

Abstract
The corticotropin-releasing factor (CRF) system is the primary central mediator of stress-like states, coordinating behavioral, endocrine, and autonomic responses to stress. Although induction of anorexia is a well documented effect of CRF receptor agonist administration, the central sites and behavioral processes underlying this phenomenon are poorly understood. The present studies addressed this question by examining the neuroanatomical, behavioral, and pharmacological mechanisms mediating decreases in feeding produced by the CRF1/CRF2 receptor agonist urocortin. Separate groups of food-restricted male Sprague Dawley rats were given infusions of urocortin (0, 50, 125, 250 ng/0.5 μl) into the lateral septum (LS) and immediately afterward were rated on a wide array of behaviors (locomotion, rearing, grooming, stereotypies) including a microstructural analysis of ingestive behavior. Intra-LS urocortin infusion dose-dependently reduced feeding and drinking while concomitantly increasing grooming, stereotypies, and ethological plus traditional measures of anxiety-like responses in the elevated plus-maze. Urocortin infusion into neighboring sites (lateral ventricle, medial caudate) had no effects. Coinfusion into the LS of the mixed CRF1/CRF2 receptor antagonist d-Phe-CRF(12–41) (0, 100, 1000 ng/0.5 μl) or the novel selective CRF2 receptor antagonist Astressin2B (0, 500, 1000 ng/0.5 μl) blocked urocortin-induced effects, but the CRF1-selective antagonist NBI27914 (0, 500, 1000 ng/0.5 μl) had no effect, although it completely reversed the behavioral sequelae of CRF when infused into the basolateral amygdala. These results indicate that one of the modes through which the CRF system promotes anorexia is the recruitment of stress-like states after stimulation of CRF2 receptors within the LS.
Introduction
Corticotropin-releasing factor (CRF) receptor agonists potently reduce food intake across species (Zorrilla et al., 2003). Yet, whether these anorectic effects are dissociable from anxiety-promoting actions remains unclear, because the CRF system is the primary central mediator of stress responses and contributes to the etiology of stress-related psychiatric illness (Dautzenberg and Hauger, 2002; Reul and Holsboer, 2002; Bale and Vale, 2004). Given the current epidemic of obesity in the United States, there is great interest in identifying agents that reduce ingestion without producing such deleterious side effects; CRF2 receptor agonists have been proposed as such (Zorrilla et al., 2003).
The CRF system exerts a complex regulatory influence over feeding, the nature of which is dependent on receptor subtype, peripheral versus central sites, exogenous versus endogenous mechanisms, and gene knock-out strategies versus acute pharmacological manipulations. Despite these numerous factors, there is consensus that stimulation of CRF1 receptors reduces feeding and recruits anxiety-like behaviors in multiple paradigms regardless of the method of receptor targeting (genetic modification or drug infusion) or the brain region studied (Zorrilla et al., 2003). The function of the CRF2 receptor, however, is less straightforward. A plethora of evidence suggests that CRF2 receptor stimulation decreases feeding, but the role of CRF2 in stress-like behaviors remains unclear. Some studies of CRF2 receptor knock-out mice indicate that deletion of this receptor increases anxiety-like responses, leading to the suggestion that stimulation of CRF2 provides a tonic inhibitory influence over stress-like states that may facilitate the “coping” responses after exposure to stress (Bale et al., 2000; Coste et al., 2000; Kishimoto et al., 2000). Yet, because basal expression of CRF was increased in CRF2 knock-out animals, an alternative explanation is that their enhanced stress-like responses were attributable to increased stimulation of CRF1 receptors rather than CRF2 deletion itself. Recent studies with intracerebroventricular delivery of the CRF2-selective endogenous agonists urocortin2 and urocortin3 reveal that these peptides cause a delayed-onset reduction in feeding and gastric emptying, prompting the hypothesis that drugs targeting this receptor may cause anorexia without anxiety-like effects (Zorrilla et al., 2003; Czimmer et al., 2006). Nevertheless, several reports indicate that stimulation of CRF2 receptors recruits anxiety-like behaviors (Moreau et al., 1997; Ho et al., 2001; Bakshi et al., 2002; Spina et al., 2002; Hammack et al., 2003; Henry et al., 2006).
One factor contributing to the apparent discrepancy between these theories of CRF2 receptor function is the paucity of studies combining site-specific manipulations of CRF2-containing regions (particularly extrahypothalamic sites) with a detailed observational analysis of the behaviors accompanying the alterations in consumption (acute microstructural analysis of feeding and concomitant rating of other behaviors to simultaneously assess feeding and stress-like effects). The present studies addressed these issues and also examined the relative contributions of CRF receptor subtypes to these effects by characterizing the mechanisms through which the CRF1/CRF2 agonist urocortin (UCN) reduces feeding after microinfusion into the lateral septum (LS), a site that has a high concentration of CRF2 receptors and UCN-containing terminals and has been found recently to mediate UCN-induced anorexia (Bakshi et al., 2001; Wang and Kotz, 2002).
Materials and Methods
Animals
Ninety-three male Sprague Dawley rats (Charles River Laboratories, Wilmington, MA), weighing 290–320 g at the beginning of each experiment, were used in the present studies. Rats were pair housed in clear plastic cages in a temperature- and humidity-controlled vivarium. During experiments, animals were kept on a 20 h food-restriction schedule (food removed at 5:00 P.M., food returned at 1:00 P.M., or on test days, at the beginning of the test session). Lights in the animal colony were on a 12 h light/dark cycle (lights on at 7:00 A.M.), and all testing occurred between 9:00 A.M. and 1:00 P.M. On arrival, rats were handled daily for 5 min by the experimenters to minimize stress during experiments. Animal facilities were approved by the Association for the Assessment and Accreditation of Laboratory Animal Care; protocols were in accordance with the Guiding Principles in the Care and Use of Animals provided by the American Physiological Society and the guidelines of the National Institutes of Health. All efforts were made to prevent animal suffering and minimize the number of animals used for the studies.
