
Abstract
Amygdalar NMDA receptor activation has been implicated in the acquisition of fear memories in rats. However, little is known about the role of endogenous modulators of the NMDA receptor, such as polyamines, in pavlovian fear-conditioning learning. Therefore, in the present study we investigated whether the immediate pretraining or post-training bilateral infusion of arcaine, an antagonist of the NMDA receptor polyamine-binding site, or spermidine, an agonist of the NMDA receptor polyamine-binding site, into the amygdala affected classical fear conditioning in rats. Bilateral microinjections of arcaine (0.0002-0.2 nmol) decreased, whereas spermidine (0.002-20 nmol) increased, contextual and auditory fear conditioning. Arcaine coadministration, at a dose that had no effect per se, reversed the facilitatory effect of spermidine. These results provide evidence that endogenous and exogenous polyamines modulate the acquisition or early consolidation (or both) of the fear-conditioning task in the amygdala.
- spermidine
- arcaine
- polyamines
- amygdala
- fear conditioning
- learning
- memory
Introduction
In pavlovian or classical fear conditioning, an animal learns to exhibit defensive responses to a neutral conditioned stimulus (CS) after its association with a noxious unconditioned stimulus (US). The amygdala has been implicated in the acquisition (Maren et al., 1996; Lee and Kim, 1998; Antoniadis and McDonald, 2000; Wilensky et al., 2000; Goosens and Maren, 2001), extinction (Falls et al., 1992; Lee and Kim, 1998), and in the expression of fear conditioning (Kim et al., 1993). Although the exact role of this structure in fear memory storage and formation has been a matter of intense debate in the literature (Cahill et al., 1999; Maren, 2000; Vazdarjanova, 2000), some agreement has been reached regarding the role of amygdalar ionotropic NMDA and non-NMDA glutamatergic receptors in the modulation of acquisition of fear memories and expression of fear. Whereas amygdalar NMDA and non-NMDA receptor activation seems necessary to the acquisition of fear memories (Fanselow and Kim, 1994; Rogan et al., 1997; Lee and Kim, 1998), the activation of non-NMDA receptors seems critical for the expression of fear (Fendt, 2001; Walker and Davis, 2002). Accordingly, pretraining intra-amygdala infusions of NMDA receptor antagonists disrupt fear conditioning (Miserendino et al., 1990; Fanselow et al., 1991, 1994; Campeau et al., 1992; Maren et al., 1996; Lee and Kim, 1998; Walker and Davis, 2002). However, little is known about the role of endogenous modulators of the NMDA receptor in the fear-conditioning learning (Walker et al., 2002).
Although the modulatory role of polyamines in the NMDA receptor function has been long known (Williams et al., 1991; Rock and Macdonald, 1995), the effects of polyamines on learning and memory have only recently been examined. High doses of polyamines (125-250 nmol, i.c.v.) cause hippocampal damage and learning impairment in rats (Conway, 1998) and potentiate dizocilpine-induced impairment of a learning task in rats (Shimada et al., 1994). These data support the view that high cerebral levels of polyamines are neurotoxic (Anderson et al., 1975; Halonen et al., 1993) and impair learning and memory.
Nevertheless, the systemic pretraining injection of polyamines improves social memory (Mikolajczak et al., 2002) and attenuates 3-[(±)-2-carboxypiperazine-4-yl)-propyl-1-phosphonic acid-induced learning deficits (Meyer et al., 1998) in a 14-unit T-maze. The post-training intrahippocampal or intra-amygdala administration of low doses of spermidine (0.02-20 nmol) improves inhibitory avoidance performance of rats (Rubin et al., 2000, 2001) and attenuates retrieval deficits induced by glutamatergic and cholinergic antagonists in maze tasks (Kishi et al., 1998a,b). As expected, polyamine antagonists impair passive avoidance behavior and prevent the facilitatory effects of spermidine on memory (Rubin et al., 2000, 2001). Therefore, it seems that the conflicting effects of polyamines, mainly spermidine, on learning and memory are attributable to the use of a wide range of doses of this compound. Although toxic amounts (>125 nmol, i.c.v.) of polyamines impair learning and memory, low doses (range, 0.02-40 nmol) facilitate memory consolidation and retrieval.
