a pseudocolor representation of in situ hybridization fluorescent staining for sensorin mRNA (blue is low intensity, and red is high intensity) in sensory neuron cell bodies in control and at 0, 2, and 12 h after 5-HT. ANOVA indicated a significant effect of treatment (df = 3, 16; F = 21.2; p<0.001). After being normalized to staining intensity in control, staining at 0 h did not change but was significantly greater at 2 and 12 h (F = 3.434 and p<0.05; F = 16.228 and p<0.01, respectively). Scale bar, 30 μm.
The secretion of sensorin during this 2 h period was critical for LTF. We found previously that exogenous anti-sensorin Ab binds to and inactivates sensorin (Hu et al., 2004a). The antibody blocked LTF when it was applied after 5-HT for 22 h. Incubation with anti-sensorin Ab for only 2 h (0–2 h after 5-HT) was sufficient to block LTF, whereas incubation with anti-sensorin Ab for 2 h at a later time (3–5 h after 5-HT) did not block LTF (Fig. 1C). Incubation with anti-sensorin Ab between 0 and 2 h (n = 11 cultures) reduced the change in EPSP amplitude produced by 5-HT to 103.6 ± 3.3% compared with an increase of 154.7 ± 6.2% after a 2 h incubation with control rabbit IgG (n = 9). In contrast, EPSP amplitudes increased by 146.6 ± 6.3% (n = 13 cultures) when cultures were incubated with anti-sensorin Ab for 2 h beginning at 3 h after 5-HT. Thus, newly expressed sensorin secreted from the distal processes of sensory neurons immediately after 5-HT is critical for LTF.
The rapid increase in sensorin protein immediately after 5-HT was not correlated with a rapid increase in sensorin mRNA (Fig. 1D). Compared with controls (n = 5 cultures; staining intensity normalized to 100% for controls), hybridization staining for sensorin mRNA immediately after 5-HT (0 h; n = 5 cultures) was not significantly different (103 ± 8%). At 2 h after the treatment with 5-HT (n = 5 cultures), when sensorin was secreted to deplete the protein from axons and varicosities, staining for sensorin mRNA increased to 133 ± 12% above controls. By 12 h after 5-HT, the staining for both sensorin protein (n = 6 cultures) and mRNA (n = 5 cultures) was elevated significantly above controls: nearly twofold greater for protein and mRNA in the cell body and nearly threefold greater for sensorin protein in axons and varicosities (Fig. 1A,B,D). Thus, 5-HT produced two periods of increased sensorin protein synthesis that are likely to be mediated by different mechanisms: a rapid transcription-independent increase that contributed to the sensorin secreted from distal sites immediately after 5-HT and a second phase of increased synthesis known to persist up to 24 h after 5-HT (Santarelli et al., 1996; Liu et al., 2003) that was preceded by an increase in sensorin mRNA.
PI3K and rapamycin regulate the rapid increase in sensorin expression by 5-HT
How does 5-HT regulate the rapid synthesis of sensorin? Several kinase activities are activated by 5-HT. Recent evidence suggests that PI3K activity is required for LTF, and rapamycin, a potent inhibitor of specific translation, blocks LTF produced by 5-HT or sensorin (Hu et al., 2004a; Udo et al., 2005). Because PI3K and rapamycin can regulate protein synthesis at distal sites in neurons (Beaumont et al., 2001; Tang et al., 2002; Cammalleri et al., 2003; Schratt et al., 2004; Takei et al., 2004; Lee et al., 2005), we examined whether PI3K- and rapamycin-sensitive pathways participated in the rapid changes in sensorin expression immediately after 5-HT applications.
The PI3K inhibitor LY294002 (10 μm) (Fig. 2) or rapamycin (50 nm) (Fig. 3) blocked the rapid increase in sensorin immediately after treatment with 5-HT (0 h). When compared with controls (n = 6 cultures; staining in each compartment normalized to 100%), the PI3K inhibitor (n = 9 cultures) blocked the increase in sensorin staining in the cell body (99 ± 7%), in the axon (99 ± 6%), and in varicosities (98 ± 7%). Treatment with only 5-HT (n = 7 cultures) resulted in a significant increase in sensorin staining in the cell body (186 ± 14%), in the axon (482 ± 20%), and in the varicosities (287 ± 11%). Treatment with the PI3K inhibitor alone failed to alter sensorin staining in all compartments (Fig. 2 A, B). In contrast, blocking PKA activity during 5-HT with the PKA inhibitor KT5720 had only a modest effect on the rapid increase in sensorin protein (images not shown). Compared with controls (normalized to 100% for each compartment; n = 6 cultures), applications of 5-HT in the presence of KT5720 (n = 8 cultures) increased staining for sensorin in the cell body (144 ± 6%), axon (343 ± 13%), and varicosities (240 ± 9.0%). These changes in sensorin produced by 5-HT in the presence of KT5720 were comparable with the increases produced by 5-HT alone (n = 7 cultures) in the cell body (163 ± 9%), axon (492 ± 28%), and varicosities (269 ± 10%). Although interfering with PKA during this period affects other processes necessary for LTF, it appears that PI3K activity selectively mediates the rapid increases in sensorin with 5-HT.
