

Abstract
The embryonic sympathetic nervous system consists of predominantly noradrenergic neurons and a very small population of cholinergic neurons. Postnatal development further allows target-dependent switch of a subset of noradrenergic neurons into cholinergic phenotype. How embryonic cholinergic neurons are specified at the prenatal stages remains largely unknown. In this study, we found that the expression of transcription factor Tlx3 was progressively restricted to a small population of embryonic sympathetic neurons in mice. Immunostaining for vesicular acetylcholine transporter (VAChT) showed that Tlx3 was highly expressed in cholinergic neurons at the late embryonic stage E18.5. Deletion of Tlx3 resulted in the loss of Vacht expression at E18.5 but not E12.5. By contrast, Tlx3 was required for expression of the cholinergic peptide vasoactive intestinal polypeptide (VIP), and somatostatin (SOM) at both E12.5 and E18.5. Furthermore, we found that, at E18.5 these putative cholinergic neurons expressed glial cell line-derived neurotrophic factor family coreceptor Ret but not tyrosine hydroxylase (Ret+/TH−). Deletion of Tlx3 also resulted in disappearance of high-level Ret expression. Last, unlike Tlx3, Ret was required for the expression of VIP and SOM at E18.5 but not E12.5. Together, these results indicate that transcription factor Tlx3 is required for the acquisition of cholinergic phenotype at the late embryonic stage as well as the expression and maintenance of cholinergic peptides VIP and SOM throughout prenatal development of mouse sympathetic neurons.
Introduction
The sympathetic nervous system is a major component of the autonomic nervous system that maintains the homeostasis of the internal environment during diverse physiological conditions (Cannon, 1939; Janig and McLachlan, 1992). Mature mammalian and avian sympathetic ganglia are composed of two types of neurons. The vast majority of sympathetic neurons are noradrenergic and express molecular markers, such as tyrosine hydroxylase (TH) and dopamine β-hydroxylase (DBH). By contrast, a small subset of sympathetic neurons use acetylcholine as transmitter and can be discriminated by the expression of choline acetyltransferase (ChAT) and vesicular acetylcholine transporter (VAChT) (Elfvin et al., 1993; Ernsberger and Rohrer, 1999). Diversity of mature sympathetic neurons is also indicated by the restricted expression of peptides in distinct subpopulations (Elfvin et al., 1993; Gibbins, 1995). How the diversity of the sympathetic neurons is generated during development is a major question for studying neural development (Le Douarin and Kalcheim, 1999; Howard, 2005; Huber, 2006).
Substantial progress has been made in understanding the extrinsic signals and intrinsic transcription factors that control the acquisition of noradrenergic phenotype in sympathetic neurons (Apostolova and Dechant, 2009; Cane and Anderson, 2009; Rohrer, 2011). Transcription factors Phox2b and Hand2 are required to maintain the noradrenergic traits, whereas Gata3 is required for the survival of differentiated noradrenergic neurons (Schmidt et al., 2009; Coppola et al., 2010; Tsarovina et al., 2010). For cholinergic differentiation in sympathetic neurons, there are two modes of how sympathetic neurons acquire their cholinergic phenotype (Ernsberger and Rohrer, 1999; Francis and Landis, 1999; Ernsberger, 2001). In the classic postnatal target-dependent cholinergic differentiation, the sudomotor neurons and the neurons that innervate the periosteum initially express noradrenergic phenotype, but acquire the cholinergic phenotype while losing functional adrenergic properties after contacting the target tissue (Landis, 1990; Asmus et al., 2000). The switch from noradrenergic to cholinergic phenotype is initiated by signals released from the target (e.g., sweat glands) (Apostolova and Dechant, 2009; Cane and Anderson, 2009). The glycoprotein (gp)130 receptor that is expressed in sympathetic neurons is indispensable for the target-dependent acquisition of cholinergic phenotype (Stanke et al., 2006). Prenatal expression of cholinergic properties has also been found in a very small population of developing sympathetic neurons of rodents and chick (Ernsberger et al., 1997; Schäfer et al., 1997; Schütz et al., 1998). Further studies revealed that transcription factor Phox2b and Ret, the glial cell line-derived neurotrophic factor (GDNF) family ligands (GFLs) coreceptor, are required for this embryonic cholinergic differentiation (Burau et al., 2004; Huber and Ernsberger, 2006). However, the molecular mechanism that controls the development of embryonic cholinergic neurons remains largely unknown.