To date, studies investigating the effect of low doses of polyamines on learning and memory were restricted to the consolidation phase of the inhibitory avoidance task. Because studies on the effects of polyamines on the acquisition and consolidation of other tasks are lacking in the literature, we investigated whether the intra-amygdala injection of polyamines interferes with the acquisition and consolidation of the fear-conditioning task.
Materials and Methods
Animals. Male Wistar rats (230-250 gm) were housed five to a cage on a natural day/night cycle at a temperature of 21°C with ad libitum access to water and standard lab chow (Guabi, Santa Maria, Rio Grande do Sul, Brazil).
Surgery. Rats were implanted, under thionembutal-ketamine anesthesia (20 mg·kg-1 and 50 mg·kg-1, i.p., respectively), with a 27 gauge guide cannula aimed 1 mm above the amygdala at the anterior (-3.1), lateral (5.0), and ventral (6.4) (from the dura) coordinates from the atlas of Paxinos and Watson (1986).
Apparatus. Training and contextual fear testing took place in an identical observation chamber (30 × 25 × 25 cm), located in a well-lit room (room A) with ventilation fans producing a background noise of 70 dB. The front and ceiling walls of the chamber were made of clear acrylic plastic, whereas the lateral and rear walls were made of opaque plastic. The floor of the chamber consisted of 32 stainless steel rods (3 mm diameter), spaced 1 cm apart and wired to a shock generator. The chamber was cleaned with an acetic acid 1% solution before and after each rat occupied it.
Tone testing took place in a different room (room B) using a completely different chamber. This chamber was triangular (size, 23 cm each wall) and had its floor covered with linoleum. The lateral walls of the chamber were made of wood, whereas the front wall was made of translucent acrylic plastic to allow observation of the animals. There was no background noise. The cage was cleaned with 30% ethyl alcohol before and after each rat occupied it.
Fear conditioning. Four days after surgery, the animals were subjected to a fear-conditioning training session as described by Fanselow et al. (1994). In brief, the rat was placed in the conditioning chamber of room A. Animals were habituated to the apparatus for 3 min. During this period, the number of crossings across one imaginary line that divided the box floor into two equal segments was counted. Immediately thereafter, the rats received three presentations of a CS (90 dB sound at 2000 Hz) for 10 sec, followed immediately by an US (0.6 mA foot shock) for 1 sec. The shocks were 40 sec apart. The stimulus strength and number of training CS/US pairs were chosen on the basis of pilot experiments to allow increase or decrease of freezing behavior in the testing session. After the last CS/US pairing, rats were allowed to stay in the chamber for another 60 sec before returning to their home cages, and every 4 sec an instantaneous observation of the rat (1 sec) was made to assess whether it was in freezing. Freezing responses during this period were recorded as immediate freezing (Tang et al., 2001). Immediate freezing and spontaneous locomotion were recorded only in those experiments designed to evaluate whether polyamines modulate acquisition of contextual and auditory fear conditioning.
On the following day, each rat was placed back in the cage in room A, where it was trained the day before, and an 8 min test was performed. During this time, neither shock nor tone was given, and every 4 sec an instantaneous observation of the rat was made to assess whether it was in freezing, or not, by a trained observer who was unaware of the experimental treatment conditions. Behavior was judged as freezing if there was an absence of any visible movement, except for that necessitated by respiration. The data were converted to the percentage of samples scored as freezing and analyzed by one- or two-way ANOVA, depending on the experimental design. The percentage of samples scored as freezing in the absence of tone was taken as a contextual fear-conditioning measure.
On the second day of testing (tone test), the rats were placed into the chamber into room B. After a 3 min period of habituation, the tone (2000 Hz; 90 dB) was presented continuously for 8 min, and the percentage of samples scored as freezing was recorded, as described above. The percentage of samples scored as freezing in the presence of tone was taken as an auditory fear-conditioning measure.
