
Abstract
Central respiratory chemoreceptors sense changes in CO2/H+ and initiate the adjustments to ventilation required to preserve brain and tissue pH. The cellular nature of the sensors (neurons and/or glia) and their CNS location are not conclusively established but the glutamatergic, Phox2b-expressing neurons located in the retrotrapezoid nucleus (RTN) are strong candidates. However, a direct demonstration that RTN neurons are intrinsically sensitive to CO2/H+, required for designation as a chemosensor, has been lacking. To address this, we tested the pH sensitivity of RTN neurons that were acutely dissociated from two lines of Phox2b-GFP BAC transgenic mice. All GFP-labeled cells assayed by reverse transcriptase-PCR (n = 40) were Phox2b+, VGlut2+, TH−, and ChAT−, the neurochemical phenotype previously defined for chemosensitive RTN neurons in vivo. We found that most dissociated RTN neurons from both lines of mice were CO2/H+-sensitive (∼79%), with discharge increasing during acidification and decreasing during alkalization. The pH-sensitive cells could be grouped into two populations characterized by similar pH sensitivity but different basal firing rates, as previously observed in recordings from GFP-labeled RTN neurons in slice preparations. In conclusion, these data indicate that RTN neurons are inherently pH-sensitive, as expected for a respiratory chemoreceptor.
Introduction
Central respiratory chemoreceptors subserve an important homeostatic reflex that acts to dynamically regulate breathing and CO2 excretion to maintain brain and tissue acid–base balance (Feldman et al., 2003; Nattie and Li, 2006). The definition of a central respiratory chemoreceptor continues to evolve as additional candidates are scrutinized. However, an inescapable premise is that a central chemoreceptor must be a cell that has the ability to detect changes in CO2 and/or pH. At present, the nature and number of these cells is not definitively established, and evidence suggests that both neurons and glia may possess this ability (Nattie and Li, 2006; Guyenet et al., 2010; Hodges and Richerson, 2010).
Candidate central respiratory chemoreceptors should present at least the following requisite properties: (1) intrinsic sensitivity to physiologically relevant changes in CO2/H+; (2) sensitivity to changes in CO2 that are readily demonstrable in vivo; (3) functional connectivity to respiratory centers, such that their selective activation, inhibition, or elimination produces the appropriate effect on the respiratory chemoreflex (Guyenet et al., 2010; Hodges and Richerson, 2010).
In the case of the glutamatergic, Phox2b-expressing neurons of the retrotrapezoid nucleus (RTN), all but the first of these criteria have been fulfilled (Mulkey et al., 2004; Stornetta et al., 2006; Guyenet et al., 2010). Inhibition or chemotoxic/genetic destruction of RTN neurons impairs respiratory CO2 sensitivity and raises the CO2 threshold for breathing (Dubreuil et al., 2008; Takakura et al., 2008; Marina et al., 2010), whereas their selective optogenetic stimulation drives central respiratory output in both anesthetized and conscious animals (Abbott et al., 2009; Kanbar et al., 2010). In addition, Phox2b-expressing RTN neurons respond avidly to elevated CO2/H+, both in vivo and in slices (Mulkey et al., 2004; Stornetta et al., 2006; Lazarenko et al., 2009, 2010). However, although pH-regulated excitability of RTN neurons is preserved during synaptic blockade (Mulkey et al., 2004; Lazarenko et al., 2009, 2010; Onimaru et al., 2012), these findings are not sufficient to conclude that pH sensitivity is intrinsic. Indeed, there is also evidence that at least some RTN neurons attain a fraction of their chemosensitivity indirectly, by effects of ATP released from local pH-sensitive astrocytes (Gourine et al., 2010; Wenker et al., 2010).
Here, we tested directly the intrinsic pH sensitivity of Phox2b-expressing RTN neurons following acute dissociation, under conditions that eliminate any extrinsic influences on cell properties that might be present in vivo or retained in brainstem slices. Our data indicate that the majority of isolated RTN neurons retain their characteristic sensitivity to CO2/H+, providing definitive evidence of an intrinsic pH sensitivity demanded for designation as a central respiratory chemoreceptor neuron.