Surgery
All surgeries took place within 1 week of arrival. Rats were anesthetized with sodium pentobarbital (50 mg/kg; Abbott Labs, North Chicago, IL) and treated with 0.1 ml of atropine sulfate (Phoenix Pharmaceuticals, St. Joseph, MO) to minimize respiratory distress. Using a stereotaxic apparatus (David Kopf Instruments, Tujunga, CA), stainless steel cannulas (23 gauge) were implanted bilaterally and affixed to the skull with dental cement (Lang Dental, Wheeling, IL) and skull screws (Small Parts, Miami Lakes, FL). Cannulas were aimed at the LS [anteroposterior (AP), +0.4 mm from bregma; mediolateral (ML), ±0.8 mm from midline; dorsoventral (DV), −3.5 mm from skull surface], the lateral ventricle (LV) (AP, −1.0 mm from bregma; ML, ±1.4 mm from midline; DV, −2.1 mm from skull surface), the medial caudate (MC) (AP, +0.4 mm from bregma; ML, ±2.2 mm from midline; DV, −2.5 mm from skull surface), or the basolateral amygdala (BLA) (AP, −3.0 from bregma; ML, ±5.0 mm from midline; DV, −5.8 mm from skull surface) [all coordinates are based on the atlas of Paxinos and Watson (1998)]. After surgery, rats were allowed 5–7 d to recover, during which time daily health checks and gentle handling were performed by the experimenter.
Drugs
UCN was obtained from Bachem-Peninsula Laboratories (Torrance, CA) and was dissolved with 10 mm HCl and sterile distilled water to a final pH of 6.5. The vehicle treatment for all experiments in which UCN was administered was sterile distilled water, pH 6.5. NBI27914 (NBI), a selective CRF1 receptor antagonist (Chen et al., 1996), was generously donated by Dr. D. Grigoriadis (Neurocrine Biosciences, La Jolla, CA) and was dissolved with sonication in a vehicle solution of 90% double-distilled water, 5% ethanol, and 5% cremaphor EL (Sigma, St. Louis, MO). This vehicle solution was used as the control treatment for all experiments in which NBI was administered. Astressin2B, a selective and long-acting CRF2 receptor antagonist (Rivier et al., 2002) (donated by Dr. W. Vale, Salk Institute, La Jolla, CA), was dissolved in sterile distilled water. Sterile distilled water was the vehicle treatment for all experiments in which Astressin2B was administered. All doses (see below) were calculated using the salt weight.
Microinfusion procedure
On all test days, animals were held gently, and their stylets were removed and placed into 70% ethanol. Cannulas were cleaned with a dental broach, and stainless steel injectors (30 gauge) were lowered so that they extended below the tips of the cannulas, making the final DV coordinates from skull surface −6.0 mm for the LS, −3.6 mm for the LV, −5.0 mm for the MC, and −8.8 mm for the BLA. The injectors were attached to polyethylene tubing, which was connected to 10 μl Hamilton microsyringes (Fisher Scientific, Pittsburgh, PA). The microsyringes were mounted on a motorized pump (Harvard Apparatus, Holliston, MA). A total of 0.5 μl of vehicle or drug solution was delivered per side over 93 s in each infusion. After the pump was shut off, the injectors were kept in place for an additional 60 s to let the infusion bolus absorb into the tissue. Injectors were then removed, stylets were replaced, and animals were placed immediately into test cages for behavioral testing. Two to 3 d before drug testing, all rats received a mock infusion in which injectors were lowered but no solution was administered. The mock infusion was intended to acclimate rats to the infusion procedure and to minimize stress attributable to infusions on the test days. After the mock infusion, animals were put into the test cages with food and water for 30 min to acclimate to the test environment.
Behavioral testing
On test days, after drug administration, rats were placed with food (lab chow) and water into test cages, which were identical to the rats' home cages but had a wire grate floor. Total duration, number of bouts (discrete episode of each behavior lasting continuously for at least 5 s), and mean duration of each bout were rated for 45 min for the following behaviors: feeding, drinking, rearing, and grooming. In addition, the latencies to feed and drink and the amount of food eaten (in grams) and water drunk (in milliliters) were measured. Locomotion, defined as the number of center-cage crosses, and stereotypies (perseverative oral behaviors directed toward inedible objects) were also measured. All of these behaviors were scored simultaneously and continuously by a trained experimenter (who was blind to the rats' treatments) during the test session; total food and water intake were calculated at the end of the session by subtracting the posttest amounts from the pretest amounts (correcting for the amount of food that spilled through the bottom of the cage during the test session). For testing in the elevated plus-maze (experiment 2 only), rats were placed individually into the center square of the apparatus facing the same closed arm immediately after intra-LS microinfusions. The apparatus was constructed from black Plexiglas, consisted of closed arms (bordered on three sides by a 40-cm-high wall) and open arms (no walls but bordered on three sides by a 0.5-cm-high ledge) arranged in a “+” configuration with like arms (each arm was 50 cm long × 10 cm wide) opposite each other [based on the parameters described previously (Heinrichs et al., 1992; Spina et al., 2000)], and was located in the center of a quiet, dimly lit room. After placing the rat in the plus-maze, the experimenter exited the room, and a video camera mounted on the ceiling above the apparatus recorded the rats' movements for 5 min on videotape, which was subsequently scored (by an experimenter blind to the rats' treatment condition) for the following measures: percentage of open arm time (seconds spent in open arms divided by the total number of seconds in the closed arms plus the open arms); percentage of open arm entries (number of entries into open arms divided by the total number of entries into closed arms plus open arms); number of stretch-attends (rat's hindlegs in a closed arm while head and forepaws stretch into the center or an open arm; proposed as an index of threat-assessment behavior that is increased during stress-like states) (Rodgers and Cole, 1993; Wall and Messier, 2001; Carobrez and Bertoglio, 2005). It should be noted that avoidance of novel, bright, open environments (i.e., the open arms of the elevated plus-maze) is most parsimoniously used as an operational definition of a stress-like state (File, 1996). The experimenter was blind to treatment conditions throughout all experiments.
Experimental design
Eight experiments were conducted in separate groups of experimentally naive rats. In all studies except experiment 2, rats were placed on a food-restriction schedule for the entire duration of the experiment (food removed at 5:00 P.M., food returned at 1:00 P.M., or on test days, at the beginning of the test session). Water was available ad libitum. In all experiments in which repeated tested occurred, a minimum of 48 h separated consecutive test days.
