

Abstract
The neuromodulator dopamine (DA) has multiple modes of action on neuroplasticity induction and modulation, depending on subreceptor specificity, concentration level, and the kind of stimulation-induced plasticity. To determine the dosage-dependent effects of D2-like receptor activation on nonfocal and focal neuroplasticity in the human motor cortex, different doses of ropinirole (0.125, 0.25, 0.5, and 1.0 mg), a D2/D3 dopamine agonist, or placebo medication were combined with anodal and cathodal transcranial direct current stimulation (tDCS) protocols, which induce nonfocal plasticity, or paired associative stimulation (PAS, ISI of 10 or 25 ms), which generates focal plasticity, in healthy volunteers. D2-like receptor activation produced an inverted “U”-shaped dose–response curve on plasticity for facilitatory tDCS and PAS and for inhibitory tDCS. Here, high or low dosages of ropinirole impaired plasticity. However, no dose-dependent response effect of D2-like receptor activation was evident for focal inhibitory plasticity. In general, our study supports the assumption that modulation of D2-like receptor activity exerts dose-dependent inhibitory or facilitatory effects on neuroplasticity in the human motor cortex depending on the topographic specificity of plasticity.
Introduction
Dopamine (DA) is involved in learning and memory formation, processes dependent on synaptic modifications like long-term potentiation (LTP) and long-term depression (LTD) (Huang et al., 2003; Jay, 2003; Manahan-Vaughan and Kulla, 2003; Knecht et al., 2004; Flöel et al., 2005), and an important factor in neuropsychiatric disorders like schizophrenia, restless legs syndrome, Parkinson's disease, and depression (Emilien et al., 1999). Despite advances in the understanding of the dopaminergic system, the impact of DA on synaptic plasticity is far from being completely understood. Opposing effects of DA on synaptic activity and cognition in animals and humans (Chen et al., 1996; Jay, 2003; Seamans and Yang, 2004), and nonuniform effects of DAergic pharmacotherapy in neuropsychiatric disorders point out its complex action on brain function (Kulisevsky, 2000; Obeso et al., 2000; Kaiser et al., 2002; Paulus and Trenkwalder, 2006). It has been proposed that the specific dopaminergic impact on neuroplasticity depends on subreceptor specificity, concentration levels of dopamine, and the type of cortical plasticity, e.g., focal or nonfocal plasticity (Millan et al., 2002; Centonze et al., 2003; Seamans and Yang, 2004; Collins et al., 2005; Nitsche et al., 2006; Kuo et al., 2008).
For DA concentration, it was demonstrated that either insufficient or excessive dopamine impairs cognitive functions (Cai and Arnsten, 1997; Goldman-Rakic et al., 2000; Seamans and Yang, 2004). It is believed that this inverted “U”-shaped effect of DA is mediated by D1-like receptors (Seamans and Yang, 2004). Also D2-like receptors are involved in plasticity generation and cognitive processes (Otani et al., 1998; Mehta et al., 1999; Spencer and Murphy, 2000; Manahan-Vaughan and Kulla, 2003; Gibbs and D'Esposito, 2005; Nitsche et al., 2006). However the results of different studies for the impact of D2-like activity on plasticity and behavior are conflicting (Jay, 2003): for LTD and LTP, D2-like activity has been described as enhancing (Otani et al., 1998; Spencer and Murphy, 2000; Manahan-Vaughan and Kulla, 2003) or diminishing (Frey et al., 1989; Chen et al., 1996) it. Similarly, behavioral studies in humans showed at least partially contradictory results. A positive effect of the D2-like receptor agonist (cabergoline) has been shown on encoding of a motor memory (Meintzschel and Ziemann, 2006), whereas pergolide (predominant D2-like dopamine agonist) impaired verbal associative learning (Breitenstein et al., 2006). These findings might at least partially be explained by a dosage-dependent nonlinear effect of D2-like activity on brain function.
Here we aim to characterize the dose-dependent effects of the D2-like receptor activation on neuroplasticity of healthy subjects in a placebo-controlled study design. The effect of D2-like activity on focal plasticity, induced by paired associative stimulation (PAS) (Stefan et al., 2000, 2002), and nonfocal, induced by transcranial direct current stimulation (tDCS) (Nitsche and Paulus, 2001) was explored, because it was demonstrated that DA has a discernable impact on both kinds of plasticity (Kuo et al., 2008). We hypothesize that D2-like receptor activation results in a dose-dependent inverted U-shaped impact on neuroplasticity. This effect might differ for focal and nonfocal plasticity.
