

Abstract
Although it is well established that many glutamatergic neurons sequester Zn2+ within their synaptic vesicles, the physiological significance of synaptic Zn2+ remains poorly understood. In experiments performed in a Zn2+-enriched auditory brainstem nucleus—the dorsal cochlear nucleus—we discovered that synaptic Zn2+ and GPR39, a putative metabotropic Zn2+-sensing receptor (mZnR), are necessary for triggering the synthesis of the endocannabinoid 2-arachidonoylglycerol (2-AG). The postsynaptic production of 2-AG, in turn, inhibits presynaptic probability of neurotransmitter release, thus shaping synaptic strength and short-term synaptic plasticity. Zn2+-induced inhibition of transmitter release is absent in mutant mice that lack either vesicular Zn2+ or the mZnR. Moreover, mass spectrometry measurements of 2-AG levels reveal that Zn2+-mediated initiation of 2-AG synthesis is absent in mice lacking the mZnR. We reveal a previously unknown action of synaptic Zn2+: synaptic Zn2+ inhibits glutamate release by promoting 2-AG synthesis.
Introduction
Zn2+ is, after iron, the second most abundant trace element in humans. As an essential element for living organisms, Zn2+ plays a catalytic and structural role in many enzymes and regulatory proteins (Vallee, 1988). Since the surprising discovery that Zn2+ is present in large amounts within synaptic vesicles in many areas of the brain (Maske, 1955), numerous studies have investigated the possible roles of this metal on synaptic function. These studies have revealed that synaptic Zn2+, as an allosteric modulator, inhibits GABAA, NMDA, and kainate receptors (Paoletti et al., 1997; Vogt et al., 2000; Ruiz et al., 2004; Mott et al., 2008; Nozaki et al., 2011; Veran et al., 2012), while potentiating glycine receptors (Hirzel et al., 2006). Moreover, synaptic Zn2+, as a trigger of signaling pathways, is thought to be required for mossy fiber long-term potentiation (LTP) via presynaptic and postsynaptic mechanisms (Huang et al., 2008; Pan et al., 2011); however, these results are not consistent with other studies suggesting that Zn2+ signaling does not affect mossy fiber LTP (Vogt et al., 2000; Lavoie et al., 2011). Indeed, the establishment of specific presynaptic or postsynaptic pathways of Zn2+-mediated signaling and their physiological role on neuronal processing remain poorly understood.
The uniquely high concentrations of synaptic Zn2+ in the dorsal cochlear nucleus (DCN) (Frederickson et al., 1988), an auditory brainstem nucleus with well characterized circuitry and cellular mechanisms modulating synaptic transmission (Oertel and Young, 2004; Bender and Trussell, 2011), provide an attractive model for studying the role of synaptically released Zn2+ in regulating synaptic transmission. DCN principal neurons receive Zn2+-rich excitatory parallel fiber inputs in their apical dendrites, yet the physiological consequences of synaptic Zn2+ release in this circuit are completely unknown. Here, we report an unexpected Zn2+-mediated signaling pathway wherein synaptic Zn2+ reduces presynaptic glutamate release by triggering endocannabinoid synthesis. Our results reveal a previously unknown Zn2+-mediated signaling pathway that establishes a role of synaptic Zn2+ release in regulating neurotransmission.
Materials and Methods
Slice preparation
Experiments were conducted according to the methods approved by the Institutional Animal Care and Use Committee of the University of Pittsburgh. Coronal brainstem slices were prepared from either sex ICR, mZnR wild-type (mZnR WT) and knock-out (mZnR KO), and ZnT3 wild-type (ZnT3 WT) and knock-out (ZnT3 KO) mice (P17–P25). The preparation of coronal slices containing DCN has been described in detail (Tzounopoulos et al., 2004).
Electrophysiology
In vitro recordings.
Whole-cell voltage-clamp and current-clamp recordings were obtained from visually identified fusiform cells at a temperature of 31–33°C. Fusiform cells were identified on the basis of morphological and electrophysiological criteria (Tzounopoulos et al., 2004). For DCN recordings the external solution contained the following (in mm): 130 NaCl, 3 KCl, 2.4 CaCl2, 1.3 MgSO4, 21 NaHCO3, 3.5 HEPES, and 10 glucose; saturated with 95% O2/5% CO2.
For EPSC recordings fusiform cells were voltage clamped at −70 mV using pipettes with a K+-based internal solution of the following (in mm): 113 K-gluconate, 1.5 MgCl2, 14 Tris-phosphocreatine, 9 HEPES, 0.1 EGTA, 4 Na-ATP, 0.3 Tris-GTP, and 10 sucrose. All the internal solutions were adjusted to pH 7.3, ∼285 mOsmol. Voltage-clamp experiments were not included if the series and/or input resistance changed >20% during the recording. EPSCs in fusiform cells were evoked by stimulating parallel fiber tracts (0.2 Hz) in the presence of SR95531 (GABAA receptor antagonist, 20 μm) and strychnine (glycine receptor antagonist, 0.5 μm). Fiber tracts were stimulated with voltage pulses (100 μs, 10–30 V). To measure the time course of pharmacological manipulations, EPSC peak amplitude was measured and averaged every minute, then normalized to baseline. To ensure washout of bath-applied Zn2+ we perfused the slices at a rate of 6 ml/min. Synaptic Zn2+ release was induced by electrical stimulation of the parallel fibers using a protocol consisting of 100 pulses at 100 Hz and 30 pulses at 30 Hz. For depolarization-induced suppression of excitation (DSE) experiments, baseline EPSCs were acquired at a stimulation frequency of 0.67 Hz, followed by a depolarization of 5 s to 10 mV delivered to the postsynaptic cell. EPSC amplitude was measured and averaged from 10 sweeps in each cell and normalized to the average value before depolarization. DSE is reported as a percentage of average EPSCs 1–3 s after depolarization versus before depolarization. Electrophysiological data were acquired and analyzed using pClamp (Molecular Devices), IGOR Pro (Wavemetrics), and GraphPad Prism (GraphPad Software). All means are reported ± SEM. Statistical comparisons were made using ANOVA, and paired and unpaired two-tailed Student's t tests. Statistical significance was based on p values <0.05. All means are reported ± SEM.
mEPSCs recordings.
mEPSCs were recorded at a holding potential of −70 mV in the presence of tetrodotoxin (TTX; 0.5 μm), SR95531 (20 μm), and strychnine (0.5 μm). The intracellular solution contained the following (in mm): 128 CsMeSO3, 10 HEPES, 1 EGTA, 4 MgCl2, 4 ATP, 0.3 GTP, 10 phosphocreatine, 1 Cs-EGTA, 3 ascorbate, and 0.5 QX-314 (a blocker of voltage-activated Na+ channels). Ten second blocks of mEPSCs were acquired at a sample rate of 50 kHz and low-pass filtered at 10 kHz. Negative-voltage pulses (5 mV, 50 ms) were delivered every 10 s to monitor input and access resistance; mEPSC experiments were not included if the series and/or input resistance changed >20% during the recording. mEPSCs were detected and analyzed using Mini Analysis software (Synaptosoft) with amplitude and area threshold set at 3 root mean square noise level. All events were verified by visual inspection. Amplitude values were obtained by subtracting the average baseline from the amplitude at the local maximum during the event. Rise times were measured as time difference between 10 and 90% of the peak amplitude. Decay time was calculated as a time that took the event to decay to 37% of the peak amplitude.
Fluorescence imaging
For Ca2+ imaging, slices were loaded with fura-2 AM (25 μm, TEFLabs) for 20 min at room temperature, in the presence of 0.02% pluronic acid, and then washed in artificial CSF (ACSF) for at least 20 min (Besser et al., 2009). For extracellular Zn2+ imaging, Newport Green (2 μm) was added to the slice. Slices were then loaded in a microscope chamber and perfused with PO4-free ACSF to avoid Zn2+ precipitation (Chorin et al., 2011). Fluorescent imaging measurements were acquired every 3 s (Imaging Workbench 4, INDEC BioSystems and polychrome monochromator, TILL Photonics) using a 10× objective (Olympus BX51) with 4 × 4 binning of the image (SensiCam; PCO). Regions of interest (ROI) were chosen throughout the DCN molecular layer that was not covered by the stimulating electrode, and traces from four ROIs were averaged for each slice. Cell bodies are surrounded by our selected ROIs, thus, the observed fura-2 fluorescence changes mostly represent the result of Ca2+ rises in neuronal cell bodies. Fura-2 fluorescence signals are represented as a ratio of the signal obtained using excitation of 340 nm/380 nm wavelengths and a 510 nm emission bandpass filter (Chroma Technology). Newport Green fluorescent signals were normalized to the initial fluorescent level (F0) and the signal is represented as F/F0. Bar graphs represent the averaged difference in the fluorescent signal averaged over n slices as indicated. The stimulating electrode was placed on the molecular layer of the DCN and a train of 100 pulses at 100 Hz was applied (Master-8 stimulator unit; A.M.P.I.). When required, the initial baseline was used to calculate a linear regression curve that was subtracted from the trace.
