

Abstract
Adaptation is typically associated with attenuation of the neuronal response during sustained or repetitive sensory stimulation, followed by a gradual recovery of the response to its baseline level thereafter. Here, we examined the process of recovery from sensory adaptation in layer IV cells of the rat barrel cortex using in vivo intracellular recordings. Surprisingly, in approximately one-third of the cells, the response to a test stimulus delivered a few hundred milliseconds after the adapting stimulation was significantly facilitated. Recordings under different holding potentials revealed that the enhanced response was the result of an imbalance between excitation and inhibition, where a faster recovery of excitation compared with inhibition facilitated the response. Hence, our data provide the first mechanistic explanation of sensory facilitation after adaptation and suggest that adaptation increases the sensitivity of cortical neurons to sensory stimulation by altering the balance between excitation and inhibition.
Introduction
Adaptation is a prevalent property of sensory systems across different species and modalities and is typically associated with an attenuation of neural activity in response to sustained or repetitive stimulation. Although the precise functional role of adaptation is unclear, it is thought to play a fundamental role in information processing (Wark et al., 2007) and in behavioral habituation (Gutfreund, 2012). Adaptation allows optimal response tuning to particular aspects of the stimulus by shifting the input–output relations of sensory circuits even when the overall conditions of the sensory environment are changing (Adorjan et al., 1999; Muller et al., 1999; Maravall et al., 2007). In addition, by suppressing responses to familiar stimuli, adaptation enhances the detection of novelty (Dragoi et al., 2002; Ulanovsky et al., 2003). These views support the notion that adaptation acts to suppress neuronal responses when stimulation persists.
The degree of adaptation and the time course for its recovery are very dynamic (Kohn, 2007) and may change across different cell types and in a manner that depends on both the intensity and duration of the adapting stimulation (Bonds, 1991; Greenlee et al., 1991; Nelson, 1991; Muller et al., 1999; Fairhall et al., 2001; Lundstrom et al., 2008; Wark et al., 2009; Abolafia et al., 2010; Ganmor et al., 2010; Lundstrom et al., 2010). Interestingly, psychophysical studies have shown that periodic tactile stimulation enhances human performance in amplitude, frequency, and spatial discrimination tasks when a test stimulus closely follows repetitive stimulation (Goble and Hollins, 1993, 1994; Harris et al., 2002; Tannan et al., 2006). This implicates a role for adaptation in increasing rather than suppressing the sensitivity of the somatosensory system to a test stimulus presented after adaptation.
Here, we investigated the time course of neuronal recovery from adaptation in the rodent's vibrissal system. This system serves as a model for studying neuronal processes in sensory systems in general and in the somatosensory system in particular (Petersen, 2007). Rats gather tactile information by sweeping their whiskers against objects at a rate of 5–20 Hz (Carvell and Simons, 1990; Dassonville, 1995). Within this frequency range, neuronal cells in the barrel cortex exhibit slow and quite substantial adaptation (Ahissar et al., 2000; Khatri et al., 2004; Katz et al., 2006; Heiss et al., 2008; Ganmor et al., 2010), which probably results from short-term depression of thalamocortical synapses (Chung et al., 2002; Katz et al., 2006).
In whole-cell patched neurons from layer IV and its vicinity in the barrel cortex of anesthetized rats, we studied neuronal responses to a sensory stimulant after repetitive whisker deflection. Surprisingly, despite the adaptation induced by repetitive stimulation of the principal whisker, we found not only that the response to subsequent whisker deflection recovered shortly afterward, but also that ∼30% of the neurons exhibited a significantly enhanced response during the first few hundred milliseconds after adaptation. By recording the cells under different holding currents, we found that the facilitated test response was associated with delayed recovery of inhibition relative to excitation.
Materials and Methods
Animal preparation.