Intra-LS UCN infusion: detailed behavioral analysis.
Experiment 1 examined the behavioral effects of UCN infusion into the LS. Rats were given three different doses of UCN (50, 125, 250 ng/0.5 μl/side) or vehicle into the LS (n = 8) and were immediately placed into test cages. The doses of UCN used in the present experiment were chosen based on pilot studies that characterized the behavioral effects of UCN agonist infusion into various limbic structures (Bakshi et al., 2001). A repeated-measures design was used, with dosage as a within-subjects factor. Therefore, all rats received vehicle and all three doses of UCN over 4 counterbalanced test days (with a minimum of 48 h between each test day). In separate groups of experimentally naive rats (experiment 2), the effects of intra-LS UCN infusion (0, 125 ng; n = 6–13 per treatment) on anxiety-like responses in the elevated plus maze were evaluated; each rat was acclimated to the infusion procedure with mock infusions twice before the test day but received infusions and was tested just once in the plus-maze immediately after the intra-LS microinfusion (drug treatment as a between-subjects factor).
Behavioral effects of UCN infusion into regions neighboring the LS.
To confirm that the behavioral effects observed after intra-LS infusion of UCN were localized specifically to the LS and were not attributable to diffusion of UCN to adjacent areas, the behavioral effects of UCN (0, 50, 125, 250 ng/0.5 μl/side) infused into regions proximal to the LS were determined. Thus, in experiment 3, rats (n = 7) had cannulas aimed at the LV, and in experiment 4, UCN infusions were made into the MC (n = 8). All aspects of the experimental design, infusion, and testing procedures in experiments 3 and 4 were identical to those used in experiment 1.
CRF receptor mediation of LS effects: blockade of UCN-induced behaviors.
To confirm the pharmacological specificity of UCN-induced effects to the CRF system, experiment 5 examined the ability of the mixed CRF1/CRF2 receptor antagonist d-Phe-CRF(12–41) (d-Phe) to reverse the behavioral effects of intra-LS UCN in experimentally naive rats. Thus, in two separate groups of rats, the effects of a low (100 ng; n = 6) or high (1000 ng; n = 9) antagonist dose on UCN-induced effects were examined. In both studies, rats received intra-LS infusion of vehicle or d-Phe, followed immediately by vehicle or UCN (250 ng), and then were tested immediately afterward in the behavioral observation test cages. All rats received all four treatment combinations (vehicle/vehicle, vehicle/UCN, antagonist/vehicle, antagonist/UCN) in a counterbalanced order over 4 test days that were separated by at least 48 h. To determine the relative contributions of the CRF1 and CRF2 subtypes to these effects, the abilities of a CRF1-selective antagonist [NBI; CRF1/CRF2 affinity >10,000 (Chen et al., 1996)] and a CRF2-selective antagonist [Astressin2B; CRF2/CRF1 affinity >500–1000 (Rivier et al., 2002)] to block intra-LS UCN-induced behavioral changes were assessed in experiments 6 and 7, using the same design and protocol as the d-Phe experiment. Thus, in experiment 6, the effects of either a moderate (500 ng; n = 8) or a high (1000 ng; n = 8) dose of NBI were examined; in experiment 7, the effects of 500 ng (n = 7) or 1000 ng (n = 8) of Astressin2B were studied. Antagonist doses were chosen based on our previous work indicating that this dose range prevents the behavioral effects elicited by CRF agonist infusion into other brain regions (Jochman et al., 2005).
CRF receptor mediation of BLA effects: positive control for CRF1 antagonism in LS.
To verify that the doses of NBI that failed to alter UCN-induced effects after intra-LS infusion were behaviorally active, an additional study was performed in experimentally naive rats (experiment 8; n = 8) in which NBI (0, 1000 ng) was infused into the BLA immediately before infusion of CRF (0, 200 ng) into this site, using the within-subjects counterbalanced design applied in experiments 5–7. Rats were then tested in behavioral observation cages as described above. Our previous work indicates that CRF infusions into the BLA cause dramatic reductions in feeding and increases in grooming behavior similar to those observed in the present studies with UCN into LS (Jochman et al., 2005). Based on this work, it was predicted that this dose of NBI would reverse the intra-BLA CRF-induced anorexia.
Note that because UCN is the primary ligand that is expressed in the LS, whereas CRF is the predominantly expressed ligand in the BLA (Kozicz et al., 1998), we chose to infuse UCN in LS and CRF into amygdala. Nevertheless, because UCN and CRF have very similar affinities for the CRF1 receptor (Dautzenberg and Hauger, 2002) and we used similar doses of UCN and CRF in our studies, it is reasonable to compare results of the NBI experiments across these ligands. It also should be noted that the dose of NBI that was used (1000 ng/side) is in excess of the concentration required to bind all CRF1 receptors in the BLA (Bakshi et al., 2002; D. Grigoriadis, personal communication). Given that the LS expresses far fewer CRF1 receptors than the BLA (Dautzenberg and Hauger, 2002), this dose of NBI would be more than enough to bind all CRF1 receptors in the LS.
Histology
At the end of the experiments, rats were overdosed with sodium pentobarbital (100 mg/kg) and perfused transcardially with isotonic saline followed by 10% formalin. Brains were removed, stored in formalin, frozen with powdered dry ice, and sliced into 60 μm coronal sections using a cryostat (Leica, Deerfield, IL). After staining with cresyl violet, sections were examined under a microscope for the location of injector tips. Animals whose injector tip placements fell outside the targeted brain regions were excluded from analyses of behavioral data. For experiment 2 (intracerebroventricular UCN infusions), anesthetized rats received intracerebroventricular infusion of 5 μl of Chicago Sky Blue Dye (Sigma). One minute after infusion of the dye, rats were decapitated and brains were sliced into 1 mm sections and examined for the presence of dye within ventricles distal to the infusion site. Only rats for which blue dye was observed in the third and fourth ventricles were considered to have accurate cannula placements in the LV. The experimenter was blind to rats' experimental grouping as well as the behavioral data at the time of histological analyses.
