

Abstract
Startling acoustic stimuli (SAS) can accelerate reaction times (“StartReact” effect), but the underlying mechanism remains unclear. Both direct release of a subcortically stored motor program and a subcortically mediated trigger for a cortically stored motor program have been hypothesized. To distinguish between these hypotheses, we examined the StartReact effect in humans with pure hereditary spastic paraplegia (HSP). Delayed reaction times in HSP patients in trials both with and without a SAS would argue in favor of a cortically stored response.
We instructed 12 HSP patients and 12 matched controls to respond as rapidly as possible to a visual imperative stimulus, in two different conditions: dorsiflexion of the dominant ankle; or flexion of the dominant wrist. In 25% of trials, a SAS was delivered simultaneously with the imperative stimulus. Before these tests, subjects received five SAS while standing to verify normal function of the reticulospinal tract in HSP.
Latencies of startle responses in sternocleidomastoid and tibialis anterior muscles were comparable between patients and controls. During the ankle dorsiflexion task, HSP patients had an average 19 ms delay in reaction times compared with controls. Administration of a SAS accelerated ankle dorsiflexion in both groups, but more so in the patients, which completely normalized their latencies. The wrist flexion task yielded no differences in onset latencies between HSP patients and controls.
The reticulospinal tract seems unaffected in HSP patients, because startle reflex onsets were normal. The corticospinal tract was affected, as reflected by delayed ankle dorsiflexion reaction times. These delayed onsets in HSP were normalized when the imperative stimulus was combined with a SAS, presumably through release of a subcortically stored motor program conveyed by the preserved reticulospinal tract.
Introduction
The startle reflex is an involuntary reaction to unexpected sensory input and is the fastest generalized motor reaction of humans and animals (Valls-Solé et al., 2008). Auditory startling stimuli can accelerate reaction times when delivered simultaneously with an imperative cue, a phenomenon known as “StartReact” (Valls-Solé et al., 1999). The underlying mechanism of this phenomenon is not completely clear (Serranová et al., 2012). One hypothesis to explain the StartReact effect is that a startling acoustic stimulus (SAS) acts as an additional stimulus on top of the imperative stimulus and thereby increases the energy of the sensory input, resulting in an acceleration of sensorimotor coupling. This is known as intersensory facilitation (Nickerson, 1973). The other and prevailing hypothesis for the StartReact effect is a direct release of a subcortically stored motor program by the SAS (Valls-Solé et al., 1999; Carlsen et al., 2004a), conveyed by the reticulospinal tract (Rothwell, 2006; Valls-Solé et al., 2008). This hypothesis assumes that during motor preparation, motor programs become represented in subcortical structures (Buford and Davidson, 2004; Schepens and Drew, 2004), which are then accessible to startle pathways. Yet, it has recently been proposed that the SAS could also act as a subcortically mediated trigger for a cortically stored motor program (Alibiglou and MacKinnon, 2012; Carlsen et al., 2012). This notion would imply that the cortically stored response is triggered without the usual cortical processing, and is conveyed by the corticospinal tract.
To distinguish between the abovementioned hypotheses, we have examined the StartReact effect in patients with a pure form of hereditary spastic paraplegia (HSP). HSP is a diverse group of inherited disorders that are clinically characterized by progressive spasticity, muscle weakness, and reduced proprioception of the lower extremities (Salinas et al., 2008). The common pathological feature of these conditions is retrograde axonal degeneration of the corticospinal tract and the posterior spinal columns, without cortical pathology (McDermott et al., 2000). The retrograde axonal degeneration is presumably due to abnormal axonal membrane trafficking processes, which primarily affect the distal parts of axons (Blackstone et al., 2011). Using transcranial magnetic stimulation, degeneration of the corticospinal tract in HSP is reflected by prolonged central motor conduction times, elevated cortical motor thresholds, and reduced amplitudes of motor-evoked potentials in the lower limbs (Pelosi et al., 1991; Polo et al., 1993; Bönsch et al., 2003; Nardone and Tezzon, 2003; Jørgensen et al., 2005; Sartucci et al., 2007). In contrast, motor conduction times, cortical motor thresholds and amplitudes of motor-evoked potentials to the arm muscles have been reported to be normal (Pelosi et al., 1991; Polo et al., 1993; Jørgensen et al., 2005; Sartucci et al., 2007). In HSP, the reticulospinal tract is generally assumed not to be affected by the retrograde degenerative process, but this has not been proven. Therefore, we first examined startle reflex latencies to verify the intact function of the reticulospinal tract in HSP.