All procedures involving animals were reviewed and approved by the Weizmann Institutional Animals Care Committee. Animal surgeries and in vivo recordings were performed as previously described (Katz et al., 2006). Briefly, recordings were made on young adult Wistar female rats (4–7 weeks old). For intracellular recording from the barrel cortex, after initial anesthesia with ketamine (100 mg/kg) and xylazine (10 mg/kg), a tracheotomy was made and the animals were mounted in a stereotaxic device and artificially respirated with a mixture of halothane (0.5–1%) and oxygen-enriched air. In a few experiments (6 cells recorded from 3 animals), isofluorane (0.5–1.5%) was used. As no qualitative differences were found, these cells were pooled together with the rest of the recorded neurons. A craniotomy (∼1 mm in diameter) was made above the barrel cortex (centered 2.5 mm posterior and 5.5 mm lateral to the bregma) and a portion of the dura mater was carefully removed. For intracellular thalamic recordings, animals were maintained under ketamine anesthesia (50 mg/kg, administrated at a rate of 0.3 ml/h). The levels of end-tidal CO2 and heart rate (250–450 beats/min) were monitored throughout the experiments. Body temperature was kept at 37°C using a heating blanket and rectal thermometer.
Cortical patch recordings.
Borosilicate micropipettes were pulled to produce electrodes with a resistance of 4–10 MΩ when filled with an intracellular solution containing (in mm) as follows: 136 K-gluconate, 10 KCl, 5 NaCl, 10 HEPES, 1 MgATP, 0.3 NaGTP, and 10 phosphocreatine (310 mOsm). For experiments in which current was injected to reveal inhibitory synaptic potentials, QX-314 (2 mm) was added to prevent action potentials. Intracellular signals were acquired using an Axoclamp-900A amplifier (Molecular Devices) and low passed at 3 kHz before being digitized at 10 kHz.
Thalamic recordings.
Intracellular recordings were performed using sharp electrodes filled with 2 m K-acetate and 20 mm QX-314 with a resistance of 60–100 MΩ. The craniotomy was centered 3 mm lateral and 3 mm posterior of the bregma over the ventral posteromedial nucleus (VPM). Signals were amplified using an Axoclamp-2B amplifier, low passed at 3 kHz, and digitized at 20 kHz.
Whisker stimulation and protocols.
Whiskers were trimmed to a length of 10–20 mm. The principal whisker was inserted into a 2-mm-plastic cone glued to a needle attached to the piezoelectric wafer (T220-H4–203Y; Piezosystems), which was driven by a homemade controller. Whisker displacement near the tip of the needle was measured off-line using an optical displacement measuring system (optoNCDT 1605; Micro-Epsilon). A fast-rising voltage command was used to evoke a fast whisker deflection with a constant rise time of ∼1 ms followed by a 20 ms ramp-down signal. Because of the fixed rise time, amplitude and speed of deflection grow together with the magnitude of the voltage command, following a quasi-linear relationship. The stimulation velocity and the corresponding deflection amplitude (15–65 mm/s, 15–75 μm amplitude) were adjusted to evoke clear subthreshold responses in the cortical cells.
In each trial, a train of 10 stimuli at 20 Hz (50 ms interstimulus interval) was applied. The final stimulus in the train was followed 50–1250 ms later by a single test stimulus (Fig. 1A).
Data analysis.
The recordings were analyzed using custom software written in MATLAB (The MathWorks). Postsynaptic potential (PSP) amplitude was measured as the difference between the peak membrane potential response and the mean baseline value obtained over the 10 ms before stimulation. Spike counts were calculated as the sum of spikes observed during the 5–45 ms period after whisker stimulation. For neurons in which QX-314 was not used, subthreshold responses were calculated after digital removal of the spikes, which were replaced with a spline interpolation based on the 5 samples on each side of the action potential.
As explained above, for each cell, we applied a test stimulus at different time intervals after termination of the adapting stimuli and selected the time interval in which the test response was the largest. To prevent bias in determining whether the largest test response (of 7–12 others) was higher than the control response, we used the bootstrap method to calculate the average response to the control stimulus. Briefly, all responses to the first deflection of the adapting train stimuli (“control stimulus”) were pooled (e.g., the 140 values that were taken from 20 repeats of seven test trials). From this set, we randomly drew (with repetitions) seven equal groups, and their means were computed. The largest mean was selected before performing a new iteration of 500 values. The largest average test response was considered significant if its value exceeded the confidence limits of the average control response, computed from the 500 values selected. To establish the significance level when comparing two sets of magnitudes, we used either the Wilcoxon signed rank test for paired data or the Mann–Whitney U test (Wilcoxon rank–sum test) for nonpaired values. A p value of <0.05 was considered to indicate a statistically significant difference. Based on the above measurements, a recovery index was calculated as follows:
 where Rt is the test response and Rc is the control response.
where Rt is the test response and Rc is the control response.
Except where otherwise indicated, data are presented as mean ± SEM.