Materials and Methods
Subjects.
Twelve healthy volunteers were included in each experiment [tDCS experiment, 0.125 mg of ropinirole (RP) group: six men, six women, aged 27.88 ± 5.1 years (mean ± SD); 0.25 mg of RP group: seven men, five women, aged 28.08 ± 6.2 years; 0.5 mg of RP group: five men, seven women, aged 26.73 ± 6.7 years; 1.0 mg of RP group: six men, six women; aged 25.81 ± 6.18 years and PAS experiment, 0.125, 0.5, and 1.0 mg of RP group: 6 men, 6 women, aged 28.92 ± 5.77 years]. Subjects were recruited from the campus of the Georg August University. The subjects included in the study were between 18 and 40 years old, had no history of chronic or acute neurological, psychiatric, or medical disease, no family history of epilepsy, no pregnancy, no cardiac pacemaker, no previous surgery involving implants to the head (cochlear implants, aneurysm clips, brain electrodes) and did not take acute or chronic medication or drugs. Participants were allocated randomly to tDCS and PAS groups. The order of stimulation protocol and drug condition were also randomized. Age (t test; p > 0.05) and gender (χ2; p > 0.05) did not differ significantly between groups. Before the sessions, written informed consent was obtained from all subjects. The experiments were approved by the ethics committee of the University of Goettingen, and we conform to the Declaration of Helsinki. Due to the multitude of sessions in the tDCS experiment (experiment 1), we performed a complete repeated-measures design only for the PAS experiment (experiment 2).
Nonfocal plasticity induction by tDCS (experiment 1).
We used a battery-driven constant current stimulator (Schneider Electronic) with a maximum output of 2 mA. tDCS was applied using a pair of saline-soaked surface sponge electrodes (35 cm2). One electrode was positioned over the motor cortex representational area of the right abductor digiti minimi muscle (ADM), the other electrode above the right orbit. tDCS was administered with a current strength of 1 mA for 13 (anodal tDCS, the polarity refers always to the motor cortical electrode) or 9 min (cathodal tDCS), which had been demonstrated to induce cortical excitability enhancement or inhibition, respectively, lasting for ∼1 h after the end of stimulation (Nitsche and Paulus, 2001; Nitsche et al., 2003). In addition a control experiment with sham tDCS was performed for the highest RP dosage (1 mg) to explore whether RP alone influences cortical excitability. Sham tDCS was performed by ramping current flow for the first 10 s of stimulation (as done also for real tDCS), but switching the stimulator off after 30 s (Gandiga et al., 2006).
Focal plasticity induction by PAS (experiment 2).
Peripheral nerve stimulation was delivered to the right ulnar nerve at the wrist level by a Digitimer D185 stimulator, followed by single-pulse transcranial magnetic stimulation (TMS) applied with a stimulator output resulting in motor-evoked potentials (MEPs) of ∼1 mV amplitude (“baseline-intensity”, see description below) over the motor cortex representing area of the right ADM. Ulnar nerve stimulation intensity was adjusted to 3.0 times of the individual perceptual threshold, defined as the lowest perceivable stimulus intensity. The interstimulus interval (ISI) between the peripheral and the TMS stimulus was set at 10 ms (inhibitory PAS; iPAS) or 25 ms (excitatory PAS; ePAS). Ninety pairs were delivered at 0.05 Hz over 30 min, which has been shown to induce long-lasting motor cortical excitability enhancement by ePAS protocol and inhibition by iPAS (Stefan et al., 2000).
Pharmacological intervention.
Low (0.125 and 0.25 mg in the tDCS experiments; 0.125 mg in the PAS experiments), medium (0.5 mg), or high (1.0 mg) dosages of RP (a D2/D3-specific dopamine agonist) or equivalent PLC drugs were taken by the subjects 1 h before the start of the plasticity-inducing cortical stimulation. A dose of 0.125 mg of RP was chosen as the lowest dosage, because a roughly equivalent dosage of pergolide affected tDCS-induced plasticity in a former study (Nitsche et al., 2006). The largest dosage the subjects tolerated without systemic side effects was 1 mg of RP; thus, 1 mg of RP was chosen as maximum dosage. Moreover, this dosage is similar to that RLS patients receive for therapeutic purposes. Doses of 0.25 and 0.5 mg of RP were administered to explore the dosage-dependent linearity of effects of RP. One hour after oral dosing, RP reaches maximum plasma concentration and produces prominent effects in the CNS (Brefel et al., 1998; Asquith and Egan, 1999). One hour before RP intake, 20 mg of domperidone were administered orally to minimize drug-induced systemic side effects of RP. Before and after tDCS and PAS, subjects were interviewed about possible adverse effects. The minimum interval between sessions was 1 week to avoid cumulative drug effects. Subjects were blinded to the medication condition.
