
Abstract
The adenosine 5′-triphosphate (ATP), discovered in 1929 by Karl Lohman, Cyrus Hartwell Fiske, and Yellagaprada SubbaRow, acts as an important extracellular signaling molecule. In the CNS, ATP can be released from synaptic terminals, either on its own or together with other neurotransmitters. After the release from the presynaptic terminals, ATP binds to a plethora of ionotropic and metabotropic receptors, which mediate its action as an excitatory neurotransmitter. Furthermore, ATP also acts as an important mediator in neuronal–glial communications because glial cells are endowed with numerous ATP receptors, which trigger Ca2+ signaling events and membrane currents in both macro and microglia. In addition, ATP can be released from astroglial cells, thereby acting as a mediator of glial–glial and glial–neuronal signaling.
Similar content being viewed by others

The beginning
The molecule of adenosine 5′-triphosphate, ATP, was discovered simultaneously in Heidelberg and Harvard in 1929 by Karl Lohman and by Cyrus Hartwell Fiske and Yellagaprada SubbaRow. By stroke of luck or through the inspired leadership of Otto Meyrhof who was the director of Kaiser Wilhelm Institute for Physiology in Heidelberg, Lohman succeeded to publish his results in March 1929 [42], whereas Fiske and SubbaRow promulgated their report several months later, in October 1929 [23]. In this case, however, the fate was just and although the initial discovery is credited to Lohman, the name of ATP belongs to Fiske and SubbaRow, as Lohman called his molecule “adenylpyrophosphate.” Very soon the importance of ATP as a universal source of chemical energy in biological systems was fully appreciated [41].
Incidentally, the idea that purines may serve not only important intracellular functions but also act as extracellular signaling molecules was also born in 1929 when Drury and Szent-Györgyi [14] found that purines trigger negative chronotropic effect on the heart, mediated dilatation of coronary vessels, and inhibited intestinal smooth muscle. Very soon, it also became apparent that it was ATP, which was responsible for many purine-mediated physiological reactions [13].
Thirty years after the discovery of ATP, Pamela Holton [30] made a seminal observation that ATP can be released from nerves upon stimulation. Using the firefly luminescence method for ATP detection (introduced by Strehler and Totter [57, 58]), Pamela Holton found that electrical stimulation of great auricular nerve of rabbit resulted in transient elevation of extracellular ATP (Fig. 1). Based on these experiments, she concluded that “when noradrenalin is liberated from sympathetic nerve endings ATP may also be liberated in to the tissue spaces” [30], thus establishing a foundation for the theory of purinergic neurotransmission.

Discovery of ATP release from nerve terminals. Galvanometer readings before and at 10, 20, 30, and 60 s after adding 0.5 ml test solution to the firefly enzyme. C1 and C2 perfusates collected during control period, S1 and S2 during stimulation, and AS1 and AS2 immediately after stimulation. The last three deflexions were caused by adding 2, 4, and 10 pmol ATP to the control perfusate (C2). Figure is reproduced with permission from Holton [30]
In 1970, Geoffrey Burnstock et al. [9] presented the first direct evidence that ATP may act as a transmitter in “nonadrenergic, noncholinergic” nerves in the gut and in 1972, the same Geoffrey Burnstock [7] introduced the concept of “purinergic nerves” and “purinergic neurotransmission.” It was once more Geoffrey Burnstock who, with his characteristic determination, developed and elaborated the concept of purinergic transmission in the peripheral nervous system and demonstrated that ATP fully conforms to the criteria of the definition of “neurotransmitter”: (1) ATP is synthesized and stored in presynaptic terminals; (2) it is released upon nerve stimulation; (3) released ATP can be rapidly degraded by coenzymes; and (4) pharmacological agents, which inhibited effects of endogenous ATP, also suppresses the effects of nerve stimulation. Finally, Burnstock also challenged a Dale’s principle, which postulated that each nerve cell produces and releases only one transmitter, by demonstrating that ATP may be coreleased with other neurotransmitters, such as noradrenalin or acetylcholine.