Materials and Methods
Phox2b-GFP mice.
In this study, we used two lines of BAC transgenic mice in which GFP expression was directed to Phox2b-expressing neurons; the first line (B/G) was made on-site and the second line (Jx99) was prepared by the GENSAT project (www.gensat.org) (Lazarenko et al., 2009). The characteristics of GFP expression and pH sensitivity of RTN neurons were described previously for both lines (Lazarenko et al., 2009, 2010).
Acute dissociation of RTN neurons.
Transverse brainstem slices were prepared from neonatal (P6–P10) mice of either sex (Lazarenko et al., 2009, 2010). Briefly, mice were anesthetized with ketamine (375 mg/kg) and xylazine (25 mg/kg) intramuscularly, and rapidly decapitated; brainstems were removed and slices (350 μm) cut in the coronal plane in ice-cold sucrose-substituted Ringer's solution containing the following (in mm): 260 sucrose, 3 KCl, 5 MgCl2, 1 CaCl2, 1.25 NaH2PO4, 26 NaHCO3, 10 glucose, and 1 kynurenic acid, bubbled with 95% O2 and 5% CO2.
Slices were incubated for 15 min at room temperature in PIPES buffer (in mm): 120 NaCl, 5 KCl, 1 CaCl2, 1 MgCl2, 25 d-glucose, 20 PIPES, 100% O2 (Kay and Wong, 1986), and then for 60 min at 33°C in PIPES containing trypsin (Sigma-Aldrich, type XI; 0.5 mg/ml). After enzymatic treatment, slices were rinsed and maintained in PIPES buffer at room temperature for ∼60 min. Slices were transferred to DMEM buffer (Invitrogen, Life Technologies), and the RTN region was identified and excised under a fluorescence-equipped dissecting microscope (Zeiss Discovery V20). The tissue was triturated gently in DMEM buffer using a series of fire-polished Pasteur pipettes (600, 300, and 150 μm, inner diameter) and the DMEM/neuron suspension was placed in a recording chamber on a fixed-stage fluorescence microscope (Zeiss AxioExaminer).
Single-cell reverse transcriptase-PCR of dissociated RTN neurons.
Single-cell reverse transcriptase (RT)-PCR (scPCR) was performed on dissociated RTN neurons (Lazarenko et al., 2010). Individual GFP-fluorescent cells were aspirated into pipettes containing 10× RT buffer and RNaseOUT (Superscript III, Invitrogen) and expelled (∼1 μl) into sterile tubes containing dNTPs, BSA, RNaseOUT, MgCl2, oligo-dT, and random hexamers. The pre-RT mixture was incubated at 65°C, first-strand cDNA synthesis was performed with Superscript III reverse transcriptase, RNA was digested with RNase H, and cDNA was stored at −20°C. Two rounds of conventional PCR (GoTaq, Promega) used pairs of gene-specific, intron-spanning, outside and nested primer pairs. Primers for Phox2b, GAPDH, VGlut2, and glutamic acid decarboxylase (GAD1) were described previously (Lazarenko et al., 2010); primers were prepared for tyrosine hydroxylase (TH Fo: GCCGTCTCAGAGCAGGATAC; Ro: GGGTAGCATAGAGGCCCTTC; Fn: AGGAGAGGGATGGAAATGCT; Rn: ACCAGGGAACCTTGTCCTCT) and choline acetyltransferase (ChAT Fo: AATGGGTATGGAGCCTGTTA; Ro: AATGTCTTTGTGGATGTGGG; Fn: AGCCACCAACAGCAAAGGAA; Rn: GGATAGGGGAGCAGCAACAA). We included a no-template negative control for each experiment (H2O and/or bath solution); amplification of GAPDH mRNA served as a positive control.
Electrophysiological recordings.