2-Arachidonoylglycerol quantification
A modified isotopic dilution liquid chromatography/tandem mass spectrometry (LC/MS/MS) method (Zhang et al., 2010) was used to determine 2-arachidonoylglycerol (2-AG) in DCN slices. DCN slices were incubated at 37°C for ∼30 min to allow the tissue to recover after the cutting procedure (similar to electrophysiological experiments). 2-AG concentrations were measured in control samples and samples exposed to Zn2+ (100 μm, 5 min) in the presence or absence of U73122 (5 μm). In experiments in which U73122 was used, slices were pretreated with this drug or vehicle for 30 min. After this pretreatment, both control and treated slices were transferred to a chamber with either normal solution or Zn2+-containing solution for 5 min (U73122 was also present during the 5 min treatment). Samples were then immediately flash-frozen and stored in liquid nitrogen. The tissues were later thawed and homogenized in 20 μl of H2O with 0.02% trifluoroacetic acid and 2 μl aliquots were used for protein concentration determinations using the Bradford method. Then, 180 μl of cold acetonitrile was added to the samples previously spiked with 9 pmol 2-AG-d8 internal standard and followed by homogenization. The samples were centrifuged at 14,000 g for 10 min at 4°C and the supernatant transferred into vials. Brain homogenates at a 5 mg tissue/ml were used to perform quantification quality controls by spiking samples with internal standard and 300 pmol of 1-AG and 2-AG per milligrams of tissue. Samples were thawed and homogenized in the presence of deuterated 2-AG internal standard, thus controlling for any loss during sample work-up. The internal standard corrects for losses due to degradation, extraction efficiency, binding to silica present in the glass tubes, gas phase equilibrium, and ionization efficiencies. The concentration of 2-AG was normalized to amounts of DCN protein to avoid the errors introduced by handling small amounts of tissue per DCN slice (typically <1 mg).
Standard curve preparation.
A calibration curve was prepared by serial stock dilutions of 2-AG and 1-AG. Three independent standard curves were prepared and the limit of detection (3 × signal-to-noise ratio) and limit of quantification (LOQ) concentrations calculated (LOQ was defined as the lowest concentration displaying a relative SD percentage lower than 15).
LC.
The chromatography was performed on a Shimadzu SIL-20A HPLC equipped with a degasser, autosampler, and column oven (set at 37°C) on a 2.0 mm ID, 150 mm long, 3 μm C-18 Luna column (Phenomenex). The solvent system used consisted of 0.1% acetic acid in H2O and 0.1% acetic acid in acetonitrile.
MS.
The flow after the column was split and one-third was sent to a triple quadrupole for quantification purposes and two-thirds was diverted into an LTQ Velos Orbitrap for high MS resolution ion and product ion determinations. The quantification of 1-AG and 2-AG was performed in positive ion mode using a 4000 QTrap triple quadrupole (Applied Biosystems). The following transitions were optimized and used for quantification of 1-AG and 2-AG (m/z 379 → 287) and 2-AG-d8 (m/z 387 → 295) using a collision energy of 15, a declustering potential of 50 V, an entrance potential of 8 V, and a collision cell exit potential of 15 V. High-resolution spectra and accurate mass determinations (at a resolving power of 10,000) were obtained in samples and standards for 2-AG and 1-AG and its product ions.
Experiments with KO mice
Imaging and biochemical experiments were blinded (see Figs. 4, 8). Electrophysiological experiments were partially blinded: initial experiments were not blinded (approximately half n), but all subsequent experiments were blinded (see Figs. 6, 11, 12). As we did not observe any differences in the two datasets, we have included all these experiments.
Drugs and KO mice
SR95531, NBQX disodium salt, QX-314, AM251, TTX, and WIN55, 212–2 mesylate (WIN), were purchased from Ascent Scientific. ZnCl2, strychnine, BAPTA, GDP-β-S, tricine, and tetrahydrolipstatin (THL) were purchased from Sigma-Aldrich. The 2-AG, 1-AG, and 2H8 2-AG standards were purchased from Cayman Chemical. U73122 was purchased from Tocris Cookson. Fura-2 AM and Newport Green were purchased from TEFLabs and Invitrogen, respectively. GPR39/mZnR KO mice were kindly provided by D. Moechars from Janssen, Pharmaceutical Companies of Johnson & Johnson. ZnT3 KO mice were purchased from The Jackson Laboratory.
Results
Zn2+ decreases probability of release via activation of endocannabinoid signaling
To investigate the effects of Zn2+ on parallel fiber EPSC strength, we obtained whole-cell recordings from DCN principal neurons (fusiform cells). Bath application of Zn2+ (100 μm, 5 min) significantly decreased EPSC amplitude (Fig. 1A). The inhibitory effect of Zn2+ on EPSC strength was reversed upon Zn2+ removal, suggesting that extracellular Zn2+ mediates a short-lasting decrease in synaptic strength (Fig. 1A). The observed Zn2+-induced decrease in synaptic strength is due to presynaptic and/or postsynaptic mechanisms. To investigate whether the effects of Zn2+ on synaptic strength are mediated by presynaptic mechanisms, we used two assays that are sensitive to changes in presynaptic neurotransmitter release (Pr): paired-pulse ratio (PPR) and coefficient of variation (CV) analysis (Faber and Korn, 1991; Tsien and Malinow, 1991). Paired-pulse facilitation, an increased second response to two stimuli applied in rapid succession, is thought to reflect an increase in Pr. The CV is the SD of the EPSC amplitudes normalized to the mean amplitude and varies oppositely with quantal content; the inverse square, 1/CV2, is directly proportional to quantal content (quantal content = n p, where n = number of release sites and p = Pr). Application of Zn2+ increased PPR and decreased 1/CV2, both of which indicate a decrease in Pr (Fig. 1B,C). Baclofen (2–5 μm), which is known to inhibit synaptic strength via a presynaptic mechanism (Dutar and Nicoll, 1988), caused a similar increase in PPR and a similar reduction in 1/CV2, indicating that PPR and CV assays are highly sensitive to changes in Pr (Fig. 1B,C). Conversely, 0.5 μm 2,3-dihydroxy-6-nitro-7-sulfamoyl-benzo[f]quinoxaline-2,3-dione (NBQX), which reduces synaptic strength by partially blocking postsynaptic glutamate receptors, left PPR and 1/CV2 unaffected, thus confirming that PPR and CV assays are not sensitive to changes in synaptic strength that are mediated by postsynaptic manipulations (Fig. 1B,C). Consistent with these results, application of Zn2+ decreased miniature EPSC (mEPSC) frequency without affecting mEPSC amplitude (Fig. 1D). Together, these findings show that the Zn2+-induced EPSC depression is mediated by a decrease in Pr.

Zn2+ reduces synaptic strength by lowering Pr. A, Time course of EPSC peak amplitude reversible depression by 100 μm, 5 min Zn2+ application. (During Zn2+ application: 67.9 ± 9.0% of baseline, n = 7, p < 0.01; after Zn2+ washout: 94.5 ± 9.5% of baseline, n = 7, p = 0.7). Top, Representative averaged traces of EPSCs before (pre), during Zn2+, and after removal of Zn2+ (washout). B, PPR (20 ms interstimulus interval) is increased after Zn2+ application (PPR: baseline: 1.96 ± 0.11; after Zn2+: 2.42 ± 0.13; n = 7, p < 0.01). C, Zn2+-induced EPSC depression is associated with a decrease in Pr. 1/CV2 is decreased after Zn2+ (1/CV2 in Zn2+: 0.38 ± 0.1% of baseline, n = 7, p < 0.01). Effect of baclofen (2–5 μm) on PPR and 1/CV2 (PPR: baseline: 2.00 ± 0.15; after baclofen: 2.42 ± 0.10; n = 6, p < 0.01; 1/CV2 in baclofen: 0.27 ± 0.07% of baseline, n = 6, p < 0.01). Effect of NBQX (0.5 μm) on PPR and 1/CV2 (PPR: baseline: 1.84 ± 0.25; after NBQX: 1.89 ± 0.08; n = 6, p = 0.47; 1/CV2 in NBQX: 0.90 ± 0.08% of baseline, n = 8, p = 0.55). D, Cumulative probability plot of mEPSC frequencies shows that Zn2+ application decreased frequencies (left). Cumulative probability plot of mEPSC amplitudes shows Zn2+ application did not affect amplitudes (right). Top, Sample traces of mEPSCs before and after Zn2+ application (mean mEPSC frequency: baseline: 1.89 ± 0.3 Hz; after Zn2+: 1.22 ± 0.5 Hz; n = 4, p < 0.05 paired t test; mean mEPSC amplitude: baseline: 13.79 ± 0.81 pA; after Zn2+: 13.35 ± 0.94 pA; n = 4, p > 0.12 paired t test). Summary data represent mean ± SEM. Asterisks indicate statistically significant differences, p < 0.05.