In those experiments designed to evaluate whether polyamines modulate acquisition of contextual and auditory fear conditioning, the animals received injections bilaterally into the amygdala immediately before training with 0.5 μl/side over 5 min of vehicle (100 nmol of PBS, pH 7.4), 1,4-diguanidinobutane sulfate (arcaine; 0.002-.2 nmol; Sigma, St. Louis, MO), N-[3-aminopropyl]-1,4-butanediamine trihydrochloride (spermidine; 0.002-20 nmol; Sigma), or spermidine (0.02 nmol) plus arcaine (0.002 nmol). The injectors were left in place for 1 min after infusion. The injection was performed using 30 gauge cannulas that were fitted into the guide cannula, with the tip of the infusion cannula protruding 1.0 mm beyond that of the guide cannula and, therefore, aimed at the amygdala.
In those experiments designed to evaluate whether polyamines modulate early consolidation of contextual and auditory fear conditioning, the animals received injections bilaterally into the amygdala with 0.5 μl/side over 5 min of vehicle (100 nmol PBS, pH 7.4), arcaine (0.0002-0.2 nmol), spermidine (0.02-20 nmol), or spermidine (0.2 or 2 nmol) plus arcaine (0.0002 nmol), as described above, except that injections were performed immediately after training. The 0.5 μl injection volume was chosen based on previous studies (Barros et al., 2000; Wilensky et al., 2000; Fendt, 2001; Walker et al., 2002), and injection was aimed at the basolateral complex, which has been the amygdalar structure implicated in memory consolidation. However, because it has been reported that such a volume is likely to affect the basal nucleus (Wilensky et al., 2000), we refer to the affected area as the amygdala in this entire report.
Injection placements were verified immediately after the last testing session (Fig. 1), as described previously (Rubin et al., 2001). Only data from the animals with the correct placement of cannulas were analyzed.

Drawing adapted from Paxinos and Watson (1986) showing the area (black) where the infusions were considered correctly placed. Note that infusions were bilateral.
Freezing responses in a novel context (room B) without tone. To ensure that freezing during the tone was specific, as opposed to generalization across contexts, a new set of experiments was performed evaluating the freezing in the novel context without tone. Briefly, the animals were subjected to surgery, fear-conditioning testing, and pretraining or post-training administration of arcaine (0.002-0.02 nmol) or spermidine (0.02-2 nmol), as described above. On the second day of testing, the rats were placed into the chamber in room B, and the number of freezing responses was recorded for 8 min in the absence of tone. The percentage of samples scored as freezing in room B without tone was taken as freezing in a novel context without tone.
Foot shock sensitivity test. Reactivity to the shock was evaluated in the same apparatus used for fear-conditioning training. The modified “up and down” method (Roesler et al., 1999) was used to determine the flinch, jump, and vocalization thresholds in experimentally naive animals. Each animal received an injection with vehicle, spermidine (0.02 nmol), or arcaine (0.02 nmol) and was placed on the grid and allowed a 3 min period habituation period before the start of a series of shocks (1 sec) delivered at 10 sec intervals. Shock intensities ranged from 0.1 to 0.5 mA in 0.1 mA increments. The adjustments in shock intensities were made in accordance with each animal's response. The intensity was raised by one unit when no response occurred and lowered by one unit when a response was made. A flinch response was defined as withdrawal of one paw from the grid floor, and a jump response was defined as withdrawal of three or four paws. Two measurements of each threshold (flinch, jump, and vocalization) were made, and the mean of each score was calculated for each animal.
Plus-maze and open field tests. The elevated plus-maze apparatus consisted of a wooden structure elevated 50 cm from the floor and comprising two opposite open arms, 50 × 10 cm, crossed at right angles by two arms of the same dimensions enclosed by 40 cm high walls, with an open roof. Initially, subjects were placed on the center platform of the maze facing an enclosed arm (File et al., 1996). The behaviors recorded were the: total number of entries, the percentage of time spent on either arm, and the total number of dips. The apparatus was cleaned thoroughly between the 5 min observation sessions with a 30% ethanol solution.