ATP mediates synaptic transmission in the CNS
Probably the first hints that ATP may act as a neurotransmitter in neuronal–neuronal synapses were obtained by Thomas White [68] who found the release of ATP from brain synaptosomes treated by veratridine or high extracellular K+; later, he also demonstrated depolarization-induced ATP release from synaptosomes isolated from cortex and striatum and also found that in central nervous tissue, ATP was not coreleased with acetylcholine [70, 69].
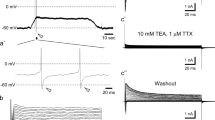
The excitatory action of ATP on neurones was found in 1983. First, robust inward currents induced by rapid application of ATP were recorded from internally dialyzed isolated sensory (nodose, vestibular, trigeminal, and dorsal root ganglia) neurones [39]. These currents were concentration-dependent (with Kd for ATP ∼5 μM) and were carried mostly by sodium ions as the substitution of the latter by Tris+, tetraethylammonium (TEA)+, or choline+ markedly suppressed the ATP-induced responses (Fig. 2).

First recordings of ATP-induced membrane currents in acutely isolated sensory neurones. a The “square pulse” application technique used for rapid application of ATP to internally dialyzed sensory neurones. The tip of the micropipette with the cell (a) is inserted into a plastic tube. The lower end of this tube can be exposed to different external solutions or to air. The suction applied to the upper end of the tube (indicated by arrow) is controlled by an electromagnetic valve (b). A preprogrammed sequence of current pulses applied to the valve allows a column of testing solution to form in the tube separated from the normal solution by air bubbles. Another sequence of pulses exposes the cell to the testing solution for the desired period of time by rapid displacement of the column along the tube. The electrical recording is unharmed because there is a thin layer of saline between the air bubbles and the walls of the tube. An invagination in the tip of the micropipette prevents the cell from damage. b ATP-activated inward current (ordinate) elicited by application of different concentrations of ATP as indicated on the graph. c The voltage-dependence of the peaks of ATP-activated current. ATP (5 μM) was applied in normal saline (circles) and in an external solution in which Na was substituted with TEA (triangles). Holding potential was −90 mV throughout. Figure is reproduced with permission from Krishtal et al. [39]
Several months later, the ATP-induced excitatory responses were also found in cultured dorsal horn neurones, impaled with microelectrodes [32]. Applications of ATP in concentrations between 1 and 10 μM triggered rapid depolarizations (Fig. 3) in about 30% of the neurones and these responses were abolished by lowering the extracellular Na+ concentration from 137 to 7.5 mM.

First recordings of ATP-induced excitatory responses in cultured dorsal horn neurones. a Chart records of intracellularly recorded responses of dorsal horn neurones to ATP. Disodium ATP (10 μM) was ejected at the moments indicated on the graph by pressure (1.5 psi) from a micropipette positioned ∼15 μm from the soma of the recorded neurone in pulses of 50, 100, and 200 ms in duration. The fast downward deflections were produced by injecting constant current hyperpolarizing pulses of 100 ms at 80 pA through the recording electrode and by providing a measure of input resistance. Resting potential=−62 mV. b Ionic-dependence of ATP-evoked depolarization of dorsal horn neurone. Voltage responses of the neurone to iontophoresis of Tris glutamate and Mg-ATP in control medium ([Na+]o=137 mM) and after switching to medium containing 7.5 mM Na+. Figure is reproduced with permission from Jahr and Jessell [32]
The year 1983 also witnessed another important discovery when Hans-Albert Kolb and Michael Wakelam identified ATP-activated single channel cation currents in membranes of myoblasts and myotubes [38]. They found several populations of channels with unitary conductances of 20, 48 (in myotubes), and 43 pS (in myoblasts), thus obtaining the first indication for heterogeneity of ATP receptors.
It took another decade before ATP-mediated synaptic transmission was experimentally identified. First, ATP-dependent synaptic currents were detected from cultured celiac ganglion cells [19, 55] and almost simultaneously, ATP-mediated excitatory postsynaptic currents and spontaneous “miniature” postsynaptic currents were detected from neurones in medial habenula from acutely isolated brain slices (Fig. 4a, [17, 18]). In all cases, the ATP-mediated synaptic currents were dissected using pharmacological approaches; in particular, the purinergic nature of the synaptic responses was confirmed based on their sensitivity to broad antagonist suramin [15] or by desensitization induced after incubation with α,β-methylene-ATP. Further investigations revealed ATP-mediated synaptic transmission in the spinal cord [3], hippocampus [44, 47], and cortex (Fig. 4b, [48, 49]).

