
Abstract
The synaptic cleft, a crucial space involved in neurotransmission, is filled with extracellular matrix that serves as a scaffold for synaptic differentiation. However, little is known about the proteins present in the matrix and their functions in synaptogenesis, especially in the CNS. Here, we report that Hikaru genki (Hig), a secreted protein with an Ig motif and complement control protein domains, localizes specifically to the synaptic clefts of cholinergic synapses in the Drosophila CNS. The data indicate that this specific localization is achieved by capture of secreted Hig in synaptic clefts, even when it is ectopically expressed in glia. In the absence of Hig, the cytoskeletal scaffold protein DLG accumulated abnormally in cholinergic postsynapses, and the synaptic distribution of acetylcholine receptor (AchR) subunits Dα6 and Dα7 significantly decreased. hig mutant flies consistently exhibited resistance to the AchR agonist spinosad, which causes lethality by specifically activating the Dα6 subunit, suggesting that loss of Hig compromises the cholinergic synaptic activity mediated by Dα6. These results indicate that Hig is a specific component of the synaptic cleft matrix of cholinergic synapses and regulates their postsynaptic organization in the CNS.
- acetylcholine receptor
- DLG
- Drosophila
- matrix
- synaptic cleft
- synaptogenesis
Introduction
In both vertebrates and invertebrates, proteins secreted to the extracellular synaptic spaces regulate clustering of neurotransmitter receptors (Broadie et al., 2011; Nakayama and Hama, 2011). For example, Agrin in vertebrates (Wu et al., 2010) and LEV-9 and OIG-4 in Caenorhabditis elegans (Gendrel et al., 2009; Rapti et al., 2011) induce clustering of AchR at cholinergic neuromuscular junctions (NMJs). In the mouse cerebellum, Cbln1, which links Neurexin to the glutamate receptor GluD2 at synapses (Matsuda et al., 2010; Uemura et al., 2010), is capable of prompting GluD2 clustering in heterologous cells. In Drosophila glutamatergic NMJs, clustering of glutamate receptors depends on the secreted protein Mind-the-Gap (Rohrbough et al., 2007). Although these secreted proteins have been identified in a few specific types of synapses, the identities of the matrix proteins that regulate the organization of cholinergic synapses in the CNS remain unknown.
The hikaru genki (hig) gene was identified by analysis of Drosophila mutants exhibiting reduced locomotor behavior. This gene is expressed in a subset of neurons in the CNS of embryos and continues to be expressed in the nervous system until the adult stage. Hig is a secretory protein with an immunoglobulin (Ig) domain and a maximum of five complement control protein (CCP) domains (Hoshino et al., 1993). Immunoelectron microscopy reveals that Hig localizes to the synaptic clefts of subsets of synapses in the adult brain and in nascent synapses during pupal stages (Hoshino et al., 1996). This protein is present in the secretory apparatus (Hoshino et al., 1996) and is secreted from cultured cells to the medium (Hoshino et al., 1999). Mutant phenotypes of hig flies can be rescued by transient expression of Hig during middle pupal stages, suggesting that Hig plays a role in differentiation of synapses (Hoshino et al., 1996).
Here, we show that Hig is a matrix protein localized at the synaptic clefts of cholinergic synapses in the CNS and regulates the distribution of AchR subunits and DLG in postsynaptic terminals. This study indicates a possible avenue for revealing the composition of synaptic cleft matrix and determining how the matrix is involved in differentiation of cholinergic synapses in the CNS.
Materials and Methods
Drosophila strains.
higdd37 flies harbor a null mutation caused by a deficiency in the hig gene (Hoshino et al., 1993). The mutant strain nAcRα-30DDAS1 (hereafter Dα6DAS1) (Watson et al., 2010) was obtained from the Bloomington Stock Center. Dα7PΔEY6, Dα7-GAL4, and UAS-Dα7-EGFP were obtained from H.J. Bellen (Fayyazuddin et al., 2006). To generate UAS-hig-GFP, hig-GFP was amplified by PCR from pCasperHS-higD30GFP, which contains the longest form of hig cDNA fused with GFP. The resulting fragment was cloned into the EcoRI site of pUASTattB (Bischof et al., 2007). Transgenic fly lines expressing UAS-hig-GFP were generated by Phi-C31-mediated site-specific integration at attP2, P18, and P40.