Then, to differentiate between the hypotheses explaining the StartReact phenomenon, we used a simple reaction time paradigm involving either voluntary ankle dorsiflexion or voluntary wrist flexion. The retrograde corticospinal axonal degeneration in the patients with HSP was expected to affect responses in the legs, but not in the arms. For a direct release of a subcortically stored motor program to explain the StartReact effect, we expected two findings. First, we expected to observe delayed simple reaction times during dorsiflexion, because of the degenerated corticospinal tract. Second, when the imperative “go” signal would be combined with a SAS, we expected to see normal latencies as, in this hypothesis, the SAS is able to launch a subcortically stored motor program conveyed by the reticulospinal tract. In contrast, delayed dorsiflexion reaction times both with and without a SAS would argue in favor of a cortically stored response or intersensory facilitation as underlying mechanism of the StartReact phenomenon.
Materials and Methods
Participants.
Twelve patients with autosomal dominant forms of HSP (9 men; mean age 51 years, range 23–68 years) were recruited from all patients with HSP who were known at the outpatient departments of Neurology and Rehabilitation of our university hospital, a tertiary referral center for HSP. All 12 patients fulfilled the diagnostic clinical criteria for “pure” HSP (Salinas et al., 2008); 8 patients had previously been tested positive for pathogenic SPAST (SPG4) mutations, and 1 patient for a pathogenic KIAA0196 (SPG8) mutation. In the other three patients, mutations in genes most frequently associated with AD-HSP (ATL1, SPAST, and/or REEP1) had been excluded. All patients were able to walk independently. In addition, 12 aged-matched healthy controls (7 men, mean 49 years, range 23–65) participated.
Ethics statement.
The study was approved by the regional medical ethics committee (Commissie Mensgebonden Onderzoek Arnhem-Nijmegen) and was conducted in accordance with the Declaration of Helsinki. All subjects gave their written informed consent before the experiment.
Clinical assessment.
Muscle tone of the triceps surae (TS) and tibialis anterior (TA) muscles was assessed using the Modified Ashworth Scale (0–5), with higher scores indicating more hypertonia (Bohannon and Smith, 1987). The TS muscles were tested both with the knee flexed and extended. Muscle strength of the TS and the TA was assessed with the Medical Research Council scale (0–5), with lower scores indicating less muscle strength (Medical Research Council, 1981). We assessed the deep sensory modalities of the legs by testing the vibration sense at the lateral malleolus and at the first metatarsophalangeal joint using the semiquantitative tuning fork (0–8; Rydel Seiffer, Neurologicals, Poulsbo, Washington), with lower scores indicating more sensory loss (Pestronk et al., 2004). Vibration sense at the ankle and forefoot was averaged to obtain one value. For all measures, the mean of both legs was determined and used for further analysis (Table 1).
Clinical assessment of patients with HSP
Assessment of motor conduction time.
The motor conduction time to the dominant TA muscle was assessed in 11 patients before participation in the experiment. These motor conduction times were collected to be used in a parallel longitudinal study in the same patient group. One of the 12 patients did not want to undergo transcranial magnetic stimulation (TMS) of the lower limbs already at baseline, whereas several others refused to participate in follow-up TMS measurements. Therefore, we were unable to collect motor conduction times to the arm muscles. Magnetic stimulation of the cortex was performed using a double 110 mm cone coil, and for lumbar root stimulation a circular 90 mm coil was used, according to the International Federation of Clinical Neurophysiology guidelines (Rossini et al., 1994; Rothwell et al., 1999). All subjects were stimulated at 100% of the maximum stimulator output. Motor cortex stimulation was assessed with slight voluntary contraction of the TA, whereas spinal root stimulation was assessed at rest. The onset of TA activity after motor cortex stimulation was taken as the total motor conduction time (TMCT). Peripheral motor conduction time (PMCT) was obtained after spinal root stimulation. In one patient with HSP, we were unable to determine the PMCT. Corticospinal motor conduction time (CMCT) was assessed by subtracting the PMCT from the TMCT.