The homeodomain transcription factor Tlx3 and Tlx1 determine glutamatergic over GABAergic cell fates and the expression of corresponding peptides and transmitter receptors in the dorsal spinal cord (Cheng et al., 2004, 2005; Xu et al., 2008; Guo et al., 2012; Hu et al., 2012). Here, we report that Tlx3 is enriched in the cholinergic subset of sympathetic neurons at the late embryonic stage and is required for their proper phenotypic development.
Materials and Methods
Mouse strains.
The generation of Tlx3 mutant mice was described previously (Shirasawa et al., 2000). Ret mutant mice were kindly donated by Dr. Frank Costantini (Columbia University Medical Center, New York, NY) and obtained from The Jackson Laboratory (Schuchardt et al., 1994). In all timed mating using both male and female mice, the morning that vaginal plugs were observed was designated as E0.5. Genotyping for Tlx3 mutant mice was done as described previously (Qian et al., 2002). Wild-type Ret allele was identified with the information provided by The Jackson Laboratory and the mutant Ret allele was amplified with the following primers that produce a 651 bp product: 5'-AGTCTCCAAGGCAATTTCAACG-3' and 5'-GCCCAATAGCAGCCAGTCC-3'. All animal procedures are contained in protocols reviewed and approved by the Animal Care Committee at the Institute of Neuroscience, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences.
In situ hybridization and immunostaining.
In situ hybridization experiments were performed as described previously (Huang et al., 2010). Riboprobe for mouse Vacht (NM_021712, 1.14 Kb) and Vip (NM_011702, 1.04 kb) were used. Sst in situ probe was described previously (Huang et al., 2008).
Immunostaining was performed as described previously (Huang et al., 2010). The following antibodies were used: rabbit anti-Tlx3 antibody (1:400) (Huang et al., 2010), guinea pig anti-Tlx3 (1:1000, kind gift from C. Birchmeier, Max Delbrück Center for Molecular Medicine, Berlin, Germany), goat anti-VAChT (1:50, Santa Cruz Biotechnology), rabbit anti-VAChT (1:500, SYSY), goat anti-VIP (1:200, Santa Cruz Biotechnology), goat anti-SOM (D-20; 1:300, Santa Cruz Biotechnology), mouse anti-TH (1:500, Sigma-Aldrich), rabbit anti-TH (1:500, Pel-Freez), goat anti-Ret (1:25, R&D Systems), goat anti-Phox2b (1:50, Santa Cruz Biotechnology), rabbit anti-SOM (H-106) (1:50, Santa Cruz Biotechnology), mouse anti-CGRP (1:500, Sigma-Aldrich).
Cell counting.
Stellate ganglion from three pairs of control and mutant embryos (10 μm) were analyzed with various molecular markers. Immunoreactive cells with signal highly above the background were counted and the values were presented as mean ±SEM. The differences in values were considered to be significant at p < 0.05 by Student's t test.