Antibody production.
To generate anti-Hig antibody, a fragment of hig cDNA encoding the N-terminal fragment (amino acids 40–266) was amplified by PCR from GH23986 (BDGP) and cloned into pET15b (Novagen). Hexahistidine fusion proteins were expressed in Escherichia coli BL21 (DE3), purified, and injected into guinea pigs. To generate anti-Dα6 antibody, DNA encoding the cytoplasmic loop (amino acids 392–460) was amplified by PCR from GH15518 (BDGP) and cloned into pGEX4T-1 (GE Healthcare). GST fusion proteins were expressed in E. coli BL21 (DE3), purified, and injected into rabbits. Anti-Hig and anti-Dα6 antibodies were purified by affinity binding to the respective antigens.
Immunohistochemistry.
Brains of adult male flies were dissected in PBS, fixed with 4% paraformaldehyde (PFA) in PBS on ice for 1 h, and stained with the following antibodies: anti-Hig (1:1000), anti-Bruchpilot (nc82, 1:20, DSHB), anti-ChAT (ChAT4B1, 1:20, DSHB), anti-DVGLUT (1:500, Mahr and Aberle, 2006), anti-Rdl (1:1000, Enell et al., 2007), anti-GABARbII (1:1000, Enell et al., 2007), anti-DLG (4F3, 1:100, DSHB), anti-DN-cadherin (Ex#7, 1:20, DSHB), or anti-GFP (1:500, Invitrogen). AlexaFluor (-488, -568, -594, and -647)-conjugated antibodies (1:200, Invitrogen) were used as secondaries. For staining with anti-Dα7 (1:1000) and anti-Dα6 (1:1000), dissected brains were fixed with 4% PFA in PBS for 10 min on ice. Samples were observed by sequential scans on an SP2 confocal microscope (Leica) or ELYRA S.1 microscope (Zeiss). Quantitation of confocal images was performed using the Leica confocal software, with fluorescence intensity compared with that of control samples stained simultaneously in the same tube. At least seven samples were examined for the purposes of statistical analysis.
Insecticide assay.
Insecticide resistance assays were performed as previously reported (Watson et al., 2010) with slight modifications. Spinosad was dissolved at appropriate concentrations in 2:1 acetone:10% sucrose solution, and then 0.1 ml of each solution was applied to vials (inner diameter 2.1 cm) containing 5 ml of blue agar (1% agar, 1% Brilliant Blue FCF). Vials were dried overnight. Ten male flies (3- to 5-d-old) were introduced to each vial, and mortality was examined over 24 h. To assess the amount of food ingested by surviving flies, the bodies were separated from heads, homogenized in PBS, and centrifuged at 13,000 rpm for 25 min. After the supernatants were recentrifuged, absorbance was measured at 625 nm.
Results
Hig localizes predominantly at cholinergic synapses
Hig was detected most abundantly in synaptic regions that labeled positively for the presynaptic active-zone protein Bruchpilot (Brp; Wagh et al., 2006) in the adult brain (Fig. 1A). This Hig signal disappeared in the hig mutant higdd37, confirming the specificity of the anti-Hig antibody (Fig. 1A). To identify the neurotransmitter associated with the Hig-positive synapses, we performed double immunolabeling for several synaptic markers and Hig (Fig. 1B–E). The antennal lobes are well-analyzed regions containing synapses that use acetylcholine, glutamate, and GABA as neurotransmitters (Liu and Wilson, 2013). Synaptic Hig was closely associated with ChAT, a presynaptic marker of cholinergic neurons, whereas vesicular glutamate transporter (VGLUT) and GABA receptors (Rdl and GABAbRII) were largely excluded from Hig-positive zones (Fig. 1B–E). To confirm the distribution of Hig at cholinergic synapses, we used structured illumination microscopy to obtain high-resolution images of synapses, taking advantage of the surface localization of the antennal lobes in the brain. We expressed Dα7-GFP, a GFP-tagged AchR subunit Dα7, using a Dα7-GAL4 driver line; when the center of each signal was focused, the Dα7-GFP signals were closely associated with Hig (Fig. 1F). Structured illumination microscopy also revealed fine images of a single synapse in which Hig was present between the presynaptic marker Brp and the postsynaptic marker Dα7-GFP, indicating that Hig was localized to the synaptic cleft (Fig. 1G). The microglomerulus of the mushroom body (MB) calyx is a specialized synaptic complex that uses