Immediately after the plus-maze evaluation, the animals were transferred to the central area of a round open field (56 cm in diameter), which had its floor divided into 10 equal areas. Five areas of the apparatus had their borders limited by the walls of the arena and were considered as peripheral areas. The remaining five areas that had no contact with the walls of the apparatus were considered as central areas. The latency to leave the central area and the number of peripheral and central areas crossed were recorded for 5 min by an observer who was not aware of the subjects' conditions. Plus-maze and open field evaluation was performed on a separate subset of animals that was not used in other experiments.
Results
Figure 2, A and B, shows the effects of the pretraining intra-amygdala administration of arcaine (0.002-0.2 nmol) (a competitive antagonist at the NMDA polyamine-binding site) on freezing to context and to tone, respectively. Statistical analysis (one-way ANOVA) revealed that arcaine decreased freezing to context (F(3,36) = 7.76; p < 0.001) (Fig. 2A) and freezing to tone (F(3,36) = 4.23; p < 0.05) (Fig. 2B). Post hoc analysis (Student-Newman-Keuls test) showed that 0.02 nmol of arcaine impaired the acquisition of auditory and contextual fear conditioning. The post-training administration of arcaine also impaired freezing to context and to auditory CS (Fig. 3A,B). Statistical analysis (one-way ANOVA) revealed that arcaine decreased contextual freezing (F(4,42) = 2.89; p < 0.05) (Fig. 3A) and freezing to tone (one-way ANOVA: F(4.42) = 3.04; p < 0.05) (Fig. 3B). Post hoc analysis (Student-Newman-Keuls test) revealed that post-training administration of arcaine (0.02 nmol) impaired early consolidation of contextual fear conditioning, whereas administration of 0.002-0.02 nmol of arcaine impaired auditory fear conditioning.

Effect of pretraining intra-amygdala arcaine administration on freezing to context (A) and to tone (B). *p < 0.05 compared with vehicle by the Student-Newman-Keuls test. Data are the means + SEM percentage of freezing averaged across all context (A) or tone (B) trials (n = 9-11 animals in each group).