Results
Facilitation of sensory response after an adapting stimulus
Intracellular recordings were obtained from 29 neurons in the primary somatosensory cortex of anesthetized rats. The average recording depth was 878 ± 22 μm from the pia surface (range, 760–1200 μm), well within layer 4 and its vicinity (Brecht and Sakmann, 2002; Heiss et al., 2008). The principal whisker was mechanically stimulated at 20 Hz (adapting stimulus), with the first stimulation in the train considered the control stimulus. The test stimulus had the same shape and amplitude of the adapting stimuli and was delivered 50–1250 ms after they were completed.
The average responses of a neuron to a test stimulus presented at four different time intervals after the repetitive train are presented in Figure 1A, B. As expected from previous studies, repetitive stimulation induced profound adaptation of both the synaptic response (Fig. 1A) and the firing response (Fig. 1B) of the cell. For this cell, the sensory response fully recovered 250–350 ms after the adaptive train (Fig. 1C,D). For each recorded neuron, a recovery index was calculated at each time interval by subtracting the average amplitude of the control response from the average amplitude of the synaptic response to the test stimulation and dividing the difference by the sum of both (Fig. 1E; see Materials and Methods). The time course of this index, when averaged across the population (Fig. 1E, black circles), displayed a fast response recovery after adaptation and was similar to that of the cell shown in Figure 1A–D. However, when looking at the entire time course of each neuron separately (Fig. 1E, colored circles), we found that, in some cells, the recovery index at particular time intervals was >0, indicating that the test response exceeded the control response and suggesting that, for some of the neurons, the response was facilitated. For different neurons, the facilitation occurred at different time intervals after termination of the adapting stimuli.

Rapid recovery of sensory response after adaptation. A, An average subthreshold response of a neuron during 20 Hz adapting whisker stimulation (vertical black lines) followed, at four different time intervals, by a single test whisker stimulus (vertical gray line). Spikes were removed digitally before the data were averaged. B, Corresponding average spike peristimulus time histograms extracted from the same neuron as shown in A. Bin size, 10 ms. C, Average peak voltage of the response to a test stimulus delivered at different time intervals after termination of the adapting stimuli (mean ± SEM, the