Data analysis
For experiments 1–4, in which UCN was infused into the LS, LV, or MC, separate one-factor ANOVAs were used to analyze each behavioral measure, with drug dosage as the within-subjects factor and each behavioral measure as different dependent variables. Subsequent Newman–Keuls post hoc tests were performed to determine dosage effects in each site. For experiments 5–7, in which rats were infused intra-LS with d-Phe, NBI, or Astressin2B and then infused intra-LS with UCN, two-factor ANOVAs were run for each behavioral measure, with antagonist pretreatment and drug treatment as the within-subjects factors. Similarly, separate within-subjects two-factor ANOVAs were used to analyze data from experiment 8 (intra-BLA CRF antagonists vs CRF). Post hoc comparisons were made using Tukey's test; the α level was set at 0.05.
Results
Experiments 1 and 2: UCN in LS reduces ingestive behaviors and recruits stress-like responses
Figure 1 depicts the effects of intra-LS UCN infusion on ingestive behaviors; all graphs, except J and I, depict data from the entire 45 min test session. ANOVA indicated a main effect of UCN administration on total food intake (F(3,18) = 3.38; p < 0.04), with the highest dose of UCN significantly reducing grams of food ingested (p < 0.05) (Fig. 1A). No effects were observed on the latency to begin eating or the total number of eating bouts, suggesting that UCN infusion into LS did not alter the motivation to begin feeding or the ability to initiate feeding bouts. UCN significantly reduced the total amount of time spent feeding (F(3,18) = 3.15; p < 0.05; main effect of UCN), which was most apparent during the middle portion (16–30 min after infusion) of the test session (Fig. 1G) and reached statistical significance for the highest dose (p < 0.05). Moreover, examination of the average duration of a feeding bout revealed that UCN markedly reduced this measure (F(3,18) = 14.2; p < 0.01) at all three doses (p < 0.05) during the middle portion (minutes 16–30 after infusion) of the test session (Fig. 1I). Thus, UCN infusion into LS caused a significant reduction in food intake that was not accompanied by alterations in feeding latency or number of feeding bouts but was associated with reduced average feeding bout duration. This profile of results suggests that intra-LS UCN did not affect motivation or ability to initiate feeding but significantly reduced the ability to maintain feeding once it had been initiated.

Microstructural analysis of eating (top row) and drinking (bottom row) after UCN infusion into LS. Shown are effects on total food/water intake over 45 min (A, B), latency to eat/drink (C, D), total number of eating/drinking bouts in the entire test session (E, F), total time spent eating/drinking during the entire session (G, H), and average duration of an eating/drinking bout during the middle portion (minutes 16–30 after infusion) of the test session (I, J). Values represent mean ± SEM for each treatment. *p < 0.05, compared with vehicle. VEH, Vehicle. UCN doses are in nanograms per 0.5 μl/side.
A similar pattern of results was seen for drinking behavior. Water intake was also decreased by intra-LS UCN infusion (F(3,18) = 5.89; p < 0.01), with both the middle and highest dose reaching statistical significance (p < 0.05) (Fig. 1B). No effects were observed on latency to drink or total number of drinking bouts, but a main effect of UCN treatment was indicated by ANOVA for total time spent drinking (F(3,18) = 7.37; p < 0.01), with all three doses producing this effect (p < 0.05 for 50 and 125 ng doses and p < 0.01 for 250 ng dose). This effect was likely caused by the significant decrease in the average duration of a drinking bout produced by intra-LS UCN (F(3,18) = 4.56; p < 0.02), which was statistically significant at the middle and highest doses (p < 0.05), and mirrored the profile observed for feeding (Fig. 1J). Thus, as with feeding, intra-LS UCN decreased total amount of water drunk, total time spent drinking, and average drinking bout duration without affecting latency to drink or total number of drinking bouts. As with feeding, this profile suggests that UCN did not alter the motivation or ability to initiate drinking but, rather, disrupted the maintenance of this behavior.
Figure 2 illustrates the effects of intra-LS UCN administration on exploratory behaviors and stereotypies. These measures were rated to evaluate the effects of intra-LS UCN on noningestive behaviors and to determine the global behavioral profile of UCN administration in the rats. No effects were seen on locomotion (cage crossings), suggesting that the decrease in ingestive behaviors in these rats was not attributable to a change in general activity levels. Rearing was slightly decreased by intra-LS UCN (F(3,18) = 8.08; p < 0.01) at the highest dose (p < 0.05) (Fig. 2B). Although the total number of grooming bouts were not affected, the total time spent grooming (F(3,18) = 4.79; p < 0.02) was significantly increased by the highest dose (p < 0.01) (Fig. 2D) and was accompanied by a strong trend toward an increase in the average duration of an individual grooming bout (F(3,18) = 2.59; p < 0.08) (Fig. 2E). Interestingly, this grooming profile was complementary to that seen for ingestive behaviors, with UCN altering the maintenance rather than the initiation of this displacement behavior. In addition, infusion of UCN into the LS markedly increased the incidence of oral and whole-body stereotypies: noningestive licking (licking of any objects including the self that are not food or water), biting (of inedible objects), self-gnawing, wet dog shakes, and freezing (F(3,18) = 3.52; p < 0.04), particularly at the highest dose (p < 0.05) (Fig. 2F). Thus, the same dose of UCN that decreased food intake after intra-LS administration did not increase overall activity levels but did recruit several displacement behaviors that are often associated with stress (Berridge et al., 1999).

Effects of UCN infusion into LS on noningestive behaviors over a 45 min test session: A, locomotion; B, rearing; C, total number of grooming bouts; D, total time spent grooming; E, average duration of a grooming bout; F, total number of stereotypy bouts. Values represent mean ± SEM for each treatment. +p < 0.08, *p < 0.05, **p < 0.01, compared with vehicle. VEH, Vehicle. UCN doses are in nanograms per 0.5 μl/side.