Experimental setup and protocol.
First, subjects received five SAS while standing. The SAS were given through binaural earphones and consisted of 50 ms white noise with an intensity of 116 dB (sound pressure level; SPL). The SAS was generated by a custom-made noise generator. Second, participants performed a warned simple reaction task. For this test, participants sat in a chair placed in front of two blocks with light-emitting diodes (LEDs). Illumination of the first LED block formed a warning signal and participants were instructed to perform ankle dorsiflexion with the dominant ankle as soon as the second LED block was lit (i.e., imperative stimulus). Warning periods (1–3.5 s) and intertrial periods (6–10 s) were variable. Participants performed 20 trials, in 25% of which a SAS was given simultaneously with the imperative stimulus. Third, participants performed another reaction time task where they had to flex the dominant wrist. The participant's arm was secured in a semiprone position with the palm facing inward, to a custom-made wrist manipulandum that moved in the transverse plane with an axis of rotation at the wrist joint (Carlsen et al., 2004b). Again, in 25% of the series of 20 trials a SAS was given simultaneously with the visual go signal.
Data collection.
Electromyographic (EMG) data were collected from the dominant TA muscle, dominant gastrocnemius medialis (GM) muscle, the dominant flexor carpi radialis (FCR), and left sternocleidomastoid (SCM) muscles (ZeroWire, Aurion). EMG signals were sampled at 2000 Hz and full-wave rectified and low-pass filtered at 30 Hz (zero-lag, second order Butterworth filter). The applied filtering technique resulted in systematic, small reduction of the detected latencies (on average 7 ms) for trials both with and without a SAS. Furthermore, a triaxial accelerometer was placed at the dominant foot and hand. Accelerometer data were collected to ensure that the SAS did not only result in shortened EMG onsets, but also in shortened movement onsets. Accelerometer signals were sampled at 2000 Hz and low-pass filtered at 30 Hz (zero-lag, second order Butterworth filter).
Data analysis.
Auditory startle reflexes were defined as the presence of a short latency response in the sternocleidomastoid muscle within 100 ms after the SAS (Brown et al., 1991b; Thevathasan et al., 2011). The response had to exceed, for at least 20 ms, a threshold of 2 SD above mean background activity, as calculated over a 500 ms period just before the SAS. Reflex activity in the TA muscle and FCR had to occur within 120 ms after the SAS to exclude any voluntary component (Brown et al., 1991a). For every participant we assessed for each muscle whether one or more startle reflexes occurred. For every muscle, the percentage of patients demonstrating at least one startle reflex is reported, in addition to the latency of the first occurring response.
Two reaction time parameters were assessed: EMG reaction time and accelerometer reaction time. Onset latencies of the muscles of interest were determined using a semiautomatic computer algorithm that selected the first instant at which the mean EMG activity exceeded a threshold of 2 SD above the mean background activity, as calculated over a 500 ms period just before the imperative go signal. Onsets were first selected by the computer algorithm, then visually approved and (when necessary) corrected (Carpenter et al., 2004; Queralt et al., 2008). Average EMG onset latencies were calculated separately for trials with and without a SAS. The onset of foot and wrist acceleration was determined in the same manner. For the ankle dorsiflexion task, we also calculated the interval between the EMG activity of the TA and GM by subtracting the TA onset from the GM onset.
Statistical analysis.
We tested for differences in onset latencies of startle reflexes between patients with HSP and controls using unpaired t tests. Differences in the rates of occurrence of startle reflexes between patients and controls were tested using a χ2 test. Reaction time parameters were analyzed using a repeated-measures ANOVA, with SAS (SAS-no SAS) as a within-subjects factor and group (HSP-controls) as a between-subjects factor. The α level was set at 0.05. In addition, the 95% confidence interval (CI) of the mean difference between patients and controls is presented, both for trials with and without a SAS. The statistical analyses were performed using SAS 9.2 for Windows (SAS Institute) and IBM SPSS Statistics 20 for Windows.