Given the role of endocannabinoid retrograde signaling in determining the Pr of DCN parallel fiber synapses (Tzounopoulos et al., 2007; Tzounopoulos and Kraus, 2009; Sedlacek et al., 2011; Zhao and Tzounopoulos, 2011; Zhao et al., 2011), we tested whether endocannabinoids are involved in Zn2+-mediated modulation of Pr. We found that the specific cannabinoid receptor 1 (CB1R) antagonist AM-251 (1 μm) prevented Zn2+-mediated EPSC depression and the concomitant changes in PPR and 1/CV2 (Fig. 2A–C). Bath application of AM-251 (1 μm) did not affect baseline synaptic transmission, indicating lack of tonic activation of CB1Rs and lack of tonic endocannabinoid signaling in modulating baseline Pr (Fig. 2D). These results suggest that Zn2+-dependent endocannabinoid signaling mediates Pr reduction, thus revealing an unexpected link between Zn2+ and endocannabinoid signaling.

Zn2+-induced decrease in Pr is mediated by endocannabinoid signaling. A, Zn2+-induced EPSC depression is blocked by 1 μm AM-251 (EPSC amplitude in control: 66.4 ± 4.8% of baseline, n = 8, p < 0.01; in AM-251: 91.4 ± 7% of baseline, n = 6, p = 0.3). B, PPR (20 ms interstimulus interval) is increased after Zn2+ application in control conditions, but this increase is blocked in the presence of AM-251 (PPR in control: baseline: 1.88 ± 0.12; after Zn2+: 2.36 ± 0.11, n = 8, p < 0.01; PPR baseline in AM-251: 1.86 ± 0.2; after Zn2+: 1.91 ± 0.12; n = 6; p = 0.19. C, 1/CV2 is decreased after Zn2+ application in control conditions, but this decrease is blocked in the presence of AM-251 (1/CV2 in Zn2+: 0.45 ± 0.12%, n = 8, p < 0.01; 1/CV2 in Zn2+ + AM-251: 0.97 ± 0.11% of baseline, n = 6, p = 0.78). D, Time course of baseline EPSC amplitude before and after application of AM-251 (EPSC amplitude after AM-251 application: 107.6 ± 10.2% of baseline, n = 5, p = 0.34). Summary data represent mean ± SEM. Asterisks indicate statistically significant differences, p < 0.05.
Endocannabinoid modulation of Pr has both postsynaptic (endocannabinoid synthesis and release) and presynaptic (activation of cannabinoid receptors, CB1Rs) components (Alger, 2012). Therefore, we investigated whether Zn2+-dependent modulation of Pr is mediated through direct presynaptic activation or allosteric modulation of CB1Rs, or through initiation of a postsynaptic signaling cascade leading to endocannabinoid synthesis and release. Intracellular application of the calcium chelator BAPTA (10 mm) in fusiform cells blocked Zn2+-induced depression of EPSCs and the concomitant changes in PPR and 1/CV2, indicating that an increase in postsynaptic Ca2+ is necessary for Zn2+-mediated modulation of Pr (Fig. 3A–C). While BAPTA also chelates intracellular Zn2+ (Hyrc et al., 2000), subsequent experiments using the extracellular Zn2+ chelator tricine (Figs. 4, 9, 11) are consistent with a lack of dependence of our proposed mechanism on an intracellular Zn2+ increase. In addition, loading fusiform cells with GDP-β-S (0.5 mm), a nonhydrolyzable analog of GDP, blocked the effects of Zn2+ on EPSC amplitude, PPR, and 1/CV2, suggesting that postsynaptic activation of G-protein-coupled receptors (GPCRs) mediate Zn2+-induced activation of endocannabinoid signaling (Fig. 3A–C; see also figure legend for control experiments on the lack of effects on GDP-β-S on baseline synaptic transmission). Given that endocannabinoid synthesis from the postsynaptic neuron can be induced by elevations in intracellular Ca2+, by activation of GPCRs, or by concerted activation of GPCRs and elevations in intracellular Ca2+ (Wilson and Nicoll, 2001; Kim et al., 2002; Zhao and Tzounopoulos, 2011), our results indicate that Zn2+ likely promotes endocannabinoid production through a postsynaptic activation of GPCRs and a rise in intracellular Ca2+.

Zn2+-induced decrease in Pr requires rises in postsynaptic Ca2+ and postsynaptic activation of a GPCR pathway. A, Zn2+-induced EPSC depression is blocked by 10 mm intracellular BAPTA or 0.5 mm intracellular GDP-β-S (BAPTA: EPSC amplitude: 99.9 ± 8.5% of baseline, n = 5, p = 0.3; GDP-β-S: EPSC amplitude: 105.87 ± 4.39% of baseline, n = 7, p = 0.38). Our previous studies have shown that GDP-β-S does not affect parallel fiber EPSCs in fusiform cells; yet, application of GDP-β-S decreases the input resistance of fusiform cells (Zhao and Tzounopoulos, 2011). Thus, to avoid possible complications in the interpretation of our findings due to changes in input resistance and to allow diffusion of GDP-β-S in fusiform cells, we investigated the effect of GDP-β-S on Zn2+-mediated modulation of Pr after the GDP-β-S-mediated effect on input resistance reached a steady state: Zn2+ application took place ∼30 min after break-in of the cell. B, PPR (20 ms interstimulus interval) was not increased after Zn2+ application in the presence of BAPTA and GDP-β-S (BAPTA PPR: baseline: 1.95 ± 0.1; PPR after Zn2+: 1.92 ± 0.1; n = 5; p = 0.49; GDP-β-S: PPR baseline: 1.83 ± 0.1; PPR after Zn2+: 1.82 ± 0.1; n = 7; p = 0.8). C, 1/CV2 was not decreased after Zn2+ application in the presence of BAPTA and GDP-β-S (BAPTA: 1/CV2: 0.8 ± 0.12% of baseline, n = 5, p = 0.61; GDP-β-S: 1/CV2: 0.87 ± 0.13% of baseline, n = 7, p = 0.78). Summary data represent mean ± SEM.
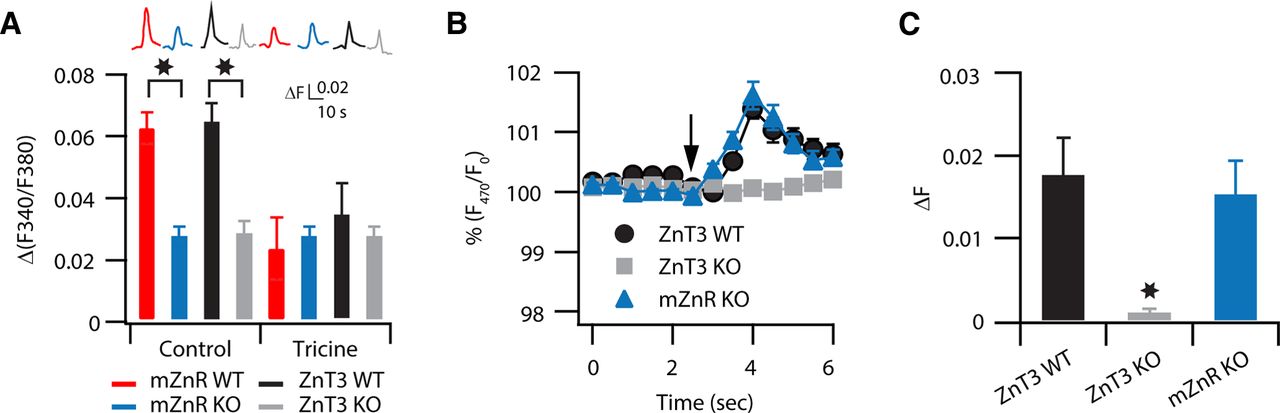
mZnR, a Zn2+-sensing Gq-protein-coupled receptor, mediates Zn2+-induced increase in intracellular Ca2+ in DCN neurons. A, Intracellular Ca2+ rise was monitored following a 100 Hz, 1 s stimulation of the molecular layer of the DCN in the presence or absence of tricine (10 mm, top traces). Slices from WT and KO mice were loaded with fura-2. The conditions for the different traces are indicated at the bottom of the bar graph. Bar graph showing averaged changes of Ca2+ rises. The residual Ca2+ response in the slices from mZnR KO and ZnT3 KO mice is not further attenuated by tricine and is similar to the response observed in the presence of tricine in slices from WT mice (ΔF/F: mZnR WT Control: 0.062 ± 0.005, n = 8; mZnR KO Control: 0.027 ± 0.003, n = 9, p < 0.01; mZnR WT in tricine: 0.023 ± 0.01, n = 9, p < 0.01 when compared with mZnR WT control; mZnR KO in tricine: 0.027 ± 0.003, n = 7, p = 0.48 when compared with mZnR WT in tricine; ZnT3 WT control: 0.064 ± 0.006, n = 4; ZnT3 KO control: 0.028 ± 0.004, n = 8, p < 0.01 when compared with ZnT3 WT control; ZnT3 WT in tricine: 0.034 ± 0.01, n = 8, p < 0.01 when compared with WT control; ZnT3 KO in tricine: 0.026 ± 0.004, n = 8, p = 0.52 when compared with ZnT3 KO control). B, Representative response illustrating extracellular Zn2+-dependent changes in 2 μm Newport Green (a nonpermeable, extracellular Zn2+ sensor) fluorescence in the DCN molecular layer after 1 s, 100 Hz stimulation of the parallel fibers in a slice from WT, ZnT3 KO, and mZnR KO mice. C, Average responses showing that Newport Green fluorescence in response to 1 s, 100 Hz stimulation of the parallel fibers was abolished in ZnT3 KO mice, but was normal in mZnR KO mice (ΔF: ZnT3 WT: 0.018 ± 0.004, n = 7 slices; ZnT3 KO: 0.001 ± 0.0004, n = 11 slices, p < 0.01 compared with ZnT3 WT; mZnR KO: 0.015 ± 0.004, n = 8 slices). Summary data represent mean ± SEM. Asterisks indicate statistically significant differences, p < 0.05.