same cell as A; recovery time was measured from the last stimulus of the adapting train). D, Average spike count (sum of spikes in a window of 5–45 ms after whisker stimulation) calculated for control whisker stimulation (gray line) and test stimulation delivered at different time intervals after the adapting train. E, Recovery indices for the population average (black circles) and for all cells (colored circles) calculated for a test stimulus delivered at different time intervals after the adapting train.
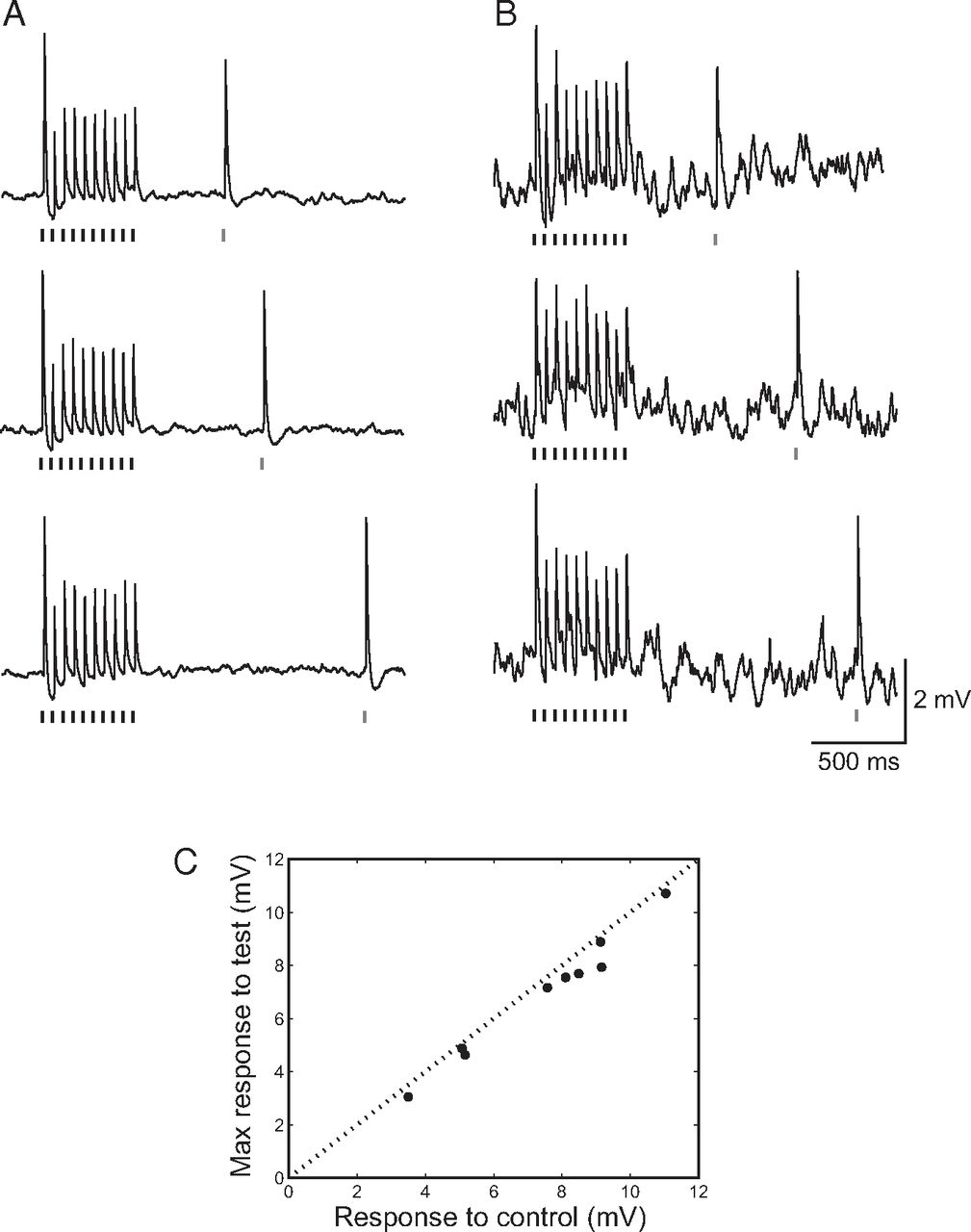
Figure 2 presents two cells that demonstrated a significantly facilitated response to a test stimulus after repetitive stimulation of the principal whisker. Facilitation at particular time intervals was clearly evident both in the subthreshold responses (increases of 109% and 21%, as shown Figure 2A and Figure 2E, middle rows, p = 8*10−12 and p = 0.005, respectively) and in the peak spike count (reaching an increase of 119% and 42% for the time intervals showed in Figure 2B and Figure 2F, respectively). In neurons where a facilitated response to the test stimulation was found, the time interval for maximal facilitation was, in most cases, the same for both the subthreshold responses and the spike count. See Figure 2C, D and Figure 2G, H for the response curves of the neurons illustrated in Figure 2A, B and Figure 2E, F, respectively. Test responses peaked at different times for these two cells as well as for others (Fig. 1E), and this may explain why the population-averaged data showed no clear peak at a particular time interval after the adapting stimulation (Fig. 1E).

Postadaptation facilitation of sensory response in the barrel cortex. A, E, Average subthreshold responses of two neurons during 20 Hz adapting whisker stimulation (vertical black lines) followed by delivery of a single test whisker stimulus (vertical gray line) at three different time intervals and their corresponding average peristimulus time histograms (B, F; bin size, 10 ms). A clearly facilitated response to the test stimulus was found in these cells (middle rows). Subthreshold data were averaged after digital removal of spikes from the raw traces. C, G, Average peak voltage of the subthreshold response to test stimuli delivered at different time intervals after termination of the adapting stimulus (mean ± SEM) and average spike count (sum of spikes in a window of 5–45 ms after whisker stimulation) calculated for control whisker stimulation (gray line) and test stimulation delivered at different time intervals (D, H). The data correspond to the presented cells.