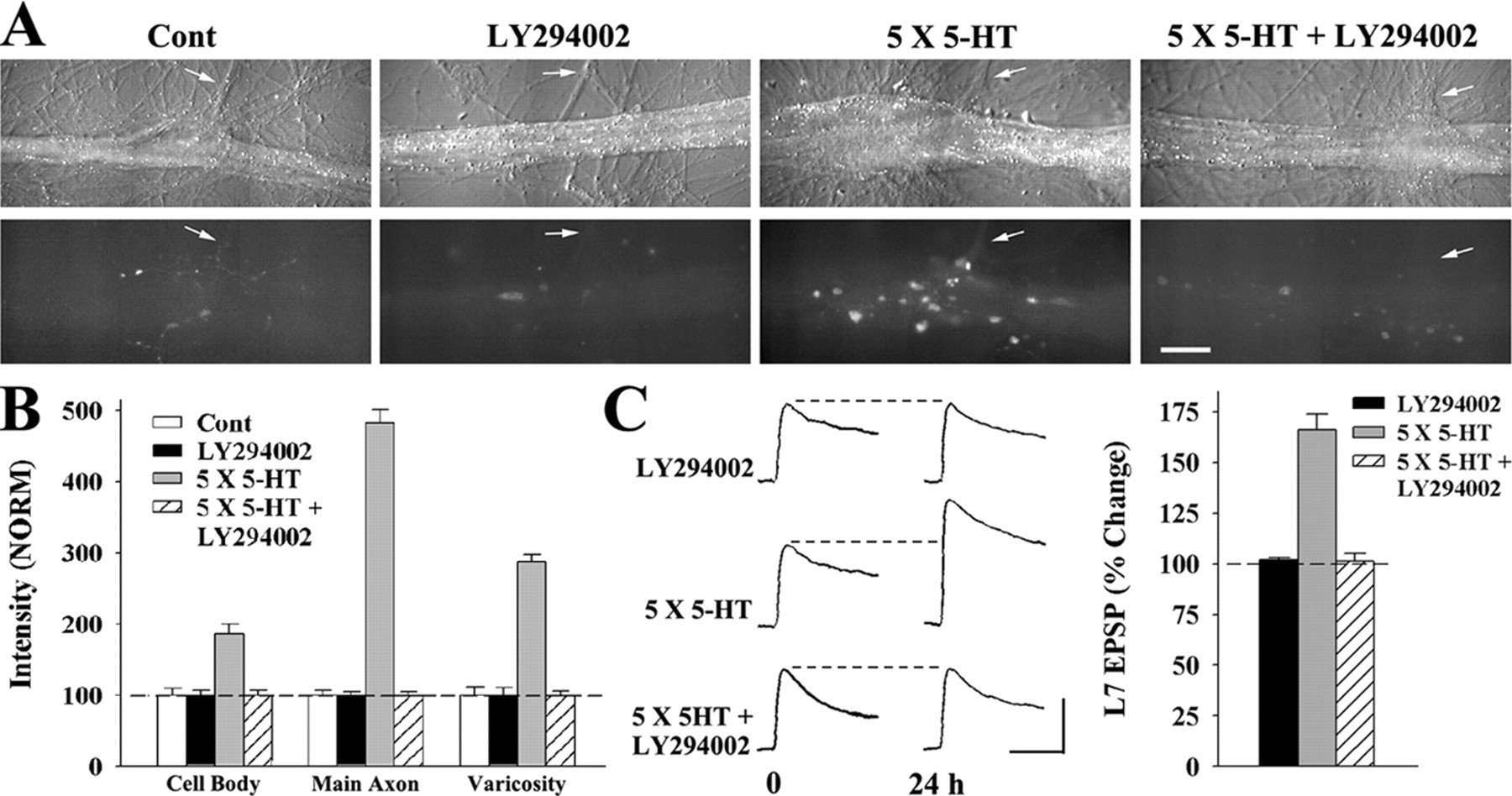
PI3K activity is required for the rapid increase in sensorin. A, Nomarski contrast (top) and sensorin immunofluorescent (bottom) images of cocultures immediately after (0 h) 5-HT in the presence or absence of LY294002 mock applications in the presence or absence of LY294002. Staining in the axons (arrows) and varicosities was reduced significantly when 5-HT was applied in the presence of the drug. Scale bar, 50 μm. B, Summary of sensorin immunostaining immediately after (0 h) the respective treatments. Staining for each compartment after drug or 5-HT treatments was normalized to the average staining for that compartment in control (Cont; dashed line at 100%). ANOVA indicated a significant effect of treatment (df = 6, 50; F = 85.383; p<0.001). Staining in cell body, axon, and varicosities increased significantly with 5-HT alone (F = 11.598 and p<0.01; F = 191.68 and p<0.01; F = 51.743 and p<0.01, respectively). In the presence of LY294002, 5-HT failed to increase staining in all compartments. Treatment with LY294002 alone had little effect on sensorin staining. C, LY294002 blocked LTF produced by 5-HT. EPSPs were recorded before and 24 h after the respective treatments. ANOVA indicated a significant effect of treatment (df = 2, 23; F = 59.819; p<0.001). LY294002 blocked the increase in EPSP produced by 5-HT (F = 46.836; p<0.01). LY294002 did not affect baseline synaptic transmission. Calibration: vertical, 20 mV; horizontal, 25 ms.

