
Abstract
The aim of this research was to assess the antinociceptive activity of the transient receptor potential (TRP) channel TRPV1, TRPM8, and TRPA1 antagonists in neurogenic, tonic, and neuropathic pain models in mice. For this purpose, TRP channel antagonists were administered into the dorsal surface of a hind paw 15 min before capsaicin, allyl isothiocyanate (AITC), or formalin. Their antiallodynic and antihyperalgesic efficacies after intraperitoneal administration were also assessed in a paclitaxel-induced neuropathic pain model. Motor coordination of paclitaxeltreated mice that received these TRP channel antagonists was investigated using the rotarod test. TRPV1 antagonists, capsazepine and SB-366791, attenuated capsaicin-induced nociceptive reaction in a concentration-dependent manner. At 8 μg/20 μl, this effect was 51% (P<0.001) for capsazepine and 37% (P<0.05) for SB-366791. A TRPA1 antagonist, A-967079, reduced pain reaction by 48% (P<0.05) in the AITC test and by 54% (P<0.001) in the early phase of the formalin test. The test compounds had no influence on the late phase of the formalin test. In paclitaxel-treated mice, they did not attenuate heat hyperalgesia but N-(3-aminopropyl)-2-{[(3-methylphenyl)methyl]oxy}-N-(2-thienylmethyl) benzamide hydrochloride salt (AMTB), a TRPM8 antagonist, reduced cold hyperalgesia and tactile allodynia by 31% (P<0.05) and 51% (P<0.01), respectively. HC-030031, a TRPA1 channel antagonist, attenuated tactile allodynia in the von Frey test (62%; P<0.001). In conclusion, distinct members of TRP channel family are involved in different pain models in mice. Antagonists of TRP channels attenuate nocifensive responses of neurogenic, tonic, and neuropathic pain, but their efficacies strongly depend on the pain model used.
摘要
目的
评价瞬时受体电位通道(TRP通道)TRPV1、 TRPA1和TRPM8拮抗剂在小鼠神经源性、 持续性和神经病理性疼痛模型中的作用。
方法
通过辣椒素实验、 异硫氰酸烯丙酯(AITC)实验和福尔马林实验, 评估TRP通道拮抗剂在小鼠神经源性疼痛模型中的镇痛作用; 通过建立紫杉醇诱导的小鼠神经病理性疼痛模型, 对TRP通道拮抗剂的抗痛觉(冷、 热、 触觉)过敏效应进行评估; 通过旋转法实验对小鼠的运动协调性进行评估。
结论
TRP通道家族包含了不同的小鼠疼痛模型。 TRP通道拮抗剂能减轻神经源性、 持续性和神经病理性疼痛, 但是其镇痛效果与疼痛模型有关。
Similar content being viewed by others

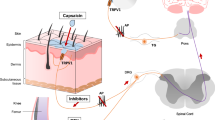

1 Introduction
Major interest in transient receptor potential (TRP) channels is because of the variety of biological functions in which they participate. Their localization on sensory neurons determines photo and thermoreception, as well as taste and smell perception. Since TRP channels play a crucial role in hypersensitivity to thermal, mechanical, and chemical stimuli during inflammation and after nerve injury, they are regarded as novel and promising drug targets for the treatment of inflammatory and neuropathic pain in humans (Brederson et al., 2013; Sałat et al., 2013b).
Recent neurobiological studies clearly indicate that some of the so-called ’thermosensitive’ TRP channels, i.e. vanilloid 1 (TRPV1), ankyrin-repeat 1 (TRPA1), and melastatin 8 (TRPM8), play a pivotal role in nociception. Noxious heat, capsaicin, or acidity can activate TRPV1 channels, whereas TRPA1 is stimulated by cold, oxidative stress byproducts, and chemical irritants, such as allyl isothiocyanate (AITC), cinnamaldehyde, acrolein, or formaldehyde (Eid et al., 2008; Sałat et al., 2013b). Cooling properties of menthol are explained by the activation of TRPM8 and TRPA1. In view of this, the inhibition of the aforementioned TRP channels by specific ligands holds promise for their use as not only a novel class of analgesics but also pharmacological tools for further investigation of the role of TRP channels in various pain models in mammals, particularly in chemically induced acute neurogenic, tonic, and neuropathic pain models.
It is well known that TRP channel agonists such as capsaicin and AITC evoke neurogenic pain by stimulating TRPV1 and TRPA1 channels, respectively. In contrast to this, nociception evoked by formalin and paclitaxel is not so clearly understood as it involves several distinct mechanisms. It was previously shown that formalin-induced pain is evoked by the activity of numerous mediators, channels, and receptors (Santos and Calixto, 1997; Coste et al., 2012), as well as nerve fiber types (Rios et al., 2013) and signaling pathways (Coste et al., 2012). Paclitaxel in turn is a chemotherapeutic agent, which causes toxic painful neuropathy and pain that are usually resistant to analgesic drugs (Nieto et al., 2012; Hara et al., 2013).
Accumulating data indicate that some TRP channels, i.e. TRPV1 (Hara et al., 2013), TRPV4 (Alessandri-Haber et al., 2004; Levine and Alessandri-Haber, 2007), TRPA1-and TRPV4-mediated glutathione-sensitive mechanisms (Materazzi et al., 2012) might be involved in the development and maintenance of paclitaxel-evoked neuropathic pain, although other molecules, such as sigma-1 receptors (Nieto et al., 2012), cannabinoid receptors (Authier et al., 2009), calcium channels (Authier et al., 2009) and CCL2 chemokines (Pevida et al., 2013), are also considered.
In the last years, numerous antagonists of thermo-TRP channels have been developed (Sałat et al., 2013b), but they have been mainly tested using in vitro and ex vivo assays, and there is rather conflicting and limited knowledge about their influence on the nociceptive threshold in behavioral models of acute and chronic pain in rodents (Walker et al., 2003). Moreover, in these previous studies only a systemic route of administration of TRP channel antagonists was used.