Monitoring of motor cortical excitability.
TMS-elicited MEPs were recorded to measure excitability changes of the representational motor cortical area of the right ADM. Single-pulse TMS was conducted by a Magstim 200 magnetic stimulator (Magstim Company) with a figure-of-eight magnetic coil (diameter of one winding = 70 mm, peak magnetic field = 2.2 tesla). The coil was held tangentially to the skull, with the handle pointing backwards and laterally at an angle of 45° from midline. The optimal position was defined as the site where stimulation resulted consistently in the largest MEPs. Surface EMG was recorded from the right ADM with Ag–AgCl electrodes in a belly-tendon montage. The signals were amplified and filtered with a time constant of 10 ms and a low-pass filter of 2.5 kHz, then digitized at an analog-to-digital rate of 5 kHz and further relayed into a laboratory computer using the Signal software and CED 1401 hardware (Cambridge Electronic Design). The intensity was adjusted to elicit, on average, baseline MEPs of 1 mV peak-to-peak amplitude and was kept constant for the poststimulation assessment unless adjusted (see below).
Experimental procedures.
Both experiments (tDCS and PAS) were conducted in a repeated-measurement design. For the tDCS experiments, different subject groups participated in each drug dosage condition. For the PAS experiments, a complete cross-over design was performed. The subjects were seated in a comfortable chair, with head and arm rests. TMS was applied over the left motor cortical representational area of the right ADM where it produced consistently the largest MEPs in the resting ADM (optimal site). The intensity of the magnetic cortical stimulus was adjusted to elicit MEPs with a peak-to-peak amplitude of on average 1 mV (baseline 1). One hour after intake of RP or the equivalent placebo medication, a second baseline (baseline 2) was determined to control for a possible influence of the drug on cortical excitability and adjusted if necessary (baseline 3). In the tDCS experiment, the motor cortical DC stimulation electrode was then fixed above the left motor cortex and the other was fixed at the contralateral forehead. In the PAS experiment, the interventional PAS protocol as described above was used. Immediately after tDCS and PAS, 25 MEPs were recorded every 5 min at 0.25 Hz for 0.5 h, and then every 30 min up to 2 h after the end of each stimulation. For the drug conditions, TMS recordings were performed at four additional time points: same day evening (se), next morning (nm), next afternoon (na), and next evening (ne). The electrode and coil positions were marked with a waterproof pen to guarantee identical positions during the whole course of the experiment. Under PLC conditions, the aftereffects of tDCS and PAS were evaluated until 120 min after the stimulation, because we did not expect longer-lasting aftereffects in these conditions (Fig. 1). In the control experiment, where 1 mg of RP was combined with sham tDCS, cortical excitability was monitored for 60 min after sham stimulation.

Experimental course of the present study. TMS was applied over the left motor cortical representational area of the right ADM with an intensity to elicit MEPs with a peak-to-peak amplitude of on average 1 mV (baseline—BL 1). One hour after intake of RP, but not after placebo (PLC) medication, a second baseline (baseline 2—BL 2) was determined to control for a possible influence of the drug on cortical excitability and adjusted if necessary (baseline 3—BL 3). Two methods of plasticity induction were used, tDCS (experiment 1) and PAS (experiment 2). In the tDCS experiment, a current strength of 1 mA for 13 (anodal tDCS) or 9 min (cathodal tDCS) was applied. In the PAS experiment, ePAS (ISI of 25 ms) and iPAS (ISI of 10 ms) were used (for more details, see Materials and Methods). Immediately after tDCS and PAS, MEPs were recorded. For the drug conditions, TMS recordings were performed up to next evening after intervention. Under PLC, the aftereffects of tDCS and PAS were evaluated until 120 min after the stimulation.
Data analysis and statistics.