Cell-attached patch recordings were obtained from GFP-fluorescent dissociated neurons (Perkins, 2006; Lazarenko et al., 2009, 2010) that were superfused with two solutions for testing pH sensitivity. The first used HCO3−-buffered solution, containing the following (in mm): 130 NaCl, 3 KCl, 2 MgCl2, 2 CaCl2, 1.25 NaH2PO4, 26 NaHCO3, and 10 glucose; it was bubbled with 95% O2 and 5% CO2 to attain a pH of 7.3. For acidification to pH 7.1, the CO2 was raised to 8% (balance O2) and for alkalization, the HCO3− was raised to 45 mm (substituted for NaCl, and bubbled with 95% O2/5% CO2). The second was a HEPES-based buffer that contained the following (in mm): 140 NaCl, 3 KCl, 2 MgCl2, 2 CaCl2, 10 HEPES, 10 glucose; pH was adjusted between 7.0 and 8.0 by addition of HCl or NaOH. Recordings were performed at room temperature under voltage clamp at a holding potential of −60 mV (Perkins, 2006). Patch electrodes (3–6 MΩ) were filled with (mm): 120 KCH3SO3, 4 NaCl, 1 MgCl2, 0.5 CaCl2, 10 HEPES, 10 EGTA, 3 Mg-ATP, 0.3 GTP-Tris, pH 7.2. Recordings were obtained using pCLAMP, a MultiClamp amplifier, and Digidata 1440A analog-to-digital converter (Molecular Devices). Firing rate histograms were generated by integrating action potential discharge in 10 s bins using Spike2 software.
Data analysis.
We determined pH sensitivity of individual RTN neurons by plotting firing rate versus bath pH and calculating the pH value at which firing rate was reduced to half of that obtained at pH 7.0 (pH50) using linear regression analysis (Microsoft Excel). The frequency distribution plot of pH50 values was fitted with a double Gaussian fit using Prism software (GraphPad). All data are expressed as mean ± SE. Statistical significance was determined using Student's t test, with a significance level of p < 0.05.
Results
We performed experiments on dissociated neurons from two BAC-transgenic lines in which GFP is expressed under the control of the Phox2b promoter. In the Jx99 line, GFP is expressed in all brainstem Phox2b-expressing neurons including, but not limited to, those within the RTN (Lazarenko et al., 2009). By contrast, GFP expression is restricted to the RTN in the rostral ventrolateral medulla of the B/G line, and therein, only to a subset of Phox2b-expressing RTN neurons (∼38%) (Lazarenko et al., 2009). In brainstem slice preparations, from both lines of mice, nearly all GFP-positive neurons in the RTN are sensitive to changes in extracellular pH (∼95%) (Lazarenko et al., 2009, 2010). In this work, we tested whether this chemosensitivity is retained in acutely dissociated GFP-expressing RTN cells.
Acutely dissociated and neurochemically defined RTN neurons are sensitive to CO2
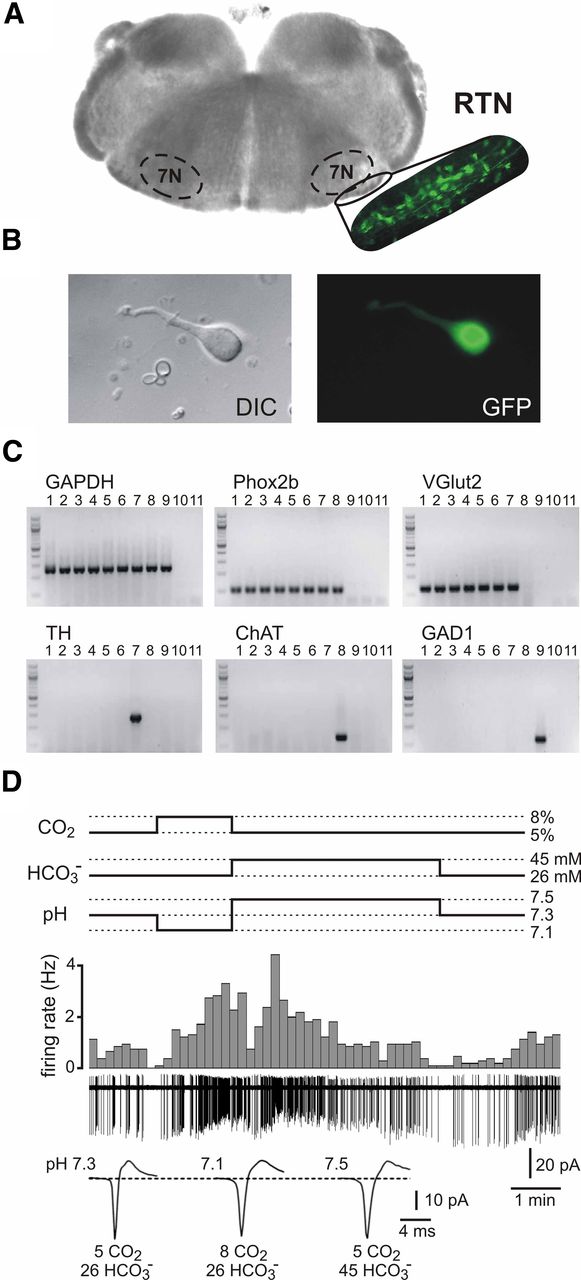