Zn2+-induced decrease in Pr is mediated by a mZnR-dependent increase of 2-AG synthesis
Since endocannabinoid production is tightly linked to Gq-coupled receptors in the DCN (Zhao and Tzounopoulos, 2011), and because the previously orphan Gq-coupled receptor 39 (GPR39) has emerged as a putative metabotropic Zn2+-sensing receptor (mZnR) (Besser et al., 2009), we evaluated whether mZnR activation is involved in the Zn2+-triggered promotion of endocannabinoid production in the DCN. As activation of mZnRs is thought to lead to Zn2+-mediated increases in intracellular Ca2+ in the hippocampus and in cortical neurons (Besser et al., 2009; Saadi et al., 2011; but see Evstratova and Toth, 2011), we determined whether synaptically released Zn2+ leads to an mZnR-mediated increase in intracellular Ca2+ in the DCN. We measured intracellular Ca2+ responses in the molecular layer of DCN slices from WT and from genetically modified mice lacking GPR39 (Moechars et al., 2006) (mZnR KO mice). While parallel fiber stimulation (100 Hz, 1 s) caused robust intracellular Ca2+ responses in slices from mZnR WT mice, measurements from littermate mZnR KO mice displayed significantly reduced Ca2+ responses (Fig. 4A). To determine whether reduced Ca2+ responses in mZnR KO mice are due to the lack of Zn2+-triggered, mZnR-mediated signaling, we measured the sensitivity of the Ca2+ responses to tricine: an extracellular Zn2+ chelator (Paoletti et al., 2009). Application of 10 mm tricine reduced Ca2+ responses in WT mice and left Ca2+ responses unaffected in mZnR KO mice; tricine also reduced Ca2+ responses in WT mice to levels similar to those observed in mZnR KO mice (Fig. 4A). Because tricine only reduces Ca2+ responses in WT mice, but not in KO mice, we conclude that the observed differences in Ca2+ responses are due to Zn2+-mediated, mZnR-dependent signaling.
To further support that vesicular Zn2+ triggers Zn2+-mediated signaling via mZnR, we used a mouse genetic model lacking the transporter that loads Zn2+ into synaptic vesicles (ZnT3 KO) mice (Cole et al., 1999). ZnT3 KO mice lack vesicular Zn2+ staining and vesicular Zn2+ release (Cole et al., 1999; Qian and Noebels, 2005; Chorin et al., 2011). As expected, while ZnT3 WT mice displayed DCN vesicular Zn2+ release after parallel fiber stimulation, DCN vesicular Zn2+ release was undetectable in ZnT3 KO mice (Fig. 4B,C). In response to parallel fiber stimulation, ZnT3 KO mice displayed reduced Ca2+ responses when compared with WT mice; in addition, tricine did not affect Ca2+ responses in ZnT3 KO mice (Fig. 4A). These results indicate that a significant proportion of the Ca2+ response following stimulation of the parallel fibers requires both vesicular Zn2+ and the mZnR. While fura-2 can also detect changes in intracellular Zn2+, previous studies have established that activation of mZnRs by extracellular Zn2+ leads to a phospholipase C (PLC)-dependent increase in intracellular Ca2+ that is not associated with Zn2+ influx (Besser et al., 2009; Chorin et al., 2011; Saadi et al., 2012). Finally, mZnR KO mice displayed normal DCN vesicular Zn2+ release (Fig. 4B,C). Together these findings suggest that mZnRs are functionally expressed in the DCN neurons and participate in Zn2+-triggered increases in intracellular Ca2+ responses.
Given the dependence of the Zn2+-induced reduction of Pr on postsynaptic Ca2+ increases and GPCR activation (Fig. 3A), as well as the mZnR-mediated increase in Ca2+ responses in DCN neurons (Fig. 4A), we tested whether mZnRs are necessary for the Zn2+-induced reduction of synaptic strength. We found that Zn2+-mediated modulation of EPSC amplitude, PPR, and 1/CV2 were absent in mZnR KO mice (Fig. 5A–C). In contrast, littermate WT mice displayed reversible Zn2+-induced reductions on EPSC amplitude and Pr (Fig. 5A–C). PPR levels were not different between WT mZnR and mZnR KO mice, suggesting that the absence of mZnR does not affect baseline Pr (Fig. 5B, p = 0.22). Importantly, mEPSC analysis revealed that basal quantal properties—mini EPSC frequency and amplitude—were not different between mZnR WT and KO mice (Fig. 6A,B). Moreover, mEPSC rise and decay times were not different between WT and KO mice (Fig. 6 C,D). These results indicate that the lack of effect of Zn2+ on reducing synaptic strength and Pr in mZnR KO mice is not due to differences in basal synaptic properties between WT and KO mice. The lack of Zn2+-mediated changes in Pr in mZnR KO mice was not caused by nonfunctional endocannabinoid signaling in mZnR KO mice. This was determined by the fact that depolarization-induced suppression of excitation (DSE is induced by postsynaptic depolarization leading to rises in postsynaptic Ca2+ levels that, in turn, trigger endocannabinoid synthesis and Pr reduction) was not different between mZnR WT and KO mice (Fig. 6E). DSE in mZnR KOs was blocked by AM-251, confirming that it is mediated by CB1Rs (n = 5, data not shown). These results reveal that baseline Pr and depolarization-induced endocannabinoid signaling are intact in mZnR KO mice. Finally, application of 2 μm WIN (a CB1R agonist) caused a quantitatively and kinetically similar decrease in EPSC amplitude in WT and KO mice (Fig. 6F), suggesting that CB1R function is not altered between mZnR WT and mZnR KO mice. Because Zn2+-induced modulation of Pr and the Zn2+-induced promotion of endocannabinoid signaling are absent in mZnR KO mice, we conclude that mZnRs are necessary for the Zn2+-induced reduction of Pr; this Pr reduction is likely occurring via the activation of postsynaptic endocannabinoid synthesis.