acetylcholine as a neurotransmitter (Fig. 1I,J) (Yasuyama et al., 2002) and is surrounded by noncholinergic synapses (Fig. 1K). This complex comprises an axon terminal of the projection neuron at its center and the dendrites of MB Kenyon cells (Fig. 1H) (Kremer et al., 2010). In most microglomeruli, a discrete pattern of Hig staining was observed in the surrounding regions of ChAT-positive projection neuron axon terminals (Fig. 1I). These immunohistochemical data from antennal lobe, MB, and other brain regions, including the optic lobes (see Fig. 3A,B), indicated that Hig predominantly localized to cholinergic synapses in the brain. To further analyze the discrete pattern of Hig in the microglomeruli, brains were simultaneously stained for Dα7 and Hig or Brp and Hig. Hig did not simply diffuse over the entire space of the synaptic clefts but was instead juxtaposed with Dα7 on the postsynaptic membrane (Fig. 1H,J); it also colocalized with Brp, an active-zone marker in the presynaptic terminal (Fig. 1H,K). We conclude that Hig is predominantly distributed in the cholinergic synapses and localizes at the synaptic cleft region adjacent to the active zone.

Hig localizes in the synaptic clefts of cholinergic synapses in the brain. A, Distribution of Hig in the brain. Frontal sections of the anterior and posterior brain regions are shown. Hig was observed in the entire synaptic region labeled with Brp in the WT brain, whereas Hig signals disappeared in higdd37. al, Antennal lobe; cx, calyx of the MB. B–E, Double labeling of synapses in the antennal lobe with antibodies against synaptic proteins and Hig. Hig was closely associated with ChAT (B) but not VGLUT (C), Rdl (D), or GABAbRII (E). Single glomeruli surrounded by dotted squares are magnified in the three right panels. F, G, High-resolution images of synapses in the antennal-lobe glomerulus obtained by structured illumination microscopy. Each Hig-positive synapse in focus is colabeled with Dα7-GFP, driven by Dα7-GAL4 (F). In most synapses, Hig (red at the top, gray at the bottom) localizes between Brp (blue) and Dα7-GFP (green) (G). Serial optical images were obtained at Z intervals of 0.3 μm. Scale bar, 100 nm. H, Schematic model for the distribution of Brp, Hig, and Dα7 in a microglomerulus formed by a single presynaptic bouton (pb) of olfactory projection neurons and the dendrites (d) of MB Kenyon cells (for the structure of the microglomerulus and distribution of synaptic markers, see Kremer et al., 2010). I–K, Distribution pattern of Hig in microglomeruli of the calyx. The single microglomerulus in a dotted square is enlarged in the three lower panels. Most Hig-positive microglomeruli were associated with ChAT (I). Discrete signals of Hig in microglomeruli were juxtaposed with Dα7 (J) and the active-zone marker Brp (K). K, Arrowheads indicate synapses without Hig, which are not cholinergic.
Hig is extracellularly diffused and trapped by cholinergic synapses
Because Hig is predominantly distributed in the cholinergic synapses, we predicted that the hig mutant phenotype could be rescued by expression of transgenic hig in the cholinergic neurons. To test this idea, we expressed GFP-tagged Hig (Hig-GFP) in the higdd37 background under the control of several GAL4 drivers. The higdd37 homozygotes exhibited reduced longevity, and this phenotype was rescued by expression of Hig-GFP in all neurons (elav-Gal4) and cholinergic neurons (Cha-Gal4), as expected (Fig. 2A). However, the longevity of higdd37 was also rescued by hig expression in glutamatergic neurons (OK371-Gal4 and VGlut-Gal4) as well as in glial cells (Eaat-Gal4 and Repo-Gal4) (Fig. 2A). By contrast, Gal4 lines driving expression in a small number of neurons (5HTR1B-Gal4, Yuan et al., 2005; Ddc-Gal4, Liu et al., 2012) were unable to rescue the mutant phenotype. Nevertheless, when expression of Hig-GFP was enhanced by addition of one more copy of the hig transgene, Ddc-Gal4 could prolong longevity (Fig. 2A). These results indicate that Hig does not need to be expressed in cholinergic neurons. Indeed, endogenous Hig was detected in the cell bodies of a large number of glutamatergic and cholinergic neurons (Fig. 2B).