Results
Motor conduction times
TMCTs in patients with HSP were on average 35.8 ± 5.1(SD) ms and PMCTs were on average 15.6 ± 1.6 ms, resulting in mean CMCTs of 20.2 ± 5.1 ms (95% CI: 17.1–23.3 ms). These CMCT values were significantly delayed compared with reference data obtained in healthy subjects (13.8 ± 1.3 ms; 95% CI: 13.2–14.4 ms; Sartucci et al., 2007; and 13.8 ± 1.5 ms; 95% CI: 13.4–14.2 ms; Furby et al., 1992).
Startle reflex
SAS while standing clearly induced startle reflexes in the SCM muscle with similar onset latencies in patients with HSP (52 ± 10 ms, rate of occurrence 75%) and control subjects (52 ± 16 ms, rate of occurrence 92%). Neither onset latencies (t(18) = 0.72, p = 0.943) nor rates of occurrence (χ2(1) = 1.2, p = 0.273) differed between the groups. Startle reflexes were also seen in the wrist flexor, both in patients (75 ± 14 ms, rate of occurrence 67%) and controls (84 ± 20 ms, rate of occurrence 58%). The onset latencies (t(13) = 1.064, p = 0.307) and rates of occurrence (χ2(1) = 0.178, p = 0.673) did not differ between the groups either. In the TA muscle, we also recorded similar onset latencies between patients (87 ± 14 ms, rate of occurrence 50%) and controls (94 ± 17 ms, rate of occurrence 42%). Again, neither onset latencies (t(9) = 0.805, p = 0.442) nor rates of occurrence (χ2(1) = 0.168, p = 0.682) differed between the groups.
Ankle dorsiflexion reaction time
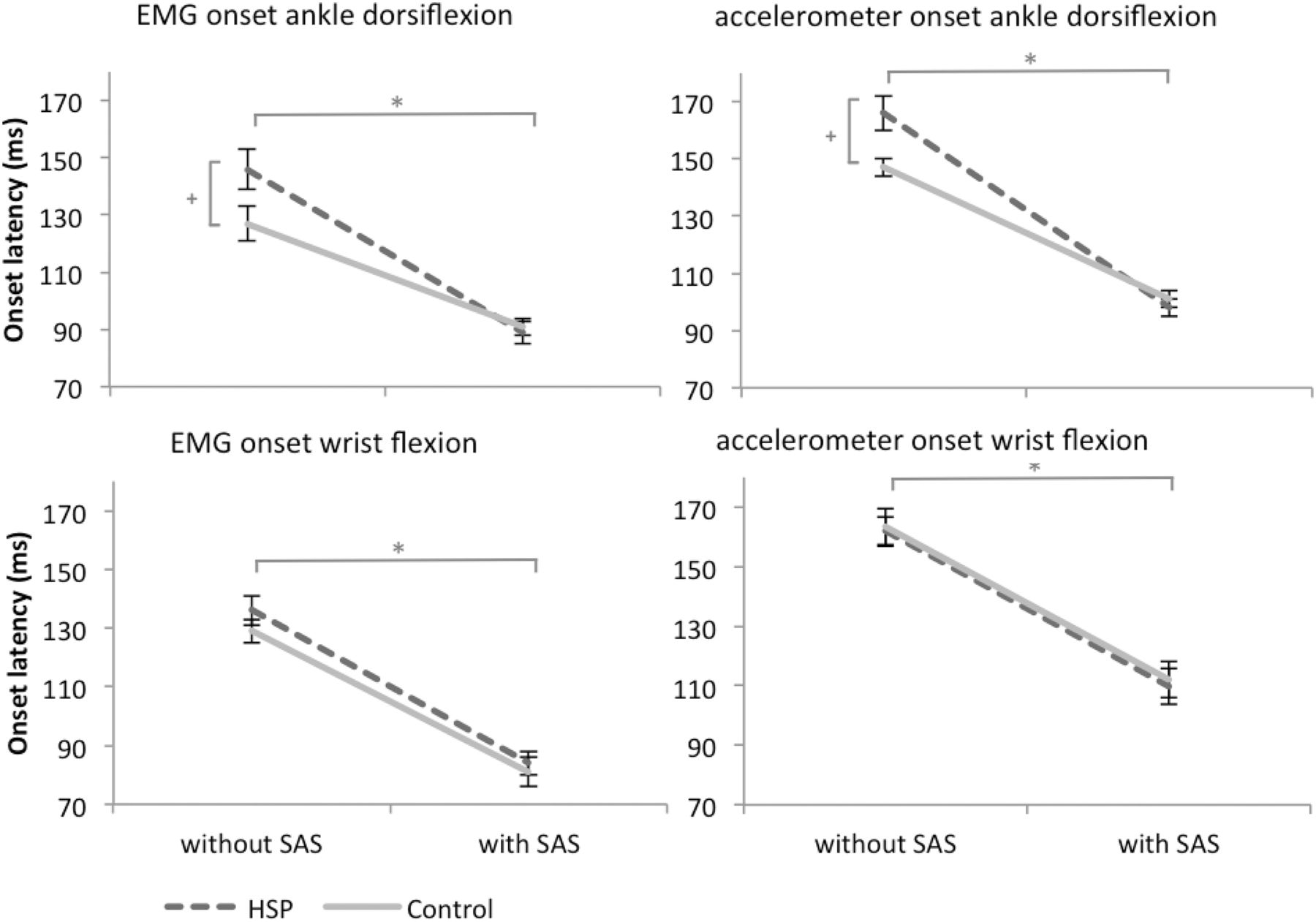
Mean EMG traces of a representative patient with HSP and a control subject during the simple reaction task involving voluntary dorsiflexion of the foot are shown in Figure 1. We found no consistent differences in the EMG activation pattern between patients with HSP and controls. In all participants, an asynchronous activation pattern of the TA and GM was observed both in trials with and without a SAS (Fig. 1). The onset latency in the TA was 146 ± 23 ms in patients with HSP compared with 127 ± 15 ms in controls in trials without a SAS (see Fig. 3). Administration of a SAS accelerated