The present research aimed to establish the antinociceptive activity of TRP channel antagonists using various pain models in mice. These compounds injected into the dorsal surface of the hind paw of a mouse were tested in acute (neurogenic) pain models (capsaicin test and AITC test) and in the formalin (tonic) pain model. The intraperitoneal route of compounds’ administration was applied only in a model of painful toxic neuropathy induced by paclitaxel to assess the effect of various TRP channel antagonists on thermal hyperalgesia and tactile allodynia, and to investigate the potential role of TRP channels in pain hypersensitivity caused by paclitaxel.
To establish the effect of particular TRP channels on the development and maintenance of pain in these in vivo models, capsazepine, a nonselective TRPV1 antagonist (Walker et al., 2003), SB-366791, a selective TRPV1 antagonist (Niiyama et al., 2009), HC-030031 and A-967079 which are both TRPA1 antagonists (Chen et al., 2011), and AMTB, a TRPM8 antagonist (Lashinger et al., 2008) were assessed in behavioral assays.
2 Materials and methods
2.1 Animals
Adult male albino Swiss (CD-1) mice weighing 18–24 g were used in the present study. The animals were housed in standard laboratory cages (20 cm× 30 cm×15 cm), in groups of 10 mice/cage, at room temperature of (22±2) °C, under light/dark (12 h:12 h) cycle. The mice had free access to food and water before experiments. During the in vivo tests the temperature of the room and humidity were controlled. For the experiments, the animals were selected randomly. Each group consisted of 8–10 animals/dose, and each mouse was used only once. The behavioral measures were scored by trained observers blind to experimental conditions. The experiments were performed between 8:00 a.m. and 3:00 p.m. Immediately after the assay, the animals were euthanized by cervical dislocation. All experimental procedures were carried out according to the guidelines of the Local Ethics Committee of the Jagiellonian University in Cracow (ZI/595/2011), Poland.
2.2 Chemicals
Capsaicin, AITC, and formalin injected into the dorsal surface of the hind paw of a mouse cause a prompt response at the application site. This effect is called neurogenic inflammation. It appears within seconds and lasts for tens of minutes. Therefore, the antinociceptive activity of TRP channel antagonists in these three pain models was assessed after their local administration 15 min before the administration of algogens. The antinociceptive activity of TRP channel antagonists after their intraperitoneal injection was investigated in the paclitaxel-induced neuropathic pain model. Neuropathy caused by this anticancer drug develops within both the central and peripheral nervous systems. In this model of neuropathic pain, TRPV1, TRPA1, and TRPM8 ligands were evaluated for their potential antiallodynic and antihyperalgesic properties.
Paclitaxel, capsaicin, capsazepine, A-967079, AITC, and cremophor EL were provided by Sigma Aldrich (Poznań, Poland). HC-030031 and N-(3-aminopropyl)-2-{[(3-methylphenyl) methyl]oxy}-N-(2-thienylmethyl)benzamide hydrochloride salt (AMTB) were purchased from Tocris Bioscience (Warszawa, Poland). To establish a mouse model presenting neuropathy, paclitaxel was dissolved in ethanol (100% (v/v); Polskie Odczynniki Chemiczne, Gliwice, Poland) at 10% of the final desired volume and vortexed for 2 min. An equal volume of cremophor EL (10% of the final volume) was then added and the mixture was vortexed for the next 10 min. Prior to injection, ice-cold physiological saline (80% of the final volume) was added to make up a final volume and the solution was maintained on ice during dosing. This protocol was modified from Nieto et al. (2012).
AITC was diluted in corn oil (Sigma Aldrich, Poznań, Poland) to obtain 0.1% (w/w) solution, whereas capsaicin was dissolved in ethanol at 5% (w/w) of the final desired volume and vortexed for 2 min. Then physiological saline (Polfa, Kutno, Poland) was added (95% of the final volume) and the mixture was vortexed again. Formalin (37% (w/w) formaldehyde solution) was purchased from Polskie Odczynniki Chemiczne (Gliwice, Poland). It was diluted to obtain a 5% (w/w) solution, which was then used to induce a nociceptive reaction.
TRP channel antagonists tested in behavioral assays were dissolved in 0.3% (w/w) dimethyl sulfoxide (DMSO; Polskie Odczynniki Chemiczne, Gliwice, Poland) and vortexed for 5 min. They were either administered intraperitoneally (i.p.) 30 min before pain tests (refers to paclitaxel-induced neuropathic pain model) or injected into the dorsal surface of the hind paw of a mouse 15 min before algogens (formalin, AITC, or capsaicin). The doses of the test compounds used for in vivo experiments were chosen based on the results of our previous preliminary studies, as well as available literature data (Zhao et al., 2012). In each test, control animals received equivalent injections of the respective vehicle solutions.
2.3 Behavioral testing paradigm
2.3.1 Capsaicin test
After an adaptation period (15 min), the mice received 1.6 μg of capsaicin dissolved in 20 μl of physiological saline and ethanol (5:1, v/v). Capsaicin was injected into the dorsal surface of the right hind paw of a mouse. The test compounds were administered by the same route 15 min before capsaicin. In this assay, the animals were observed individually for 5 min following capsaicin injection. Pain-related behavior, i.e. the amount of time spent on licking, biting, flinching, or lifting the injected paw was measured using a chronometer (Sałat et al., 2009).
2.3.2 AITC test
After an adaptation period (15 min), 20 μl of 0.1% (w/w) AITC solution was injected into the dorsal surface of the right hind paw of each mouse. The test compounds were administered using the same route 15 min before AITC. In this test, the animals were observed individually for 20 min following AITC injection. Pain-related behavior, i.e. the amount of time spent on licking, biting, flinching, or lifting the injected paw, was measured using a chronometer (Zhao et al., 2012).