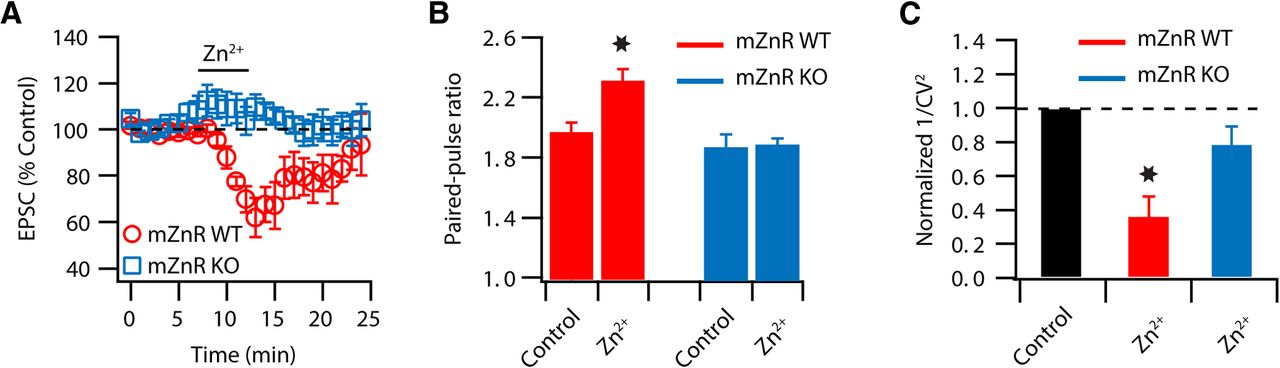
mZnR is necessary for Zn2+-induced decrease in Pr. A, Zn2+-induced reversible depression of EPSCs was observed in WT mice but not in mZnR KO littermates (EPSC amplitude mZnR WT: 65.7 ± 8% of baseline, n = 6, p < 0.01; mZnR KO: 107.4 ± 5.6% of baseline, n = 5, p = 0.27). B, PPR (20 ms interstimulus interval) is increased after Zn2+ application in mZnR WT mice but is unaffected in mZnR KO littermates (mZnR WT PPR baseline: 1.97 ± 0.05; PPR after Zn2+: 2.32 ± 0.06; n = 6, p < 0.01; mZnR KO: PPR baseline: 1.88 ± 0.07; PPR after Zn2+: 1.90 ± 0.03; n = 5, p = 0.75). C, 1/CV2 is decreased after Zn2+ application in mZnR WT mice but is unaffected in mZnR KO littermates (mZnR WT 1/CV2: 0.36 ± 0.11% of baseline, n = 6, p < 0.01; mZnR KO: 1/CV2: 0.79 ± 0.11% of baseline, n = 5, p = 0.15). Summary data represent mean ± SEM. Asterisks indicate statistically significant differences, p < 0.05.

Quantal properties and depolarization-induced endocannabinoid signaling are not altered in mZnR KO mice. A, Cumulative probability plot of mEPSC frequencies shows that frequencies are not different between mZnR WT and KO mice (mean mEPSC frequency mZnR WT: 3.03 ± 0.45 Hz, n = 5; mZnR KO: 3.52 ± 0.59 Hz; n = 5, p = 0.51). Top, Sample traces of mEPSCs from mZnR WT and KO mice. B, Cumulative probability plot of mEPSC amplitudes shows that amplitudes are not different between mZnR WT and KO mice (mean mEPSC amplitude mZnR WT: 16.62 ± 2.09 pA, n = 5; mZnR KO: 15.56 ± 0.65 pA; n = 5, p = 0.64). C, Cumulative probability plot of mEPSC rise times shows that rise times are not different between mZnR WT and KO mice (mean mEPSC rise time mZnR WT: 0.63 ± 0.05 ms, n = 5; mZnR KO: 0.66 ± 0.05 ms; n = 5, p = 0.61). D, Cumulative probability plot of mEPSC decay times shows that decay times are not different between mZnR WT and KO mice (mean mEPSC decay time: mZnR WT: 1.3 ± 0.09 ms, n = 5; mZnR KO: 1.43 ± 0.07 ms; n = 5, p = 0.28). E, Time course of DSE, induced by 5 s depolarization to 10 mV in mZnR WT and KO mice (mZnR WT DSE: 37 ± 3.1% of control, n = 5; mZnR KO DSE: 41 ± 5.3% of control, n = 6, p = 0.35). F, Time course of 2 μm WIN on baseline synaptic transmission of mZnR WT and KO mice (mZnR WT: 31.8 ± 0.77% of baseline, n = 5, mZnR KO: 30.84 ± 1.9%, n = 5, p = 0.32). Summary data represent mean ± SEM.
To determine the specific endocannabinoid that leads to mZnR-mediated endocannabinoid signaling and subsequent Pr reduction, we used a combined electrophysiological and biochemical approach. 2-AG, the endocannabinoid that mediates retrograde signaling in the DCN, is synthesized following PLC activation and the cleavage of diacylglycerol by a diacylglycerol lipase (DGL) (Zhao et al., 2009). To determine whether Zn2+-induced changes in Pr are mediated through the 2-AG biosynthetic pathway, we first investigated the effect of pharmacological blockade of PLC and DGL on Zn2+-mediated Pr modulation. We observed that incubation of slices with either a PLC inhibitor (5 μm U73122), or a DGL inhibitor (10 μm THL) blocked Zn2+-induced depression of EPSC amplitude and the concomitant Zn2+-induced changes in PPR and 1/CV2 (Fig. 7A–C). Moreover, bath application of both these inhibitors did not affect baseline synaptic transmission, or CV of evoked EPSCs, indicating that their inhibitory effect on Zn2+-induced Pr reduction is not due to a nonspecific change of baseline Pr that occludes or blocks further changes in Pr by Zn2+ (Fig. 7D–F). Together, these results suggest that application of Zn2+ leads to a PLC-dependent production of 2-AG.

Zn2+-induced decrease in Pr is mediated by a PLC- and DGL-dependent metabolic pathway. A, Zn2+-induced reversible depression of EPSCs was blocked by bath application of 5 μm U73122 or 10 μm THL (EPSC amplitude control: 67.60 ± 6.10% of baseline, n = 5, p < 0.01; U73122: 98.30 ± 3.66% of baseline, n = 6, p = 0.33; THL: 105.40 ± 7.70% of baseline, n = 6, p = 0.42). B, PPR (20 ms interstimulus interval) is increased after Zn2+ application in control conditions, but this increase is blocked in the presence of either U73122 or THL (PPR Control baseline: 1.91 ± 0.08; after Zn2+: 2.44 ± 0.16; n = 5; p < 0.01; U73122 baseline: 2.09 ± 0.08; after Zn2+: 2.06 ± 0.09; n = 6; p = 0.59; THL baseline: 1.78 ± 0.11; after Zn2+: 1.72 ± 0.10; n = 6; p = 0.3). C, 1/CV2 is decreased after Zn2+ application in control but this decrease is blocked in the presence of either U73122 or THL (1/CV2: after Zn2+ 0.46 ± 0.11% of baseline, n = 6, p < 0.01; + U73122 1.27 ± 0.24% of baseline, n = 6, p = 0.41; + THL: 1.13 ± 0.26% of baseline, n = 6, p = 0.56). D, Bath application of U73122 does not affect baseline synaptic transmission (EPSC amplitude: 104.9 ± 2.9%, of baseline, n = 5, p = 0.25). E, Bath application of THL does not affect baseline synaptic transmission (EPSC amplitude: 101.5 ± 2.1%, n = 5, p = 0.35). F, Bath application of either U73122 or THL does not affect 1/CV2 (1/CV2 U73122: 1.03 ± 0.08% of baseline, n = 5, p = 0.52; THL: 0.99 ± 0.12% of baseline, n = 5, p = 0.59). Summary data represent mean ± SEM. Asterisks indicate statistically significant differences, p < 0.05.
Next, we used isotopic dilution LC/MS/MS to quantify 2-AG levels and their dependence on Zn2+. The two different isomers of AG (2-AG and 1-AG) were chromatographically resolved and base peak separations were achieved (Fig. 8A). Consistent with previous reports (Zhang et al., 2010), the isotopically labeled 2-AG-d8 had a 0.12 min shorter retention time (Fig. 8A). The formation of endogenous 2-AG was confirmed by co-elution with internal standard (Fig. 8A) and by high-resolution MS and MS/MS data obtained on the 2-AG peak, where a similar fragmentation pattern was observed between the standard and the DCN tissue samples (Fig. 8B,C). In agreement with our hypothesis on the enhancing effect of Zn2+ on the 2-AG metabolic pathway (Fig. 7), HPLC-MS analysis showed that DCN 2-AG levels increased significantly after Zn2+ (100 μm for 5 min) application (Fig. 8D). Moreover, Zn2+-mediated increases in 2-AG levels were abolished when slices were either pretreated with the PLC inhibitor (U73122, 5 μm for 30 min before Zn2+ application) or in mZnR KO mice (Fig. 8D). These results indicate that exogenous Zn2+ leads to increased 2-AG synthesis via a PLC-dependent pathway. Given that 2-AG synthesizing enzymes have been localized in the dendrites of DCN fusiform cells (Zhao et al., 2009), our results suggest that activation of mZnR leads to the activation of postsynaptic PLC and DGL leading to enhanced postsynaptic 2-AG production.