Secreted Hig-GFP is captured by the cholinergic synapses. A, Rescue of the hig phenotype by ectopic expression of Hig-GFP. The indicated GAL4 lines were used to drive Hig-GFP expression in cholinergic, glutamatergic, or dopaminergic neurons, or glia, in the hig mutant background. The genotype of the control is higdd37; UAS-hig-GFP/ +. UAS-hig-GFP on the third chromosome was used for most of the experiments, unless otherwise stated. Ddc + 1st: UAS-hig-GFP/ Y; Ddc-Gal4, higdd37/ higdd37; +/+. Ddc + 3rd: +/ Y; Ddc-Gal4, higdd37/ higdd37; UAS-hig-GFP/+. Ddc + 1st 3rd: UAS-hig-GFP/ Y; Ddc-Gal4, higdd37/ higdd37; UAS-hig-GFP/+. Error bars indicate ± SEM. B, Expression of endogenous Hig in cholinergic and glutamatergic neurons. Cholinergic and glutamatergic neurons were labeled with nuclear GFP driven by Cha-Gal4 and VGlut-Gal4, respectively. Filled and open arrowheads indicate Hig-expressing neurons with and without GFP signals, respectively. C, Synaptic localization of Hig-GFP ectopically expressed in the glia. Eaat1-GAL4 was used for glial expression of membrane-bound mCD8-GFP and Hig-GFP. Hig-GFP is detected in most synaptic regions labeled with Brp in both the anterior (ant) and posterior (post) brain sections, whereas mCD8-GFP labels glial membranes (yellow arrowheads). In MB calyx of the posterior section, Hig-GFP was detected in cholinergic microglomeruli (arrows) but excluded from noncholinergic synapses outside microglomeruli (inset, arrowheads). Scale bar: C, inset, 5 μm. D, Microglomerulus-specific localization of endogenous Hig and pan-neuronally expressed Hig-GFP. As in C, inset, endogenous Hig (top) and Hig-GFP driven by elav-Gal4 (bottom) were only detected in cholinergic microglomeruli, but not in other synapses (arrowheads). mCD8-GFP driven by elav-Gal4 labeled the whole synaptic region of the calyx (middle).
The results of these rescue experiments and the synaptic distribution of Hig suggested that secreted and diffused Hig is eventually integrated into cholinergic synaptic clefts. To test this possibility, we examined the distribution of Hig-GFP expressed by a glia-specific driver (Fig. 2C). Secreted Hig-GFP was detected in most synaptic regions throughout the brain, notably in the cholinergic microglomeruli of the MB calyx (Fig. 2C), whereas Hig-GFP was not detected in noncholinergic synapses outside of the microglomeruli (Fig. 2C, inset). This microglomerulus-specific localization was also observed for endogenous Hig and Hig-GFP driven by elav-Gal4 in all neurons (Fig. 2D, insets). Thus, Hig is produced in cholinergic and noncholinergic neurons, secreted to extracellular spaces, and finally trapped in cholinergic synaptic clefts by a specific mechanism.
Hig regulates the synaptic distribution of Dα7, Dα6, and DLG
Because Hig is required for normal locomotor activity and longevity, we predicted that the neural function of cholinergic synapses would be partly impaired in the hig mutant. We therefore examined the distribution of several proteins localized to cholinergic synapses in the higdd37 brain. Although no obvious changes in the distribution of ChAT and acetylcholine esterase were observed in higdd37 brains (data not shown), levels of the AchR subunit Dα7, which was well colocalized with Hig in most brain regions, were significantly reduced throughout the brain, including MB calyx (Fig. 3A,G). In addition, levels of another AchR subunit Dα6 were reduced; Dα6 colocalized with Hig in many regions, including the layers of the medulla neuropil of the optic lobe and the MB calyx (Fig. 3B,C,G). Clustering of neurotransmitter receptors on the postsynaptic membranes is mediated by cytoskeletal scaffold proteins (Renner et al., 2008). Therefore, we examined the distribution of DLG, a Drosophila ortholog of PSD-95. At low magnification, the overall staining pattern of DLG in most brain regions did not differ significantly between wild-type (WT) and hig mutants (Fig. 3D). However, a higher-magnification view revealed elevated fluorescent intensity of DLG staining in a punctate pattern throughout the brain; each of the puncta was juxtaposed with the presynaptic terminals of cholinergic neurons (Fig. 3E). More conspicuously, DLG accumulated abnormally at microglomeruli in the MB calyx of higdd37, although the level of DN-cadherin was unchanged (Fig. 3F,G). Accumulation of Dα7, Dα6, and DLG in higdd37 brains was restored to WT levels by pan-neuronal expression of Hig-GFP (Fig. 3G). These results indicate that Hig is required for the proper accumulation of AchR subunits and the scaffold protein DLG in postsynaptic terminals of cholinergic synapses.