To more directly examine the relationship between stress-like and ingestive behaviors induced by UCN infusion into the LS, the effects of intra-LS infusion of a dose of UCN that failed to decrease food intake (125 ng) was assessed on behavior in the elevated plus-maze. UCN infusion significantly decreased the percentage of time spent in the open arms (F(1,17) = 8.29; p < 0.02) and the percentage of open arm entries (F(1,17) = 6.19; p < 0.03) (Fig. 3A,B), consistent with the profile that is produced by stress in this paradigm and which is presumed to indicate an anxiety-like state in the rat (Koob et al., 1993). Moreover, a pronounced increase in threat-assessment behavior, as indexed by the number of stretch-attends (Fig. 3C), was produced by intra-LS UCN (F(1,17) = 7.68; p < 0.02). These increases in stress-like behaviors in the elevated plus-maze were not accompanied by general behavioral activation, because UCN infusion had no effect on the total number of arm entries (Fig. 3D). Thus, a potent induction of anxiety-like behaviors was recruited by intra-LS infusion of a dose of UCN that was half the concentration of that needed to reduce food intake.

Effects of intra-LS UCN infusion in the elevated plus-maze during a 5 min test session: A, percentage of time spent in open arms; B, percentage of entries into open arms; C, risk assessment indexed by the total number of stretch-attends; D, total entries into open and closed arms. Values represent mean ± SEM for each treatment. *p < 0.05, compared with vehicle. VEH, Vehicle. UCN doses are 125 ng/0.5 μl/side.
Experiments 3 and 4: no effects of UCN infusion into regions neighboring the LS
To determine the anatomical specificity of the LS UCN effects, the behavioral sequelae of UCN infusion into regions neighboring the LS were measured. Figure 4 illustrates the lack of any behavioral effects after UCN infusion into the LV or MC, contrasting with the potent behavioral effects following intra-LS UCN infusions (as described above). Thus, the behavioral effects of intra-LS UCN infusion could not be reproduced with infusion of UCN into regions proximal to the LS, indicating that these behavioral effects were localized specifically to the LS and were not caused by diffusion of UCN to the neighboring regions studied. For the sake of brevity, only data for time spent feeding, time spent drinking, time spent grooming, and stereotypy bouts are presented; however, no effects were observed on any behavior with UCN infusion into LV or MC. The LS data are the same as those presented in Figures 1 and 2 but are presented again here adjacent to the LV and MC results to illustrate the contrast between the sites.

Behavioral sequelae of UCN infusion into sites adjacent to the LS during a 45 min test session. Shown are effects on total time spent eating (A), total time spent drinking (B), total time spent grooming (C), and total number of stereotypy bouts (D). Values represent mean ± SEM for each treatment. *p < 0.05, **p < 0.01, compared with vehicle. VEH, Vehicle. UCN doses are in nanograms per 0.5 μl/side.
Experiments 5–7: mixed CRF1/CRF2 or CRF2-specific antagonists, but not a selective CRF1 receptor antagonist, block LS UCN effects
The effects of infusing the mixed CRF1/CRF2 antagonist d-Phe, the CRF1-selective antagonist NBI, or the CRF2-selective antagonist Astressin2B into the LS on UCN-induced behaviors are depicted in Figure 5. For the sake of brevity, only the results from the high-dose antagonist studies (1000 ng of each) are displayed; however, the results of the low-dose antagonist experiments are also described below. Food intake and grooming data are shown to represent the two domains that intra-LS UCN altered (ingestive behavior and displacement behaviors).

Effects over a 45 min test session of intra-LS CRF receptor antagonists on UCN-induced suppression of food intake (A) and increases in total time spent grooming (B). Values represent mean ± SEM for each treatment. **p < 0.01, compared with VEH/VEH; +p < 0.05, compared with VEH/UCN. VEH, Vehicle; ANTAG, antagonist. UCN doses are 250 ng/0.5 μl/side.
Food intake
Coinfusion of the CRF1/CRF2 antagonist d-Phe into the LS blocked intra-LS UCN-induced decreases in ingestive behavior, as indicated by a significant main effect of UCN treatment (F(1,8) = 26.7; p < 0.01) and a significant antagonist × agonist interaction (F(1,8) = 13.1; p < 0.01) (Fig. 5A). Post hoc comparisons of means revealed that UCN (250 ng) markedly reduced grams of food eaten (p < 0.01), replicating the effect observed in the original dose–response studies. In the d-Phe (1000 ng) plus UCN condition, food intake values were significantly higher than those for the vehicle plus UCN condition (p < 0.05) and were almost back to control (vehicle/vehicle) levels although the antagonist on its own did not affect food intake (F(1,8) = 0.1; NS). Similarly, pretreatment with the selective CRF2 receptor antagonist Astressin2B also reversed UCN-induced anorexia. ANOVA indicated a main effect of agonist treatment (F(1,7) = 37.2; p < 0.01) and a significant antagonist × agonist interaction (F(1,7) = 5.36; p < 0.05). Intra-LS UCN on its own was again found to reduce food intake compared with the vehicle-alone condition (p < 0.01), and pretreatment with Astressin2B significantly increased gram intake values in UCN-treated rats (p < 0.05) so that they no longer differed significantly from those of the vehicle/vehicle condition. In contrast to the mixed or CRF2-selective antagonists, the CRF1 antagonist NBI failed to alter UCN-induced decreases in food intake. A main effect of agonist treatment was found with ANOVA (F(1,7) = 63.6; p < 0.01), but no interaction between antagonist and agonist treatment was seen (F(1,7) = 0.6; NS). Post hoc analyses indicated that both the UCN-alone (p < 0.01) and the NBI/UCN (p < 0.01) conditions markedly reduced food intake compared with the vehicle/vehicle condition; there was no difference between the UCN-alone and NBI/UCN conditions. Thus, either a mixed CRF1/CRF2 or a selective CRF2 receptor antagonist reverses the reduction in ingestive behavior elicited by intra-LS UCN, but a selective CRF1 antagonist has no effect.