Rapamycin blocks the rapid increase in sensorin produced by 5-HT. A, Nomarski contrast (top) and sensorin immunofluorescent (bottom) images of cocultures immediately after (0 h) mock applications in the presence or absence of rapamycin (Rapa) or 5×5-HT in the presence or absence of Rapa. Staining in the axons (arrows) and varicosities was reduced significantly when 5-HT was applied in the presence of the drug. Scale bar, 50μm. B, Summary of sensorin immunostaining immediately after (0 h) the respective treatments. Staining for each compartment after drug or 5-HT treatments was normalized to the average staining for that compartment in control (Cont; dashed line at 100%). ANOVA indicated a significant effect of treatment (df = 6, 48; F = 55.004; p<0.001). Staining in cell body, axon, and varicosities increased significantly with 5-HT (F = 11.037 and p<0.01;F=161.508 and p<0.01;F=102.023andp<0.01,respectively). In the presence of Rapa, 5-HT failed to increase staining in all compartments. Treatment with rapamycin alone had little effect on sensorin staining. C, Rapamycin blocked LTF produced by 5-HT. EPSPs were recorded before and 24 h after the respective treatments. ANOVA indicated a significant effect of treatment (df = 2, 21; F = 13.694; p < 0.001). Rapamycin blocked the increase in EPSP produced by 5-HT (F = 18.419; p < 0.01). Rapamycin alone did not affect baseline synaptic transmission. Calibration: vertical, 20 mV; horizontal, 25 ms.
Incubation with LY294002 also blocked LTF produced by 5-HT (Fig. 2C). The change in EPSP amplitude 24 h after 5-HT was blocked by the PI3K inhibitor (n = 12; 101.4 ± 3.7%) when compared with treatment with 5-HT only (n = 7; 166.0 ± 8.1%). Treatment with LY294002 alone did not alter baseline EPSP amplitude (n = 7; 102.1 ± 1.0%). Thus, PI3K activity during this early phase is required for both the increase in sensorin synthesis and LTF.
Rapamycin applied during 5-HT applications, which blocked LTF, also blocked the rapid increase in sensorin synthesis in all parts of the sensory neuron (Fig. 3). Compared with controls (n = 7 cultures; normalized to 100%), rapamycin (n = 7 cultures) blocked the 5-HT-induced increase in sensorin staining in the cell body (99 ± 10%), in the axon (102 ± 7%), and in the varicosities (101 ± 8%). Treatment with 5-HT produced the expected increases in staining in each compartment, and rapamycin produced no significant reduction in staining when compared with controls (Fig. 3 A, B). Rapamycin also blocked LTF produced by 5-HT (Fig. 3C). Rapamycin blocked the change in EPSP amplitude 24 h after 5-HT (n = 10 cultures; 104.9 ± 4.5%) when compared with an increase in EPSP amplitude of 147.4 ± 7.4% after treatment with 5-HT alone (n = 7 cultures). Rapamycin alone (n = 7 cultures) did not alter baseline EPSP amplitude (101.2 ± 1.8%). The rapid changes in sensorin produced by 5-HT also was regulated by rapamycin-sensitive protein synthesis.