MEP amplitude means were calculated for each time bin, including baseline 1, 2, and 3 and poststimulation values. The postintervention MEPs were normalized and are given as ratios of the baseline determined immediately before intervention (baseline 3). For the tDCS experiment, an ANOVA model for repeated measures for the time bins up to 120 min after tDCS were calculated with the dependent variable MEP amplitude, with RP dosage (0.125, 0.25, 0.5, 1.0) as between-subject factor and the within-subject factors time course, tDCS (anodal and cathodal tDCS), and medication (real vs placebo). For the PAS experiment, we performed a repeated-measure ANOVA with the within-subject factors PAS (25 and 10 ms of stimulation interval), RP dosage (0.125, 0.5, 1.0, and placebo), and time course for MEPs up to 120 after stimulation. In difference to the tDCS study, the PAS experiment did not include a between-subject factor, because it was performed as a complete crossover study. Student's t tests (paired samples, two-tailed, p < 0.05) were performed to compare the MEP amplitudes before and after the interventional brain stimulations in each intervention condition and between drug conditions for each time point. A p value of <0.05 was considered significant for all statistical analyses. All results are given as mean and SEM. For the sham tDCS 1 mg of RP control study, we performed a one-factorial repeated-measures ANOVA with the dependent variable MEP amplitude and the within-subject factor time course. The Mauchly test of sphericity was checked and the Greenhouse-Geisser correction was performed, when appropriate.
Results
Two subjects experienced transient nausea and vomiting under the highest dose of RP, but the symptoms subsided before the neuroplasticity-inducing intervention. The remaining subjects tolerated the drugs well. Peak-to-peak baseline MEP amplitudes remained unaffected by RP, i.e., MEP amplitudes did not differ significantly before and after drug intake in all conditions (p ≥ 0.05, Student's t tests, paired, two-tailed) (see also supplemental material, available at www.jneurosci.org). A slight effect of 1.0 mg of RP on cortical excitability might be noticed; however, this was far from being significant. Moreover, MEP amplitudes were stable under the highest RP dose (1.0 mg) without additional plasticity-inducing stimulation (sham tDCS condition), confirming that RP alone did not affect corticospinal MEP amplitudes, as monitored by single-pulse TMS (Fig. 2).

Effect of D2-like activation under highest RP dosage (1 mg) on cortical excitability (control experiment). The control experiment with sham tDCS was performed for the highest RP dosage (1 mg) to observe whether RP alone influences cortical excitability. Shown are baseline-standardized MEP amplitudes after plasticity induction by cathodal and anodal tDCS under placebo medication condition and after 1.0 mg of RP intake without tDCS (Sham stimulation condition). Filled symbols indicate significant deviations of the MEP amplitudes from baseline values (Student's t test, two-tailed, paired samples, p < 0.05).
Dose-dependent effect of ropinirole on nonfocal tDCS-induced plasticity (experiment 1)