Dα7 and Dα6 levels decreases, but DLG level increases in the synaptic regions of adult higdd37 brain. A, Reduction of Dα7 localization at synapses of the optic lobe (top) and calyx microglomeruli (bottom) in higdd37. Hig was well colocalized with Dα7. B, Decrease of Dα6 localization at subsets of synapses in the optic lobe of higdd37. Magnified views of dotted areas of the medulla are shown in the middle panels for WT, and in the bottom center panel for higdd37. Strong Hig staining is found in specific layers of synapses that also contain Dα6 signals (arrowheads). Dα6 in these synapses (bracket) is significantly decreased in higdd37. Dα6 signals disappeared in Dα6DAS1 mutant brains, confirming the specificity of the anti-Dα6 antibody. C, Reduction in Dα6 signals at the synapses of the calyx microglomeruli in higdd37. D–F, Abnormal accumulation of DLG in higdd37. D, Frontal optical sections of WT and higdd37 brains stained for DLG, which labels synapses. The dotted square in the synaptic region is enlarged in E. E, Abnormal accumulation of DLG at cholinergic synapses in higdd37. Strong DLG signals (arrowheads) are adjacent to the presynaptic marker Syt-GFP, expressed under the control of cha-Gal4 (dotted squares, and insets for magnified views). Scale bar: E, inset, 1 μm. F, Abnormal accumulation of DLG in the calyx of higdd37. The intensity of DN-cadherin in higdd37 is comparable with that in the WT. G, Quantitative analyses of synaptic proteins Dα7, Dα6, DLG, and DN-cadherin in the calyx of WT, higdd37, and rescued flies (elav-Gal4/Y; higdd37; UAS-hig-GFP/+). Intensity of fluorescence in higdd37 is normalized to the WT values. Error bars indicate ± SEM. **p < 0.01, compared with WT (ANOVA followed by Bonferroni's post hoc test). NS, Not significantly different.
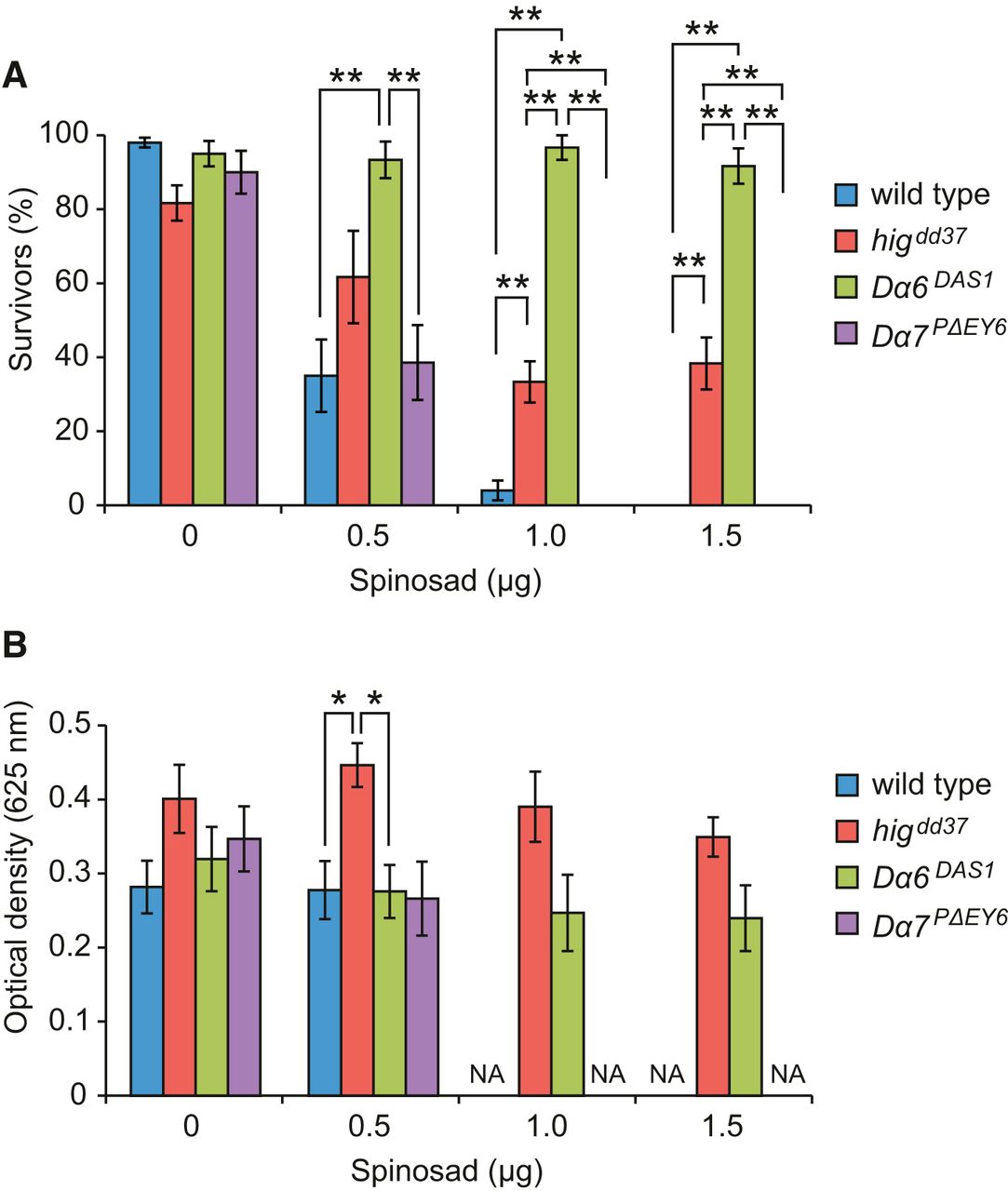
hig mutant flies are resistant to a Dα6 agonist