Inhibitor of PI3K or rapamycin blocks the 5-HT-induced increase in sensorin at distal sites in the absence of sensory neuron cell body.A,B, In the absence of the cell body (at each arrow), 5-HT produced a rapid increase in sensorin immediately after 5-HT (0 h) that was reversed by 2 h. Shown are Nomarski contrast (top) and sensorin immunofluorescent (bottom) images of cocultures (A) at 0 and 2 h after mock applications (Cont) or 5-HT. Staining in the axons and varicosities increased significantly with 5-HT at 0 h, but not at 2 h after 5-HT. Scale bar, 50μm. Summary of sensorin immunostaining in axon and varicosities after control or 5-HT treatments (B) indicates that 5-HT regulated sensorin levels even in the absence of cell bodies. Staining for each compartment at 0 and 2 h after 5-HT was normalized to the average staining in that compartment for control (dashed line at 100%).ANOVA indicated a significant effect of treatment (df = 2, 20; F = 15.267; p<0.001). At 0 h, there was a significant increase in staining in the axon and varicosities (F = 144.826, p<0.01; F = 40.098, p<0.01, respectively). By 2 h, staining in both axon and varicosities reverted back to control levels. C, D, In the absence of the sensory neuron cell body, both LY294002 and rapamycin blocked the rapid increases in sensorin produced by 5-HT. Shown are Nomarski contrast (top) and sensorin immunofluorescent (bottom) images of cocultures (C) at 0 h after mock applications (Cont) or 5-HT applications in the presence or absence of the drug. Staining in the axons (arrows) and varicosities that increased significantly with 5-HT at 0 h are blocked by either LY294002 or rapamycin. Scale bar, 50 μm. Summary of sensorin immunostaining in axon and varicosities immediately after control or 5-HT treatments (D) indicates that changes produced by 5-HT in the absence of the cell bodies were blocked by the drugs. Staining for each compartment at 0 h after treatments is normalized to the average staining in that compartment for control (dashed line at 100%). ANOVA indicates a significant effect of treatment (df = 3, 22; F = 17.536; p<0.001. At 0 h, there is a significant increase in staining in the axon and varicosities with 5-HT alone (F = 74.414 and p<0.01; F = 28.293 and p<0.01, respectively). Both LY294002 and rapamycin blocked the increases produced by 5-HT in both axon and varicosities so that the staining in each compartment was not different from control and significantly lower than that produced by 5-HT alone (F = 79.17 and 30.715 and p<0.01, respectively, comparing 5-HT with 5-HT plus LY294002; F = 80.138 and 28.561 and p<0.01, respectively, comparing 5-HT with 5-HT plus rapamycin).
PI3K and rapamycin regulate sensorin levels at isolated sensory neuron processes
After the cell body is removed, 5-HT produces a translation-dependent form of LTF at isolated sensory neuron synapses (Liu et al., 2003; Grabham et al., 2005). We examined whether 5-HT also produces a rapid increase in sensorin in isolated axons and varicosities and whether the PI3K activity and rapamycin regulate that increase in expression. At 3 h after removal of the sensory neuron cell bodies the cultures were treated with 5-HT or control applications in the presence or absence of either LY294002 or rapamycin. Cultures were fixed immediately after washout of the last 5-HT application (0 h) or 2 h later (2 h). Then, the cultures were processed for sensorin immunostaining.
In the absence of the sensory neuron cell body, 5-HT produced a significant increase in sensorin staining in the axon and varicosities of sensory neuron at 0 h, which returned to or below baseline at 2 h after the treatment with 5-HT (Fig. 4A,B). Compared with controls (n = 7 cultures; staining in each compartment normalized to 100%), 5-HT produced increases in staining at 0 h (n = 8 cultures) in the axon (380 ± 17%) and in the varicosities (254 ± 18%). Staining in the axon was reduced to control levels (99 ± 6%) and in the varicosities to below control levels (58 ± 3%) at 2 h after 5-HT (n = 8 cultures). The increases in sensorin staining at 0 h in the axon and varicosities in the absence of the sensory neuron cell body were blocked by the PI3K inhibitor LY294002 and rapamycin (Fig. 4C,D). Compared with controls (n = 6 cultures; staining in each compartment normalized to 100%), the PI3K inhibitor (n = 7 cultures) and rapamycin (n = 7 cultures) blocked the increase in sensorin staining produced by 5-HT at 0 h in the axon (102 ± 7 and 100 ± 6%, respectively) and in the varicosities (99 ± 13 and 101 ± 12%, respectively). Sensorin staining was increased by 5-HT (n = 6 cultures) both in the axon (359 ± 23%) and in the varicosities (271 ± 13%). The rapid local translation of sensorin and its release from distal processes are regulated by 5-HT and are likely to contribute to LTF. Both PI3K activity and rapamycin regulate translation of sensorin in all parts of the sensory neuron.
Type II PKA regulates secretion of sensorin and the autocrine response of sensorin
The activation of PKA by 5-HT appears to affect sensory neurons upstream of the release of sensorin and activation of its autocrine responses (Hu et al., 2004a). In Aplysia neurons, type I and type II PKA are concentrated in different cellular compartments. Type I PKA is concentrated in the sensory neuron cell body, and type II PKA is concentrated at synaptic sites at which its interactions with anchoring proteins (AKAPs) are required for producing LTF. Incubation with a permeable form of a peptide (S-Ht31) that interferes with type II PKA–AKAP interaction blocked LTF (Liu et al., 2004). We therefore examined whether this interaction is required for sensorin secretion.