Grooming duration
Significant main effects of agonist treatment were observed in the d-Phe (F(1,8) = 24.6; p < 0.01), the NBI (F(1,7) = 67.7; p < 0.01), and the Astressin2B (F(1,7) = 51.3; p < 0.01) studies, with UCN significantly increasing total grooming duration compared with vehicle/vehicle values (p < 0.01) (Fig. 5B). A significant interaction between antagonist and agonist treatments was seen in the d-Phe study (F(1,8) = 5.7; p < 0.04), with post hoc comparisons of means indicating that d-Phe/UCN grooming values were significantly lower than those for the vehicle/UCN condition (p < 0.05) and that d-Phe/UCN values did not differ significantly from those of the vehicle/vehicle condition. A similar profile was seen with Astressin2B: there were trends toward a main effect of antagonist treatment (F(1,7) = 4.4; p < 0.07) and an antagonist × agonist interaction (F(1,7) = 3.3; p < 0.1), with coinfusion of the CRF2 antagonist causing grooming duration values in UCN-treated rats to be statistically indistinguishable from those of the vehicle/vehicle condition. In contrast, NBI failed to alter UCN-induced grooming (F(1,7) = 0.2, p < 0.69, main effect of antagonist; F(1,7) = 0.2, p < 0.68, antagonist × agonist interaction), with NBI/UCN values that were nearly identical to those of the vehicle/UCN condition and were significantly higher than those of the vehicle/vehicle condition (p < 0.01). Thus, the mixed CRF1/CRF2 antagonist and the selective CRF2 antagonist, but not the CRF1 antagonist, blocked the behavioral effects of UCN infusion into LS.
Low-dose antagonist experiments
In all three low-dose antagonist studies, significant main effects of UCN treatment were seen for all the measures described above for experiment 1, replicating the findings of this experiment. For the sake of brevity, only the effects of feeding and grooming are described. In all three studies, UCN significantly reduced feeding (p < 0.01) while increasing displacement behaviors such as grooming (p < 0.01). Coinfusion of a low dose (100 ng) of d-Phe into the LS had no effects on any UCN-induced behaviors. An interaction between pretreatment and treatment was seen after coinfusion of Astressin2B at a lower dose (500 ng) for food intake (F(1,7) = 6.4; p < 0.04), which during post hoc analyses revealed that food intake values for the Astressin2B/UCN condition were significantly higher than those for the UCN-alone condition (p < 0.05), although they still differed significantly from those of the vehicle/vehicle treatment (p < 0.05). Thus, the lower dose of the CRF2-selective antagonist caused a partial reversal of the UCN-induced anorectic effect. A similar trend was observed for grooming duration, with 500 ng of Astressin2B tending to partially reduce UCN-induced grooming, although this effect did not reach statistical significance. In contrast, the lower dose of the CRF1 antagonist NBI (500 ng) had no effects on any UCN-induced behaviors.
Experiment 8: CRF1 receptor antagonist does block behavioral effects of intra-BLA CRF administration
To verify that the doses of NBI used in the LS were sufficiently high to produce behavioral effects, experiment 8 tested the hypothesis that NBI would block the effects of CRF microinfusion into the BLA, which we previously showed to cause decreases in feeding and increases in grooming similar to those of UCN in the LS (Jochman et al., 2005). Figure 6 displays the results of this experiment. Only data for food intake and grooming duration are shown, because these are representative of the two main domains in which CRF agonists were found to affect behavioral responses. For food intake, ANOVA revealed main effects of antagonist pretreatment (F(1,7) = 7.5; p < 0.03) and CRF treatment (F(1,7) = 13.6; p < 0.01) and a significant interaction between antagonist and agonist (F(1,7) = 9.2; p < 0.02). Post hoc comparisons of means indicated that intra-BLA CRF infusion reduced food intake (p < 0.01), replicating our previously reported effect (Jochman et al., 2005), and that coinfusion of NBI completely blocked this reduction (p < 0.01), with values in the NBI/CRF condition indistinguishable from vehicle/vehicle levels (Fig. 6A). For time spent grooming, ANOVA indicated main effects of pretreatment (F(1,7) = 8.3; p < 0.03) and treatment (F(1,7) = 7.9; p < 0.03) and a trend toward an interaction between those factors (F(1,7) = 3.5; p < 0.1). Comparisons of means corroborated our previous finding (Jochman et al., 2005) that intra-BLA CRF infusion dramatically elevates grooming duration (p < 0.05) and that pretreatment with NBI significantly reduces grooming in CRF-treated rats (p < 0.05), with NBI/CRF values not differing from those of the vehicle/vehicle condition. Thus, intra-BLA infusion of NBI, at a dose that had no effects after intra-LS infusion, completely blocked the behavioral effects of intra-BLA CRF.

Effects of intra-BLA CRF1 receptor antagonist infusion on CRF-induced suppression of food intake (A) and increases in total time spent grooming (B) over a 45 min test session. Values represent mean ± SEM for each treatment. *p < 0.05, **p < 0.01, compared with VEH/VEH; +p < 0.05, ++p < 0.01, compared with VEH/CRF. VEH, Vehicle. Doses are as follows: CRF, 200 ng/0.5 μl/side; NBI, 1000 ng/0.5 μl/side.