LC/MS/MS analysis reveals that Zn2+ induces a mZnR-dependent increase of 2-AG synthesis in the DCN. A, The two different AG isomers (2-AG and 1-AG) were chromatographically resolved and base peak separations were achieved. 2-AG and 2-AG-d8 were detected following the 379.3 to 287.2 m/z and 387.3 to 295.2 transitions, respectively, using a triple quadrupole. In parallel accurate mass determination using a Velos Orbitrap resulted in mass confirmations at the 2 ppm level for standards and for tissue-obtained 2-AG (inserts). B, 2-AG identity was confirmed by co-elution with internal standard (A) and by high-resolution MS and MS/MS data obtained on the 2-AG peak, where a similar fragmentation pattern was observed between the 2-AG standard and the DCN tissue samples. C, Molecular structures of product ions obtained upon ion trap collision-induced dissociations of 2-AG shown in B. D, Zn2+ application (100 μm for 5 min) increased 2-AG levels in a PLC and mZnR-dependent manner. HPLC-MS measurements of 2-AG levels in DCN slices from WT and mZnR KO littermates in the presence of Zn2+ and/or U73122 (5 μm, PLC inhibitor applied for 30 min before Zn2+ application; mZnR WT Control: 1 ± 0.06 pmol/μg protein, n = 8; mZnR WT in Zn2+: 2.2 ± 0.1 pmol/μg protein, n = 8, p < 0.01; mZnR WT in U73122: 0.97 ± 0.51 pmol/μg protein, n = 6 when compared with mZnR WT control; p = 0.6; mZnR WT in U73122 and Zn2+: 0.89 ± 0.37 pmol/μg protein, n = 7; p = 0.42 when compared with mZnR WT control; mZnR KO: 1.3 ± 0.08 pmol/μg protein in control, n = 6; mZnR KO in Zn2+: 1.2 ± 0.02 pmol/μg protein, n = 4, p = 0.5 when compared with mZnR KO control). Summary data represent mean ± SEM. Asterisks indicate statistically significant differences, p < 0.05.
Endogenous synaptic Zn2+ levels decrease Pr via mZnR and endocannabinoid signaling
Next, we investigated whether endogenous, synaptically released Zn2+ reduces Pr via promotion of endocannabinoid signaling. Brief bursts of presynaptic activity can promote the synthesis and release of endocannabinoids from postsynaptic cells, leading to presynaptic CB1 receptor activation that, in turn, suppress neurotransmitter release (Brown et al., 2003). This process leads to endocannabinoid-mediated short-term plasticity: short-term depression in this case. Therefore, to determine whether endogenous Zn2+ triggers endocannabinoid signaling, we investigated the role of synaptically released Zn2+ in short-term plasticity. We measured parallel fiber-evoked EPSCs at low frequencies (0.3 Hz) before and after a high-frequency stimulus train (100 Hz, 1 s). This stimulus train produced short-term facilitation in EPSC amplitude (STF; Fig. 9A1). Bath application of the extracellular Zn2+ chelator tricine after the initial stimulus train, in the same cell, significantly increased the magnitude of STF induced by an identical stimulus train (Fig. 9A1,A3). These results suggest that chelation of synaptically released Zn2+ during the high-frequency train enhanced the magnitude of induced STF. Brief trains of presynaptic activity, apart from promoting synthesis and release of endocannabinoids, trigger a build-up of calcium in presynaptic boutons resulting in forms of short-term plasticity (Zucker and Regehr, 2002), such as augmentation and post-tetanic potentiation (PTP). In many synapses, the combined effect of PTP and endocannabinoid signaling determines the size and the sign of short-term plasticity (Beierlein et al., 2007). Therefore, one possible interpretation of the effect of tricine in enhancing the amount of STF is that the chelation of Zn2+ blocks Zn2+-mediated endocannabinoid signaling and short-term depression, thereby leading to enhanced STF. This hypothesis predicts that blockade of endocannabinoid signaling should increase the amount of STF and occlude the enhancing effect of tricine. Consistent with this hypothesis, application of AM-251 increased STF and occluded the enhancement of STF produced by Zn2+ chelation (Fig. 9A2,A3). These results suggest that endogenous, synaptically released Zn2+ modulates STF by enhancing endocannabinoid signaling.

Synaptically released Zn2+ modulates STF via endocannabinoid signaling. A1, STF is enhanced after Zn2+ chelation (STF control: 58.24 ± 3.4%, n = 9; tricine: 101.5 ± 11.16%, n = 9, p < 0.01). Summary graph showing the time course of the change in EPSC amplitude after 100 Hz, 1 s parallel fiber stimulation, in control and after endogenous Zn2+ chelation with tricine (100 Hz, 1 s stimulation was applied before and after tricine for the same and for each cell; 10 mm tricine was bath applied for at least 10 min before 100 Hz, 1 s stimulation). The high-frequency stimulus was delivered under current clamp to allow postsynaptic spiking. Top, Representative traces of EPSCs before the train (pre-burst), and after the 100 Hz train (post-burst). A2, AM-251-enhanced baseline STF and blocked the enhancement of STF after Zn2+ chelation (STF control in AM-251: 90.32 ± 3.40%, n = 10, p < 0.05 when compared with control from A1; tricine in AM-251: 90.55 ± 11.16%, n = 10, p = 0.28 when compared with control in AM-251). Summary graph showing the time of the change in the EPSC amplitude after 100 Hz, 1 s parallel fiber stimulation as in A1, but now in the presence of AM-251 (pre-incubation of slices with 1 μm AM-251 for at 1 h and bath application during the experiment). Top, Representative traces of EPSCs before the train (pre-burst), and after the 100 Hz train (post-burst). A3, Average values of STF from A1 and A2. Percentage STF magnitude is the average of first three points after the train. B1, STF is revealed after Zn2+ chelation; synaptic Zn2+ modulates the threshold for STF (STF control: 6.66 ± 6.10%, n = 6, p = 0.25; tricine: 33.95 ± 3.31%, n = 6, p < 0.01). Summary graph showing the time course of the change in the EPSC amplitude after 30 Hz, 1 s parallel fiber stimulation, in control and after endogenous Zn2+ chelation with tricine (30 Hz, 1 s stimulation was applied before and after tricine for the same and for each cell; 10 mm tricine was bath applied for at least 10 min before 30 Hz, 1 s stimulation). The high-frequency stimulus was delivered under current clamp to allow postsynaptic spiking. B2, AM-251 enhanced baseline STF and blocked the enhancement of STF after Zn2+ chelation. Summary graph showing the time of the change in the EPSC amplitude after 100 Hz, 1 s parallel fiber stimulation as in B1, but now in the presence of AM-251 (pre-incubation of slices with 1 μm AM-251 for at 1 h and bath application during the experiment; STF control in AM-251: 34.27 ± 5.97%, n = 6, p < 0.01 when compared with control from B1; tricine in AM-251: 38.16 ± 6.23%, n = 6, p = 0.38 when compared with control in AM-251). B3, Average values of STF from B1 and B2. Percentage STF magnitude is the average of first three points after the train. Summary data represent mean ± SEM. Asterisks indicate statistically significant differences, p < 0.05.
If chelation of synaptic Zn2+ enhances STF (Fig. 9A) by affecting the balance between opposing forms of modulation of Pr, we expect that synaptic Zn2+ would modulate the threshold for inducing STF. To test this hypothesis, we determined the effect of Zn2+ chelation on threshold conditions under which presynaptic trains of action potentials fail to induce short-term plasticity. We varied the frequency of presynaptic bursts and investigated the effect of tricine on the resulting short-term plasticity. We found that although a 30 Hz, 1 s stimulation did not affect synaptic strength, application of tricine, under identical conditions, resulted in significant STF (Fig. 9B1, B3). Moreover, application of AM-251 alone under these conditions also revealed STF and, importantly, occluded the enhancing effect of tricine (Fig. 9B2, B3). Together, these results suggest that the release of endogenous Zn2+ during bursts of presynaptic activity promotes endocannabinoid signaling and shifts the balance of opposing short-term plasticity mechanisms toward reduction, or decreased enhancement, of synaptic strength.
Because PTP depends on baseline Pr (Zucker and Regehr, 2002), potential effects of tricine on baseline Pr would affect the amount of observed STF. However, tricine did not affect baseline EPSC strength, PPR, or CV, suggesting that its effects on short-term plasticity are not due to changes of baseline Pr (Fig. 10A–C). Moreover, these observations indicate that any release of synaptic Zn2+ during baseline synaptic transmission is not capable of promoting endocannabinoid signaling; presynaptic trains of action potentials are needed for Zn2+-induced promotion of endocannabinoid signaling. Together, these findings (Figs. 9,10) reveal a previously unknown role for synaptic Zn2+ in setting the threshold for short-term plasticity via enhancement of endocannabinoid signaling.

Tricine does not affect either baseline synaptic transmission or Pr. A, Time course of the effect of tricine (10 mm) on EPSC amplitude (EPSC amplitude: 106.4 ± 6.7% of control, n = 6, p = 0.57). B, C, PPR (PPR control: 1.71 ± 0.11; tricine: 1.72 ± 0.10, n = 6, p = 0.51) (B) and 1/CV2 (1/CV2 in tricine: 1.28 ± 0.18% of control, n = 6, p = 0.3) (C) did not change after tricine application. Summary data represent mean ± SEM. Asterisks indicate statistically significant differences, p < 0.05.