For dissociation of RTN neurons from the Jx99 line of mice, we identified the compact cluster of GFP-expressing cells ventral to the facial nucleus in brainstem slices and excised this region for enzymatic dissociation (Fig. 1A). We targeted small, fusiform-shaped GFP-labeled neurons for cell-attached recording of firing activity (Fig. 1B). This dissociation and selection procedure effectively identified neurochemically defined chemosensitive RTN neurons (Stornetta et al., 2006) because all GFP-expressing dissociated neurons examined by scPCR (n = 40) expressed Phox2b and the glutamate transporter, VGlut2 (Fig. 1C), including a subset of functionally characterized neurons assayed after recording (n = 7). On the other hand, none expressed TH or ChAT, markers for nearby Phox2b (and GFP)-expressing C1 adrenergic cells and facial motoneurons (Fig. 1C). Moreover, as described below, we find the majority of dissociated neurons show pH sensitivity typical of GFP-expressing RTN neurons identified previously in brainstem slices (Lazarenko et al., 2009, 2010).

RTN neurons dissociated from Jx99 mice are sensitive to CO2. A, Brainstem slice from Jx99 Phox2b-GFP transgenic mouse (left), with magnified fluorescent image of the RTN region (right); the facial nucleus is demarcated (7N). Note the GFP-labeled cells in the RTN region that was microdissected for dissociation. B, Image of a GFP-expressing dissociated RTN neuron under DIC (left) and fluorescence (right). C, Single-cell RT-PCR data revealed expression of Phox2b and VGlut2 in GFP-expressing dissociated RTN neurons (lanes 1–6), including three pH-sensitive neurons examined after recording (4–6); none of those RTN neurons expressed TH, ChAT, or GAD1. Control experiments verified detection of the following: TH (with Phox2b and VGlut2) in C1 neurons (lane 7); ChAT (and Phox2b) in motoneurons (lane 8), and GAD1 in striatal medium spiny cells (lane 9); GAPDH expression was seen in all cells, and negative controls included bath solution (lane 10) and water (lane 11) substituted for cell contents in all reactions. D, Firing activity of a GFP-expressing RTN neuron from Jx99 mouse during changes in pH produced by altering CO2 and HCO3−. Firing rate histograms (top traces) derived from cell-attached voltage-clamp recordings (bottom traces). Expanded time scale traces of action current waveforms (average of 30–50 spikes). Firing increased during CO2-induced bath acidification and decreased during HCO3−-induced bath alkalization.
An example of the pH sensitivity of a dissociated RTN neuron from the Jx99 line is shown in Figure 1D. This neuron fired at ∼0.8 Hz in a standard bath solution that was bubbled with 95% O2/5% CO2 and buffered to an extracellular pH of 7.3 using 26 mm HCO3−. The firing rate of the cell accelerated by ∼300% (to ∼2.5 Hz) when CO2 was increased to 8% (i.e., which lowered pH to 7.1) and the cell was nearly silenced (0.1 Hz) when exposed to a 5% CO2 solution that contained 45 mm HCO3− (i.e., which raised pH to 7.5). Thus, dissociated Phox2b-expressing RTN neurons from Jx99 mice are sensitive to changes in extracellular pH elicited by manipulating CO2 and HCO3− levels (n = 8/9), a characteristic we described previously for these cells recorded either in vivo or in brainstem slice preparations (Mulkey et al., 2004; Stornetta et al., 2006).