Because synaptic accumulation of Dα7 and Dα6 was reduced in the higdd37 mutant, we predicted that phenotypes similar to those of Dα7 and Dα6 mutants may occur in higdd37. Spinosad is a Dα6 agonist that specifically binds Dα6 and causes lethality by continuously activating AchR channels. Therefore, a null mutant of the Dα6 gene, Dα6DAS1, is highly resistant to spinosad (Fig. 4A) (Watson et al., 2010). When flies were reared on medium containing 1.0 or 1.5 μg of spinosad on the surface, higdd37 mutants survived at significantly higher frequencies than the control strains (WT and Dα7PΔEY6), but at lower frequencies than Dα6DAS1 flies (Fig. 4A). One potentially confounding factor was that, because higdd37 flies have reduced locomotor activity, they may eat smaller amounts of food than more active strains. To test this, we investigated the intake of food by measuring the amount of dye in extracts from surviving flies. Somewhat surprisingly, higdd37 flies tended to take even a larger amount of food than WT and Dα6DAS1, regardless of the content of spinosad (Fig. 4B); thus, the feeding behavior of higdd37 flies was not reduced. These data collectively indicate that higdd37 flies are partly resistant to spinosad, consistent with the notion that higdd37 impairs the cholinergic synaptic function mediated by Dα6. Thus, the matrix protein Hig organizes a molecular architecture involving Dα6, Dα7, and DLG in the postsynaptic terminals, and mutants defective in this organization exhibit reduced locomotion and longevity due impairments in the function of cholinergic synapses.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
higdd37 flies exhibit partial resistance to spinosad. A, Test for spinosad resistance. Flies (3- to 5-d-old) were reared on media with the indicated amount of spinosad for 24 h, and surviving flies were counted for each genotype. B, Feeding activity was estimated by measuring the optical absorbance of the dye in extracts from surviving flies. NA, Not applicable. Error bars indicate ± SEM. The resistance test was performed at least five times (50 flies) for each genotype. *p < 0.05 (ANOVA followed by Bonferroni's post hoc test). **p < 0.01 (ANOVA followed by Bonferroni's post hoc test).