2.3.3 Formalin test
The injection of diluted formalin into the dorsal surface of the hind paw of a mouse produces a biphasic nocifensive behavioral response, i.e. licking, biting, flinching, or lifting the injected paw. The acute (neurogenic) nociceptive phase lasts for the first 5 min, and is followed by a period of little activity during the next 10 min. The second (late) phase occurs between 15 and 30 min after formalin injection (Laughlin et al., 2002). The formalin test was performed as previously described (Sałat et al., 2013a). Before the test, the mice were allowed to acclimate in Plexiglas cages (20 cm×25 cm×15 cm) for 30 min. Then, they were pretreated with the test compounds or vehicle injected into the dorsal surface of the hind paw. After 15 min, 20 μl of 5% (w/w) formalin solution was injected by the same route. Immediately after formalin injection, the animals were placed individually under glass beakers and were observed for the next 30 min. The total time (in seconds) spent on licking, biting, flinching, or lifting the injected paw was measured in periods of 0–5, 15–20, 20–25, and 25–30 min in each experimental group, and was an indicator of nociceptive behavior.
2.3.4 Paclitaxel-induced neuropathic pain
Paclitaxel injected intraperitoneally at a single dose of 6 mg/kg body weight was applied to induce neuropathy. The development of neuropathy was assessed using the cold water test, in which hind paws of each mouse were subjected to noxious cold stimulation (water bath maintained at 4 °C) and the latency time to paw withdrawal was measured. The reduction of cold sensitivity threshold, i.e. the reduction of latency time to pain reaction, was an indicator of neuropathy (Sałat et al., 2013a). The influence of the test compounds on tactile allodynia, cold and heat nociceptive thresholds was assessed using von Frey, cold water and hot plate tests, respectively.
2.3.4.1 Development of cold hyperalgesia
In the cold water test, the development of cold hyperalgesia in paclitaxel-treated mice was measured twice, i.e. 2 h (acute hyperalgesia) and then 7 d after paclitaxel administration (late hyperalgesia). A cut-off time of 30 s was established to avoid paw tissue damage. The mice not responding within 30 s were removed from the water and assigned a score of 30 s. For each animal, the reaction time was measured two to three times. An interval of at least 15 min between the two measurements was applied. To avoid paw cooling, the hind paws were dried with cellulose paper after each measurement. The reduction of latency time to pain reaction in paclitaxel-treated mice compared with mice not treated with this drug was an indicator of neuropathy. For further pain tests, only neuropathic mice were used.
2.3.4.2 Influence on cold nociceptive threshold
The influence of the test compounds on cold nociceptive threshold was investigated using the cold water test. After the establishment of baseline paw withdrawal latencies (pain behavior), each mouse was pretreated with the test compound or vehicle. Thirty minutes later the animals were again observed for the presence of pain behavior. To avoid paw tissue damage, a cut-off time of 30 s was established. The mice not responding within 30 s were removed from the water and assigned a score of 30 s. Final results were expressed as percentage maximal possible effect (%MPE) according to the following formula (Sałat et al., 2013a): %MPE=[(tpo-tpr)/(30-tpr)]×100%, where tpo and tpr are post-drug latency and pre-drug latency, respectively.
2.3.4.3 Evaluation of mechanical nociceptive threshold
Hypersensitivity to mechanical stimuli (tactile allodynia) was assessed using an electronic von Frey unit (Bioseb, Montpellier, France) supplied with a single flexible filament applying increasing force (from 0 to 10 g) against the plantar surface of the hind paw of the mouse. The nocifensive paw withdrawal response automatically turned off the stimulus and the mechanical pressure that evoked the response was recorded. On the day of the experiment, the mice were placed individually in test compartments with a wire mesh bottom and were allowed to habituate for 1 h. After the habituation period, in order to obtain baseline values, each mouse was tested three times alternately in each hind paw, allowing at least 30 s between each measurement. Then the mice were pretreated with the test compound or the vehicle. Thirty minutes later the animals were tested again and the mean value of the paw withdrawal threshold was obtained for each mouse (Sałat et al., 2013a).
2.3.4.4 Evaluation of heat nociceptive threshold
Thermal hyperalgesia was assessed in the hot plate test as previously described (Eddy and Leimbach, 1953) with minor modifications. First, baseline latencies to pain reaction were established for each mouse. Then, the mice were treated i.p. either with the test compound or vehicle. Thirty minutes later the animals were placed on the hot plate apparatus (Hot Plate 2A Type Omega, Cracow, Poland). This apparatus has an electrically heated surface and is supplied with a temperature-controller that maintains the temperature at 55–56 °C. The time until the animal licked its hind paws or jumped was recorded by means of a stop-watch. In this assay a cut-off time was established (45 s) to avoid tissue damage, and the mice not responding within 45 s were removed from the apparatus and assigned a score of 45 s.
2.3.5 Evaluation of motor impairing properties in paclitaxel-treated mice (rotarod test)
The rotarod test was performed according to the method recently described by Salat et al. (2012a). Briefly, before drug testing the mice were trained for three consecutive days on the rotarod apparatus (May Commat RR0711, Turkey; rod diameter: 2 cm) rotating at a constant speed of 18 r/min. During each training session, the animals were placed on a rotating rod for 3 min with an unlimited number of trials. Drug testing was conducted at least 24 h after the final training trial. On the test day, 30 min before the rotarod test the mice were pretreated i.p. with the test compound or vehicle. Then, the animals were tested on the rotarod apparatus revolving at 6, 18, or 24 r/min. Motor impairments, defined as the inability to remain on the rotating rod for 1 min, were measured at each speed and were expressed as the mean time spent on the apparatus (Salat et al., 2012a).
2.4 Data analysis
Data analysis of the results was provided by GraphPad Prism Software v.5 (San Diego, CA, USA). The numerical results from behavioral tests are expressed as mean±standard error of the mean (SEM). The results were statistically evaluated using Student’s t-test or one-way analysis of variance (ANOVA), followed by Dunnett’s or Tukey’s post hoc comparisons to compare the results obtained in drug-treated and control groups. Repeated measures of ANOVA followed by Bonferroni post hoc comparison were applied for the statistical evaluation of time-courses of the development of cold hyperalgesia. In every case P<0.05 was considered statistically significant.