Effect of immediate post-training intra-amygdala arcaine administration on freezing to context (A) and to tone (B). *p < 0.05 compared with vehicle by the Student-Newman-Keuls test. Data are the means + SEM percentage of freezing averaged across all context (A) or tone (B) trials (n = 7-13 animals in each group).
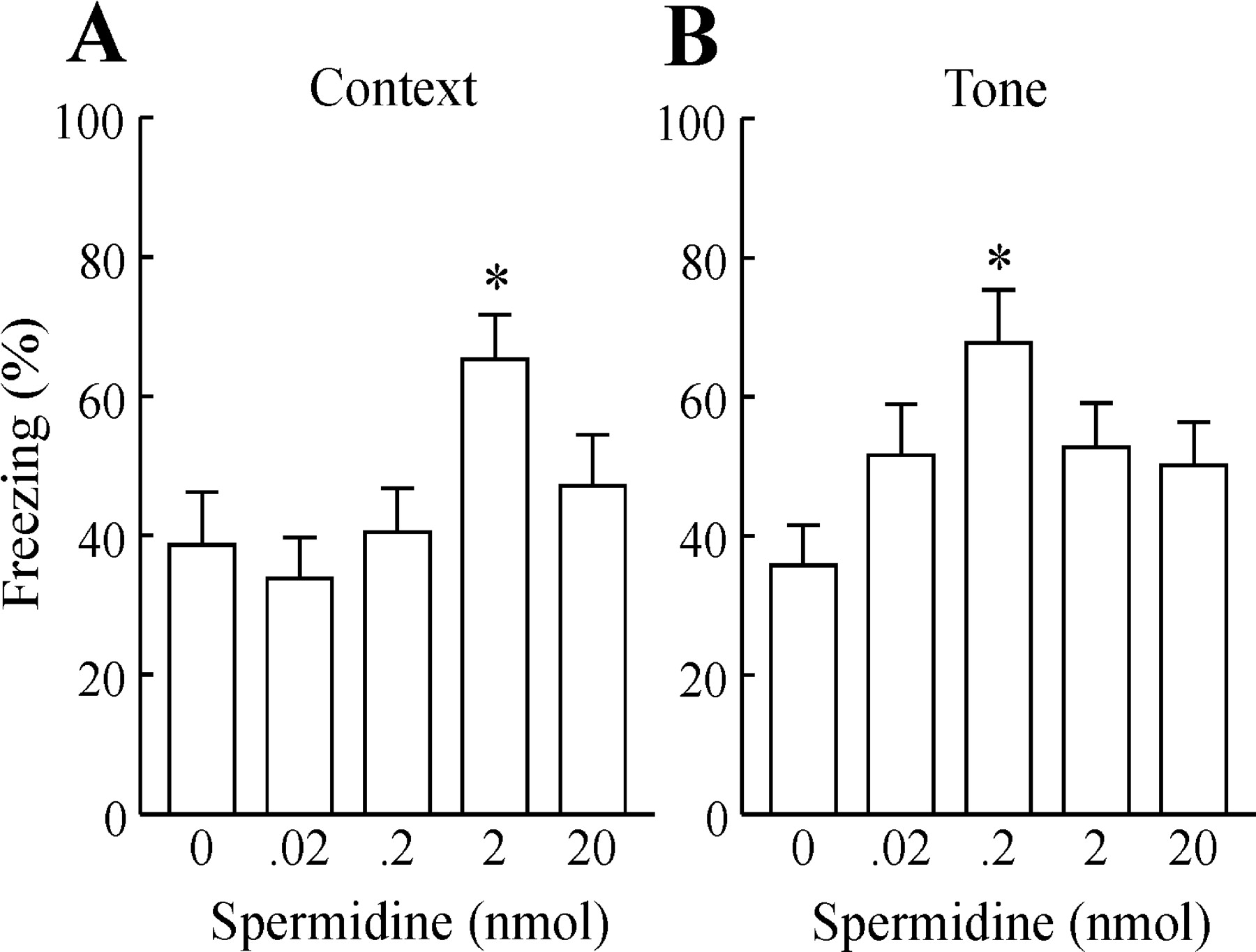
Figure 4, A and B, shows the effect of pretraining intra-amygdala administration of spermidine (0.002-20 nmol) on freezing to context and to tone (CS), respectively. Statistical analysis (one-way ANOVA) revealed a significant main effect of spermidine on contextual freezing (F(5,65) = 3.16; p < 0.05) (Fig. 4A) and on freezing to tone (F(5,65) = 4.92; p < 0.001) (Fig. 4B). Post hoc analysis (Student-Newman-Keuls test) revealed that 0.02 nmol of spermidine facilitated the acquisition of auditory and contextual fear conditioning. The post-training intra-amygdala administration of spermidine also facilitated freezing to context and to auditory CS (Fig. 5A,B). Statistical analysis (one-way ANOVA) revealed a significant main effect of spermidine on contextual freezing (F(4,60) = 3.24; p < 0.05) (Fig. 5A) and on freezing to tone (F(4,60) = 2.60; p < 0.05) (Fig. 5B). Post hoc analysis (Student-Newman-Keuls test) revealed that 2 nmol of spermidine facilitated contextual fear conditioning and 0.2 nmol of spermidine facilitated auditory fear conditioning.

Effect of pretraining intra-amygdala spermidine administration on freezing to context (A) and to tone (B). *p < 0.05 compared with vehicle by the Student-Newman-Keuls test. Data are the means + SEM percentage of freezing averaged across all context (A) or tone (B) trials (n = 9-14 animals in each group).

Effect of immediate post-training intra-amygdala spermidine administration on freezing to context (A) and to tone (B). *p < 0.05 compared with vehicle by the Student-Newman-Keuls test. Data are the meana + SEM percentage of freezing averaged across all context (A) or tone (B) trials (n = 12-14 animals in each group).