Dissociated RTN neurons retain type I and type II characteristics
In earlier recordings of GFP-expressing RTN neurons in brainstem slices from Phox2b-GFP mice, we uncovered two populations of pH-sensitive neurons that differed in the pH range over which they were active: by comparison to type I neurons, type II cells displayed higher firing rates at physiological pH and were silenced at more alkaline pH (Lazarenko et al., 2009, 2010). In this population of dissociated GFP-positive and pH-sensitive neurons (n = 26/34), we also observed these two types of cells (Fig. 2). As depicted in Figure 2A, a representative type I neuron fired at ∼2 Hz in a HEPES-based bath solution at pH 7.3, increased its firing rate to ∼3.5 Hz in pH 7.0, and was silenced at pH 7.5. A dissociated RTN neuron with characteristics of a type II cell, shown in Figure 2B, fired at ∼5 Hz under control conditions at pH 7.3; action potential discharge decreased progressively with increasing alkalization and was nearly fully suppressed only at pH 8.0, with a return to pH 7.0 evoking a strong increase in firing rate (to ∼8 Hz).

Type I and type II firing properties are retained in dissociated GFP-expressing RTN neurons. A, A relatively low level of baseline firing in a type I RTN neuron (<2 Hz) was increased by acidification (from pH 7.3 to pH 7.0) and essentially eliminated by alkalization to pH 7.5. B, In a type II cell, a comparatively high initial firing rate (∼5 Hz) was progressively suppressed with stepwise increases in bath alkalization and nearly silenced at pH 8.0. C, Frequency distribution of pH50 values fitted with a biphasic distribution with peaks at pH 7.3 and 7.6; individual neurons were designated as type I (pH50 < 7.42) or type II (pH50 > 7.42). D, Averaged pH sensitivity for dissociated Jx99 RTN neurons classified as type I (open circles) or type II neurons (solid circles). The pH sensitivity (i.e., the slope) was similar for both cell types, but type II neurons displayed higher firing rates at all pH levels.
We calculated a pH50 value for each cell (i.e., the pH at which firing frequency was reduced to half that observed at pH 7.0) and found a bimodal frequency distribution that could be approximated by two Gaussian distributions, with peaks at pH50 = 7.3 and pH50 = 7.6 (Fig. 2C). This was similar to the distributions observed in our earlier recordings of pH-sensitive GFP-labeled RTN neurons in brainstem slices (Lazarenko et al., 2009, 2010). Based on this distribution, we chose a cutoff pH50 of 7.42 to classify individual dissociated cells as type I or type II and plotted the relationship between firing rate and pH for each group (Fig. 2D). As observed for GFP-expressing RTN neurons in slices (Lazarenko et al., 2009, 2010), both groups displayed a similar pH sensitivity (i.e., slope), but type II cells were characterized by a higher firing rate at all pH levels, with greater alkalization required for complete suppression of firing activity.