ATP-mediated synaptic transmission in CNS neurones. a Voltage-dependence of ATP-mediated synaptic currents. ATP-mediated postsynaptic currents shown were recorded from a medial habenula neurone from a 24-day-old male rat in Krebs solution containing 5 mM Ca2+. Stimulus pulses (13 V, 200 μs) were delivered at 2 Hz and 53–215 currents recorded before the holding potential was stepped to the next voltage. The traces are averages of all responses (including failures). The peaks of the currents shown are plotted in the inset and fitted to a straight line using a linear regression. Peak current was measured from the single exponential fit of each current at the time indicated by the vertical line. The membrane potentials shown in this plot are −70, −50, −30, −5, +5, +20, +30, and +50 mV. Note that the voltage dependence of peak current amplitude is well fitted by a straight line with clear outward currents at positive holding potentials. Figure is reproduced with permission from Edwards et al. [18]. b Dissection of ATP-mediated component of excitatory postsynaptic currents (EPSCs) in the pyramidal neurone of layer II of somatosensory cortex. Top, changes in the amplitude of EPSC after bath application of glutamatergic antagonists NBQX and D-AP-5, cholinergic antagonist HEX, and P2X receptor antagonist NF023 as indicated on the graph. Each point represents mean±SD for six sequential trials, holding a potential of −80 mV, stimulation frequency 0.1 Hz. Bottom, the examples of EPSC (average of six traces) recorded at moments (1–5) indicated on upper graph. Figure is reproduced with permission from Pankratov et al. [48]
ATP receptors
All purinoreceptors are divided into two fundamentally distinct classes, the P1 receptors activated by adenosine and P2 receptors sensitive to ATP and its analogs (the subdivision proposed by Burnstock in 1978 [8]). The P2 receptors, in turn, are further split into two groups, the ionotropic P2X and metabotropic P2Y receptors [1, 46]. The P2X receptors are ligand-gated cationic channels, which, upon ATP-binding, undergo rapid conformational change that allows the passage of Na+, K+, and Ca2+ through the channel pore. There are seven P2X receptors subunits (P2X1–P2X7) encoded by distinct genes. These subunits may form homo- or heteromeric receptors with each functional receptor containing at least three monomers (see Fig. 5 and [35, 53, 60, 61, 65] and this issue for comprehensive reviews). The P2X7 receptor is unique in a sense that after its activation, it is followed by the appearance of a big pore, which allows the passage of molecules with molecular weight up to 900 Da. All P2X subunits are expressed in the brain and several subunits can be present within an individual neurone. Functional P2X receptors are also Ca2+-permeable and Ca2+ permeability relative to monovalent cations can range between 2 and 12 [16, 48, 53]. In this high Ca2+ permeability, the P2X receptors resemble the NMDA ones, albeit with one fundamental difference: P2X receptors can be activated at resting membrane potentials, whereas opening the NMDA ones requires membrane depolarization, which removes the Mg2+ block. As a consequence, the P2X receptors may provide an important Ca2+ influx pathway in resting neurones.

Trimeric structure of P2X2 receptors. a Imaging of P2X2 receptors. (a) A low magnification and (b) a high magnification image of P2X2 receptors bound to mica. b Atomic force microscope imaging of complexes between P2X2 receptors and anti-His6 antibodies showing images of receptors that are liganded by two antibodies. The angles between the two bound antibodies in are shown beneath the images. c Trimeric model of P2X2 receptor assembly. (a) A three-dimensional atomic force microscopy image of a receptor liganded by two anti-His6 antibodies. The angle between the bound antibodies is 121°. (b) The arrangement of antibodies bound to two His6 tags, assuming trimetric receptor architecture. Figure is modified from Barrera [4]
The P2Y receptors are classical 7-transmembrane-domain metabotropic receptors coupled to G proteins. These receptors are represented by at least ten subtypes out of P2Y1, P2Y6, P2Y11, P2Y12, P2Y13, and P2Y14 detected in the CNS [31]. Activation of neuronal P2Y receptors regulate both outwardly and inwardly rectifying K+ channels, potentiate high-voltage activated Ca2+ channels, and trigger InsP3-mediated release of Ca2+ from endoplasmic reticulum Ca2+ stores (see [31] for review).