The pretraining coadministration of arcaine, at a dose that had no effect per se (0.002 nmol), and spermidine (0.02 nmol) reversed the facilitatory effect of spermidine on freezing to context (significant spermidine or PBS × arcaine or PBS interaction: F(1,65) = 12.65; p < 0.005) (Fig. 6A) and to tone (significant spermidine or PBS × arcaine or PBS interaction: F(1,65) = 5.63; p < 0.05) (Fig. 6B), suggesting that the facilitatory effect of spermidine may involve the polyamine-binding site in the NMDA receptor. The post-training coadministration of arcaine, at the dose that had no effect per se (0.0002 nmol), also reversed the facilitatory effect of spermidine (2 nmol) on freezing to context (significant spermidine or PBS × arcaine or PBS interaction: F(2,59) = 4.53; p < 0.05) (Fig. 7A) and reversed the facilitatory effect of spermidine (0.2 nmol) on freezing to tone (significant spermidine or PBS × arcaine or PBS interaction: F(2,59) = 3.80; p < 0.05) (Fig. 7B), suggesting that the facilitatory effect of post-training spermidine may be mediated by the NMDA receptor.

Effect of the pretraining intra-amygdala coadministration of arcaine (0.002 nmol) and spermidine (0.02 nmol) on the percentage of freezing to context (A) and to tone (B). PBS represents vehicle treatment. *p < 0.05 compared with vehicle by the Student-Newman-Keuls test. Data are the means + SEM averaged across all context (A) or tone (B) trials (n = 17-18 animals in each group).

Effect of the immediate post-training intra-amygdala coadministration of arcaine (0.0002 nmol) and spermidine (0.2 or 2 nmol) on the percentage of freezing to context (A) and to tone (B). PBS represents vehicle treatment. *p < 0.05 compared with vehicle by the Student-Newman-Keuls test. Data are means + SEM averaged across all context (A) or tone (B) trials (n = 9-13 animals in each group).
The pretraining or post-training administration of arcaine or spermidine did not alter freezing in the novel context (room B) without tone (all group means were <5%) (Table 1; see F values). This finding indicates that freezing during the tone (Figs. 2B, 3B, 4B, 5B) was in response to tone and was not attributable to generalization across context.
Effect of pretraining or post-training administration of arcaine or spermidine into the amygdala on the freezing responses in a novel context (room B) without tone
Because motivational disparities in the training session may account for differences in fear conditioning retention scores at testing, experiments were performed to assess whether arcaine or spermidine affected shock threshold, anxiety, or locomotor ability of the animals. Table 2 shows that pretraining intra-amygdala administration of arcaine, or spermidine, had no immediate effect on the locomotor ability of the animals, as assessed by the similar spontaneous locomotion scores presented by all groups during the habituation period (one-way ANOVA: F(3,36) = 0.28, p > 0.05, for arcaine; F(5,65) = 0.70, p > 0.05, for spermidine). Moreover, the pretraining intra-amygdala administration of arcaine, or spermidine, did not modify immediate freezing (one-way ANOVA: F(3,36) = 0.017, p > 0.05, for arcaine; F(5,56) = 2.11, p > 0.05, for spermidine) (Table 2), suggesting that the response to the shock were the same between groups, regardless of pharmacological treatment. Accordingly, arcaine (0.02 nmol) and spermidine (0.02 nmol) did not alter foot shock sensitivity, as demonstrated by the similar flinch (one-way ANOVA: F(2,23) = 0.02; p > 0.05), jump (one-way ANOVA: F(2,23) = 0.09; p > 0.05), and vocalization (one-way ANOVA: F(2,23) = 0.43; p > 0.05) thresholds exhibited by the animals (Fig. 8). Intra-amygdala arcaine (0.02 nmol) or spermidine (0.02 nmol) administration did not alter the anxiety state of the animals measured by the elevated plus-maze test (Table 3; see F values), indicating that any difference that could arise in the testing session could not be attributed to different levels of anxiety between groups. In addition, arcaine and spermidine did not alter anxiety measures and locomotion in the open field test (Table 4; see F values). This corroborated the plus-maze data and further indicated that the intra-amygdala administration of arcaine, or spermidine, did not cause motor disabilities.
Effect of pretraining administration of arcaine or spermidine into the amygdala on the spontaneous locomotion and immediate freezing responses during the training session in room A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of intra-amygdala administration of spermidine (0.02 nmol) or arcaine (0.02 nmol) on foot shock sensitivity. PBS represents vehicle treatment. Data are the means + SEM flinch, jump, and vocalization thresholds expressed in milliamps (n = 8-10 animals in each group).