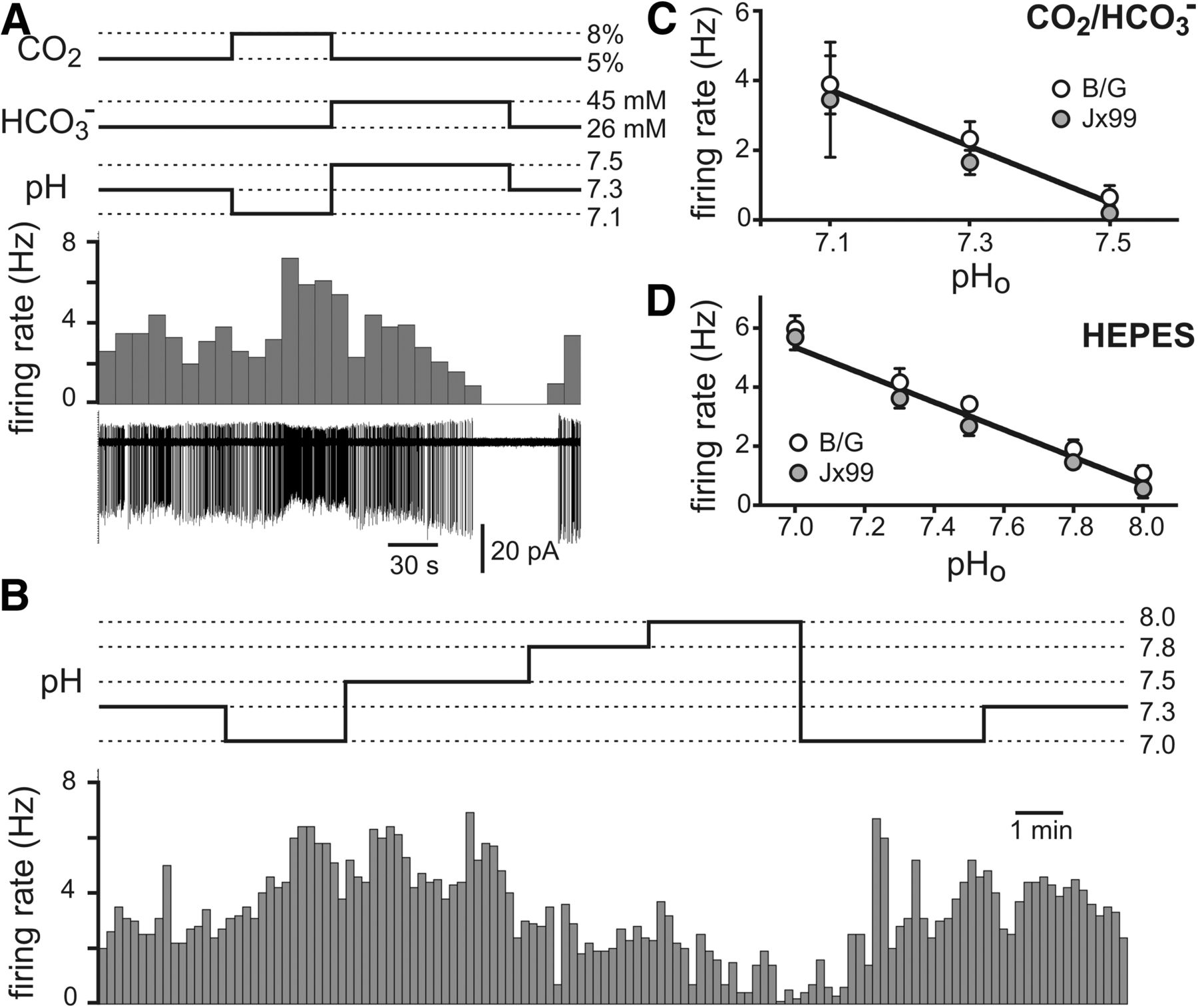
RTN neurons dissociated from B/G and Jx99 mice have essentially identical pH-sensitive firing properties
The microdissection and cell selection procedure we used to identify neurons from the Jx99 line of mice appears to have favored a sampling of chemosensitive RTN neurons, with appropriate neurochemical phenotype. However, we found that ∼21% of the recorded neurons were insensitive to pH, a higher fraction than observed previously by sampling GFP-expressing RTN neurons in brainstem slice preparations from Jx99 mice (Lazarenko et al., 2009). To increase confidence that recordings were indeed from RTN neurons, we used the B/G line of mice in which GFP-expressing neurons in this region are found only in the RTN. The yield of cells obtained from these preparations was limited (n = 13) because GFP is only present in some Phox2b-expressing RTN neurons of B/G mice. Nevertheless, we found that ∼23% of GFP-expressing cells from the B/G mice were insensitive to pH, similar to that seen in the Jx99 line. Moreover, there was no difference in initial firing rates between pH-sensitive and pH-insensitive cells (at pH 7.3 in HEPES: 3.7 ± 0.3 Hz vs 4.2 ± 0.5, n = 33 and 9 for both Jx99 and B/G lines, p = 0.38). This suggests that misidentification of RTN neurons is unlikely to account for the few pH-insensitive dissociated neurons observed with either line.