Histological analyses
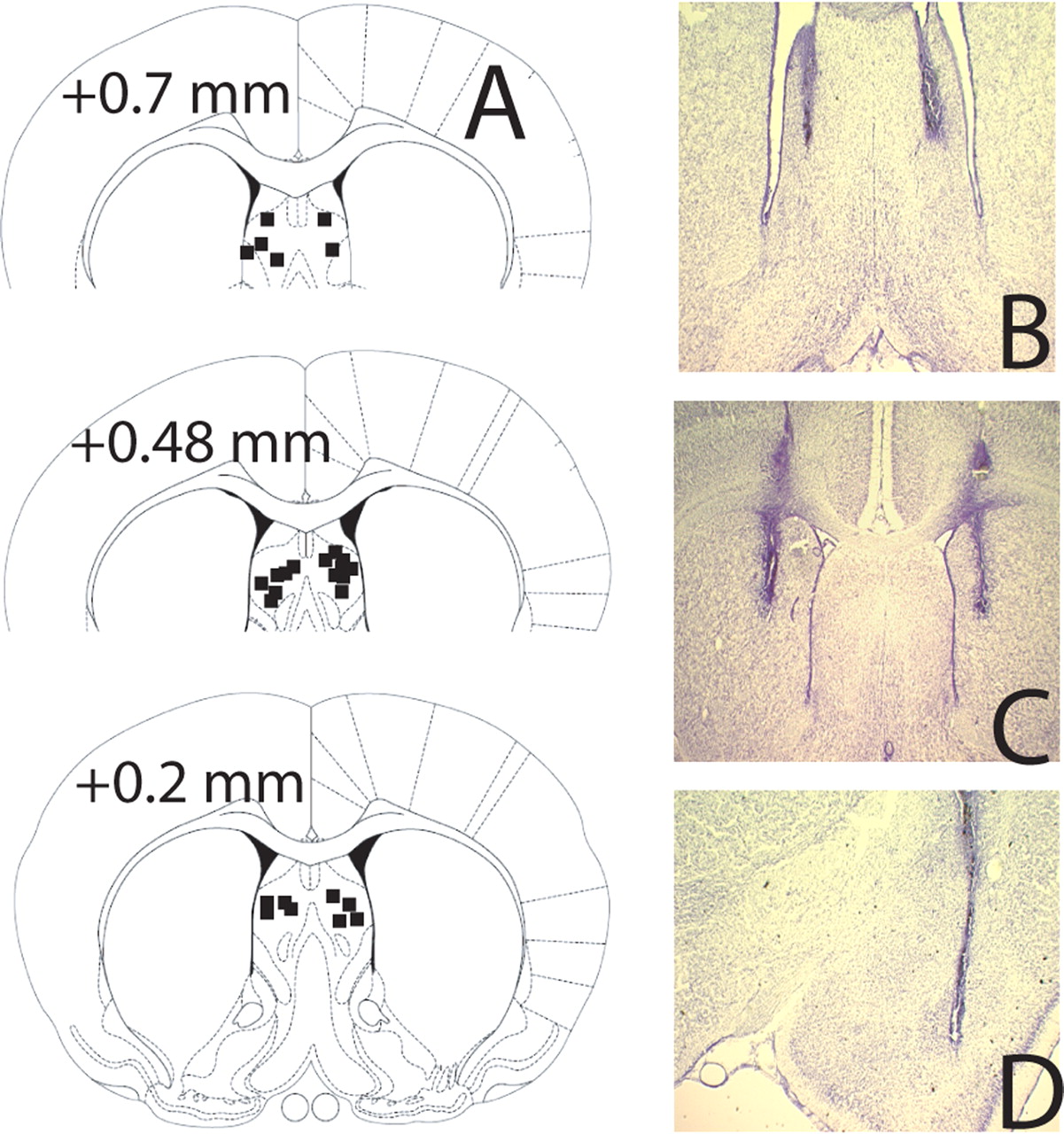
Figure 7 depicts the location of representative injector tip placements for the LS, the MC, and the BLA. An experimenter blind to the rats' treatments and behavioral data evaluated all placements to determine whether or not injectors fell within the boundaries of the targeted structures. Five LS placements (experiment 2), two MC placements (experiment 4), and one LV placement were found to be outside of the targeted regions, and the data from these rats were thus excluded from all analyses; the sample sizes given above reflect these histologically based omissions.

Location of injector tip placements within the LS (A, B), the MC (C), and the BLA (D).
Discussion
The present results provide important insights into the neuroanatomical, behavioral, and pharmacological mechanisms underlying the anorectic effects of UCN. First, the LS is a major extrahypothalamic site underlying the anorectic effects of UCN, because UCN infusion into LS but not neighboring regions reduces feeding behavior. Second, this anorectic effect does not arise from deficits in the motivation or ability to initiate feeding, but rather from a deficit in maintaining a prolonged feeding bout. Recruitment of displacement behaviors may underlie this inability to sustain ingestive responses, because doses of UCN that reduce food intake potently elicit perseverative grooming and oral stereotypies. Moreover, increased anxiety-like behaviors in the elevated plus-maze are produced by doses lower than those required to reduce food intake, additionally suggesting that reduced feeding after intra-LS UCN infusion may be secondary to the elicitation of stress-like states. Finally, the behavioral effects of intra-LS UCN are attributable to activation of CRF2 receptors, because coinfusion of a mixed CRF1/CRF2 or a CRF2-selective antagonist but not a CRF1 antagonist completely blocks these effects. Together, these findings suggest that stimulation of CRF2 receptors within the LS may promote anorexia through the induction of stress-like responses.
These findings confirm an important role for the LS in UCN-induced anorexia. It could be suggested that intra-LS UCN produces its behavioral effects via diffusion to the bed nucleus of the stria terminalis (BNST), which also contains CRF2 receptors and mediates anxiety-like behaviors (Erb and Stewart, 1999; Cooper and Huhman, 2005) such that UCN-induced anorexia is mediated by the LS but stress-like effects occur through the BNST. Yet, given that stimulation of CRF receptors in BNST also induces anorexia (Ciccocioppo et al., 2003), this possibility is not likely. It also is unlikely that intra-LS UCN diffuses to BNST because the primary route of diffusion would be up the injector tract but BNST is ventral to LS. Our previous work indicates that the infusion parameters used in the present studies allow for very anatomically restricted effects within the LS, because infusion of CRF system peptides into control sites such as the nucleus of the diagonal band (located ventral to the BNST and therefore more likely than LS to permit diffusion into BNST) fails to produce effects (Bakshi et al., 2002).
The present results are in agreement with those of Kotz and colleagues, who found that deprivation- or hypocretin-induced feeding are reduced by UCN infusion into LS (Wang and Kotz, 2002), and add to the growing body of literature indicating that CRF receptor stimulation in either hypothalamic or extrahypothalamic sites reduces food intake (Ohata et al., 2000; Currie et al., 2001; Wang et al., 2001; Daniels et al., 2004; Jochman et al., 2005; Weitemier and Ryabinin, 2006). The present studies also confirm our previous finding that CRF1 receptors within the BLA regulate feeding (Jochman et al., 2005). Importantly, the dose of CRF1 antagonist that failed to affect intra-LS UCN-induced effects is behaviorally active in our feeding paradigms because this dose administered into the BLA blocked the effects of CRF, used in the same dose range as UCN. Thus, multiple forebrain and brainstem sites appear to mediate the anorectic actions of CRF receptor agonists.