Results
Expression of Tlx3 in sympathetic cholinergic neurons
To study the expression of transcription factor Tlx3 in the developing sympathetic neurons, we performed immunostaining experiments on sympathetic ganglia of mouse embryos from E9.5 to E18.5. The first Tlx3+ cells were detectable at E10.5 and found to coexpress Phox2b, which is known to be expressed in early sympathetic progenitors (Pattyn et al., 1997, 1999), indicating that Tlx3 was highly expressed in developing sympathetic ganglia (Fig. 1A–F). Approximately 82% and 95% Phox2b+ cells expressed Tlx3 at E10.5 and E12.5, respectively (Fig. 1J). Over the next few days, Tlx3 expression was progressively restricted to a small population of sympathetic neurons (Fig. 1G-I). At E16.5 and E18.5, the percentage of Phox2b+ neurons that expressed Tlx3 reduced to 17% and 6%, respectively (Fig. 1J). This progressive restriction of Tlx3 expression in sympathetic neurons was reminiscent of the restrictive expression pattern of cholinergic marker genes in rodent and chick sympathetic neurons at prenatal stages (Ernsberger et al., 1997; Schütz et al., 1998; Burau et al., 2004; Huber and Ernsberger, 2006) (Fig. 2). Furthermore, gene expression studies performed in chicken sympathetic ganglia have shown that Tlx3 is among the genes upregulated by factors that induce cholinergic neuronal differentiation in vitro (Apostolova et al., 2007). To determine whether Tlx3 is associated with cholinergic neuronal differentiation in developing sympathetic ganglia, we performed triple staining of the stellate ganglia at embryonic stages, where a small population of cholinergic neurons has been observed at the prenatal stages (Schäfer et al., 1997; Schütz et al., 1998). VAChT and ChAT are well known molecular markers for cholinergic neurons (Ernsberger and Rohrer, 1999). In situ hybridization studies have shown that the expression level of Vacht is higher than that of Chat in sympathetic neurons (Burau et al., 2004). Thus, we focused on VAChT expression in this study. Immunostaining results showed that VAChT and TH were coexpressed in many neurons at E12.5, but in largely separated populations by E18.5 (Fig. 1K–N and data not shown). Meanwhile, the majority of VAChT+ neurons (82%) expressed Tlx3 at E18.5 (Fig. 1K–N). Vasoactive intestinal polypeptide (VIP) is known to be expressed in cholinergic neurons of stellate ganglia in newborn mice (Stanke et al., 2000). Indeed, our immunostaining results showed that the majority of VIP+ neurons (80%) expressed Tlx3 (Fig. 1O–R). Thus, most of the cholinergic neurons express Tlx3 during late embryonic stages.

Expression of Tlx3 in the embryonic sympathetic ganglia of mouse. A–I, Immunostaining of Tlx3 was performed on transverse sections of the primary sympathetic chain from wild-type mice at E10.5 (A), and stellate ganglia at E12.5 (D), E14.5 (G), E16.5 (H), and E18.5 (I). Double staining of Tlx3 and Phox2b was performed at E10.5 (A–C) and E12.5 (D–F). J, The number of Tlx3+ cells was determined in sympathetic ganglia at E10.5, E12.5, E16.5, and E18.5 relative to the total number of Phox2b+ cells. K–R, Triple immunostaining of Tlx3, TH, and VAChT (K–N) or VIP (O–R) was performed on sections of stellate ganglia at E18.5. Arrows (K–M, O–Q) and arrowheads (N, R) showed the majority of VAChT+/TH− and VIP+/TH− cells coexpressed Tlx3. Scale bars: A–F, K–R, 10 μm; G–I, 20 μm.

Different expression patterns of cholinergic and noradrenergic neuronal markers during embryonic sympathetic development. A–F, In situ hybridization was performed on sections of stellate ganglia from wild-type mice at E12.5, E16.5, and E18.5 with Vacht (A–C), and Vip (D–F) as the probes. Note a progressively restricted expression pattern of Vacht. G–I, Immunostaining of TH was performed on sections of stellate ganglia from wild-type mice at E12.5, E16.5, and E18.5. The persistently widespread expression of TH was observed during embryonic sympathetic development. Scale bars: A, D, G, 20 μm; B, C, E, F, H, I, 50 μm.
It has been reported that cholinergic sympathetic neurons in adult stellate ganglia that innervate sweat glands acquire cholinergic properties largely between P4 and P21 through target-dependent gp 130 signaling (Guidry and Landis, 1998; Ernsberger and Rohrer, 1999; Francis and Landis, 1999; Stanke et al., 2006). Those sudomotor neurons are characterized by the expression of calcitonin gene-related peptide (CGRP) (Landis and Fredieu, 1986; Anderson et al., 2006). To study whether Tlx3 was expressed in the second-wave of cholinergic neurons, we performed immunostaining experiments on postnatal stellate ganglia. The results showed that Tlx3 was clearly expressed in a subset of sympathetic neurons at P14 and P60 (Fig. 3A,B). Furthermore, double staining results showed that Tlx3+ neurons expressed CGRP at P14 and P30 (Fig. 3C–H). Tlx3 also was nicely colocalized with VIP in sympathetic neurons: most Tlx3+ neurons (87%) expressed VIP and ∼42% VIP+ neurons expressed Tlx3 at P30 (Fig. 3I–N). Meanwhile, double staining results revealed that almost all of the VIP+ neurons coexpressed VAChT at P14 and P60 (data not shown). A recent study also showed that Tlx3 was expressed in cholinergic neurons at P60 (Furlan et al., 2013). Together, Tlx3 was expressed in the postnatal emergence of cholinergic neurons as well.