ankle dorsiflexion in both groups, but a larger acceleration in HSP patients normalized the latencies (89 ± 20 ms) compared with controls (91 ± 12 ms; SAS: F(1,22) = 217.16, p < 0001; SAS × group: F(1,22) = 12.028, p = 0.002; group: F(1,22) = 1.524, p = 0.230). Latencies during trials without a SAS were on average 19 ms longer in patients than in controls (95% CI: 4–35 ms, p = 0.017), but were similar during trials with a SAS (95% CI: −18–13 ms, p = 0.731). Administration of a SAS did not change the interval between the TA and GM (SAS: F(1,22) = 1.792, p = 0.194), neither in patients (21 ± 16 ms without SAS vs 17 ± 6 ms with SAS) nor in controls (20 ± 10 ms without SAS vs 17 ± 8 ms with SAS; group: F(1,22) = 0.030, p = 0.865; SAS × group: F(1,22) = 0.030, p = 0.865).

Mean EMG traces of a representative patient with HSP and control subject during ankle dorsiflexion. Gray lines are trials with a SAS, black lines are trials without a SAS. Determined latencies are presented by a dot.
The same pattern of results was seen in the onset latencies as measured with the accelerometer on the foot; in trials without a SAS these were 166 ± 21 ms in patients with HSP versus 147 ± 10 ms in controls. A larger SAS-induced acceleration in patient with HSP normalized the latencies (98 ± 20 ms) compared with controls (101 ± 9 ms; SAS: F(1,22) = 550.42, p < 0001; SAS × group: F(1,22) = 20.493, p < 0.001; group: F(1,22) = 1.459, p = 0.240). Again, latencies during trials without a SAS were on average 19 ms longer in patients with HSP than in controls (95% CI: 5–32 ms, p = 0.011), but were similar during trials with a SAS (95% CI: −17–10 ms, p = 0.591).
Wrist flexion reaction time
Mean EMG traces of a representative patient with HSP and a control subject during the simple reaction task involving wrist flexion are shown in Figure 2. The onset latency of the FCR was accelerated by a SAS, both in patients with HSP (136 ± 17 ms to 84 ± 14 ms) and in control subjects (129 ± 18 ms to 81 ± 20 ms; SAS: F(1,22) = 144.22, p < 0.001; Fig. 3).