Effect of arcaine or spermidine administration into the amygdala on the behavior of rats in the elevated plus-maze test
Effect of arcaine or spermidine administration into the amygdala on the behavior of rats in the open field test
Discussion
In the present study, we show that amygdalar endogenous polyamines are involved in the acquisition and early consolidation of fear conditioning, because pretraining (Fig. 2) and immediate post-training (Fig. 3) intra-amygdala administration of arcaine decreased freezing to tone and context in the testing sessions biphasically. This resulted in a U-shaped dose-response curve. To our knowledge, this is the first demonstration that the post-training intra-amygdala administration of negative NMDA receptor modulators disrupts auditory fear conditioning. It is interesting that the dose of arcaine that caused an impairment in the retention of fear conditioning (0.02 nmol; after training) is the same previously reported to impair inhibitory avoidance performance (Rubin et al., 2001). These results do suggest a role for amygdalar polyamines in the consolidation of both tasks. However, the potency of arcaine varied between tasks, because 0.2 nmol of arcaine impaired inhibitory avoidance performance (Rubin et al., 2001) but not fear conditioning. The reason for the potency difference between the two tasks is unknown. However, we may speculate that it may reflect some degree of task dependency, as previously debated by Wilensky et al. (2000), or may be the result of other factors, such as the use of different shock intensities between tasks. In fact, it is known that drug effects in memory tasks are sensitive to variations in experimental conditions, such as shock intensity (Gold and Buskirk, 1976; Walker and Davis, 2000). The shift of dose-response curves because of motivational differences (i.e., shock intensity, plate temperature) is a fairly well known phenomenon, particularly in experiments investigating the analgesic effects of selected drugs (Hunskaar et al., 1986; Yeomans et al., 1996; Yeomans and Proudfit, 1996). However, whether motivational differences can alter the dose-effect function of compounds injected intracranially has not been examined. In this regard, it is interesting to note that the amnestic effect of a single dose of AP-5 (intra-amygdala) depends on shock intensity (Walker and Davis, 2000). It is, therefore, also possible that the currently reported difference of effective arcaine doses between tasks may not represent a true task dependency but a shift in the dose-response curve because of shock intensity differences.
Interestingly, the pretraining and immediate post-training intra-amygdala spermidine administration increased freezing to tone and context in the testing sessions in a dose-dependent manner, after doses from 0.002-0.02 nmol/animal (before training) and 0.02-2 nmol/animal (after training). As the dose of spermidine increased beyond 0.2 nmol/animal (before training) and 2-20 nmol (after training), freezing scores returned to control values, yielding an inverted U-shaped dose-effect curve that mirrored that obtained with the polyamine receptor antagonist (Fig. 4 and 5). Such an inverted U-shaped dose-response curve for post-training administration of spermidine (including the dose range) is also quite similar to that previously reported by us using the inhibitory avoidance paradigm (Rubin et al., 2001) and further suggests that low doses of polyamines regulate amygdalar function, probably through the modulation of NMDA receptors. It is also remarkable that arcaine, at a dose that had no effect per se on conditioning to tone and context, reversed the improvement of fear conditioning induced by pretraining and post-training administration of spermidine (Figs. 6 and 7) and suggests a role for NMDA receptors in the currently reported effects of spermidine.
There is convincing neurochemical and electrophysiological evidence that polyamines dually modulate the NMDA receptor (Williams et al., 1991; Rock and Macdonald, 1995; Williams, 1997). Polyamines, at low micromolar concentrations, enhance [3H]MK-801 and [3H]TCP binding to the NMDA receptor channel, whereas higher concentrations of polyamines do not alter the binding of these ligands, resulting in a biphasic concentration dose-response curve (Ransom and Stec, 1988; Sacaan and Johnson, 1990; Williams, 1997). Accordingly, low concentrations of polyamines enhance NMDA-evoked currents, whereas higher concentrations of polyamines produce less enhancement of, or inhibit, NMDA receptor currents (McGurk et al., 1990; Sprosen and Woodruff, 1990; Williams et al., 1990; Rock and Macdonald, 1995). It is worth noting, however, that although polyamines cause biphasic electrophysiological and neurochemical effects on the NMDA receptor in vitro and on fear conditioning, it does not imply that these phenomena are causally related. This remark is particularly relevant in view of the finding that polyamines also regulate AMPA channels biphasically (Pellegrini-Giampietro, 2003), although the effects of spermidine on these channels are inverse, compared with NMDA receptors (i.e., low concentrations of polyamines inhibit, whereas high concentrations activate, ionic conductance). Therefore, one might expect that, given the role of amygdalar non-NMDA ionotropic receptors on fear conditioning acquisition (Rogan et al., 1997), polyamines should cause impairment of fear conditioning acquisition, which is inconsistent with the currently reported spermidine-induced increase of fear-conditioning performance.