Given that exogenous Zn2+ triggers endocannabinoid synthesis that is dependent on mZnRs (Figs. 2⇑⇑⇑⇑⇑–8), we hypothesized that endogenous Zn2+-mediated reduction of STF (Fig. 9) is also dependent on mZnRs. If mZnRs are necessary for the Zn2+-induced endocannabinoid synthesis, then we expect that STF should be increased in mZnR KO mice; the enhancing effect of tricine on STF should also be absent in mZnR KO mice. In agreement with our hypotheses, STF in response to 100 Hz stimulation was enhanced in mZnR KO mice (Fig. 11A1,A3). Moreover, chelation of synaptically released Zn2+ by tricine did not increase the magnitude of STF in mZnR KO mice, suggesting that the lack of the zinc receptor, similar to the application of AM-251 in control mice in Figure 9, occludes the effect of tricine (Fig. 11A2, A3). Since depolarization-induced endocannabinoid signaling is intact in mZnR KO and baseline Pr is not affected by the absence of mZnR (Fig. 6), these results suggest that the enhancing effect of synaptic Zn2+ on short-term plasticity is dependent on mZnR activation of endocannabinoid signaling.

Synaptically released Zn2+-induced modulation of STF is absent in mZnR KO and in ZnT3 KO mice. In A1–A3 baseline STF, elicited by 100 Hz, 1 s stimulation is increased in mZnR KO mice; tricine-mediated enhancement of STF is absent in mZnR KO littermates. A1, Summary graph showing the time course of the change in EPSC amplitude after 100 Hz, 1 s parallel fiber stimulation, in control and after endogenous Zn2+ chelation with tricine in mZnR WT mice (100 Hz, 1 s stimulation was applied before and after tricine for the same and for each cell; 10 mm tricine was bath applied for at least 10 min before 100 Hz, 1 s stimulation; STF mZnR WT: 45.41 ± 6.65%, n = 10; mZnR KO: 87.28 ± 13.54%, n = 10, p < 0.01). A2, Summary graph showing the time course of the change in EPSC amplitude after 100 Hz, 1 s parallel fiber stimulation, in control and after endogenous Zn2+ chelation with tricine as in A1, but now in mZnR KO mice (STF: mZnR KO control: 87.28 ± 13.54%, n = 10; mZnR KO in tricine: 76.83 ± 17.10%, n = 10, p = 0.30). A3, Average values of STF from A1 and A2. Percentage STF magnitude is the average of the first three points after the train. In B1–B3 baseline STF elicited by 100 Hz, 1 s stimulation is increased in ZnT3 KO mice; tricine-mediated enhancement of STF is absent in ZnT3 KO littermates. B1, Summary graph showing the time course of the change in EPSC amplitude after 100 Hz, 1 s parallel fiber stimulation, in control and after endogenous Zn2+ chelation with tricine in ZnT3 WT mice (conditions are the same as in A1; STF ZnT3 WT: 39.28 ± 3.97%, n = 9; ZnT3 KO: 87.36 ± 10.92%, n = 9, p < 0.01). B2, Summary graph showing the time course of the change in EPSC amplitude after 100 Hz, 1 s parallel fiber stimulation, in control and after endogenous Zn2+ chelation with tricine in ZnT3 KO mice (conditions are the same as in A1; STF ZnT3 KO control: 87.36 ± 10.92%, n = 9; ZnT3 KO in tricine: 71.6 ± 12.29%, n = 9, p = 0.42). B3, Average values of STF from B1 and B2. Percentage STF magnitude is the average of the first three points after the train. In C1–C5 baseline STF elicited by 30 Hz, 1 s stimulation is revealed in mZnR KO and in ZnT3 KO mice. C1, C2, Summary graphs showing the time course of the change in EPSC amplitude after 30 Hz, 1 s parallel fiber stimulation, in mZnR WT and in mZnR KO littermates (STF mZnR WT: 14.35 ± 4.41%, n = 6; mZnR KO: 42.7 ± 14.10%, n = 6 p < 0.01 when compared with mZnR WT). C3, C4, Summary graphs showing the time course of the change in EPSC amplitude after 30 Hz, 1 s parallel fiber stimulation, in ZnT3 WT and in ZnT3 KO littermates (STF ZnT3 WT: 14.25 ± 6.91%, n = 6; ZnT3 KO: 49.34 ± 6.01%, n = 6, p < 0.01 when compared with ZnT3 WT). C5, Average values of STF from C1–C4. Percentage STF magnitude is the average of the first three points after the train. Summary data represent mean ± SEM. Asterisks indicate statistically significant differences, p < 0.05.
To determine the effect of elimination of synaptic Zn2+ release on endocannabinoid signaling and short-term plasticity, we also used ZnT3 KO mice, which do not display any DCN vesicular Zn2+ release (Fig. 4B,C). Given that Zn2+-chelation experiments with tricine suggest that the release of vesicular Zn2+ decreases STF levels (Fig. 9), ZnT3 KO mice are expected to show more robust STF than WT littermate mice. Moreover, we expected that the enhancing effect of tricine on STF would be occluded in ZnT3 KO mice. Indeed, comparison of STF levels between ZnT3 WT and ZnT3 KO littermates revealed increased levels of STF (100 Hz stimulation) in ZnT3 KO mice (Fig. 11B1, B3). Additionally, the enhancing effect of tricine was occluded in ZnT3 KO mice (Fig. 11 B2,B3). Together these results suggest that endogenous, synaptically released Zn2+ mediates the depressing effects in short-term plasticity observed in fusiform cells.
During milder presynaptic stimulation (30 Hz, 1 s) synaptic Zn2+ increased the threshold for STF (Fig. 9B1–B3). Based on this finding, we hypothesized that the threshold for inducing STF would be lower in mZnR KO and in ZnT3 KO mice. To test this hypothesis we investigated the amount of STF after a 30 Hz 1 s presynaptic stimulus. In accordance with our hypothesis, while slices from mZnR WT and ZnT3 WT mice showed minimal levels of STF, mZnR KO and ZnT3 KO mice showed significantly enhanced levels of STF (Fig. 11C1–C5). The observed effects in ZnT3KO mice are not due to changes in basal quantal properties, baseline Pr, or in deficient endocannabinoid signaling in ZnT3 KO mice, as WT and KO mice displayed similar mEPSC frequency, amplitude, rise time, decay time, similar levels of baseline PPR, and DSE (Fig. 12A–F). Together, our physiological, pharmacological, genetic, and biochemical results show that release of endogenous, vesicular Zn2+ reduces neurotransmitter release via mZnR-dependent, Gq-coupled receptor activation of 2-AG synthesis. While a Gq-coupled receptor promotion of endocannabinoid synthesis has been well established during the last decade, our results reveal a surprising link between synaptically released Zn2+ and retrograde endocannabinoid signaling, thus providing a significant advance toward the elucidation of the role of Zn2+ in synaptic physiology.

Quantal properties, baseline Pr and depolarization-induced endocannabinoid signaling are not altered in ZnT3 KO mice. A, Top, Sample traces of mEPSCs from ZnT3 WT and KO mice. Cumulative probability plot of mEPSC frequencies shows that frequencies are not different between ZnT3 WT and KO mice (mean mEPSC frequency WT: 3.25 ± 0.60 Hz, n = 5; KO: 3.57 ± 0.73 Hz; n = 5, p = 0.74). B, Cumulative probability plot of mEPSC amplitudes shows that amplitudes are not different between ZnT3 WT and KO mice (mean mEPSC amplitude WT: 17.50 ± 1.2 pA, n = 5; KO: 16.7 ± 0.33 pA; n = 5, p = 0.54). C, Cumulative probability plot of mEPSC rise times showing that rise times are not different between ZnT3 WT and KO mice (mean mEPSC rise time: WT: 0.67 ± 0.06 ms, n = 5; KO: 0.59 ± 0.03 ms; n = 5, p = 0.3). D, Cumulative probability plot of mEPSC decay times showing that decay times are not different between ZnT3 WT and KO mice (mean mEPSC decay time WT: 1.44 ± 0.08 ms, n = 5; KO: 1.42 ± 0.07 ms; n = 5, p = 0.84). E, PPR is similar in ZnT3 WT and KO littermates (PPR WT: 2.13 ± 0.15, n = 7; KO: 2.16 ± 0.11, n = 7, p = 0.85). F, DSE is similar in ZnT3 WT and KO mice. Time course of DSE, induced by a 5 s depolarization to 10 mV (DSE WT: 33.5 ± 4.9% of control, n = 5; KO DSE: 35 ± 3.2% of control, p = 0.54). Summary data represent mean ± SEM.
Discussion
We report that synaptic Zn2+ modulates probability of release of glutamatergic terminals via activation of retrograde endocannabinoid signaling. We propose that Zn2+-mediated modulation of Pr via endocannabinoid signaling reveals a previously unrecognized synaptic signaling pathway that includes presynaptic ZnT3 and CB1Rs as well as postsynaptic mZnRs, PLC and DGL. This signaling pathway provides a fundamental substrate for both neuromodulatory and state-dependent regulation of neuronal properties, thus shaping synaptic strength in an activity-dependent manner.