Expression of Tlx3 in the postnatal sympathetic ganglia of mouse. A, B, Immunostaining of Tlx3 was performed on sections of stellate ganglia from wild-type mice at P14 (A) and P60 (B).C–H, Double immunostaining of Tlx3 and CGRP was performed on sections of stellate ganglia from wild-type mice at P14 (C–E) and P30 (F–H). I–N, Double immunostaining of Tlx3 and VIP was performed on sections of stellate ganglia from wild-type mice at P14 (I–K) and P30 (L–N). Scale bars: C–N, 10 μm; A, B, 50 μm.
Tlx3 controls development of cholinergic neurons
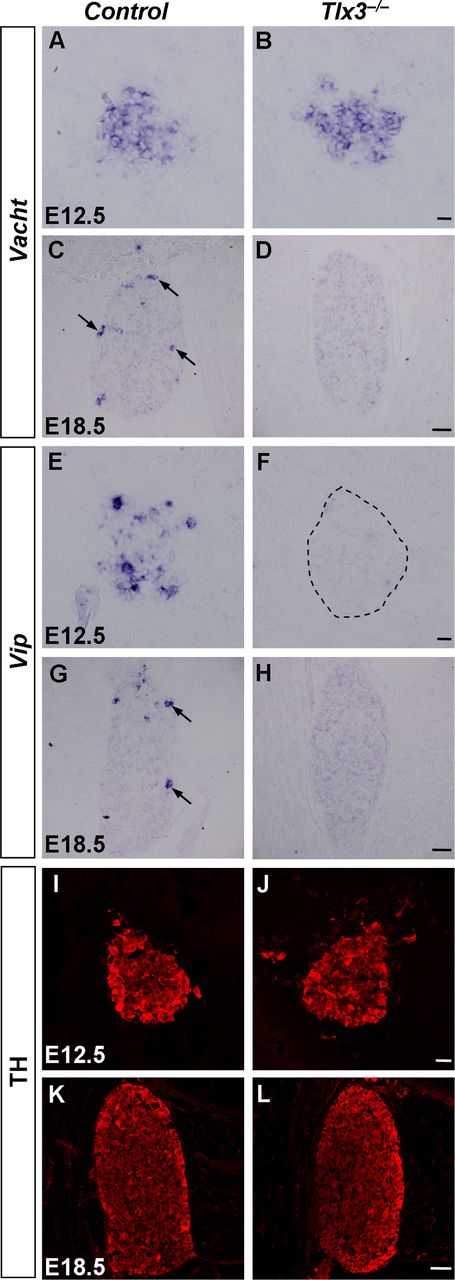
To determine whether Tlx3 was required for the development of cholinergic neurons in stellate ganglia, we examined the expression of Vacht and Vip in control (wild-type or Tlx3+/−) and Tlx3 deletion (Tlx3−/−) mice by in situ hybridization. The results showed that the early expression of Vacht at E12.5 was similar between control and Tlx3−/− mice, whereas the expression of Vacht expression at E18.5 was abolished in Tlx3−/− mice but not in control mice (Fig. 4A–D). In contrast, the expression of Vip was eliminated in Tlx3−/− mice at both E12.5 and E18.5 (Fig. 4E–H). To determine whether Tlx3 controlled the development of noradrenergic neurons in the stellate ganglion, we examined the expression of TH and transcription factors that are required for the development of noradrenergic neurons, including Phox2b, Hand2, Gata3, and Insm1, in the Tlx3−/− mice at E12.5 and E18.5. The results showed that the expression of these molecular markers were largely unaffected when Tlx3 was fully ablated (Figs. 4I–L, 5).

Loss of cholinergic properties in stellate ganglia of Tlx3−/− mice. A–H, In situ hybridization was performed on sections of stellate ganglia from control and Tlx3−/− mice at E12.5 and E18.5 with Vacht (A–D), and Vip (E–H) as the probes. The Vacht and Vip signals were absent in Tlx3−/− mice at E18.5. Note that the expression of Vacht was comparable in Tlx3−/− versus control mice, whereas the expression of Vip was abolished in Tlx3−/− mice early at E12.5. I–L, Immunostaining of TH was performed on sections of stellate ganglia from control and Tlx3−/− mice at E12.5 and E18.5. No apparent difference was observed between Tlx3 mutant and control mice. Scale bara: A, B, E, F, I, J, 20 μm; C, D, G, H, K, L, 50 μm.