ATP and neuronal–glial interactions
Purinergic transmission is particularly important for neuronal–glial integration in the CNS. It became exceedingly clear that the function of the nervous system is executed through continuous interaction of two cellular circuits, the neuronal and the glial [28, 45, 66]. Glial cells are an integral part of the synaptic contacts and they are endowed with numerous receptors allowing them to detect neuronal activity. In particular, glial cells are specifically sensitive to ATP as all types of glia, astrocytes, oligodendrocytes, Schwann cells, and microglia express ATP receptors (see, e.g., [11, 21, 27, 36, 37, 43, 54, 63]). In astrocytes and oligodendrocytes, the ATP-mediated signaling predominantly occurs through the P2Y receptors, which in turn trigger intracellular Ca2+ release and propagate Ca2+ waves, which serve as a substrate for glial excitability [24, 64]. The role for P2X1–6 receptors in astrocytes remains obscure, although P2X-mediated currents were detected in astroglial cells in culture [67] and P2X1–4 and P2X6 receptor mRNAs and proteins were detected in astrocytes in situ [22, 40]. P2X7 receptors are widespread in astrocytes and may be important upon pathological conditions [20]. Particularly important P2X and P2Y receptors are for microglia where they are abundantly expressed and may serve as sensors for neighboring cell damage, which may be associated with massive ATP release [29, 43, 51].
The second important role for ATP in neuronal–glial relations is associated with its capacity to act as a “glio” transmitter, as indeed, glial cells are capable to release several transmitters both in exocytotic and nonvesicular way ([5, 62, 66], see also Todd & Robitaille in this issue). ATP released by astrocytes can signal onto the neighboring astrocytes, thus assisting propagating Ca2+ waves in astroglial syncytium [11, 26]. Similarly, astrocyte-originated ATP can signal onto neurones. For example, in neuronal–glial cocultures prepared from hippocampus, ATP secreted by astrocytes was shown to inhibit glutamatergic synapses through activation of presynaptic P2Y receptors [71]. Alternatively, astroglial release of ATP may cause (through ATP degradation) an accumulation of adenosine, which in turn, may produce tonic suppression of synaptic transmission by acting on adenosine receptors [50].
ATP release
The fact that ATP molecule cannot cross the plasma membrane by diffusion was realized very early [10, 25]. This implies the necessity of specialized pathways for ATP release; furthermore, to provide for meaningful intercellular signaling, this pathway must be coupled with cellular excitation. Several release pathways can be operative in the CNS. First, ATP may be accumulated in synaptic vesicles together with other neurotransmitters and then both can be coreleased. This corelease of ATP is widespread in the peripheral nervous system [6]; in the CNS, ATP was reported to be coreleased with GABA [33, 34] and glutamate [44]. Alternatively, ATP may be released on its own from specific ATP-reach vesicles ([52], Pankratov et al., this issue), which may coexist with other neurotransmitter vesicles in the same presynaptic terminal or certain neurones can have pure ATP-releasing terminals. Which mechanism dominates in the CNS and what regional differences it may experience remains yet to be resolved. ATP may also be released by a nonvesicular mechanism, such as through gap junction hemichannels or via volume-sensitive chloride channels or indeed, via dilated P2X7 receptors [2, 12, 56, 59]. These latter mechanisms are likely to be associated predominantly with glial cells. Identifying the mechanisms and functional significance of ATP released from different elements of the CNS is of specific importance for further understanding of the role of purinergic transmission in the brain.
When coming up with this special issue, we tried to cover all fundamental aspects of purinergic transmission in the CNS, including the role of purinergic mechanisms in the regulation of CNS development (H. Zimmerman), molecular physiology, pharmacology of P2X (Roberts et al., Egan, Ford) and P2Y (Hussl & Boehm) receptors, intracellular signaling cascades involved in purinoreceptors action (Weisman & Erb), role of purinoreceptors in sensory transmission (Gu et al.), mechanisms of ATP release in central synapses (Pankratov et al.), signaling through dinucleoside polyphosphates (Delicado et al.), role of purinergic transmission in neuronal–glial interactions (Todd & Robitaille), and involvement of ATP in brain pathology (Färber & Kettenmann, Illes, Salter). We hope that this collection of essays, prepared by the leading experts in the field, will provide useful information on purinergic neurotransmission in the CNS.