3 Results
3.1 Antinociceptive activity of TRP channel antagonists in the capsaicin test
In the capsaicin test, the mean duration of the licking response in vehicle-treated mice was (49±5.7) s. Neither A-967079 nor AMTB at any concentration tested showed the antinociceptive activity. In contrast to this, the two antagonists of TRPV1 channels, capsazepine and SB-366791 (both tested at 4 µg/20 µl, 8 µg/20 µl, and 16 µg/20 µl), showed the antinociceptive activity, reducing the duration of the licking response as compared with the vehicle-treated mice (Fig. 1). The effects of both these compounds were concentration-dependent. A statistically significant antinociceptive effect was shown for 8 µg of SB-366791 (36.5%, P<0.01 vs. vehicle-treated mice), 16 µg of SB-366791 (49.7%, P<0.001 vs. vehicle-treated mice), and for all concentrations of capsazepine (31.9%, P<0.05 for 4 µg; 51.4%, P<0.01 for 8 µg; and 64.0%, P<0.001 for 16 µg). The comparison of the antinociceptive efficacies of the same doses of SB-366791 and capsazeine showed that these differences were not statistically significant.

Influence of TRP channel antagonists (each applied at 4 μg/20 μl, 8 μg/20 μl, and 16 μg/20 μl) on the duration of the nociceptive response in the capsaicin test
Test compounds were administered into the dorsal surface of the hind paw of a mouse 15 min before capsaicin. The results were statistically analyzed using one-way analysis of variance (ANOVA), followed by Dunnett’s post hoc comparison: SB-366791, F[3, 27]=7.489, P<0.001; capsazepine, F[3, 26]=11.08, P<0.0001; A-967079, F[3, 28]=0.6222, P>0.05; AMTB, F[3, 28]=0.3746, P>0.05. Results were compared to those of vehicle-treated mice: * P<0.05; ** P<0.01; *** P<0.001
3.2 Antinociceptive activity of TRP channel antagonists in the AITC test
In control animals, the mean duration of AITC-induced nocifensive response was (113±18.2) s. SB-366791, capsazepine, or AMTB did not attenuate the pain reaction induced by AITC. Only A-967079 showed antinociceptive activity in this test, reducing the duration of the nocifensive response by 30%–43.8% (vs. control mice). This effect was statistically significant only for 16 µg/20 µl (P<0.05; Fig. 2).

Influence of SB-366791, capsazepine, A-967079, and AMTB on the duration of the nocifensive response in the AITC test
Test compounds were administered into the dorsal surface of the hind paw of a mouse 15 min before AITC. The results were statistically analyzed using one-way ANOVA, followed by Dunnett’s post hoc comparison: SB-366791, F[3, 28]=0.1481, P>0.05; capsazepine, F[3, 28]=0.3393, P>0.05; A-967079, F[3, 28]=2.845, P>0.05; AMTB, F[3, 27]= 0.1008, P>0.05. Results were compared to those of vehicletreated mice: * P<0.05
3.3 Antinociceptive activity of TRP channel antagonists in the formalin test
In the early (neurogenic) phase of the test (0–5 min) only A-967079 showed antinociceptive activity (53.9% vs. control animals; statistically significant at P<0.001). The other TRP channel antagonists tested did not have antinociceptive properties either in the early or in the late phase of the formalin test (Fig. 3).

Time-course of the influence of SB-366791, capsazepine, A-967079, and AMTB (each applied at 16 μg/20 μl) on the duration of the nocifensive response assessed in the formalin test
Test compounds were injected into the dorsal surface of the right hind paw of a mouse 15 min before formalin. The results were statistically analyzed using repeated measures ANOVA, followed by Bonferroni post hoc comparison: drug effect, F[4, 105]=4.75, P<0.01; time effect, F[3, 105]= 59.06, P<0.0001; interaction, F[12, 105]=2.83, P<0.01. Results were compared to those of vehicle-treated mice at the respective time points: *** P<0.001
3.4 Antinociceptive activity of TRP channel antagonists in paclitaxel-induced neuropathic pain
3.4.1 Development of cold hyperalgesia
In the vehicle-treated mice, the mean latency time to pain reaction measured in the cold water test was (15.3±2.2) s. Two hours later in control mice not treated with paclitaxel, the mean latency time to pain reaction was (16.1±1.6) s (P>0.05 vs. latency time 2 h earlier). In contrast to this, a statistically significant reduction of pain sensitivity threshold was observed in the paclitaxel-treated group (Fig. 4). At this time point in paclitaxel-treated animals, the mean latency time to pain reaction was (5.1±0.7) s (P<0.001 vs. latency before paclitaxel administration). Since this effect was even more pronounced 7 d after paclitaxel injection ((2.8±0.4) s; P<0.001 vs. latency time before paclitaxel injection), the influence of the test compounds on thermal (cold and heat) and mechanical nociceptive threshold was assessed 7 d after paclitaxel injection.

Development of hyperalgesia in response to cold stimulus (water at 4 °C) in paclitaxel-treated mice compared with mice not treated with paclitaxel
The results were statistically analyzed using repeated measures ANOVA, followed by Bonferroni post hoc test: drug effect, F[1, 202]=61.26, P<0.0001; time effect, F[2, 202]=6.22, P<0.01; interaction, F[2, 202]=13.32, P<0.0001. Cremophor EL used as a solubiliser to prepare paclitaxel solution had no effect on animals’ pain sensitivity threshold, and there was only a marginal difference in pain reactivity between these animals and animals of the control group, so the data obtained for cremophor EL-treated mice were pooled
3.4.2 Influence of TRP channel antagonists on cold nociceptive threshold
In the cold water test in paclitaxel-treated mice only AMTB elevated the nociceptive threshold. For this compound %MPE value was 31% (significant at P<0.05). Neither capsazepine nor HC-030031 showed antihyperalgesic activity in this test. The %MPE values obtained for these compounds were 3.2% and 2.4%, respectively (Fig. 5a).