Discussion
The data of Hig presented here provide the first evidence in any organism of a synaptic matrix protein specific to cholinergic synapses of the CNS. Hig secreted from cholinergic and noncholinergic neurons diffuses but is specifically trapped at the cholinergic synaptic clefts and then organizes a molecular architecture, including DLG and AchR subunits Dα6 and Dα7 in postsynaptic terminals.
In hig mutant brains, the intensity of antibody staining for Dα6 and Dα7 was notably reduced in the synaptic regions. Consistent with this result, hig mutant flies were resistant to a Dα6 agonist, spinosad. Therefore, Dα6-mediated neurotransmission is most likely to be impaired in hig flies, and deficits in Dα6 as well as Dα7 might explain why the mutant flies exhibited reduced locomotion and longevity. However, double-null mutants of Dα6 and Dα7 exhibit apparently normal locomotor behavior (data not shown), suggesting that hig mutant phenotypes are caused by additional defects in other factors, including AchR subunits, that functionally overlap with Dα6 and Dα7. Among 10 nAChR subunits encoded by the Drosophila genome, Dα5 can be combined with Dα6 and Dα7 to form functional heteromeric channels and also forms a functional homomeric channel (Lansdell et al., 2012). Therefore, Dα5 may compensate for loss of Dα6 and Dα7 in double mutants and is therefore a candidate for the AchR subunit responsible for reduced locomotion and longevity of hig flies.
Hig contains CCP domains, which are found in proteins that regulate complement activity in the vertebrate immune system. However, many CCP proteins exist in the nervous system of vertebrates and invertebrates, suggesting that they play evolutionarily conserved roles in neural function or development. Some of these proteins have been reported to localize at specific synapses and function in their differentiation (Nakayama and Hama, 2011). In C. elegans, the CCP proteins LEV-9 and AchR depend on each other for proper synaptic localization, and the two proteins are required for normal development of the NMJ (Gendrel et al., 2009). The human CCP protein SRPX2 is associated with rolandic epilepsy and oral dyspraxia (Roll et al., 2006), and its mouse counterpart regulates formation of glutamatergic synapses in the brain (Sia et al., 2013). When these CCP proteins modulate the development of synapses, a series of CCP domains in a polypeptide possibly enables it to interact with other proteins and participates in construction of the scaffolds of the synaptic clefts. CCP proteins have been shown to form a complex with proteins containing CCP or CUB domains (Tuveson et al., 1991; Gendrel et al., 2009). Therefore, some proteins with these domains in Drosophila may be Hig binding partners that are responsible for incorporation of Hig into specific synaptic clefts. Further studies of Hig should reveal the molecular composition of the synaptic cleft matrix, which gathers extracellular and membrane proteins on the terminal structures during cholinergic synaptogenesis. Because many CCP proteins are present in the human brain, the results of such investigations may be of clinical relevance.
Footnotes
- Received April 18, 2014.
- Revision received August 14, 2014.
- Accepted August 31, 2014.
This work was supported by Japan Society for the Promotion of Science Grant-in-Aid for Challenging Exploratory Research Grant 23650170. We thank Hugo J. Bellen, Ulrich Thomas, Hermann Aberle, Dick R. Nässel, Tadashi Uemura, and the Developmental Studies Hybridoma Bank for antibodies BDGP for cDNAs; Jing Wang, Hugo J. Bellen, and the Bloomington Stock Center for fly stocks; Shin-ichi Tsunoda for the use of structured illumination microscopy system; and Nobuaki Tanaka for technical instruction of GCaMP imaging.
The authors declare no competing financial interests.
- Correspondence should be addressed to Dr. Chihiro Hama, Faculty of Life Sciences, Kyoto Sangyo University, Kita-ku, Kyoto 603-8555, Japan. hama{at}cc.kyoto-su.ac.jp
- Copyright © 2014 the authors 0270-6474/14/3413872-06$15.00/0