Blocking PKA–AKAP interaction during the first 2 h after the treatment with 5-HT blocked sensorin secretion and LTF (Fig. 5). Incubation with the permeable peptide inhibitor of PKA–AKAP interactions (S-Ht31; 10 μm) for 2 h beginning immediately after the last 5-HT application blocked the reduction in sensorin staining detected 2 h after 5-HT (Fig. 5 A, B). Incubation with the permeable form of the control peptide (10 μm; S-Ht31P, which has a single amino acid substitution, thereby preventing its inhibition of PKA–AKAP interaction) failed to block the decline in sensorin staining at 2 h after 5-HT. Compared with the high level of staining at 0 h after 5-HT (n = 6; staining in each compartment normalized to 100%), incubation with active peptide (n = 8 cultures) blocked the expected decline in sensorin staining in the axon (90 ± 5%) and in the varicosities (87 ± 5%). In contrast, treatment with the control peptide for 2 h after 5-HT did not affect the expected decline in staining in each compartment; staining intensity declined to 21 ± 2% in the axon and to 25 ± 2% in the varicosities. Incubation with the active peptide during the same time period (0–2 h after 5-HT) also attenuated LTF (Fig. 5C). Incubation with S-Ht31 peptide significantly reduced the change in EPSP amplitude 24 h after 5-HT (115.3 ± 3.8%; n = 15 cultures), whereas incubation with the inactive peptide S-Ht31P produced an increase in EPSP amplitude of 160.9 ± 6.9% (n = 10 cultures). Both the decline in sensorin staining, an indicator of sensorin secretion, and LTF required the proper functioning of type II PKA. In contrast, blocking PI3K activity during the 2 h period after 5-HT did not interfere with the decline in sensorin by 2 h. Compared with the high level of sensorin staining at 0 h after 5-HT (normalized to 100% in each compartment; n = 8 cultures), the reduction in sensorin staining was not blocked in the axon (23 ± 2%) or in the varicosities (28 ± 3%) when the PI3K inhibitor LY294002 was present for 2 h after the treatment with 5-HT (n = 9 cultures). These reductions were comparable with those seen in the axons (22 ± 2%) and varicosities (25 ± 2%) 2 h after 5-HT alone (n = 7 cultures). Thus, the activity of type II PKA was critical for the decline in sensorin (secretion) 2 h after PI3K- and rapamycin-sensitive activities produced an increase in sensorin synthesis.

Type II PKA activity is required for the release of sensorin after 5-HT. A, Nomarski contrast (top) and sensorin immunofluorescent (bottom) images of cocultures immediately after 5×5-HT and 2 h after 5×5-HT and incubated for those 2 h with the active peptide inhibitor of type II PKA interaction with AKAPs (S-Ht31) or with the inactive peptide (S-Ht31P). Staining in the axons (arrows) and varicosities was reduced significantly only when cultures were incubated with the inactive peptide. The active peptide inhibitor blocked the decline in sensorin immunostaining. Scale bar, 50μm. B, Summary of sensorin immunostaining in the sensory neuron axon and varicosities after the respective treatments. The active peptide inhibitor S-Ht31 blocked the decline in sensorin staining 2 h after 5-HT. ANOVA indicated a significant effect of treatment (df = 2, 19; F = 16.43; p<0.001). Compared with the high staining produced at 0 h after 5-HT alone (normalized to 100%), staining declined significantly in axons and varicosities (F = 94.766 and F = 65.153, respectively; p<0.01) when the cultures were incubated with the control peptide (S-Ht31P). Incubation with the active peptide (S-Ht31) blocked the decline, and staining was not significantly different from the high level of staining produced immediately after 5-HT. C, Incubation with the active inhibitor between 0 and 2 h after 5-HT blocked LTF. EPSPs were recorded before (0) and 24 h after 5×5-HT. Immediately after 5-HT, the cultures were incubated for 2 h with the control peptide (S-Ht31P) or the active inhibitor peptide (S-Ht31). The active peptide attenuated the increase in EPSP at 24 h. Calibration: vertical, 20 mV; horizontal, 25 ms. ANOVA indicated a significant effect of treatment (df = 1, 23; F = 21.907; p<0.001). The increase in EPSP amplitude with 5-HT and incubation with the inactive peptide were significantly greater than the change produced by 5-HT and incubation with the active inhibitor (F = 39.423; p<0.01).