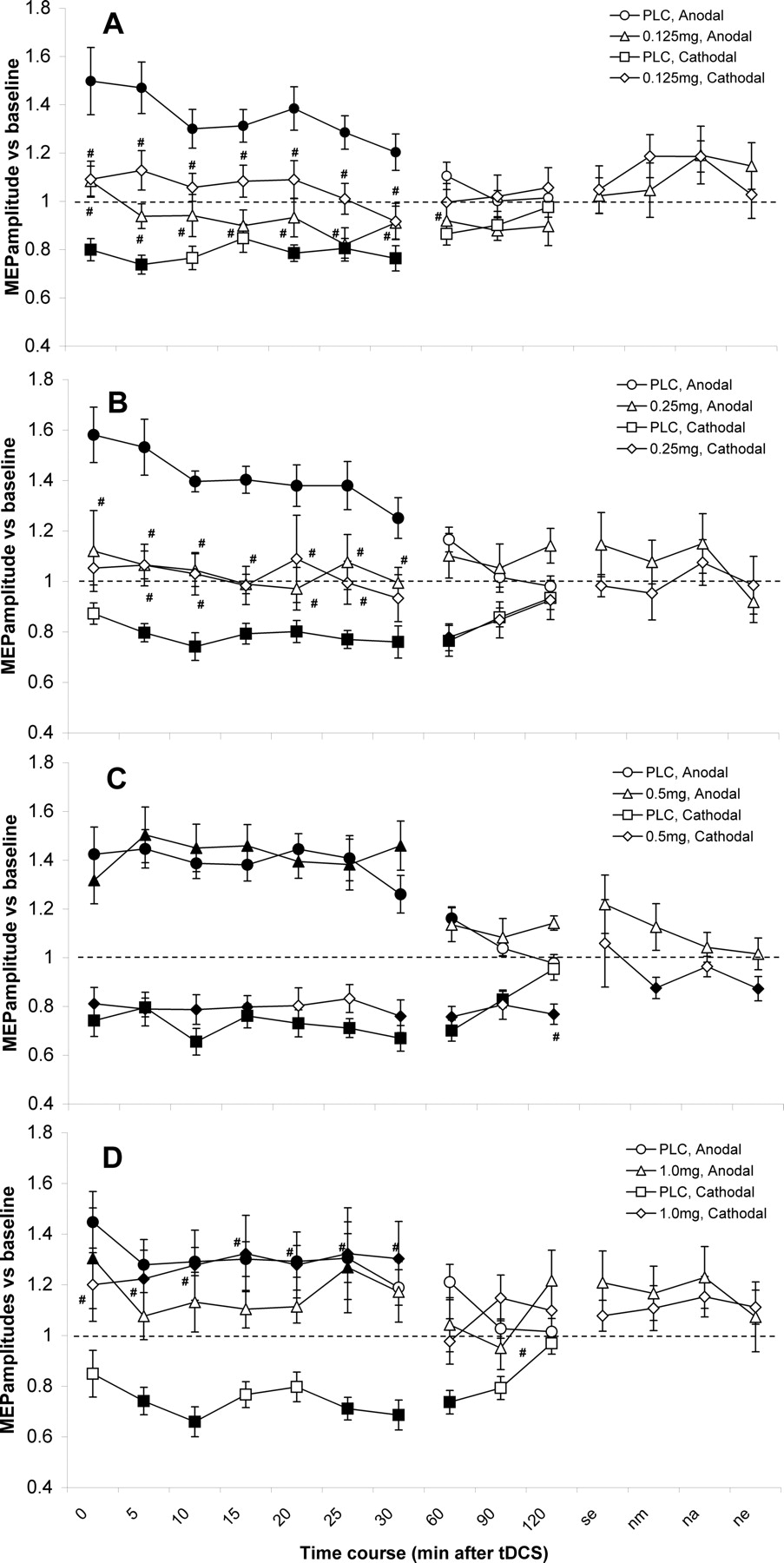
The ANOVA revealed a significant four-way interaction of time course × tDCS × dosage × medication (Table 1). Under placebo medication, anodal and cathodal stimulation significantly increased and decreased, respectively, cortical excitability until 1 h after the end of stimulation, confirming previous studies (Fig. 3). As revealed by the post hoc t tests (paired, two-tailed, p < 0.05), under 0.125 mg of RP, the cathodal tDCS-elicited excitability diminution was abolished and differed significantly from the placebo medication condition, while the anodal tDCS-induced excitatory effect tended to be reversed to inhibition. Similarly, the excitatory and inhibitory effects of tDCS on cortical excitability were suppressed after 0.25 mg of RP. Compared with baseline MEPs, medium dosage RP (0.5 mg) selectively enhanced and prolonged the inhibitory effects generated by cathodal tDCS until the evening 1 d after tDCS. When a high dose of RP was administered, the anodal tDCS-elicited neuroplastic excitability enhancement was diminished and the cathodal tDCS-induced excitability reduction inversed into facilitation, which lasted up 30 min after DC stimulation (Fig. 3). Together, these results show that RP dose-dependently modified the tDCS-induced plastic excitability changes: whereas the low (0.125 and 0.25 mg) and high (1.0 mg) dosages of RP impaired both cathodal and anodal tDCS-induced neuroplasticity, compared with PLC conditions, the medium dosage did not influence the anodal tDCS-induced excitability enhancement, but prolonged the cathodal tDCS-generated inhibition, resulting in an inverted U-shaped dose–response curve (see Fig. 5).
Results of the ANOVAs conducted for experiment 1 (tDCS) and experiment 2 (PAS)

Dose-dependent effect of D2-like activation on nonfocal plasticity induced by anodal and cathodal tDCS (experiment 1). The time course plots show the effect of different doses of RP on tDCS-induced nonfocal neuroplasticity. Shown are baseline-standardized MEP amplitudes after plasticity induction by cathodal and anodal tDCS under 0.125, 0.25, 0.5, and 1.0 mg of RP or PLC up to the evening of the poststimulation day. As shown by TMS-elicited MEP amplitudes, RP produced a biphasic response, where low (0.125 and 0.25 mg) and high (1.0 mg) dosages impaired both cathodal and anodal tDCS-induced neuroplasticity, compared with the placebo (PLC) condition. Under medium dosage (0.5 mg), RP does not influence the anodal tDCS-elicited aftereffects. In contrast, a prolonged inhibition was observed in the cathodal tDCS condition. Shown are the mean ± SEM amplitudes vs baseline across time following anodal or cathodal tDCS for 0.125 (A), 0.25 mg (B), 0.5 (C), and 1.0 mg (D) of RP and PLC conditions. Filled symbols indicate significant deviations of the post-tDCS MEP amplitudes from baseline values, and “#” symbols mark significant deviations of drug versus PLC conditions with regard to identical time points and tDCS polarities (Student's t test, two-tailed, paired samples, p < 0.05). ne, Next evening; nm, next morning; na, next afternoon; se, same evening.