The properties of the pH-sensitive, GFP-expressing dissociated neurons we recorded from the B/G line of mice were also essentially identical to those obtained from the Jx99 line. Thus, we found GFP-expressing RTN neurons that altered their firing rate during changes in extracellular pH evoked by altering CO2/HCO3− or fixed acid in a HEPES-based buffer (Fig. 3A,B). The limited sample size precluded a population analysis of type I and type II cells from B/G mice. However, direct comparison of averaged firing rate responses to changes in pH from the entire pool of GFP-labeled RTN neurons, depicted in Figure 3C for CO2/HCO3−-based bath solutions and in Figure 3D for HEPES-based bath solutions, indicates that pH-dependent firing properties of GFP-expressing neurons from the Jx99 and B/G lines are essentially identical.
{kind=link}
{kind=link}
{kind=link}
Dissociated RTN neurons from B/G and Jx99 mice have identical pH-sensitive firing properties. A, Firing rate histogram (top) and cell-attached recording of firing activity (bottom) from a GFP-expressing RTN neuron from B/G mouse during pH changes evoked by altering CO2 and HCO3− levels. B, Type II, pH-dependent firing behavior recorded in an RTN neuron from the B/G mouse line. C, D, Averaged pH sensitivity of firing for all dissociated GFP-labeled RTN neurons from B/G (open circles) and Jx99 (solid circles) mice recorded in CO2/HCO3− buffer (C) and HEPES buffer (D). The pH sensitivity was not different in cells from the two lines of mice, regardless of buffer conditions.
Discussion
The identity of the specific group(s) of brainstem neurons that sense changes in CO2/H+ for purposes of regulation of ventilation remains controversial, in part because all the key characteristics required for definitive designation of a neuron as a central respiratory chemoreceptor have not been met for any candidate cell type (Guyenet et al., 2010; Hodges and Richerson, 2010). In this work, we demonstrated that Phox2b-expressing RTN neurons retain their pH sensitivity after acute dissociation, establishing that these putative respiratory chemoreceptor cells are indeed intrinsically sensitive to CO2/H+. Thus, these data indicate that RTN neurons possess this property required of a central chemosensor (Hodges and Richerson, 2010).
We found that pH response characteristics of Phox2b-expressing RTN neurons, previously described in brainstem slices (Lazarenko et al., 2009, 2010), were well preserved after dissociation. Notably in this respect, the firing of GFP-labeled RTN cells isolated from Phox2b-GFP mice was sensitive to extracellular acidification and alkalization, whether evoked by changes in CO2/HCO3− or by changes in fixed acid. Indeed, the pH sensitivity of these dissociated GFP-expressing RTN cells (∼5 Hz/pH in HEPES; ∼8 Hz/pH in CO2/HCO3−) was essentially identical to that we recorded previously from RTN neurons in slice preparations (∼6 Hz/pH in HEPES) (Guyenet et al., 2005; Lazarenko et al., 2009), and also similar to the most sensitive of other presumptive chemosensor cell types recorded under similar conditions in vitro (Wang et al., 1998, 2001; Putnam et al., 2004). In addition, two populations of dissociated RTN neurons with similar pH sensitivity but differences in baseline firing rates could be readily discerned, as described for these GFP-labeled cells in slices (Lazarenko et al., 2009, 2010). This latter observation suggests that the differences in baseline excitability that define these two populations of RTN neurons also reflect properties intrinsic to the cells. The intrinsic ionic mechanisms that account for the shared pH sensitivity and the distinct baseline firing properties in chemosensitive Phox2b-expressing RTN neurons remain to be determined.