Mean EMG traces of a representative patient with HSP and control subject during wrist flexion. Gray lines are trials with a SAS, black lines are trials without a SAS. Determined latencies are presented by a dot.

Mean onset latencies (SE) during the simple reaction time tasks involving voluntary ankle dorsiflexion (top graphs) and wrist flexion (bottom graphs). *Significant differences between trials with and without a SAS. +Significant SAS × group interaction.
There were no differences in the latencies of the FCR responses between the patients and the controls either with or without a SAS (group: F(1,22) = 0.652, p = 0.428; SAS × group: F(1,22) = 0.247, p = 0.624). The same pattern was seen when analyzing the onset latencies of the wrist accelerometer data; a SAS accelerated the latencies both in patients with HSP (162 ± 16 ms to 110 ± 14 ms) and in controls (163 ± 22 ms to 111 ± 22 ms; SAS: F(1,22) = 351.964, p < 0.001), with no differences between the groups (group: F(1,22) = 0.042, p = 0.840; SAS × group: F(1,22) < 0.001, p = 0.988).
Startle reflexes during reaction time tasks
During the ankle dorsiflexion task, 67% of the SAS-trials was accompanied by a startle reflex in the SCM muscle, both in patients with HSP and in controls. During the wrist flexion task, 60% of the SAS-trials were accompanied by a SCM-reflex in patients with HSP, and 67% of the SAS-trials in controls. When only SAS trials with a concurrent SCM-reflex were analyzed, the effect sizes and levels of significance were the same as when all SAS trials were used for analysis.
Discussion
This study aimed to investigate the mechanisms underlying the StartReact effect by comparing onset latencies of voluntary ankle dorsiflexion and wrist flexion with and without a SAS in patients with HSP and age-matched healthy controls. Patients with HSP did have significantly delayed corticospinal motor conduction times to the leg muscles compared with reference values of healthy control subjects. In contrast, in patients with HSP, startle reflexes in the TA muscle were not different from those in healthy controls with regard to both onset latencies and rates of occurrence. Simple reaction times of voluntary ankle dorsiflexion were delayed in the patients compared with the controls. However, when this task was combined with a SAS, reaction times in the patients were accelerated to a larger extent, resulting in completely normalized EMG and movement onset latencies. When the reaction time task involved voluntary wrist flexion instead of ankle dorsiflexion, we recorded no differences in onset latencies between patients and controls, regardless of whether a SAS was applied. This pattern of results is consistent with the hypothesis that a SAS accelerates reaction times through a release of a subcortically stored motor program, conveyed by the reticulospinal tract.
Reticulospinal integrity in HSP
To test the function of the reticulospinal tract in patients with HSP, we used SAS to elicit startle reflexes in the SCM muscle, a muscle that is known to respond well to SAS. Furthermore, we recorded startle reflexes in the TA and FCR. The reticulospinal tract did not seem to be affected in the patients, as onset latencies and reflex occurrence in all three muscles were not different from controls. To our knowledge, this is the first study to test the function of the reticulospinal tract in patients with HSP. The finding that not all patients and control subjects expressed startle reflex activity can be considered as a limitation of this method of assessing reticulospinal tract function. However, to our knowledge, there is no alternative for in vivo assessment of the functional integrity of the reticulospinal tract. Moreover, we found no indication that disease severity was anyhow related to the absence of TA startle reflexes in the patients.