We compared the control response with the maximal test response, rather than with the response at a fixed recovery time, because of the variability in the time at which maximum facilitation occurred in different neurons (Fig. 3A; n = 29). To prevent any statistical bias when comparing the maximal test response with the control response, the control response was calculated using bootstrap analysis (see Materials and Methods). Our analysis indicated that, for 62% (18 cells) of the cells, the maximal recovery response was larger than the corresponding control response, and a significant facilitation was found in 34% of all the tested cells (n = 10, Fig. 3A, black circles). For these cells, the average facilitation of the synaptic potential was 29 ± 10%. Facilitation was also found for the average spike count in response to the test stimulus (Fig. 3B) in neurons recorded either intracellularly (n = 12, subpopulation of that presented in Fig. 3A) or extracellularly (n = 5). The average maximal number of evoked spikes in response to the test stimulus (calculated using bootstrap analysis) increased by 44 ± 20% (n = 17, p = 0.006) compared with the control. The lower number of cells recorded intracellularly in Figure 3B compared with Figure 3A reflects the fact that, in 10 cells, the recording was performed in the presence of QX-314 to prevent neuronal firing and that 7 additional cells were excluded because they exhibited a very low firing rate in response to whisker stimulation (<0.1 spike/trial).

Population data of postadaptation facilitation of sensory response in cortical neurons. Average maximal PSP (A) and spike count (B) in response to test stimulation plotted against the corresponding average control responses (see Materials and Methods). C, Facilitation occurs over a wide range of time intervals after adapting stimulation. The recovery index for recorded neurons plotted against the time interval during which it was maximal. A, C, Cells whose PSP responses were significantly larger than their control response are marked in black. D, Correlation between the magnitude of facilitation (recovery index) and the degree of adaptation (adaptation index).
The average time interval from termination of the adapting stimulus until maximal subthreshold facilitation was ∼700 ms (Fig. 1E). However, upon examining this phenomenon in individual neurons, we found that the time interval varied considerably, between 250 and 1150 ms, across the population (Fig. 3C). Despite this wide range of time intervals, a higher magnitude of facilitation tends to occur after shorter time intervals (p = 0.012).
It is not clear why facilitation appears in some neurons and not others. To address this question, we examined various factors that could possibly have introduced variability. Cortical neurons were recorded at different depths (760–1200 μm below the pia) and showed a large range of response latencies (8–16 ms) to principal whisker deflection. However, no correlation was found between the depth of the recorded neurons or the latency of the control response and the magnitude of facilitation at the subthreshold level (p = 0.45 and p = 0.47, respectively). Likewise, no significant difference was found between the average depths of the recordings (908 ± 50 μm vs 862 ± 20 μm; p = 0.56) or the latency of the control response of the neurons (11 ± 0.8 ms vs 10.5 ± 0.7 ms; p = 0.5) between neurons exhibiting significant versus nonsignificant facilitated response to the test stimulus, respectively. In addition, no correlation was found between the half-width response to control stimulation and the magnitude of the facilitatory response to the test stimulus (p = 0.23). Moreover, the prestimulus membrane potentials of control and test stimuli were compared within cells, and no significant difference was found within all recorded neurons. To test whether postadaptation facilitation is robust to changes in stimulation features, such as the magnitude and frequency of the adapting train, additional experiments were conducted in a subset of neurons (n = 8). Doubling the magnitude of whisker deflection or changing its frequency (10, 20, and 30 Hz) did not qualitatively change the nature of the test response (data not shown); namely, all the examined cells maintained their facilitatory behavior regardless of the frequency or magnitude of whisker stimulation. Only a weak negative correlation was found between the magnitude of facilitation and the degree of adaptation (the adaptation index was calculated by subtracting the averaged response to the last three whisker deflections of the repetitive train from the control response and dividing the difference by the sum of both, r = 0.4, p = 0.03; Fig. 3D).
Thalamic cells do not show facilitation after adaptation
Sensory inputs from the whiskers ascend to layer 4 of the primary somatosensory cortex in the lemniscal pathway through the VPM of the thalamus (Gil et al., 1999; Pierret et al., 2000; Bruno and Sakmann, 2006; Yu et al., 2006; Wang et al., 2010). Although our recordings were not confined to a specific layer, they were centered on layer 4; thus, the observed cortical facilitation could result from the response of VPM neurons. To address this possibility, we made intracellular recordings in the VPM (n = 9) and used a similar stimulation protocol as in our cortical recordings. We found that, although thalamic cells responses clearly adapted to repetitive stimulation (Fig. 4A,B for two representative cells and Fig. 4C for the maximal response across the population), they exhibited no postadaptation synaptic facilitation. Indeed, the response of most thalamic cells to test stimulus did not fully recover to baseline levels after adaptation within the time intervals we examined (Fig. 4C; mean maximal recovered response ± SEM, 6.9 ± 0.8 mV; control response ± SEM, 7.5 ± 0.8 mV; p = 0.004). Likewise, we found no significant difference between the jitter of control and test thalamic responses (p = 0.2, data not shown), suggesting that cortical facilitation is unlikely to be explained by changes in the synchronization of thalamic cells. In summary, these results imply that subcortical mechanisms are unlikely to account for the observed facilitation in the cortex.