Type II PKA activity is required for sensorin-induced LTF. A, EPSPs were recorded in L7 before (0) and 24 h after (24) the indicated treatments. Cultures were exposed to 1×5-HT (5 min), followed immediately after rinsing with a 2 h incubation with sensorin. Beginning 60 min before sensorin application the cultures were incubated with either the control peptide (S-Ht31P) or the active peptide (S-Ht31). 5-HT was added during the last 5 min and then rinsed out before sensorin was added. The active peptide blocked LTF, whereas the control peptide did not. Calibration: vertical, 20 mV; horizontal, 25 ms. B, Summary of changes in EPSP amplitudes produced by 1×5-HT plus sensorin in the presence of control or active peptide. ANOVA indicated a significant effect of treatment (df = 1, 17; F = 29.507; p<0.001). LTF was blocked (F = 22.312; p < 0.01) when the active peptide (S-Ht31) was present during the 5 min application of 5-HT.
In addition to its role in sensorin secretion, PKA activity also was required for sensorin to produce LTF. Sensorin itself (2 h incubation) produced LTF only when preceded by a single 5 min application of 5-HT. Blocking PKA activity with KT5720 during the 5-HT application blocked sensorin-induced LTF (Hu et al., 2004a). We examined whether interfering with type II PKA interaction with AKAPs during the 5-HT application also attenuates sensorin-induced LTF.
Incubation with the active peptide (S-Ht31) before the 5 min application of 5-HT blocked sensorin-induced LTF (Fig. 6). The change in EPSP amplitude 24 h later was only 107.2 ± 5.4% (n = 10). In contrast, when incubated with the inactive peptide (S-Ht31P), sensorin induced a change in EPSP amplitude 24 h later of 145.1 ± 5.8% (n = 9 cultures). Incubation with LY294002 during the single application of 5-HT failed to block sensorin-induced LTF (data not shown). Thus, proper functioning of type II PKA is critical for sensorin to produce LTF.

PI3K activity during 5-HT and type II PKA activity immediately after 5-HT are required for activation and translocation of MAPK. A, Activation and translocation of MAPK by 5-HT were blocked when LY294002 was present during 5-HT application or when the active peptide S-Ht31 was present immediately after 5-HT. Compared with 5×5-HT, overall staining in the cell body and staining in the nucleus were lower when applied in the presence of LY294002. Incubation with the control peptide (S-Ht31P) after 5-HT did not interfere with the staining produced by 5-HT, whereas the active peptide (S-Ht31) reduced staining overall and in the nucleus. B, Summary of phosphorylated MAPK-like immunostaining in the whole cell body and nucleus after the respective treatments. ANOVA indicated a significant effect of treatment (df = 3, 43; F = 151.172; p<0.001). Compared with 5×5-HT alone, 5×5-HT plus LY294002 or 5-HT plus S-Ht31 (active peptide) significantly reduced staining in the cell body (F = 17.318 or F = 17.051, respectively; p<0.01) and in the nucleus (F = 45.23 or F = 46.696, respectively; p<0.01). Incubation with the inactive peptide (S-Ht31P) did not affect staining that normally was produced by 5×5-HT alone. Compared with incubation with the inactive peptide, S-Ht31 reduced staining in both the cell body overall and the nucleus (F = 18.596 and F = 59.932, respectively; p<0.01).
Activation of PI3K and type II PKA is required for MAPK activation and translocation
5-HT activates and translocates p42/44 MAPK (Martin et al., 1997; Purcell et al., 2003) that is mediated by secreted sensorin (Hu et al., 2004a). Our data indicate that the sensorin secretion that produces LTF requires first a PI3K-dependent increase in sensorin synthesis, followed by a PKA-dependent regulation of secretion and of the ability of sensorin to produce LTF (Figs. 2, 3, 4, 5, 6). Because sensorin mediates MAPK activation and translocation, we predict that MAPK activation by 5-HT would be blocked when LY294002 is applied during 5-HT applications or when the peptide inhibitor of type II PKA–AKAP interactions (S-Ht31) is applied immediately after the last application of 5-HT. We used immunocytochemistry for detecting the phosphorylated forms of p42/44 MAPK (Murray et al., 1998) 1 h after the last application of 5-HT (Martin et al., 1997; Hu et al., 2004a).
Incubation with LY294002 during the applications of 5-HT or S-Ht31 immediately after the last application of 5-HT blocked MAPK phosphorylation and translocation into sensory neuron nuclei that are produced by 5-HT (Fig. 7). Incubation with the inactive peptide S-Ht31P failed to block MAPK phosphorylation and translocation. After 5-HT, the staining intensity over the entire cell body and in the nucleus was significantly higher (nearly twofold and more than threefold, respectively) when compared with staining in cultures treated with 5-HT in the presence of LY294002 (Fig. 7A,B, top portions). Incubation with the active peptide after 5-HT also reduced MAPK phosphorylation compared with that observed when incubated with the inactive peptide after 5-HT. Staining in the cell body and the nucleus was twofold and nearly fourfold greater when cultures were incubated with inactive peptide S-Ht31P after 5-HT. Thus, the activation and translocation of p42/44 MAPK by 5-HT require the sequential activities of PI3K and type II PKA.