Corticospinal degeneration in HSP
We were able to confirm the characteristic length-dependent, retrograde dysfunction of the corticospinal tract in HSP (Salinas et al., 2008) using reaction time tasks involving both voluntary ankle dorsiflexion and wrist flexion. In the selected patients, ankle dorsiflexion was delayed and wrist flexion not, which is coherent with the prolonged CMCTs to the lower limbs in patients with HSP. These prolonged CMCTs are in line with the literature as well as with the accepted notion of retrograde degeneration of the corticospinal tract (Pelosi et al., 1991; Polo et al., 1993; Bönsch et al., 2003; Nardone and Tezzon, 2003; Jørgensen et al., 2005; Sartucci et al., 2007). In the patients, the delay in ankle dorsiflexion reaction times (19 ms) was greater than the delay in CMCTs (6–7 ms). As patients were stimulated at 100% of the stimulator output during the TMS procedure, the CMCTs likely reflect the conduction time in the least affected corticospinal axons. Reasonably, the conduction time is longer in axons that are more affected. During the reaction time task involving ankle dorsiflexion, it is unlikely that the response involved excitation of as many neurons and at the exact same time compared with the TMS-evoked response. Hence, it is conceivable that, with degeneration of the corticospinal tract, reaction times to a visual stimulus exhibit longer delays than those measured with TMS, because the (first) corticospinal neurons to depolarize are not the least affected.
The origin of the StartReact effect
In the present experiment, we accelerated reaction times using a SAS, a phenomenon known as StartReact (Valls-Solé et al., 2008). Several observations strongly argue against the accelerated latencies simply being startle reflexes. First, during the ankle dorsiflexion task, the interval between TA en GM activation was not influenced by a SAS. This finding suggests that the SAS released the motor program without changing the characteristic agonist-antagonist activation pattern. If the SAS-accelerated latencies had been due to a startle reflex, TA and GM would have been activated synchronously, resulting in a shorter interval between both muscles. Our observation of a constant agonist-antagonist interval during StartReact is also in line with the literature. Valls-Solé et al. (1999) investigated the effect of a SAS on two stereotyped EMG patterns: the triphasic agonist-antagonist-agonist burst pattern of wrist flexion and the rising on tiptoes from standing position. When a SAS was accompanied with the imperative signal, the onsets of these movements were significantly accelerated, while leaving the movement-specific EMG pattern fully intact. This observation was reproduced by Carlsen et al. (2004a). A second argument against the SAS-accelerated latencies being startle reflexes is the observation that EMG activity in the TA was only observed during the ankle dorsiflexion task and not during the wrist flexion task, and wrist flexor activity was observed only during the wrist flexion task and not during the ankle dorsiflexion task. This finding suggests that the SAS released a specific motor program and that the accelerated latencies were not the expression of startle reflexes.
Previously, three mechanisms have been proposed to explain the occurrence of the StartReact effect. One hypothesis states that a SAS may act as an additional stimulus on top of the imperative stimulus, thereby increasing the energy of the sensory input, resulting in an acceleration of sensorimotor coupling. This hypothesis, known as intersensory facilitation (Nickerson, 1973), involves the corticospinal tract both in trials with and without a SAS. Yet, the degeneration of the corticospinal tract in patients with HSP did not lead to an impaired StartReact effect in the present study. In line with previous studies that have refuted this hypothesis (Carlsen et al., 2007; Valls-Solé et al., 2008), our results demonstrate that the StartReact effect cannot be explained by intersensory facilitation.
The currently dominant hypothesis states that the accelerated motor responses are due to the SAS directly releasing a subcortically stored pre-prepared motor program, which is then conveyed by the reticulospinal tract (Rothwell, 2006; Valls-Solé et al., 2008). Yet, another hypothesis that has recently been proposed is that a SAS acts as a subcortically mediated trigger for a cortically stored motor program (Alibiglou and MacKinnon, 2012; Carlsen et al., 2012), which mechanism would involve ascending reticular-cortical pathways and the corticospinal tract. This suggestion came from two studies using TMS. Both studies showed a significant delay in the StartReact effect when TMS was applied over the motor cortex (Stevenson et al., 2011; Alibiglou and MacKinnon, 2012). Although these results may support the involvement of cortical pathways in mediating the rapid release of a planned movement by a SAS, they do not rule out the possibility that the TMS-induced delay was due to reduced reticulospinal excitability through inhibitory effects of corticoreticular projections. A recent study did indeed provide evidence that the reticular formation can be mediated by TMS (Fisher et al., 2012). Thus, we believe that our finding that patients with HSP did not show an impaired StartReact effect in the TA muscle strongly argues in favor of a SAS releasing a subcortically stored motor program that is conveyed by an intact reticulospinal tract. This notion does not imply that the cortex has no influence on the subcortical release of motor programs by a SAS, as subcortical motor preparation most likely involves cortical processing (Marinovic et al., 2013).