References
Abbracchio MP, Burnstock G (1994) Purinoceptors: are there families of P2X and P2Y purinoceptors? Pharmacol Ther 64:445–475
Arcuino G, Lin JH, Takano T, Liu C, Jiang L, Gao Q, Kang J, Nedergaard M (2002) Intercellular calcium signaling mediated by point-source burst release of ATP. Proc Natl Acad Sci USA99:9840–9845
Bardoni R, Goldstein PA, Lee CJ, Gu JG, MacDermott AB (1997) ATP P2X receptors mediate fast synaptic transmission in the dorsal horn of the rat spinal cord. J Neurosci 17:5297–5304
Barrera NP, Ormond SJ, Henderson RM, Murrell-Lagnado RD, Edwardson JM (2005) Atomic force microscopy imaging demonstrates that P2X2 receptors are trimers but that P2X6 receptor subunits do not oligomerize. J Biol Chem 280:10759–10765
Bezzi P, Gundersen V, Galbete JL, Seifert G, Steinhauser C, Pilati E, Volterra A (2004) Astrocytes contain a vesicular compartment that is competent for regulated exocytosis of glutamate. Nat Neurosci 7:613–620
Bodin P, Burnstock G (2001) Purinergic signalling: ATP release. Neurochem Res 26:959–969
Burnstock G (1972) Purinergic nerves. Pharmacol Rev 24:509–581
Burnstock G (1978) A basis for distinguishing two types of purinergic receptor. In: Straub RW, Bolis L (eds) Cell membrane receptors for drugs and hormones: a multidisciplinary approach. Raven, New York, pp 107–118
Burnstock G, Campbell G, Satchell D, Smythe A (1970) Evidence that adenosine triphosphate or a related nucleotide is the transmitter substance released by non-adrenergic inhibitory nerves in the gut. Br J Pharmacol 40:668–688
Chaudry IH (1982) Does ATP cross the cell plasma membrane. Yale J Biol Med 55:1–10
Cotrina ML, Lin JH, Lopez-Garcia JC, Naus CC, Nedergaard M (2000) ATP-mediated glia signaling. J Neurosci 20:2835–2844
Darby M, Kuzmiski JB, Panenka W, Feighan D, MacVicar BA (2003) ATP released from astrocytes during swelling activates chloride channels. J Neurophysiol 89:1870–1877
Drury AN (1936) The physiological activity of nucleic acid and its derivatives. Physiol Rev 16:292–325
Drury AN, Szent-Györgyi A (1929) The physiological activity of adenine compounds with special reference to their action upon mammalian heart. J Physiol (London) 68:213–237
Dunn PM, Blakeley AG (1988) Suramin: a reversible P2-purinoceptor antagonist in the mouse vas deferens. Br J Pharmacol 93:243–245
Edwards FA (1994) ATP receptors. Curr Opin Neurobiol 4:347–352
Edwards FA, Gibb AJ, Colquhoun D (1992) ATP receptor-mediated synaptic currents in the central nervous system. Nature 359:144–147
Edwards FA, Robertson SJ, Gibb AJ (1997) Properties of ATP receptor-mediated synaptic transmission in the rat medial habenula. Neuropharmacology 36:1253–1268
Evans RJ, Derkach V, Surprenant A (1992) ATP mediates fast synaptic transmission in mammalian neurons. Nature 357:503–505
Fellin T, Pozzan T, Carmignoto G (2006) Purinergic receptors mediate two distinct glutamate release pathways in hippocampal astrocytes. J Biol Chem 281:4274–4284
Fields RD, Stevens B (2000) ATP: an extracellular signaling molecule between neurons and glia. Trends Neurosci 23:625–633
Franke H, Grosche J, Schadlich H, Krugel U, Allgaier C, Illes P (2001) P2X receptor expression on astrocytes in the nucleus accumbens of rats. Neuroscience 108:421–429
Fiske CH, SubbaRow Y (1929) Phosphorous compounds of muscle and liver. Science 70:381–382
Giaume C, Venance L (1998) Intercellular calcium signaling and gap junctional communication in astrocytes. Glia 24:50–64
Glynn IM (1968) Membrane adenosine triphosphatase and cation transport. Br Med Bull 24:165–169