Effect of intraperitoneally administered test compounds (capsazepine (40 mg/kg), HC-030031 (25 mg/kg), and AMTB (25 mg/kg)) on pain sensitivity threshold in paclitaxel-treated neuropathic mice assessed 7 d after paclitaxel (PACLI) injection
(a) Antihyperalgesic activity in the cold water test. Results are shown as %MPE. Statistical analysis: paired Student’s t-test, * P<0.05 (vs. baseline values). (b) Effect of the test compounds on tactile allodynia measured using von Frey test. Statistical analysis: one-way ANOVA, followed by Tukey’s post hoc comparison; F[4, 73]=26.67, P<0.0001. Result significance vs. control mice not treated with paclitaxel: *** P<0.0001; result significance vs. baseline values obtained for paclitaxel-treated mice: ## P<0.01, ### P<0.001. (c) Antihyperalgesic activity in the hot plate test. Statistical analysis: one-way ANOVA, followed by Tukey’s post hoc comparison; F[4, 73]=35.04, P<0.0001. Results significance vs. control mice not treated with paclitaxel: *** P<0.001; result significance vs. baseline values obtained for paclitaxel-treated mice: ### P<0.001
3.4.3 Influence of TRP channel antagonists on tactile allodynia
In the von Frey test in control mice not treated with paclitaxel, the mean paw withdrawal threshold in response to mechanical stimulation was (3.0±0.2) g. In paclitaxel-treated mice, a statistically significant reduction of pain reactivity threshold was observed (47% vs. non-treated mice, P<0.001). Capsazepine had no effect on mechanical nociceptive threshold, whereas HC-030031 and AMTB elevated it by 62% (P<0.001) and 51% (P<0.01), respectively (Fig. 5b).
3.4.4 Influence of TRP channel antagonists on heat hyperalgesia
In the hot plate test in mice not treated with paclitaxel, the mean latency time to pain reaction was (12.8±0.8) s. The treatment with paclitaxel reduced the latency time to pain reaction to (8.9±0.5) s (P<0.001 vs. non-treated subjects). In the hot plate test, none of the test compounds attenuated heat hyperalgesia in neuropathic mice. In capsazepine-treated mice, as well as in mice treated with HC-030031 or AMTB, a significant reduction of latency time to pain reaction was observed (64%, 81%, and 63%, respectively; P<0.001 vs. baseline values obtained for paclitaxel-treated mice; Fig. 5c).
3.5 Influence of TRP channel antagonists on motor performance
In the rotarod test, the influence of intraperitoneally administered TRP channel antagonists (capsazepine (40 mg/kg body weight), HC-30031 (25 mg/kg body weight), and AMTB (25 mg/kg body weight)) on motor coordination of neuropathic mice was measured 7 d after paclitaxel administration. None of the test compounds induced motor impairments in paclitaxel-treated animals either at 6 r/min (F[3, 31]=0.6674, P>0.05), 18 r/min (F[3, 31]=0.8201, P>0.05), or 24 r/min (F[3, 31]=0.6256, P>0.05).
4 Discussion
The results of this in vivo study confirmed the involvement of the ‘thermosensitive’ TRP channels, TRPV1, TRPA1, and TRPM8 in nociception induced by selected chemical, thermal (heat and cold), and mechanical stimuli. Antagonists of these channels investigated in the present research proved their ability to attenuate nocifensive responses in behavioral models of acute (neurogenic) pain and chronic (neuropathic) pain in mice. This finding suggests that antagonists of TRP channels could be potential novel analgesics used in acute and neuropathic pain. It is, however, worth noting that their antinociceptive efficacy was strongly dependent on the kind of nociceptive stimulation applied.
To assess the role of TRP channels in nociception related to neurogenic inflammation (capsaicin test, AITC test, the first phase of the formalin test), the antagonists of TRPV1, TRPA1, and TRPM8 were administered into the dorsal surface of the hind paw to test their antinociceptive activity directly at the site of their application. An intraperitoneal route of administration was used in the paclitaxel-induced neuropathic pain to evaluate antiallodynic and antihyperalgesic properties of TRP channel antagonists in neuropathy caused by this antineoplastic agent.
In rodents neurogenic inflammation can be elicited by a variety of chemical stimulants, including AITC, capsaicin, or formalin. Although the behavioral symptoms of this phenomenon (edema of the injected paw and pain which induces licking or biting responses) are similar regardless of the kind of chemical used, it is assumed that these three substances have distinct mechanisms of action. Capsaicin is an agonist of TRPV1 channels, AITC selectively stimulates TRPA1 channels (Lopes et al., 2013; Sałat et al., 2013b), whereas formalin not only stimulates TRPA1 channels (McNamara et al., 2007) but it has also other pronociceptive mechanisms (Coste et al., 2012).
We have found that only the treatment with the TRPV1 antagonists, capsazepine and SB-366791, effectively attenuates capsaicin-induced nociception. This finding is in line with previous studies showing that mice lacking TRPV1 channels demonstrate a reduced nociceptive response to capsaicin, while they have normal reaction to other painful stimuli (Bandell et al., 2004). In rodents the administration of capsaicin into the dorsal surface of the hind paw determines hyperalgesia in response to thermal and mechanical stimuli (Callsen et al., 2008; O’Neill et al., 2012). This effect is due to TRPV1 stimulation, but capsaicin can also increase oxidative stress, reduce intracellular glutathione levels (Westlund et al., 2010), and inhibit TASK-1, TASK-3, and TRESK two-pore domain potassium channels (Beltrán et al., 2013). These different mechanisms of capsaicin’s action can at least in part explain the results of this present study, in which we demonstrated that a non-selective and competitive antagonist of TRPV1, capsazepine (Santos and Calixto, 1997; Lopes et al., 2013), attenuated pain symptoms of neurogenic inflammation elicited by capsaicin more effectively than SB-366791, a selective TRPV1 channel antagonist. Although these differences in efficacy of the two compounds were not statistically significant, they might suggest that capsaicin-induced alterations underlying the development of neurogenic inflammation not only depend on the stimulation of TRPV1, but also involve other signaling pathways. For capsazepine, additional mechanisms of action have been demonstrated. In vitro this compound inhibits potassium, calcium, and sodium channels (Docherty et al., 1997), and it acts as an antagonist of nicotinic receptors (Ray et al., 2003). It also decreases the expression of nitric oxide synthase in lipopolysaccharide-stimulated macrophages (Oh et al., 2001), and is able to abolish the inhibitory effect of capsaicin on endogenous opioid peptide binding to μ and κ receptors (Islam, 2011). In our research, the antinociceptive effect of capsazepine was shown after its local administration, which proved a peripheral antinociceptive activity observed at the injection site. Previously we also demonstrated that capsazepine exerted antinociceptive activity after systemic administration (Salat et al., 2012b). These ‘off-target’ effects of capsazepine may be in part the basis for the variations in potency seen for capsazepine and SB-366791. On the other hand, one must be cautious about over-interpreting the results on relative potency of these two drugs since only three concentrations were compared in this present study. Differences in bioavailability and/or differential clearance of the TRP channel antagonists should rather be excluded as determinants of the relative potency in the capsaicin test in view of the local application of the drugs used.