Discussion
Our findings indicate that the secretion of a neuropeptide that is critical for LTF at Aplysia sensory neuron synapses requires the previous actions of two signaling pathways: the PI3K pathway and type II PKA. The rapid increase of sensorin protein synthesis, including local translation at distal sensory neuron varicosities, is regulated by PI3K activity and is blocked by rapamycin. The newly synthesized sensorin that is secreted from distal processes is mediated by activation of type II PKA during the period immediately after the treatment with 5-HT. The secretion of sensorin and type II PKA activates MAPK and produces LTF. Thus, the sequential and interconnected activities of PI3K, PKA, and MAPK are regulated by 5-HT and sensorin to produce LTF.
PI3K and rapamycin regulate the synthesis of sensorin
Sensorin protein synthesis in the sensory neuron is enhanced significantly and rapidly by 5-HT. The rapid increase in sensorin protein is not correlated with an increase in sensorin mRNA. Moreover, a significant increase in protein expression is detected even when 5-HT is applied after the cell bodies of sensory neurons are removed. Thus, 5-HT induces an increase in sensorin protein synthesis throughout the sensory neuron, including the distal varicosities.
In Aplysia sensory neurons, 5-HT leads to the activation of several kinases. Activation of PKA (Kandel and Schwartz, 1982; Greenberg et al., 1987; Schacher et al., 1988, 1993; Wu et al., 1995; Chang et al., 2000; Liu et al., 2004) and PKC (Sacktor and Schwartz, 1990; Sossin and Schwartz, 1992; Sutton and Carew, 2000; Manseau et al., 2001; Sutton et al., 2004) contribute to short-, intermediate-, or long-term facilitation at sensory neuron synapses. However, PKC activity, required for short- and intermediate-term facilitation, does not appear to be involved with sensorin expression or secretion. Blocking PKC activity during this period is not required for LTF produced by 5-HT or sensorin (Wu et al., 1995; Hu et al., 2003, 2004a). PKA activation during this period is required for long-term forms of facilitation (Greenberg et al., 1987; Schacher et al., 1988, 1993; Liu et al., 2004). However, blocking PKA activity during the 5-HT applications has only a modest effect on the increase in sensorin expression. MAPK activation, which is critical for LTF (Martin et al., 1997; Purcell et al., 2003; Hu et al., 2004a), is not likely to be involved with the early rapid increase in sensorin. MAPK activation occurs later and only after significant secretion of sensorin (Martin et al., 1997; Purcell et al., 2003; Hu et al., 2004a) (see below).
Another candidate pathway that might regulate sensorin synthesis is PI3K, recently implicated in LTF (Udo et al., 2005). PI3K activation can be initiated via the same G-protein-coupled receptors that activate PKA or PKC (Khan et al., 2001; Pepio et al., 2002; Brock et al., 2003). Repeated applications of 5-HT would then lead to phosphorylation of a PI3K substrate Akt/protein kinase B (J. Liu, J.-Y. Hu, J. Schwartz, and S. Schacher, unpublished observations). One of the downstream consequences of PI3K activity is the activation of rapamycin-sensitive protein synthesis (Yanow et al., 1998; Dufner and Thomas, 1999), which also is required for LTF (Casadio et al., 1999; Hu et al., 2004a). The rapamycin-sensitive synthesis of sensorin also could be affected by 5-HT by a pathway that is independent of PI3K (Khan et al., 2001). PI3K activity and rapamycin-sensitive synthesis are important at distal sites, because both LY294002 and rapamycin block the increase in sensorin at varicosities even in the absence of sensory neuron cell bodies. The local accumulation of sensorin mRNA at varicosities (Schacher et al., 1999; Hu et al., 2002) can serve as the template for the local translation of sensorin. In the absence of the sensory neuron cell body, protein synthesis-dependent LTF and the initial formation of new presynaptic varicosities (Liu et al., 2003; Grabham et al., 2005) are likely to be mediated by the increase in the local synthesis and secretion of sensorin (Hu et al., 2004a,b). Thus, both the machinery for protein synthesis and its regulation are distributed at various locations in the sensory neuron. Sensorin joins the list of proteins for which the local synthesis contributes to long-term synaptic plasticity (Miller et al., 2002; Steward and Worley, 2002; Huang et al., 2005; Smith et al., 2005). The stimulus-induced regulation of neuropeptide synthesis is also observed for BDNF after stimuli that produce long-lasting forms of LTP in the hippocampus (Hall et al., 2000; Pang et al., 2004).