Role of subcortical structures in motor preparation
Our results indicate that during motor preparation of voluntary ankle dorsiflexion and wrist flexion, motor programs become represented at subcortical levels which can be launched by a suitable reticular input. Indeed, studies in monkeys and cats provided evidence that motor preparation is not restricted to the cerebral hemispheres, and identified the pontomedullary reticular formation (pmRF) as one of the subcortical structures that subserves motor preparation as well (Buford and Davidson, 2004; Schepens and Drew, 2004). As the pmRF is also a key structure in the startle reflex circuitry (Davis et al., 1982), it may play a pivotal role in the release of pre-prepared motor programs resulting in the StartReact effect.
The extent to which the reticular system is involved in motor preparation probably varies depending on the type of movement. It has been hypothesized that the reticular system is involved in grasping, but not in all tasks that require individuated finger movements (Honeycutt et al., 2013). Accordingly, the StartReact effect was absent in the first dorsal interosseous (FDI) muscle during index finger abduction, whereas a startle did accelerate FDI latencies during grasping (Honeycutt et al., 2013). It remains speculative why the reticular formation is differently involved in the preparation of various movements. As the reticular formation is a key structure in postural control (Stapley and Drew, 2009), the representation of intended movements at a reticular level might enhance their integration with anticipatory postural adjustments for the upcoming actions. Again, this does not imply that the motor cortex is not involved in anticipatory processes (Spieser et al., 2013).
The results from the present and previous StartReact studies provide strong evidence for the existence of potent reticulospinal control over coordinated movements of the hand and foot (Riddle and Baker, 2010). As such, the reticulospinal system may be responsible for some of the functional recovery observed after acute corticospinal lesions (Baker, 2011). Recent experiments tested this idea by making focal unilateral pyramidal tract lesions in macaque monkeys (Zaaimi et al., 2012). After initial flaccid paralysis, grip function of the contralesional hand quickly recovered, which could not be attributed to corticospinal recovery. Interestingly, at 6 months postlesion, they demonstrated strengthening of reticulospinal connections to the forearm flexor but not to the extensor muscle groups. This pattern mirrors the predominant recovery of upper extremity flexor function as observed in patients with corticospinal lesions such as stroke. The gain in reticulospinal output to forearm flexors in stroke patients is also supported by a recent study that showed normal StartReact responses in stroke patients during elbow flexion, whereas excessive flexor activity was seen in SAS trials involving elbow extension (Honeycutt and Perreault, 2012).
We suggest that in the case of HSP, there may indeed be some degree of neuroplasticity through the reticulospinal system, thereby bypassing the dysfunctional corticospinal tract. Voluntary motor control through this bypass likely relies on intact corticoreticular pathways that originate from the premotor cortex, descend through the corona radiata, and terminate at the pmRF (Yeo et al., 2012). This may explain why patients with HSP (in whom these pathways are likely unaffected) generally retain voluntary, but less refined control over their leg movements, much more so than people with a severely affected corticospinal and corticoreticular tract after supratentorial stroke.
Conclusions and future perspectives
The results of this study in patients with pure hereditary spastic paraplegia support the hypothesis that the StartReact phenomenon can be attributed to the direct release of a subcortically stored pre-prepared motor program. Future studies may focus on the plasticity of the corticoreticulospinal pathways in humans and their role in motor control, as this may be a rather neglected substrate for functional recovery following lesions of the corticospinal tract.
Footnotes
This work was funded by a Radboud University Nijmegen Medical Centre Research grant to J.N., and a Netherlands Organization for Scientific Research Veni Research Grant 916.10.106 to V.W. B.R.B was supported by a research Grant from the Netherlands Organization for Scientific Research (016.076.352). We thank Roland Loeffen for technical assistance.
The authors declare no competing financial interests.
- Correspondence should be addressed to Jorik Nonnekes, Radboud University Medical Centre, PO Box 9101, 6500 HB Nijmegen, The Netherlands. jorik.nonnekes{at}radboudumc.nl





{kind=link}
{kind=link}
{kind=link}