In mice the administration of AITC into the dorsal surface of the hind paw evokes neurogenic inflammation with paw edema formation, significant hyperalgesia and allodynia. AITC is an activator of TRPA1 channels (Ren and Dubner, 1999; Bautista et al., 2005; Merrill et al., 2008; Sandkühler, 2009). It causes a rapid nociceptive response in experimental animals. This can explain the fact that in the present study the attenuation of AITC-induced neurogenic pain was demonstrated only for a TRPA1 antagonist, A-967079, but not for TRPV1 or TRPM8 antagonists.
A formalin model of tonic pain assumes a biphasic nociceptive response of animals. The first phase of the test is directly associated with the stimulation of nociceptors and the development of neurogenic inflammation. The second phase is dependent on peripheral inflammation and central sensitization of pain (Laughlin et al., 2002). Based on the dependence of the first phase responses on the stimulation of TRP-expressing peripheral nerve endings, the effect of TRP channel antagonists was assessed in the formalin model. In the first phase of this test, only A-967079 reduced neurogenic pain, while other test compounds were not effective. None of the TRP channel antagonists was effective in the late phase. The effectiveness of A-967079 in the neurogenic phase of pain caused by formalin is in line with previous studies by McNamara et al. (2007) who showed that another TRPA1 antagonist, HC-030031, attenuated pain responses in both the early and the late phases of the formalin test. Although Santos and Calixto (1997) demonstrated the antinociceptive activity of intradermal capsazepine in the formalin test, this compound was not effective in our study. Their results do not coincide with those obtained in the present work, and this might be explained by methodological differences (lower concentrations of formalin, a different channel antagonist used at a high dose, and its route of administration—intraperitoneal). Indeed, behavioral responses in the formalin assay increase with increasing concentrations of formalin at 2%–5% (Sawynok and Liu, 2003). This might in part explain the lack of efficacy of SB-366791, capsazepine, and AMTB in our study, in which a relatively high concentration of formalin (5%) was used. Moreover, it must be emphasized that formalin-induced pain involves numerous mediators, channels, and receptors (Santos and Calixto, 1997; Coste et al., 2012), as well as nerve fiber types (Rios et al., 2013) and signaling pathways (Coste et al., 2012), so a selective inhibition of only one type of TRP channels might be insufficient to obtain effective antinociception. In addition, the local administration of TRP channel antagonists used in our study, and hence their strictly peripheral, local action might explain the lack of efficacy observed in the second phase of the test during which central sensitization of pain plays a pivotal role.
In rodents paclitaxel induces painful peripheral neuropathy with hypersensitivity to touch and cold. There are conflicting data regarding the development of heat hyperalgesia in this pain model (Bennett, 2010; Pascual et al., 2010; Xiao et al., 2012) and it is thought that the symptoms observed strongly depend on the strain, as well as different methodology used in each laboratory (Pascual et al., 2010). In order to exclude potential false positive results of pain tests, we evaluated the impact of TRP channel antagonists on motor coordination before the assessment of the their antinociceptive activity. No motor deficits were observed in the rotarod test. This indicated that the activity observed in pain tests was not due to neuromuscular blockade.
The onset of sensory symptoms after a single injection of paclitaxel appears within a few hours after its administration and is transient (until 24 h after paclitaxel administration). Full hypersensitivity is usually observed 7–14 d after a single injection of paclitaxel (Andoh et al., 2013; Rigo et al., 2013). We have shown that TRPM8 is involved in cold hyperalgesia in mice treated with paclitaxel. In the present study, acute hyperalgesia (2 h after paclitaxel injection) was not due to the influence of the solvent, as animals treated with the solvent that was used for paclitaxel preparation had reaction latency similar to that of vehicle-treated non-neuropathic mice.
Pain tests were conducted when hypersensitivity was sustained on Day 7 after paclitaxel injection. AMTB, a TRPM8 antagonist, efficaciously attenuated pain in response to low temperature. Neither HC-030031 nor capsazepine proved to be effective in the cold water test. The involvement of TRPA1 channels in the development of cold hyperalgesia in response to paclitaxel was shown previously either using genetic blockade (knockout TRPA1-/-animals), or TRPA1 antagonists (Chen et al., 2011; Materazzi et al., 2012; Zhao et al., 2012). In our work, this mechanism has not been confirmed, but our results are consistent with those presented by Bautista et al. (2006).
It is assumed that TRPA1 acts as an integrator of numerous endogenous and exogenous chemical and mechanical stimuli (Kerstein et al., 2009). HC-030031, a TRPA1 antagonist, attenuates mechanical hypersensitivity in some models of chronic inflammatory and neuropathic pain (complete Freund’s adjuvant and spinal nerve ligation models) (Eid et al., 2008). In our study we found a significant reduction of mechanical nociceptive threshold in paclitaxel-treated mice compared to non-treated littermates. Tactile allodynia was alleviated by the administration of HC-030031, as well as AMTB. This suggests that both TRPA1 and TRPM8 are responsible for the increased pain sensitivity in response to mechanical stimulation in paclitaxel-treated mice. In contrast to this, we did not demonstrate the antiallodynic activity of capsazepine in the von Frey test and this finding remains in agreement with reports of other authors (Chen et al., 2011). Tactile allodynia is mediated through Aβ-afferent fibers (Pascual et al., 2010). This can explain the observed lack of efficacy of capsazepine in the von Frey test, as this agent has no influence on Aβ-fibers. It is also worth noting that species differences of the antiallodynic activity of capsazepine in guinea pigs, mice, and rats were reported by Walker et al. (2003). They showed that capsazepine did not reverse chronic pain symptoms and was not able to attenuate mechanical hyperalgesia in mice and rats in contrast to its effectiveness in guinea pigs.