Expression of some molecular markers in Tlx3−/− mice. A–D, Immunostaining of Phox2b was performed on sections of stellate ganglia from control and Tlx3−/− mice at E12.5 and E18.5.E–P, In situ hybridization was performed on sections of stellate ganglia from control and Tlx3−/− mice at E12.5 and E18.5 with Hand2 (E–H), Gata3 (I–L), and Insm1 (M–P) as the probes. No apparent differences were seen in the control and Tlx3−/− mice. Scale bars: A, B, E, F, I, J, M, N, 20 μm; C, D, G, H, K, L, O, P, 50 μm.
Some VIP-expressing neurons in the stellate ganglia of rat are also known to express the neuropeptide somatostatin (SOM) (Masliukov and Timmermans, 2004). To determine whether SOM or the gene Sst that encodes SOM was expressed in embryonic cholinergic neurons, we performed in situ hybridization and immunostaining experiments. The results showed that Sst was consistently expressed in a small portion of neurons in embryonic stellate ganglia (Fig. 6A–D). Furthermore, most of the VAChT-expressing cholinergic neurons (83%) expressed SOM and ∼64% of VIP+ neurons expressed SOM at E18.5 (Fig. 6E–L). We also found that most SOM+ neurons (89%) expressed Tlx3 at E18.5 (Fig. 6M–P). Finally, we examined whether Tlx3 was required for the expression of SOM or Sst and found that the expression of SOM or Sst at both E12.5 and E18.5 was eliminated in Tlx3−/− mice (Fig. 6Q–T). Together, these results showed that Tlx3 is required for the embryonic development of cholinergic neurons in the stellate ganglion by specifying the expression of Vacht, Vip, and SOM. The neonatal lethality of Tlx3 mutant mice (Shirasawa et al., 2000) prevented us from studying whether Tlx3 controls the development of the second-wave of cholinergic neurons at the postnatal stages.

SOM or Sst was expressed in cholinergic sympathetic neurons and their expression was abolished in Tlx3−/− mice. A-D, In situ hybridization was performed on sections of primary sympathetic chain at E11.5 (A), and stellate ganglia at E12.5 (B), E16.5 (C), and E18.5 (D) with Sst as the probe. Strong signals were detected in a small subset of neurons in the embryonic sympathetic ganglia. E–H, Triple immunostaining of SOM, VAChT, and TH was performed on sections of stellate ganglia from wild-type mice at E18.5. The majority of SOM+ cells were VAChT+/TH− cholinergic neurons (arrows and arrowheads). I–L, Triple immunostaining of SOM, VIP, and TH was performed on sections of stellate ganglia from wild-type mice at E18.5. A large portion of VIP+/TH− cells expressed SOM (arrows and arrowheads). M–P, Triple immunostaining of Tlx3, SOM, and TH was performed on sections of stellate ganglia from wild-type mice at E18.5. Most SOM+/TH− cells expressed Tlx3 (arrows and arrowheads). Q–T, Expression of SOM or Sst in the stellate ganglia from Tlx3−/− mice. Double-staining of SOM and TH was performed on sections of E12.5 stellate ganglia from control and Tlx3−/− mice (Q, R). In situ hybridization was performed on sections of stellate ganglia from control and Tlx3−/− mice at E18.5 (S, T). Note a complete loss of expression of SOM or Sst in Tlx3−/− mice at both E12.5 and E18.5.Scale bars: E-R, 10 μm; A–C, 20 μm; D, S, T, 50 μm.