Dose-dependent effects of ropinirole on focal PAS-induced plasticity (experiment 2)
As shown by the results of the ANOVA, the interaction between “time course” and “PAS” was significant (Table 1). The missing significance of the remaining interactions is most probably caused by the independency of iPAS from RP dosage. After placebo medication intake, a significant enhancement of cortical excitability compared with baseline values was observed for ePAS and a significant inhibition for iPAS. These iPAS and ePAS-induced excitability shifts remained significant until 30 min after stimulation. RP had a significant effect on focal plasticity induced by PAS. Under 0.125 mg of RP, as revealed by the post hoc t tests (paired, two-tailed, p < 0.05), the ePAS-induced aftereffects were abolished. After administration of the medium dose of RP (0.5 mg), the excitability enhancement accomplished by ePAS was significant until 30 min after stimulation, and did not differ from the placebo condition. The high dosage of RP (1.0 mg), similarly to the low dose, abolished the ePAS-elicited excitatory effects. Conversely, the iPAS protocol decreased motor cortex excitability similarly independent of the drug condition. Therefore, as can be seen in the Figures 4 and 5, the dose–response curve for ePAS is inverted U-shaped. The efficacy of ePAS in inducing motor cortex excitability enhancements follows a nonlinear dose–response relationship. In contrast, for iPAS, no significant dose-dependent effect was observed (Fig. 5).

Dose-dependent effect of D2-like receptor activation on focal plasticity induced by PAS25 and PAS10 (experiment 2). The time course plots showing the effect of different doses of RP on PAS-induced focal neuroplasticity. Shown are baseline-standardized MEP amplitudes after plasticity induction by ePAS (ISI of 25 ms) and iPAS (ISI of 10 ms) under 0.125 (A), 0.5 (B), and 1.0 mg (C) of ropinirole up to the evening of the poststimulation day. Under placebo medication, ePAS enhances while iPAS diminishes excitability significantly for up to 30 min after stimulation. Under the medium ropinirole dosage, the PAS-generated excitability enhancement remains largely unaffected, whereas the high and low doses abolish this excitatory effect. For iPAS-induced excitability reduction, no significant dose-dependent effect was observed. A prolonged inhibition was observed until 60 min after stimulation in the iPAS condition under 0.125 mg. Filled symbols indicate significant deviations of the poststimulation MEP amplitudes from baseline with regard to each drug dose. The “#” symbols indicate significant differences of the drug versus PLC conditions with regard to identical time points (Student's t test, two-tailed, paired samples, p < 0.05). Error bars indicate SEM. ne, Next evening; nm, next morning; na, next afternoon; se, same evening.

Dose-dependent effect of D2-like receptor activation on nonfocal and focal plasticity induced by tDCS and PAS. D2-like receptor activation by ropinirole has an inverted U-shaped effect on neuroplasticity induced by tDCS and ePAS. High or low D2-like agonist dosage impaired plasticity. No dose-dependent alterations on iPAS-induced aftereffects were observed. Each column represents the mean of baseline-standardized MEP amplitudes ± SEM until 30 min after stimulation.
Discussion
Modulation of the D2-like receptor exerts a dose-dependent nonlinear effect on neuroplasticity in the human motor cortex, which differs for the type of plasticity induced.
The dose-dependent effect of D2-like receptor activation on plasticity
Under placebo medication, cathodal tDCS and iPAS lowered motor cortex excitability, whereas anodal tDCS and ePAS enhanced it. The duration of these aftereffects was equivalent to those in foregoing studies (Stefan et al., 2000; Nitsche and Paulus, 2001; Nitsche et al., 2003). RP alone did not affect cortical excitability significantly.
The main result of the current study is that dependent on dosage ropinirole results in an inverted U-shaped dose–response curve. Substantial literature points to an inverted U-shaped relation between cognitive performance and DA or D1 stimulation levels (Williams and Goldman-Rakic, 1995; Verma and Moghaddam, 1996; Zahrt et al., 1997; Muly et al., 1998; Robbins, 2000; Floresco and Phillips, 2001). Studies exploring the relationship between D2-like receptor activity and cortical function are rare. However, D2/D3 agonists have an inverted U-shaped effect on yawning behavior in rats (Collins et al., 2005). Our results suggest that not only DA, and D1-like receptor agonists exert such an inverted U-shaped response, but D2-like activation also produces such effects. In the current study, both too little and too much D2-like activation impaired nonfocal plasticity induced by tDCS and focal, ePAS-generated plasticity. These results argue for an optimal range of D2-like receptor activity for its adequate action on brain function.