Recovery of sensory response in the thalamus showed no facilitation. A, B, Average responses of two neurons during 20 Hz adapting whisker stimulation (vertical black lines) followed by a single whisker stimulation (vertical gray line) at three different time intervals. C, Average maximal response of nine thalamic cells to test stimulation plotted against their corresponding average response to control stimulation. Control response was calculated from all the trials using the bootstrap method (see Materials and Methods).
Slow recovery from adaptation of inhibitory inputs compared with excitatory inputs may explain the facilitation
As our thalamic recordings strongly suggest that the observed cortical facilitation is not the result of the facilitation of thalamic inputs, it most likely emerged through intracortical mechanisms. In particular, we tested whether the enhanced recovery response was the result of synaptic mechanisms. The enhanced response could result from either facilitation of evoked excitation or reduced inhibitory inputs. To distinguish between these possibilities, the responses of a subset of cells were recorded at different holding currents; depolarizing the cells to reduce the driving force of excitation while increasing the driving force of inhibition, or by hyperpolarizing the cells to increase the driving force of excitation while decreasing that of inhibition. In addition, to examine the response of the cells at their resting potential, trials were also recorded with no current injection. In these cells (n = 14), recordings were made with pipettes filled with QX-314 to prevent the neurons from firing action potentials (Connors and Prince, 1982). In these experiments, after an adapting train of whisker stimuli, a single test stimulus was applied at a fixed time interval (Fig. 5A–C). This interval was predetermined for each cell by examining its maximal test response, similarly to the former experiments.

Facilitatory response after adaptation is accompanied by reduced inhibitory input and a full recovery of excitation. A–C, An average response of a neuron to control and test whisker stimuli when zero current (resting, A), negative (hyperpolarization, B), and positive (depolarization, C) currents were injected. D–F, Population data for the magnitude of the responses to test versus control whisker stimulation for the three different injected currents.
Figure 5A shows an example cell recorded at its resting potential. In this cell, the response to a test stimulus applied 250 ms after termination of the first adapting train was facilitated by ∼60%. An average facilitation of ∼30% was measured at the resting potential for 14 neurons (Fig. 5D, mean control PSP ± SEM, 7.34 ± 1 mV; mean test PSP ± SEM, 9.6 ± 1.3 mV, p = 0.0004). However, when the example cell was hyperpolarized to unveil its excitatory inputs, no significant difference was found between its control and test responses (Fig. 5B), and this was also the case for the whole population (Fig. 5E, mean PSP ± SEM of whole population for the control, 19.6 ± 3.1 mV and for the test, 19.9 ± 3.2 mV, p = 0.46). Hence, the facilitation that we observed at the resting potential was not a result of facilitating excitatory inputs. In contrast, when the example cell was depolarized, the inhibitory test response was reduced by ∼36% compared with its control inhibitory response (Fig. 5C), and the average inhibitory response across the population was significantly reduced by ∼27% (mean test response ± SEM, 5.8 ± 0.9 vs mean control response ± SEM, 8.4 ± 1 mV, p = 0.0002, Fig. 5F). This strongly suggests that the facilitation observed at the resting potential results from reduced inhibition. For most cells (10 of 14), the baseline potential just before the control and test stimuli was not significantly affected by adaptation in the absence of current injection or when positive or negative currents were applied, which indicates that a change in the input resistance of the cells is not a major player in the facilitation of the response.
Discussion
To the best of our knowledge, this is the first report of postadaptation facilitation in the somatosensory system and the first study in which the mechanism underlying such behavior in any sensory system is unveiled. We found that the firing and synaptic responses to a sensory stimulus after adapting stimulation of the primary vibrissa are enhanced. The subthreshold postadaptation response was significantly enhanced in 34% of the intracellularly recorded neurons of the rat barrel cortex located in layer IV and its vicinity. Importantly, the time at which the response was facilitated varied across the population, ranging between 250 and 1150 ms after the adapting stimulation. Recordings from a subset of cells strongly suggest that the underlying mechanism is slower recovery of evoked inhibitory inputs compared with excitatory ones.