A major concern in shock motivated learning tasks, particularly in those that investigate the effect of drugs on the acquisition of a given task, is whether pharmacological treatment affects motivational aspects of learning, such as shock sensitivity and anxiety. Such a concern is particularly relevant in studies involving the amygdala because of its involvement in fear expression and the fact that the fear response to acute stressful situations is generally accompanied by anxiety, sympathetic activation, and release of endogenous antinociceptive substances (Jensen and Smith, 1982; Snow et al., 1982; Fanselow, 1985; Lester and Fanselow, 1985; Netto et al., 1987). In the present study, we demonstrate that neither arcaine nor spermidine (at doses that affected fear conditioning) altered shock sensitivity, anxiety measures, or motor ability of the animals. Special care regarding this point was taken, because additional control behavioral measures within the tests (immediate freezing and spontaneous locomotor behavior) were assessed, as well as different specific behavioral tests for shock sensitivity, anxiety, and locomotion, to discard pharmacologically induced motivational differences between groups. The immediate freezing, although initially planned as a within-test measure of shock reactivity in our experiments, may also be interpreted as a measure of short-term contextual memory. Therefore, one might propose that the animals seemingly acquired, but failed to retain, conditioned fear or that, alternatively, immediate freezing is an unconditioned effect immune to NMDA receptor manipulations. These interpretations are rather interesting and reasonable, but because a high percentage of freezing (>70%) was observed in our experiments, it is also possible that ceiling effects obscured group differences, particularly facilitatory effects of pretraining administration of polyamines on immediate freezing. Therefore, at least for the moment, the meaning of this finding is uncertain, and its clarification awaits additional studies with specifically designed experimental protocols. Notwithstanding, considering the remarks above, all behavioral measures indicated that polyamine agonists and antagonists specifically affect acquisition or early consolidation (or both) of the fear-conditioning task and do not alter the motivational aspects of learning. Moreover, we also demonstrated that tone conditioning was specific and not a result of context generalization.
It is worth pointing out that the presently described arcaine-induced impairment of fear conditioning may have potential therapeutic applications, because the pharmacological disruption of consolidation processes has been considered in the treatment of the post-traumatic stress disorder (Walker et al., 2002).
In summary, in the present study we describe, for the first time, the modulatory effects of endogenous and exogenous polyamines (at low doses) on the acquisition and early consolidation (or both) of the fear-conditioning task. The involvement of NMDA receptors in the currently reported effects of polyamines is suggested, but additional studies are necessary to clarify this point.
Footnotes
- Received July 22, 2003.
- Revision received January 16, 2004.
- Accepted January 16, 2004.
This study was supported by Conselho Nacional de Desenvolvimento Cientifico e Tecnológico (CNPq) (500096/2003-1 and 500120/2003-0), Programa Institucional de Bolsas de Inciação Científica/CNPq-Universidade Federal de Santa Maria, Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, and Fundação de Amparo à Pesquisa do Estado do Rio Grande do Sul (01/1338.9). We thank Dr. Marino Bianchin for helpful comments about this manuscript and Gustavo Petri Guerra for technical assistance.
Correspondence should be addressed to Dr. Maribel A. Rubin, Departamento de Química, Centro de Ciĉncias Naturais e Exatas, Universidade Federal de Santa Maria, 97105-900, Santa Maria, Rio Grande do Sul, Brazil. E-mail: marubin{at}smail.ufsm.br.
Copyright © 2004 Society for Neuroscience 0270-6474/04/242328-07$15.00/0