Evidence for an optimal range of D2-like activation for induction and consolidation of plasticity
The results of the present study are indicative for a narrow range of optimal D2-like receptor stimulation for not interfering with neuroplasticity. Under a medium dosage, RP did not negatively affect plasticity. Together with the results of a former study, where it was shown that the D2-like receptor antagonist sulpiride abolishes the tDCS effects (Nitsche et al., 2006), this strengthens the evidence for an impact of D2-like receptors on nonfocal plasticity. These results are accordant to animal studies demonstrating that LTP and LTD are enhanced by D2-like receptor activation (Otani et al., 1998; Spencer and Murphy, 2000; Manahan-Vaughan and Kulla, 2003). Moreover, D2-like receptor activation produced a prolongation of the cathodal tDCS-induced inhibitory aftereffects, similar to a study performed with the predominant D2-like agonist pergolide (Nitsche et al., 2006), and animal experiments (Seamans and Yang, 2004). Also for ePAS-induced focal plasticity, the medium RP dosage does not induce strong changes, compared with placebo medication. For ePAS, this might be caused by a minor impact of D2-like receptors on this kind of plasticity by this dosage, because, in contrast to its effects on tDCS-induced plasticity, the D2-antagonist sulpiride does not extinguish ePAS-generated plasticity (Nitsche et al., 2009).
If so, why did RP low and high dosages have a plasticity-abolishing effect for nonfocal facilitatory and inhibitory plasticity as well as for facilitatory focal plasticity? One reason might be the capability of DA agonists for activating presynaptic and postsynaptic DA receptors (Yamada and Furukawa, 1980; Lomanowska et al., 2004; Millan et al., 2004). D2-like presynaptic dopamine receptors serve as autoreceptors, i.e., are self-regulatory, being able to regulate DA neurotransmission, modulate membrane excitability, and influence synthesis, release, and metabolism of DA (Schmitz et al., 2003). DA autoreceptor activation inhibits dopaminergic neurons (Wolf and Roth, 1990; Usiello et al., 2000; Centonze et al., 2003). It is possible that the D2-like agonist RP activated predominantly presynaptic or postsynaptic D2-like receptors depending on dosage. Indeed, low doses of the DA agonist apomorphine preferentially activate presynaptic dopamine receptors, which reduce the firing rate, synthesis, and release of DA by a negative feedback mechanism, whereas higher doses stimulate postsynaptic receptors (Yamada and Furukawa, 1980). Similarly, low doses of D2-like agonist quinpirole resulted suppress motor activity by decrease of DA release induced by presynaptic receptor activation (Lomanowska et al., 2004). Therefore we suggest that low-dose ropinirole in our study preferentially activated presynaptic DA autoreceptors, thus lowering extracellular dopamine concentration and reducing postsynaptic DA activity. Since DA is important for the induction of NMDA-dependent neuroplasticity, as elicited by tDCS and PAS (Stefan et al., 2002; Wolters et al., 2003; Nitsche et al., 2006; Kuo et al., 2008), this might explain the abolished plasticity under low-dose RP. Accordingly, Breitenstein et al. (2006) have suggested that D2-like autoreceptor activation by pergolide might cause impaired learning in humans. With a medium dosage of RP, presynaptic DA inhibition might be compensated for by postsynaptic D2-like receptor effects. In this regard, it is of importance that D2-like receptors reduce NMDA-receptor as well as GABAergic activation (Seamans and Yang, 2004). We hypothesize that the medium dosage-elicited deactivation of NMDA receptors reduces spontaneous background activity, but is not sufficiently strong to suppress stimulation-induced plasticity. Therefore, the plasticity-inducing effect of both, tDCS and PAS, might be preserved due to an enhanced signal-to-noise ratio accomplished by medium-dosed RP. Additionally reduced GABAergic activity might contribute to strengthen stimulation-induced plasticity. Indeed, D2-like receptor activation has been shown to be important for tDCS-induced plasticity before: inhibitory and facilitatory effects of tDCS are abolished by D2-block via sulpiride (Nitsche et al., 2006). However, under high-dosage D2-agonism, the inhibitory effect of D2-like receptor activation on postsynaptic membranes and NMDA receptors might suffice to suppress also stimulation-induced plasticity.