Mechanisms of adaptation
The degree of tactile adaptation of cortical neurons strongly depends on the frequency of stimulation (Ahissar et al., 2000; Castro-Alamancos, 2002; Chung et al., 2002; Khatri et al., 2004; Webber and Stanley, 2006), laminar location (Ahissar et al., 2001; Ego-Stengel et al., 2005; Katz et al., 2006), and cell type (Zhu and Connors, 1999; Brecht and Sakmann, 2002; Heiss et al., 2008). Because blind patch recordings were used in this study, some of these factors may explain the variability in the amount of adaptation we observed (Figs. 1, 2, and 5, example cells). Adaptation is also affected by stimulation strength (Ganmor et al., 2010) and the depth of anesthesia (Katz et al., 2012). Nevertheless, the dynamics of adaptation in our recordings are similar to that found in other in vivo intracellular recording studies (Brecht and Sakmann, 2002; Heiss et al., 2008), showing a widening of the postsynaptic response during the train and, in some cells, an increased response to the second stimulus in the train (Fig. 2A). Widening of the synaptic response increases firing jitter during the train (Fig. 2A,B) (Garabedian et al., 2003; Gabernet et al., 2005; Webber and Stanley, 2006). The widening of the response during the train partially reflects reduced synchrony and a weakening of thalamic inputs (Khatri et al., 2004; Heiss et al., 2008; Ganmor et al., 2010; Wang et al., 2010) but may also result from greater adaptation of inhibitory compared with excitatory inputs (Gabernet et al., 2005; Heiss et al., 2008), a process that increases the integration time window of excitation. Although the mechanisms responsible for the adaptation of inhibitory inputs are unclear, they may include reduced presynaptic excitation of the inhibitory neurons and/or depression of their output inhibitory synaptic transmission, which we previously proposed as the main mechanism based on measurements of synaptic delay (Heiss et al., 2008). This conjecture is in accordance with the maintained firing of inhibitory cells during repetitive whisker stimulation observed in both in vivo and in vitro studies (Khatri et al., 2004; Gabernet et al., 2005), even when whiskers were stimulated at higher frequencies than those used here.
Recovery from adaptation
Only a few studies (Chung et al., 2002; Ganmor et al., 2010) have explored recovery from adaptation in the vibrissa system. Our data indicate that recovery from adaptation is at least one order of magnitude more rapid than was previously reported by Chung et al. (2002). This discrepancy may stem from the different experimental conditions applied in each study, such as the depth and type of anesthesia (urethane vs halothane used in our study) as well as the intensity of the stimulation. Thalamic cells are known to adapt profoundly more when the depth of anesthesia is increased (Katz et al., 2012), which may cause a slower recovery. In addition, we previously showed that stimulus intensity substantially affects both the degree of adaptation and its recovery rate (Ganmor et al., 2010). In particular, at a low stimulation intensity, the thalamic cell response fully recovered from adaptation only after several seconds, whereas at a much higher intensity (similar to that used in this study) the response recovered in less than 1 s.
Mechanisms of enhanced recovery response
Adaptation usually reduces the neuronal response to subsequent stimulation. However, in the primary visual cortex, the firing response may be enhanced after adaptation (Dhruv et al., 2011; Wissig and Kohn, 2012). The mechanisms for this phenomenon remain unknown, and as far as we know such behavior was not previously reported in other sensory modalities. In approximately one-third of the cells that we recorded from the barrel cortex, we found a significant facilitation of the response during the recovery period from adaptation. A priori, several possibilities could explain this behavior. Facilitation could result from increased input resistance during the recovery period. Increased input resistance will produce a larger response, even when the strength of sensory-evoked synaptic input after the test stimulus is similar to or even smaller than that evoked by the control stimulus. In such a case, a larger change in membrane potential is expected upon injection of positive or negative currents after the adapting stimulation compared with that measured just before it. However, we found no significant change in the postadaptation membrane potential (at rest and under injected current) in the majority of the cells, suggesting that the facilitation we observed was not the result of an increased input resistance after adaptation.
Direct thalamic inputs are also unlikely to contribute to facilitation during the recovery period. This conclusion is supported not only by our thalamic recordings (Fig. 4), but also by in vitro studies indicating that these inputs are profoundly depressed (Gil et al., 1997; Gabernet et al., 2005) and do not show any paired pulse facilitation, as well as by the recovery response of thalamic cells after repetitive whisker stimulation in the study by Chung et al. (2002).