Previous evidence indicates that stimulation of either CRF1 or CRF2 receptors is sufficient to reduce ingestive behavior (Smagin et al., 1998; Cullen et al., 2001; Zorrilla et al., 2003; Bale and Vale, 2004). The present results provide the novel finding that the contributions of these two receptors may be anatomically segregated, with CRF1 receptors acting within the amygdala and CRF2 receptors acting within the LS in a parallel manner to regulate feeding. This notion of functionally homogenous but anatomically heterogeneous actions of CRF receptor subtypes also describes the nature of CRF1 and CRF2 receptor regulation of stress-induced defensive behaviors (Bakshi et al., 2002). The Edinger-Westphal nucleus (EW), which is the primary source of UCN to the LS, regulates ingestive behavior, and UCN infusion into other efferents of this nucleus also reduces feeding (Bittencourt et al., 1999; Weitemier and Ryabinin, 2005, 2006). Given that reduced feeding is accompanied by stress-like behaviors after intra-LS UCN and that stressors potently activate UCN-containing neurons of the EW, it is possible that the EW–LS projection represents an important anatomical substrate mediating stress-induced anorexia (Gaszner et al., 2004). One might also speculate that the periaqueductal gray (PAG), which receives projections from the LS and amygdala, has a well validated role in stress-induced behaviors, and has been implicated in the anorectic actions of certain peptides (Marchand and Hagino, 1983; Jenck et al., 1986; Kyrkouli et al., 1987; Kask et al., 2002), may participate in intra-LS UCN-induced anorexia. It would be of interest in future studies to determine whether the behavioral sequelae of intra-LS UCN infusion or EW stimulation are antagonized by blockade of PAG activity.
To our knowledge, the present studies provide the first detailed characterization of acute feeding microstructure with simultaneous evaluation of stress-related behaviors after site-specific microinfusion of UCN and offer important new information regarding the precise behavioral mechanisms through which CRF receptor stimulation reduces feeding. Intra-LS UCN significantly reduced food intake in the absence of changes in the latency to begin feeding or the total number of feeding bouts, suggesting that alterations in the motivation or ability to begin feeding did not account for the anorectic effect of UCN within the LS. Instead, the average duration of individual feeding bouts was significantly reduced. A similar profile was observed for drinking, indicating that UCN-induced effects were not specific to feeding and that CRF2 receptor stimulation within LS disrupted the maintenance rather than the initiation of ingestive behaviors. Although it remains to be determined whether CRF2 receptor stimulation in LS triggers satiety mechanisms, as has been suggested for intracerebroventricular infusion of urocortin2 (Inoue et al., 2003), the present findings suggest that this may not be the case after intra-LS UCN.
An alternative explanation is that feeding and drinking bouts were terminated prematurely because of the recruitment of noningestive behaviors that may reflect a stress-like interoceptive state. Microstructural analysis of grooming revealed that the same doses of UCN that reduced feeding bout duration markedly increased the average duration of a grooming bout. Although grooming can indicate satiety (Smith and Gibbs, 1992), it should be noted that the pattern of grooming elicited by intra-LS UCN did not fit the profile of the typical satiety sequence, because it was not accompanied by other crucial markers of satiety such as a reduction in meals or increased latency to initiate feeding. Instead, unusually long perseveration during each individual grooming episode and a concomitant elicitation of oral stereotypies were observed, suggesting that CRF2 receptor stimulation within LS promotes prolonged expression of displacement behaviors that may interfere with the maintenance of ingestive responses. The notion that perseverative grooming and oral stereotypies are indicators of stress-like states has been proposed previously (Berridge et al., 1999); however, to the best of our knowledge, the present results provide the first evidence for the proposition that such a mechanism underlies the anorectic effects of CRF2 receptor stimulation with LS. This proposed mechanism of CRF2-mediated anorexia is in agreement with a previous report demonstrating that decreases in feeding after intracerebroventricular CRF infusion were significantly correlated with stress-like behaviors such as freezing and were reversed by selective antagonism of CRF2 receptors (Pelleymounter et al., 2000).
Additional evidence for the hypothesis that the anorectic actions of UCN in LS are secondary to the recruitment of stress-like states is provided by the finding that a lower dose of UCN than that required to reduce food intake was sufficient to reduce open-arm exploration in the elevated plus-maze. This hypothesis is in contradiction to previous conceptualizations of the role of the CRF2 receptor in ingestive behavior, which have argued that stimulation of this receptor reduces feeding without inducing anxiety-like effects (Zorrilla et al., 2003, 2004), a notion that is based primarily on the finding that intracerebroventricular administration of urocortin2 and urocortin3 can cause delayed reductions in stress-like responses (Spina et al., 1996; Valdez et al., 2002, 2003, 2004; Venihaki et al., 2004). Yet, multiple studies, in addition to the present findings, indicate that particularly when administered site-specifically into extrahypothalamic regions including the LS, UCN or urocortin2 promotes stress-like behaviors through the activation of CRF2 receptors (Moreau et al., 1997; Radulovic et al., 1999; Pelleymounter et al., 2000, 2002, 2004; Spina et al., 2002; Risbrough et al., 2003; Henry et al., 2006).
CRF2 receptor stimulation within some sites (i.e., hypothalamic) could exert selective effects on feeding, but in other (extrahypothalamic) regions such as the LS, activation of these anorectic effects cannot be separated from the induction of stress-related behaviors. Given the current epidemic of obesity in the United States and the exigency to identify novel anorectic agents, one must be mindful of the possibility that stimulation of certain populations of CRF2 receptors might promote anorexia via the induction of stress-like states when considering the potential utility of CRF2 receptor agonists in this capacity.
Footnotes
-
This work was supported by National Institutes of Health Grant MH40855, the University of Wisconsin HealthEmotions Research Institute, and Meriter Hospital. We thank Dr. Dimitri Grigoriadis (Neurocrine Biosciences, La Jolla, CA) for the kind gift of NBI27914 and Dr. Wylie Vale (Salk Institute, La Jolla, CA) for the generous donation of Astressin2B.
- Correspondence should be addressed to Dr. Vaishali P. Bakshi, Department of Psychiatry, University of Wisconsin-Madison, 6001 Research Park Boulevard, Madison, WI 53719. vbakshi{at}wisc.edu





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}