There are reports showing that paclitaxel can evoke heat hyperalgesia in mice (Xiao et al., 2012). In line with this, heat hyperalgesia in response to paclitaxel was observed in the present research, but the antinociceptive activity of TRP channel antagonists was not shown. None of the test compounds was able to restore the physiological threshold for thermal sensitivity in the hot plate test. In the case of capsazepine, its aforementioned species-dependent efficacy might at least in part explain the lack of antihyperalgesic properties in the hot plate test. The lack of effects of capsazepine may be also due to either inadequate dose or the temperature used. These issues might be a limitation of the present study. Interestingly, all TRP channel antagonists that were tested lowered thermal nociceptive threshold in this assay. This proalgesic effect is rather unexpected and it requires further research. The temperature applied in the hot plate test likely invokes not only TRPV1 channel but also additional ‘thermosensitive’ TRP and other ion channels, such as TRPV4 (VRL-2). The use of such a relatively high temperature could and probably has skewed the results with respect to the ability of TRPV1 antagonists to attenuate pain associated with heat stimulation. This issue is also discussed here as a potential limitation/confounding factor in the interpretation of the data obtained in the hot plate test, and indicates that the activity of TRPV2 antagonists should be investigated as a next step of our studies.
In summary, previous reports from both in vivo and in vitro studies indicated that the ‘thermosensitive’ TRP channels TRPV1, TRPA1, and TRPM8 might be emerging drug targets for novel analgesic drugs (Sałat et al., 2013b). The results of our research performed in mice have demonstrated that the antagonists of these channels might attenuate pain responses related to neurogenic inflammation elicited by capsaicin, AITC, and formalin. In addition, they are able to diminish cold as well as tactile allodynia but not heat hyperalgesia in neuropathic pain induced by paclitaxel. In neuropathic mice, the antinociceptive activity of these compounds is not accompanied by motor impairing properties, so they seem to be very promising lead structures in the search for novel analgesic drugs and pharmacological tools to study mechanisms of nociception.
Compliance with ethics guidelines
Kinga SAŁAT and Barbara FILIPEK declare that they have no conflict of interest.
All institutional and national guidelines for the care and use of laboratory animals were followed.
References
Alessandri-Haber, N., Dina, O.A., Yeh, J.J., et al., 2004. Transient receptor potential vanilloid 4 is essential in chemotherapy-induced neuropathic pain in the rat. J. Neurosci., 24(18):4444–4452. [doi:10.1523/JNEUROSCI.0242-04.2004]
Andoh, T., Sakamoto, A., Kuraishi, Y., 2013. Effects of xaliproden, a 5-HT1A agonist, on mechanical allodynia caused by chemotherapeutic agents in mice. Eur. J. Pharmacol., 721(1–3):231–236. [doi:10.1016/j.ejphar.2013.09.030]
Authier, N., Balayssac, D., Marchand, F., et al., 2009. Animal models of chemotherapy-evoked painful peripheral neuropathies. Neurotherapeutics, 6(4):620–629. [doi:10.1016/j.nurt.2009.07.003]
Bandell, M., Story, G.M., Hwang, S.W., et al., 2004. Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron, 41(6):849–857. [doi:10.1016/S0896-6273(04)00150-3]
Bautista, D.M., Movahed, P., Hinman, A., et al., 2005. Pungent products from garlic activate the sensory ion channel TRPA1. PNAS, 102(34):12248–12252. [doi:10.1073/pnas.0505356102]
Bautista, D.M., Jordt, S.E., Nikai, T., et al., 2006. TRPA1 mediates the inflammatory actions of environmental irritants and proalgesic agents. Cell, 124(6):1269–1282. [doi:10.1016/j.cell.2006.02.023]
Beltrán, L.R., Dawid, C., Beltrán, M., et al., 2013. The pungent substances piperine, capsaicin, 6-gingerol and polygodial inhibit the human two-pore domain potassium channels TASK-1, TASK-3 and TRESK. Front. Pharmacol., 4:141. [doi:10.3389/fphar.2013.00141]
Bennett, G.J., 2010. Pathophysiology and animal models of cancer-related painful peripheral neuropathy. Oncologist, 15(Suppl. 2):9–12. [doi:10.1634/theoncologist.2009-S503]
Brederson, J.D., Kym, P.R., Szallasi, A., 2013. Targeting TRP channels for pain relief. Eur. J. Pharmacol., 716(1–3): 61–76. [doi:10.1016/j.ejphar.2013.03.003]
Callsen, M.G., Moller, A.T., Sorensen, K., et al., 2008. Cold hyposensitivity after topical application of capsaicin in humans. Exp. Brain Res., 191(4):447–452. [doi:10.1007/s00221-008-1535-1]
Chen, Y., Yang, C., Wang, Z.J., 2011. Proteinase-activated receptor 2 sensitizes transient receptor potential vanilloid 1, transient receptor potential vanilloid 4, and transient receptor potential ankyrin 1 in paclitaxel-induced neuropathic pain. Neuroscience, 193:440–451. [doi:10.1016/j.neuroscience.2011.06.085]
Coste, O., Möser, C.V., Sisignano, M., et al., 2012. The p21-activated kinase PAK 5 is involved in formalin-induced nociception through regulation of MAP-kinase signaling and formalin-specific receptors. Behav. Brain Res., 234(1):121–128. [doi:10.1016/j.bbr.2012.06.017]
Docherty, R.J., Yeats, J.C., Piper, A.S., 1997. Capsazepine block of voltage-activated calcium channels in adult rat dorsal root ganglion neurones in culture. Br. J. Pharmacol., 121(7):1461–1467. [doi:10.1038/sj.bjp.0701272]
Eddy, N.B., Leimbach, D., 1953. Synthetic analgesics. II. Dithienylbutenyl- and dithienylbutylamines. J. Pharmacol. Exp. Ther., 107(3):385–393.