These assumptions can explain the effects of RP on tDCS and ePAS, but deliver no explanation for the missing effect of RP on iPAS. It might be suggested that D2-like activity is not important for this kind of plasticity. However, the D2-antagonist sulpiride abolishes iPAS-induced plasticity (Nitsche et al., 2009). Thus D2-like activation is important for, but not nonlinearly connected to focal inhibitory plasticity. Why do the effects of RP on focal, iPAS-induced, and nonfocal, cathodal tDCS-generated inhibitory plasticity differ? This dissociation of effects might be caused by the different kinds of plasticity induction accomplished. tDCS induces plasticity by slight modifications of spontaneous activity and postsynaptic membrane potential (Bindman et al., 1964; Purpura and McMurtry, 1965) and might thus be much more prone to slight changes of these factors accomplished by modifications of dopaminergic activity than iPAS, which exerts its effects by induction of timing-dependent suprathreshold presynaptic and postsynaptic activity.
Some potentially limiting aspects of the study should be mentioned: RP alone did not modulate MEPs significantly, but a slight trend for a reduction of MEPs under the highest RP dosage can be seen. In principle, this slight excitability diminution could have had an impact on subsequent plasticity induction. However, since the effect of RP on cortical excitability was minor, compared with excitability changes altering subsequent plasticity induction in former studies (Nitsche et al., 2007), this is improbable in our opinion. Subject blinding might have been compromised because of the different duration of monitoring the aftereffects and systemic side effects of RP in the different experimental sessions. However, because (1) only two subjects experienced side effects, and this only under the highest dosage of RP, (2) the multitude of sessions per subject makes it difficult to identify the specific experimental condition, and (3) the participants were not aware of proposed results, blinding might be somewhat compromised, but still be present. In principle, it cannot be ruled out that beyond focality of stimulation, PAS and tDCS differ by other factors, like neuronal pools affected by the stimulation protocols, and the specific effects of the stimulation protocols on pharmacology and spontaneous activity. However, we presume that the most important difference between tDCS and PAS is the difference of focality of stimulation. PAS is relatively focal because its effects are restricted to somatosensory afferents and intrinsic interneurons of the primary motor cortex (Stefan et al., 2000; Weise et al., 2006). In contrast, tDCS is thought to modulate also premotor afferences (Boros et al., 2008), and other neurons, including the proximal aspect of pyramidal tract neurons (Purpura and McMurtry, 1965; Ardolino et al., 2005).
General remarks
Here we demonstrate an inverted U-shaped effect of D2-like receptor modulation on neuroplasticity in humans. The identification of this nonlinear effect contributes to the understanding of how an inappropriate increase of dopaminergic drug dosage does provide not only no added benefit, but leads to a functional impairment. This knowledge might be useful to develop tailor-made drugs and optimize individual therapies for patients suffering from diseases affecting the dopaminergic system in future. However, further studies are needed to explore directly if the nonlinear effect of dopamine on cortical function is the same in patients as in the healthy subjects studied here.
Two interesting aspects of importance for future studies should be mentioned. (1) Genetic factors, for instance polymorphisms of the catechol O-methyltransferase, influence the impact of monoaminergic drugs on cognition in humans, and shift the individual dose–response relation between drugs and performance (Mattay et al., 2003). Thus it is possible that individual genetic variants may influence the specific impact of dopaminergic activity on plasticity. (2) The impact of dopaminergic drugs on cognition is age dependent. Increased age reduces dopaminergic signaling, and receptor density (Volkow et al., 1994; Flöel et al., 2005). Thus, the effect of RP on plasticity, as accomplished in the present study, might be specific for young healthy subjects, and differ in older subjects. Further studies for clarifying these aspects are necessary.
Footnotes
-
K.M.-S. is supported by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Brazil. This project was further supported by the Federal Ministry of Education and Research (BMBF 01GQ0432), Bernstein Center for Computational Neuroscience Goettingen, and the framework of the national priority program (Schwerpunktprogram) SPP1226 Nicotine: Molecular and Physiological Mechanisms in the Central Nervous System (www.nicotine-research.com) funded by the German Research Foundation (Deutsche Forschungsgemeinschaft, Grant NI 683/4-1).
- Correspondence should be addressed to Michael A. Nitsche, Department of Clinical Neurophysiology, Georg August University, Robert-Koch-Strasse 40, 37075 Göttingen, Germany. mnitsch1{at}gwdg.de





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}