Type II PKA regulates sensorin secretion and responses that are critical for LTF
Sensorin secretion is enhanced significantly by repeated applications of 5-HT (Hu et al., 2004a). Medium collected from cocultures 2 h after the treatment with 5-HT has significant levels of sensorin. This “conditioned medium” containing secreted sensorin will evoke LTF when it is applied to naive cocultures after a single application of 5-HT (sensorin-induced LTF) [Hu et al. (2004a), their Fig. 3]. The significant decline in the newly synthesized sensorin in the axons and varicosities of the sensory neuron during the 2 h period after the treatment with 5-HT corresponds with the period when sensorin is secreted. The secretion during this period is critical for LTF because anti-sensorin Ab applied during this 2 h period blocked LTF, whereas incubation for a 2 h period with anti-sensorin Ab beginning at 3 h after the treatment with 5-HT failed to interfere with LTF. Which signaling pathway regulates sensorin secretion immediately after the treatment with 5-HT?
Type II PKA activity, specifically the phosphorylation of substrates near the membrane mediated via interactions between the regulatory subunit of type II PKA (RII) with anchoring proteins (AKAPs) (Moita et al., 2002; Liu et al., 2004), is required for the enhanced secretion of sensorin. This activity might include phosphorylation of vesicle proteins that would facilitate the enhanced secretion of the neuropeptide (Goodman et al., 1996; Han et al., 1999; Jovanovic et al., 2000; Angers et al., 2002). Because cAMP is enhanced in a persistent manner by the repeated applications of 5-HT (Bernier et al., 1982; Greenberg et al., 1987; Muller and Carew, 1998; Liu et al., 2004), the increased PKA activity at varicosities could regulate the large release of sensorin needed to produce LTF. Type II PKA activity at or near the membrane also regulates the ability of sensorin to activate autocrine responses in the sensory neurons (Hu et al., 2004a) and to produce LTF. PKA activity near the membrane may lead to the phosphorylation of sensorin receptors, molecules involved with receptor trafficking, or the associated molecules for activating downstream signaling pathways (Meyer-Franke et al., 1998; Du et al., 2000; Patterson et al., 2001).
Our results suggest that the sequential activation by 5-HT of specific signaling pathways is critical for the development of LTF. The type II PKA-dependent secretion of the newly synthesized sensorin follows the rapid PI3K- and rapamycin-dependent increase in sensorin synthesis at distal sites. Sensorin secretion and the type II PKA-dependent regulation of the autocrine responses of sensorin subsequently lead to the activation of MAPK (Hu et al., 2004a; Ormond et al., 2004). Because all of these signaling pathways are present in the distal processes of the sensory neurons, local synapse-specific changes in the synthesis and secretion of sensorin and activation of downstream signaling pathways can participate in the “tagging” of activated synapses for expressing long-term synapse-specific plasticity (Frey and Morris, 1997; Casadio et al., 1999). Later changes in sensorin mRNA expression, perhaps produced by MAPK translocation and persistent PKA activity in the sensory neuron nuclei that regulate the activity of transcription factors (Dash et al., 1990; Bacskai et al., 1993; Martin et al., 1997; Casadio et al., 1999; Chain et al., 1999; Roberson et al., 1999; Davis et al., 2000; Scott et al., 2002), might contribute to the second wave of increased sensorin synthesis detected by 12 h after 5-HT. A second wave of protein synthesis 6–12 h after 5-HT is critical for the long-lasting form of LTF detected at 72 h (Casadio et al., 1999; Chain et al., 1999; Sherff and Carew, 1999). This second increase in sensorin protein may produce a proportional increase in the constitutive secretion of sensorin needed to maintain the synaptic changes. Long-term maintenance of synaptic contacts requires the constitutive secretion of sensorin (Hu et al., 2004b). Thus, combined with the earlier changes produced by the activation of the various signaling pathways by 5-HT and sensorin, long-term changes in synapse function and structure are initiated and maintained.
Footnotes
This research was supported by National Institutes of Health (NIH) Grants MH 60387 and NS 42159. Animals were provided by the National Center for Research Resources for Aplysia, which is supported by NIH Grant RR-10294, at the University of Florida (Miami, FL). We thank Drs. Jinming Liu and James Schwartz for their technical advice and their comments on previous drafts of this manuscript.
Correspondence should be addressed to Dr. Samuel Schacher, Center for Neurobiology and Behavior, Columbia University College of Physicians and Surgeons, New York State Psychiatric Institute, 1051 Riverside Drive, New York, NY 10032. E-mail: sms2{at}columbia.edu.
DOI:10.1523/JNEUROSCI.4258-05.2006
Copyright © 2006 Society for Neuroscience 0270-6474/06/261026-10$15.00/0





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}