Guthrie PB, Knappenberger J, Segal M, Bennett MV, Charles AC, Kater SB (1999) ATP released from astrocytes mediates glial calcium waves. J Neurosci 19:520–528
Haas S, Brockhaus J, Verkhratsky A, Kettenmann H (1996) ATP-induced membrane currents in ameboid microglia acutely isolated from mouse brain slices. Neuroscience 75:257–261
Haydon PG (2001) GLIA: listening and talking to the synapse. Nat Rev Neurosci 2:185–193
Hoffmann A, Kann O, Ohlemeyer C, Hanisch UK, Kettenmann H (2003) Elevation of basal intracellular calcium as a central element in the activation of brain macrophages (microglia): suppression of receptor-evoked calcium signaling and control of release function. J Neurosci 23:4410–4419
Holton P (1959) The liberation of adenosine triphosphate on antidromic stimulation of sensory nerves. J Physiol 145:494–504
Illes P, Ribeiro JA (2004) Neuronal P2 receptors of the central nervous system. Curr Top Med Chem 4:831–838
Jahr CE, Jessell TM (1983) ATP excites a subpopulation of rat dorsal horn neurones. Nature 304:730–733
Jo YH, Role LW (2002) Coordinate release of ATP and GABA at in vitro synapses of lateral hypothalamic neurons. J Neurosci 22:4794–4804
Jo YH, Schlichter R (1999) Synaptic corelease of ATP and GABA in cultured spinal neurons. Nat Neurosci 2:241–245
Khakh BS (2001) Molecular physiology of P2X receptors and ATP signalling at synapses. Nat Rev Neurosci 2:165–174
Kirischuk S, Moller T, Voitenko N, Kettenmann H, Verkhratsky A (1995) ATP-induced cytoplasmic calcium mobilization in Bergmann glial cells. J Neurosci 15:7861–7871
Kirischuk S, Scherer J, Kettenmann H, Verkhratsky A (1995) Activation of P2-purinoreceptors triggered Ca2+ release from InsP3-sensitive internal stores in mammalian oligodendrocytes. J Physiol (London) 483(Pt 1):41–57
Kolb HA, Wakelam MJ (1983) Transmitter-like action of ATP on patched membranes of cultured myoblasts and myotubes. Nature 303:621–623
Krishtal OA, Marchenko SM, Pidoplichko VI (1983) Receptor for ATP in the membrane of mammalian sensory neurones. Neurosci Lett 35:41–45
Kukley M, Barden JA, Steinhauser C, Jabs R (2001) Distribution of P2X receptors on astrocytes in juvenile rat hippocampus. Glia 36:11–21
Lippman F (1941) Metabolic generation and utilization of phosphate bond energy. Enzymology 1:99
Lohman K (1929) Uber die Pyrophosphatfraktion im Muskel. Naturwissenschaften 17:624–625
Moller T, Kann O, Verkhratsky A, Kettenmann H (2000) Activation of mouse microglial cells affects P2 receptor signaling. Brain Res 853:49–59
Mori M, Heuss C, Gahwiler BH, Gerber U (2001) Fast synaptic transmission mediated by P2X receptors in CA3 pyramidal cells of rat hippocampal slice cultures. J Physiol 535:115–123
Newman EA (2003) New roles for astrocytes: regulation of synaptic transmission. Trends Neurosci 26:536–542
North RA (2002) Molecular physiology of P2X receptors. Physiol Rev 82:1013–1067
Pankratov Y, Castro E, Miras-Portugal MT, Krishtal O (1998) A purinergic component of the excitatory postsynaptic current mediated by P2X receptors in the CA1 neurons of the rat hippocampus. Eur J Neurosci 10:3898–3902
Pankratov Y, Lalo U, Krishtal O, Verkhratsky A (2002) Ionotropic P2X purinoreceptors mediate synaptic transmission in rat pyramidal neurones of layer II/III of somato-sensory cortex. J Physiol 542:529–536
Pankratov Y, Lalo U, Krishtal O, Verkhratsky A (2003) P2X receptor-mediated excitatory synaptic currents in somatosensory cortex. Mol Cell Neurosci 24:842–849