The role of Zn2+ and mZnR in modulating synaptic transmission
Our studies provide the first evidence for inhibitory effects of Zn2+-mediated signaling on Pr. We demonstrate a synaptic Zn2+-mediated negative feedback loop that reduces Pr at DCN parallel fiber synapses through a signaling pathway linking Zn2+ release to endocannabinoid synthesis. Bath application of exogenous Zn2+ as well as release of endogenous Zn2+ caused a reversible reduction of Pr. This inhibitory effect of Zn2+ is opposite to the effect that has been observed in mossy fiber synapses (Li et al., 2001; Huang et al., 2008), where exogenous Zn2+ application leads to LTP of synaptic response via presynaptic increases in Pr (Huang et al., 2008; Pan et al., 2011). Given that mZnRs are likely present in mossy fiber synapses (Besser et al., 2009; Chorin et al., 2011), the lack of the inhibitory effect of Zn2+ on Pr is likely due to the absence of endocannabinoid signaling in mossy fiber synapses (Hofmann et al., 2008). Our results suggest that the synaptic effects of Zn2+ are synapse specific and, similar to other neuromodulatory systems, Zn2+-mediated effects depend on the molecular composition of specific synapses.
While previous studies have shown that Zn2+ is released in an activity and Ca2+-dependent manner (Qian and Noebels, 2005, 2006), there has been a controversy about whether synaptic Zn2+ is released or whether Zn2+ creates a layer in the extracellular space that maps onto the synaptic Zn2+ stained region (Zn2+ veneer theory) (Kay, 2003; Kay and Tóth, 2008). Indeed, the amount of synaptic Zn2+ that reaches the postsynaptic membrane during synaptic stimulation remains unknown with estimates from ∼0 or no Zn2+ release (Kay, 2003; Kay and Tóth, 2008), to >100 μm (Vogt et al., 2000). Here, we focused on determining whether synaptically released (endogenous) Zn2+, regardless of the actual concentration reached in synaptic cleft, was sufficient to trigger endocannabinoid signaling. The results presented in Figures 9 and 11 show that endogenous Zn2+ reduces Pr via endocannabinoid signaling in the DCN. Thus, our pharmacological and genetic manipulations support a ZnT3/mZnR–dependent, endocannabinoid-mediated decrease of Pr as a result of release of endogenous Zn2+ in response to brief stimulus trains.
While recent results have indicated that synaptically released Zn2+ induced by a single action potential blocks NMDAR responses (Pan et al., 2011), our results show that mZnR-dependent triggering of endocannabinoid signaling requires trains of presynaptic action potentials. The requirement for trains of presynaptic action potentials is consistent with the recent proposal that Zn2+-containing synaptic mossy fiber vesicles populate the reserve, not the readily released, pool (Lavoie et al., 2011). Although the detailed physiological conditions leading to mZnR-mediated signaling remain undetermined, our findings suggest that, in response to a brief 30 Hz train, endogenous Zn2+ levels set the threshold for eliciting short-term plasticity (Figs. 9B1, 11C). While in vivo recordings from DCN granule cells have not been obtained, sensory stimulation can produce bursts of spikes in cerebellar granule cells at very high firing frequencies (>200 Hz) (Chadderton et al., 2004). Given the strong parallels between DCN and cerebellar granule cells (Oertel and Young, 2004), the brief 30 Hz stimulation that we used is expected to be well within the physiological range of parallel fiber activity, thus suggesting that Zn2+-mediated endocannabinoid pathway can be engaged by physiologically relevant stimuli.
Our results demonstrate that GPR39 (mZnR) is necessary for Zn2+-mediated endocannabinoid synthesis; it is likely that synaptically released Zn2+ mediates its effects on endocannabinoid signaling and Pr via direct activation of GPR39 (mZnR; Fig. 13). GPR39, a previously orphan GPCR was initially proposed to mediate obestatin signaling and to regulate food intake (Zhang et al., 2005). However, the absence of GPR39 in the hypothalamus (Jackson et al., 2006) and the identification of Zn2+, rather than obestatin, as its putative endogenous ligand (Yasuda et al., 2007) have raised questions regarding the physiological role of GPR39. Recent studies have suggested that synaptically released Zn2+ triggers metabotropic signaling via mZnR in the hippocampus (Besser et al., 2009), enhancing the activity of the potassium chloride cotransporter 2 in CA3 pyramidal neurons (Chorin et al., 2011). We show that during synaptic activation of excitatory terminals, similar to group 1 mGluRs (Maejima et al., 2001; Varma et al., 2001; Kushmerick et al., 2004), the mZnR is necessary for 2-AG synthesis and thus, contributes to the overall signaling profile of excitatory synaptic transmission that is tightly coupled to postsynaptic endocannabinoid synthesis. Because the 2-AG synthesizing enzymes, DGLα and DGLβ, have been localized to the dendrites of DCN fusiform cells (Zhao et al., 2009), our electrophysiological, imaging, and biochemical results are consistent with a postsynaptic localization of mZnRs.

Schematic representation illustrating the mechanism of Zn2+-induced decrease in Pr. Synaptically released Zn2+, mZnR activation, rises in postsynaptic Ca2+ and PLC stimulation are necessary for 2-AG synthesis. 2-AG is produced by cleavage of diacylglycerol by DGL. 2-AG diffuses to the presynaptic terminal where it decreases Pr.
The role of Zn2+ in brain function
The allosteric modulation of NMDA and glycine receptors by Zn2+ has suggested a role for Zn2+ in learning, and motor control, respectively (Hirzel et al., 2006; Adlard et al., 2010). Synaptically released Zn2+ has also been associated with Alzheimer's disease and pain processing (Deshpande et al., 2009; Duce et al., 2010; Nozaki et al., 2011). Although initial studies did not uncover cognitive deficits in ZnT3KO mice (Cole et al., 2001), recent evidence reveals impaired fear memory (Martel et al., 2010), impaired spatial memory and impaired behavior dependent on hippocampus and perirhinal cortex (Martel et al., 2011), accelerated aging-related decline of spatial memory (Adlard et al., 2010), as well as impairments in spatial working memory and contextual discrimination (Sindreu et al., 2011). Our studies, by demonstrating a Zn2+-mediated negative feedback loop that reduces Pr at central auditory synapses—through a specific pathway linking mZnR activation, endocannabinoid synthesis, and Pr inhibition—pave the way for novel in vivo and behavioral experimental paradigms aimed at further defining the role of Zn2+ on brain function.
In sensory systems, the levels of synaptic Zn2+ in somatosensory cortex can be rapidly and dynamically regulated by sensory experience, although the physiological consequences of these changes have not been determined (Brown and Dyck, 2003; Nakashima and Dyck, 2009, 2010). Although it has been known for 25 years that DCN parallel fibers contain very large amounts of synaptic Zn2+ (Frederickson et al., 1988), the physiological role of this metal in modulating auditory circuits has not been previously evaluated. Based on our observations, we propose that synaptic Zn2+, by providing a negative feedback loop that reduces synaptic strength, may decrease the responsiveness of the cochlear nucleus to acoustic stimuli: thus, synaptic Zn2+ release is expected to reduce the gain of input–output transformations between the auditory nerve and the cochlear nucleus. This is consistent with the proposed inhibitory role of Zn2+ in visual and auditory processing centers (Hirzel et al., 2006; Chappell et al., 2008), thus supporting the notion that Zn2+ is an inhibitory neuromodulator in sensory systems.
In conclusion, our results reveal the existence of a cellular pathway linking synaptically released Zn2+ and endocannabinoids: a previously unrecognized connection between two fundamental neuromodulatory systems, leading to activity-dependent inhibition of synaptic strength.
Footnotes
This work was supported by National Institutes of Health (NIH) Grants DC007905 (T.T.), NS043277 (E.A.), HL058115 (B.A.F.), HL64937 (B.A.F.), DK072506 (B.A.F.), HL103455 (B.A.F.), and AT006822 (F.J.S.); by Hemsley Trust Grant BSF2011126 from the U.S.–Israel Binational Science Foundation (E.A., M.H.); and by NIH Training Grant F32DC011664 (T.P.-R.). We thank Veronica Choi, Courtney Pedersen, and Karen Hartnett for technical assistance. We thank Drs. Karl Kandler, Carlos Aizenman, and Paul Rosenberg for critical reading of earlier versions of this manuscript. GPR39/mZnR KO mice were kindly provided by Dr. Moechars from the Janssen Pharmaceutical companies of Johnson & Johnson.
- Correspondence should be addressed to Thanos Tzounopoulos, Department of Otolaryngology, University of Pittsburgh School of Medicine, 10021 BST3, Pittsburgh, PA 15261. thanos{at}pitt.edu





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}