We found a number of dissociated GFP-labeled neurons that were not demonstrably sensitive to pH (∼21%), a contrast with the nearly universal pH sensitivity observed for GFP-expressing RTN neurons in slices from Phox2b-GFP mice (Lazarenko et al., 2009, 2010). Although it is possible that some pH-insensitive neurons were derived from other nearby Phox2b-expressing cell groups (e.g., C1 adrenergic neurons) (Stornetta et al., 2006), we think this is unlikely. First, all dissociated GFP-expressing neurons from Jx99 mice assayed by scPCR presented the neurochemical signature identified for chemosensitive RTN neurons in vivo (i.e., Phox2b+, VGlut2+, TH−, ChAT−) (Stornetta et al., 2006), including one pH-insensitive cell. Moreover, we obtained a similar fraction of pH-insensitive neurons from the B/G line of mice in which only RTN neurons in this region express GFP. Another possibility is that the pH-sensing mechanism is present on neuronal processes that are occasionally lost during the isolation procedure. In this regard, RTN neurons possess dendrites that extend along the ventral medullary surface (Mulkey et al., 2004; Stornetta et al., 2006) and which may survive cell isolation unevenly. A third possibility is that RTN contains a subpopulation of Phox2b-expressing neurons that are not inherently pH-sensitive. In this respect, pH-dependent release of ATP from astrocytes may contribute to an indirect form of neuronal chemosensitivity within the RTN (Gourine et al., 2010; Wenker et al., 2010), and such a mechanism would be lost in this dissociated neuronal preparation.
By way of caveat, RTN neuronal responses to changes in CO2/H+ in vitro are a fraction of those observed for the same RTN neurons examined in vivo (∼35%, when corrected to 37°C, 17 Hz/pH vs 48 Hz/pH) (Guyenet et al., 2005, 2012). This may reflect, in part, the underdeveloped respiratory chemoreflex in neonatal animals commonly used for in vitro studies. Other factors in addition to temperature and age may also account for these discrepancies (e.g., loss of neuromodulatory influences in vitro), precluding direct quantitative comparisons of in vitro neuronal responses to those of the entire respiratory network in vivo (Putnam et al., 2004; Guyenet et al., 2012). Finally, other lines of evidence suggest that glial cell-derived ATP contributes to CO2 sensitivity of RTN neurons, even in neonates (Gourine et al., 2010; Wenker et al., 2010). The relative importance of this purinergic paracrine mechanism versus the direct chemosensitivity uncovered in the current experiments is uncertain, but the convergence of both mechanisms provides an attractive explanation for the exquisite CO2 sensitivity of RTN neurons in vivo.
In conclusion, an expanding dataset implicates RTN neurons as important central respiratory chemoreceptors (Guyenet et al., 2010). For example, optogenetic activation of RTN neurons in vivo stimulates breathing in a manner that is mutually occlusive with CO2 (Abbott et al., 2009). Conversely, selective chemotoxic lesions depress breathing and raise the CO2 apneic threshold (Takakura et al., 2008), and inhibition of RTN neurons in conscious rats eliminates up to 65% of the chemoreflex (Marina et al., 2010). Moreover, Phox2b-expressing RTN neurons are uniquely destroyed in mouse models of human central congenital hypoventilation syndrome, in which respiratory chemosensitivity is absent at birth and recovers only modestly thereafter (Dubreuil et al., 2008; Ramanantsoa et al., 2011). Importantly, RTN neuronal firing activity is sensitive to CO2/H+, both in vitro and in vivo (Mulkey et al., 2004; Guyenet et al., 2005; Stornetta et al., 2006; Lazarenko et al., 2009, 2010), and this work now demonstrates that CO2/H+ sensitivity is a property that is inherent to RTN neurons.
Footnotes
- Received December 2, 2012.
- Revision received March 20, 2013.
- Accepted March 27, 2013.
This work was supported by NIH Grants HL074011 (P.G.G.) and HL108609 (D.A.B.).
- Correspondence should be addressed to Dr. Douglas A. Bayliss, University of Virginia School of Medicine, Department of Pharmacology, 1300 Jefferson Park Avenue, Jordan Hall Room 5013, Charlottesville, VA 22908-0735. bayliss{at}virginia.edu
- Copyright © 2013 the authors 0270-6474/13/337756-06$15.00/0