Pascual O, Casper KB, Kubera C, Zhang J, Revilla-Sanchez R, Sul JY, Takano H, Moss SJ, McCarthy K, Haydon PG (2005) Astrocytic purinergic signaling coordinates synaptic networks. Science 310:113–116
Raivich G (2005) Like cops on the beat: the active role of resting microglia. Trends Neurosci 28:571–573
Robertson SJ, Edwards FA (1998) ATP and glutamate are released from separate neurones in the rat medial habenula nucleus: frequency dependence and adenosine-mediated inhibition of release. J Physiol 508:691–701
Robertson SJ, Ennion SJ, Evans RJ, Edwards FA (2001) Synaptic P2X receptors. Curr Opin Neurobiol 11:378–386
Robitaille R (1995) Purinergic receptors and their activation by endogenous purines at perisynaptic glial cells of the frog neuromuscular junction. J Neurosci 15:7121–7131
Silinsky EM, Gerzanich V, Vanner SM (1992) ATP mediates excitatory synaptic transmission in mammalian neurones. Br J Pharmacol 106:762–763
Stout CE, Costantin JL, Naus CC, Charles AC (2002) Intercellular calcium signaling in astrocytes via ATP release through connexin hemichannels. J Biol Chem 277:10482–10488
Strehler BL, Totter JR (1952) Firefly luminescence in the study of energy transfer mechanisms. I. Substrate and enzyme determination. Arch Biochem Biophys 40:28–41
Strehler BL, Totter JR (1954) Determination of ATP and related compounds: firefly luminescence and other methods. Methods Biochem Anal 1:341–356
Suadicani SO, Brosnan CF, Scemes E (2006) P2X7 receptors mediate ATP release and amplification of astrocytic intercellular Ca2+ signaling. J Neurosci 26:1378–1385
Surprenant A. (2004) P2X purinergic receptor. In: Encyclopedia of biological chemistry, vol 3. Elsevier, pp 183–187
Surprenant A, Buell G, North RA (1995) P2X receptors bring new structure to ligand-gated ion channels. Trends Neurosci 18:224–229
Takano T, Kang J, Jaiswal JK, Simon SM, Lin JH, Yu Y, Li Y, Yang J, Dienel G, Zielke HR, Nedergaard M (2005) Receptor-mediated glutamate release from volume sensitive channels in astrocytes. Proc Natl Acad Sci U S A 102:16466–16471
Verkhratsky A, Steinhauser C (2000) Ion channels in glial cells. Brain Res Brain Res Rev 32:380–412
Verkhratsky A, Orkand RK, Kettenmann H (1998) Glial calcium: homeostasis and signaling function. Physiol Rev 78:99–141
Vial C, Roberts JA, Evans RJ (2004) Molecular properties of ATP-gated P2X receptor ion channels. Trends Pharmacol Sci 25:487–493
Volterra A, Meldolesi J (2005) Astrocytes, from brain glue to communication elements: the revolution continues. Nat Rev Neurosci 6:626–640
Walz W, Gimpl G, Ohlemeyer C, Kettenmann H (1994) Extracellular ATP-induced currents in astrocytes: involvement of a cation channel. J Neurosci Res 38:12–18
White TD (1978) Release of ATP from a synaptosomal preparation by elevated extracellular K+ and by veratridine. J Neurochem 30:329–336
White TD (1984) Characteristics of neuronal release of ATP. Prog Neuropsychopharmacol Biol Psychiatry 8:487–493
White T, Potter P, Wonnacott S (1980) Depolarisation-induced release of ATP from cortical synaptosomes is not associated with acetylcholine release. J Neurochem 34:1109–1112
Zhang JM, Wang HK, Ye CQ, Ge W, Chen Y, Jiang ZL, Wu CP, Poo MM, Duan S (2003) ATP released by astrocytes mediates glutamatergic activity-dependent heterosynaptic suppression. Neuron 40:971–982
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
North, R.A., Verkhratsky, A. Purinergic transmission in the central nervous system. Pflugers Arch - Eur J Physiol 452, 479–485 (2006). https://doi.org/10.1007/s00424-006-0060-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00424-006-0060-y