Expression of Ret in developing stellate ganglia of Tlx3 deletion mice
Ret has been shown to regulate cholinergic properties in embryonic mouse sympathetic neurons (Burau et al., 2004). To determine the relationship between Tlx3 and Ret in cholinergic development, we first examined whether they are colocalized in the same cells. Immunostaining results showed that the majority of Ret+/TH− neurons (87%) expressed Tlx3 in wild-type mice (Fig. 7A–D). Triple staining results further showed that most Ret+/TH− neurons (92%) were indeed cholinergic neurons (Fig. 7E–H). Then we examined whether Tlx3 was required for the expression of Ret in the putative cholinergic (Ret+/TH−) neurons by analyzing the Tlx3−/− mice. Interestingly, the small number of putative cholinergic (Ret+/TH−) neurons disappeared, whereas the majority of Ret+/TH+ neurons were unaffected in Tlx3−/− mice at E18.5 (Fig. 7I–L, arrows). It was unclear whether the disappearance of cholinergic (Ret+/TH−) neurons in Tlx3−/− mice was due to the downregulation of high-level expression of Ret in these neurons or cell death. The relatively constant number of Ret+ neurons in control and Tlx3 deletion mice (control: 70 ± 2 vs Tlx3−/−: 71.3 ± 1.4 cells/section) suggested that Ret+/TH− neurons may have changed their fate from cholinergic to noradrenergic phenotype (Ret+/TH+) by gaining the expression of TH (Fig. 7I,K). Because the number of Ret+/TH− neurons was small (∼8 neurons/section), it remains to be determined whether cell type switch had indeed occurred. Nevertheless, the above results indicated that Tlx3 is required to maintain the high-level expression of Ret in embryonic cholinergic neurons in the sympathetic ganglion.

Tlx3 and Ret in regulating the acquisition of cholinergic properties. A–D, Triple immunostaining of Tlx3, Ret, and TH was performed on sections of stellate ganglia from wild-type mice at E18.5. E–H, Triple immunostaining of VAChT, Ret, and TH was performed on sections of stellate ganglia from wild-type mice at E18.5. I–L, Double-staining of Ret and TH was performed on sections of stellate ganglia from control and Tlx3−/− mice at E18.5 (I, K). The expression of Ret in the control and Tlx3−/− mice was shown in (J, L). M–T, In situ hybridization was performed on sections of stellate ganglia from control and Ret−/− mice at E12.5 and E18.5 with Sst (M–P), and Vip (Q–T) as the probes. Note that the expression of Sst and Vip was detectable in Ret−/− mice at E12.5, but abolished at E18.5. Scale bars: A–H, 10 μm; M, N, Q, R, 20 μm; I–L, O, P, S, T, 50 μm.
Ret controls peptide genes Sst and Vip in developing stellate ganglia
To determine whether Ret is required for the expression of peptides Vip and Sst, we performed in situ hybridization experiments. The results showed that the expression of Vip and Sst remained present in Ret−/− mice at E12.5 whereas their expression was eliminated by E18.5 (Fig. 7M–T). This indicated that Ret coordinately controlled the expression of transmitter and peptide phenotypes in stellate ganglia at the late embryonic stage. This is distinct from the action of Tlx3, which controls the expression of these peptides since E12.5.
Discussion
In this study, we found that high-level expression of transcription factor Tlx3 was widely present in early embryonic stellate ganglionic neurons and became progressively enriched in cholinergic neurons at late embryonic and postnatal stages. Analysis of the Tlx3−/− mice revealed that Tlx3 was required for Vacht expression at E18.5 but not at E12.5, although it was required for the expression of cholinergic peptides VIP and SOM at both stages. Furthermore, we found that Tlx3 controlled the expression of Ret at a high level, and Ret expression was required for the expression of Vip and Sst mRNAs at the late prenatal stage. These results shed light on the molecular mechanisms underlying differentiation of cholinergic neurons in the embryonic sympathetic nervous system.