The enhanced response during the recovery period from adaptation may also result from increased evoked excitation or from reduced evoked inhibition. Several in vitro studies suggested that intracortical excitatory synapses in the granular, infragranular, and supragranular layers undergo synaptic facilitation (Gil et al., 1997; Varela et al., 1997; Beierlein and Connors, 2002; Lee and Sherman, 2008), whereas others showed that these synapses are mostly depressed (Feldmeyer et al., 2002; Petersen, 2002; Williams and Atkinson, 2007). Interestingly, monosynaptic connections in the prefrontal cortex, studied in brain slices, show similar apparent dynamics to those described in this study using sensory stimulation; namely, some facilitation during the early part of the train stimulation, followed by clear depression, and finally an enhanced response to test stimulation delivered several hundred milliseconds later (Markram et al., 1998; Wang et al., 2006). In addition, both facilitation and depression of monosynaptic connections between inhibitory neurons in the somatosensory cortex, depending on the presynaptic interneuron type, were described in vitro (Ma et al., 2012). Finally, despite the dominant depressing behavior of excitatory synapses in S1, as was described in brain slices, synaptic dynamics in the intact animal might be modulated by the presence of neuromodulators (Gil et al., 1997; Tsodyks and Markram, 1997; Oldford and Castro-Alamancos, 2003).
We found no evidence to support the possibility that facilitation of excitation could have caused the observed facilitated postadaptation response. Indeed, the recovery response under hyperpolarization, which unveiled the excitatory inputs, was similar in amplitude to the control response (Fig. 5E). However, we found that the recovery of inhibitory inputs was slower than that of excitatory inputs, and this imbalance led to facilitation during the recovery period from adaptation (Fig. 5C,F).
It remains unclear why facilitation was found only in approximately one-third of the cells and why it peaked at different times. We suggest that the optimal time for facilitation depends on the recovery dynamics of evoked excitatory and inhibitory inputs, their maximum strength, and the baseline conductance. If the test stimulation is applied too early, excitation may be too weak to evoke a large response, whereas if it is delivered too late, recovered inhibitory inputs may suppress the evoked excitatory inputs. Variability in the dynamic properties of the synapses across different cells will affect the time of maximal facilitation, and this may explain the large variability in the time of maximal recovery response (Fig. 3).
Another intriguing and probably related question is why evoked excitation of the facilitated response is not amplified by the recurrent cortical circuits despite the reduction in inhibition and the observed facilitation in cortical firing (Figs. 2 and 3). The reason is probably that, at any given time point during the recovery period, only a small fraction of the population exhibits enhanced responses (Fig. 3) and their contribution is insufficient to produce a large change in cortical excitation.
Possible functions of adaptation enhanced response
Although it remains unclear whether the priming effect of adaptation on subsequent cortical response has any perceptual effect in rodents, repetitive tactile stimulation in humans enhances amplitude and frequency discrimination (Goble and Hollins, 1993, 1994; Harris et al., 2002; Tannan et al., 2006) as well as spatial discrimination (Tannan et al., 2006). It was suggested that increased discriminability in the somatosensory system results from changes in thalamic synchrony that affect the thalamocortical transformations without increasing the responsiveness of cortical cells (Wang et al., 2010). Our data suggest that enhanced perception after adaptation may also be caused by an additional intracortical mechanism in which the altered balance between excitation and inhibition increases the responsiveness of cortical neurons in the somatosensory system. Importantly, it was proposed (Harris et al., 2002) that the primary somatosensory cortex, which is probably involved in enhanced human performance after adapting tactile stimulation (as mentioned above), is also related to working memory tasks. If so, postadaptation facilitation arising from the imbalance between excitation and inhibition and caused by short-term synaptic plasticity may be an essential component of the storage of working memory in the primary somatosensory cortex.
Footnotes
This work was supported by the Israel Science Foundation Grants 1160/11 and 1565/10, the Minerva Foundation, and the Deutsche Forschungsgemeinschaft SFB 889. We thank Boaz Mohar and Inbal Meir for their important comments and the members of the I.L. laboratory for their helpful contributions.
The authors declare no competing financial interests.
- Correspondence should be addressed to Dr. Ilan Lampl, Department of Neurobiology, Weizmann Institute of Science, Rehovot 76100, Israel. ilan.lampl{at}weizmann.ac.il





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}