Transcriptional control of neurotransmitter phenotypes in sympathetic ganglia
Diverse sympathetic neurons are derived from the multipotent neural crest progenitors (Goridis and Rohrer, 2002). For the postnatal acquisition of cholinergic phenotype in sympathetic neurons, it is well known that target-derived signals are responsible for the switch of the noradrenergic phenotype to the cholinergic fate (Landis, 1990; Asmus et al., 2000). However, cholinergic phenotype was observed in a small population of embryonic sympathetic neurons (Ernsberger et al., 1997; Schäfer et al., 1997; Schütz et al., 1998). The development of these embryonic cholinergic neurons is still poorly understood. It has been reported that transcription factor Phox2b was required for the early cholinergic differentiation at E11.5, because cholinergic markers Chat and Vacht were absent in the sympathetic ganglia of Phox2b knock-out mice (Huber and Ernsberger, 2006). Because the expression of noradrenergic markers and generic neuronal properties were impaired in sympathetic neurons of these Phox2b knock-out mice (Pattyn et al., 1999), it is possible that a general deficiency of neuronal differentiation or cell death could underlie the absence of cholinergic markers in these mice (Huber and Ernsberger, 2006). Furthermore, a recent study reported that ongoing Phox2b expression was not required for the maintenance of cholinergic differentiation during embryogenesis (Coppola et al., 2010). Ret is also required for the acquisition of cholinergic properties in embryonic sympathetic neurons (Burau et al., 2004). In the present study, we showed that transcription factor Tlx3 controls the cholinergic neuronal development and is required for the expression of Ret at a high level (Figs. 4, 6, 7). Furthermore, we found that Ret controlled the expression of cholinergic peptide genes Vip and Sst in embryonic sympathetic neurons (Fig. 7M–T). The fate of prenatal cholinergic neurons in stellate ganglia and potential targets they innervate remain unknown (Guidry and Landis, 1998; Anderson et al., 2006; Schütz et al., 2008; Apostolova and Dechant, 2009; Cane and Anderson, 2009). Further studies on fate mapping and neuronal projection are needed to clarify these issues. What roles Tlx3 plays in controlling the postnatal emergence of cholinergic neurons remains unknown. Because Tlx3 knock-out mice die shortly after birth (Shirasawa et al., 2000), it also will be of interest to study whether Tlx3 is required for the acquisition of the postnatal cholinergic properties by analyzing the Tlx3 conditional knock-out mice.
Differential regulation of transmitter and peptide in cholinergic neurons
The peptide VIP is expressed in the cholinergic neurons of newborn mice (Stanke et al., 2000). We showed here that the peptide SOM was expressed in embryonic cholinergic neurons as well (Fig. 6E–H). By analyzing the gene deletion mice, we found that the expression of Vip and Sst mRNAs required Tlx3 but not Ret at the early embryonic stage (E12.5) (Figs. 4E,F, 6Q,R, 7M,N,Q,R), suggesting that Tlx3 may be required for the initiation whereas Ret is only needed for the maintenance of VIP and SOM expression. On the other hand, Tlx3 and Ret are both required for Vacht expression at the late (E18.5 or newborn mice) but not early (E12.5 or E13) embryonic stage (Burau et al., 2004) (Fig. 4A–D), suggesting these genes are both required for the maintenance but not the initiation of Vacht expression. Thus, cholinergic transmitter and the peptide phenotypes in the prenatal sympathetic ganglia are differentially regulated. Such differential regulation was also observed in the target-dependent acquisition of cholinergic properties as well. In chick sympathetic ganglia, the timing of Vip expression correlates with the innervation of peripheral targets by sympathetic fibers (Ernsberger et al., 1997). Interestingly, the expression of Vip but not Chat in these sympathetic ganglia depends on the cytokines signaling through heterodimeric gp130/LIFRβ receptors (Geissen et al., 1998; Duong et al., 2002). In contrast to that found in chick, gp130 signaling is essential for the postnatal target-dependent expression of both VIP and VAChT in sympathetic stellate ganglia of the mouse (Stanke et al., 2006). Two recent studies also reported differential dependence of the cholinergic loci Vacht and Chat vs Vip gene on Satb2 and p38 activities that are essential for cholinergic trans-differentiation of sympathetic neurons (Apostolova et al., 2010; Loy et al., 2011). Thus, differential regulation of transmitter and neuropeptide phenotypes may be a general phenomenon in both embryonic and postnatal development of cholinergic neurons.
Footnotes
This work was supported by the “Strategic Priority Research Program” of the Chinese Academy of Sciences (Grant No. XDA01020306), and the 973 Program from The Ministry of Science and Technology of China (2011 CBA00400). We thank Dr. M.-m. Poo for critical comments on this paper, and Drs S. Shirasawa and S. Korsmeyer for Tlx3-null mice.
- Correspondence should be addressed to Dr Leping Cheng, Institute of Neuroscience and State Key Laboratory of Neuroscience, Shanghai Institutes for Biological Sciences, Chinese Academy of Science, 320 Yue Yang Road,Shanghai 200031, China. lpcheng{at}ion.ac.cn





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}