Eid, S.R., Crown, E.D., Moore, E.L., et al., 2008. HC-030031, a TRPA1 selective antagonist, attenuates inflammatory- and neuropathy-induced mechanical hypersensitivity. Mol. Pain, 4:48. [doi:10.1186/1744-8069-4-48]
Hara, T., Chiba, T., Abe, K., et al., 2013. Effect of paclitaxel on transient receptor potential vanilloid 1 in rat dorsal root ganglion. Pain, 154(6):882–889. [doi:10.1016/j.pain.2013.02.023]
Islam, M.S., 2011. Transient Potential Receptor Channels. Advances in Experimental Medicine and Biology. Springer, Berlin, Germany, p.704–720. [doi:10.1007/978-94-007-0265-3]
Kerstein, P.C., del Camino, D., Moran, M.M., et al., 2009. Pharmacological blockade of TRPA1 inhibits mechanical firing in nociceptors. Mol. Pain, 5:19. [doi:10.1186/1744-8069-5-19]
Lashinger, E.S., Steiginga, M.S., Hieble, J.P., et al., 2008. AMTB, a TRPM8 channel blocker: evidence in rats for activity in overactive bladder and painful bladder syndrome. Am. J. Physiol. Renal Physiol., 295(3):F803–F810. [doi:10.1152/ajprenal.90269.2008]
Laughlin, T.M., Tram, K.V., Wilcox, G.L., et al., 2002. Comparison of antiepileptic drugs tiagabine, lamotrigine, and gabapentin in mouse models of acute, prolonged, and chronic nociception. J. Pharmacol. Exp. Ther., 302(3): 1168–1175. [doi:10.1124/jpet.302.3.1168]
Levine, J.D., Alessandri-Haber, N., 2007. TRP channels: targets for the relief of pain. Biochim. Biophys. Acta, 1772(8):989–1003. [doi:10.1016/j.bbadis.2007.01.008]
Lopes, S.C., da Silva, A.V., Arruda, B.R., et al., 2013. Peripheral antinociceptive action of mangiferin in mouse models of experimental pain: role of endogenous opioids, KATP-channels and adenosine. Pharmacol. Biochem. Behav., 110:19–26. [doi:10.1016/j.pbb.2013.05.016]
Materazzi, S., Fusi, C., Benemei, S., et al., 2012. TRPA1 and TRPV4 mediate paclitaxel-induced peripheral neuropathy in mice via a glutathione-sensitive mechanism. Pflugers Arch., 463(4):561–569. [doi:10.1007/s00424-011-1071-x]
McNamara, C.R., Mandel-Brehm, J., Bautista, D.M., et al., 2007. TRPA1 mediates formalin-induced pain. PNAS, 104(33):13525–13530. [doi:10.1073/pnas.0705924104]
Merrill, A.W., Cuellar, J.M., Judd, J.H., et al., 2008. Effects of TRPA1 agonists mustard oil and cinnamaldehyde on lumbar spinal wide-dynamic range neuronal responses to innocuous and noxious cutaneous stimuli in rats. J. Neurophysiol., 99(2):415–425. [doi:10.1152/jn.00883.2007]
Nieto, F.R., Cendán, C.M., Sánchez-Fernández, C., et al., 2012. Role of sigma-1 receptors in paclitaxel-induced neuropathic pain in mice. J. Pain, 13(11):1107–1121. [doi:10.1016/j.jpain.2012.08.006]
Niiyama, Y., Kawamata, T., Yamamoto, J., et al., 2009. SB366791, a TRPV1 antagonist, potentiates analgesic effects of systemic morphine in a murine model of bone cancer pain. Br. J. Anaesthesiol., 102(2):251–258. [doi:10.1093/bja/aen347]
Oh, G.S., Pae, H.O., Seo, W.G., et al., 2001. Capsazepine, a vanilloid receptor antagonist, inhibits the expression of inducible nitric oxide synthase gene in lipopolysaccharide-stimulated RAW264.7 macrophages through the inactivation of nuclear transcription factor-kappa B. Int. Immunopharmacol., 1(4):777–784. [doi:10.1016/S1567-5769(01)00012-1]
O’Neill, J., Brock, C., Olesen, A.E., et al., 2012. Unravelling the mystery of capsaicin: a tool to understand and treat pain. Pharmacol. Rev., 64(4):939–971. [doi:10.1124/pr.112.006163]
Pascual, D., Goicoechea, C., Burgos, E., et al., 2010. Antinociceptive effect of three common analgesic drugs on peripheral neuropathy induced by paclitaxel in rats. Pharmacol. Biochem. Behav., 95(3):331–337. [doi:10.1016/j.pbb.2010.02.009]
Pevida, M., Lastra, A., Hidalgo, A., et al., 2013. Spinal CCL2 and microglial activation are involved in paclitaxelevoked cold hyperalgesia. Brain Res. Bull., 95:21–27. [doi:10.1016/j.brainresbull.2013.03.005]
Ray, A.M., Benham, C.D., Roberts, J.C., et al., 2003. Capsazepine protects against neuronal injury caused by oxygen glucose deprivation by inhibiting Ih. J. Neurosci., 23(31):10146–10153.
Ren, K., Dubner, R., 1999. Inflammatory models of pain and hyperalgesia. ILAR J., 40(3):111–118. [doi:10.1093/ilar.40.3.111]
Rigo, F.K., Dalmolin, G.D., Trevisan, G., et al., 2013. Effect of ω-conotoxin MVIIA and Phα1β on paclitaxel-induced acute and chronic pain. Pharmacol. Biochem. Behav., 114-115:16–22. [doi:10.1016/j.pbb.2013.10.014]
Rios, E.R., Rocha, N.F., Carvalho, A.M., et al., 2013. TRP and ASIC channels mediate the antinociceptive effect of citronellyl acetate. Chem. Biol. Interact., 203(3):573–579. [doi:10.1016/j.cbi.2013.03.014]
Sałat, K., Filipek, B., Wieckowski, K., et al., 2009. Analgesic activity of 3-mono-substituted derivatives of dihydrofuran-2-one in experimental rodent models of pain. Pharmacol. Rep., 61(5):807–818. [doi:10.1016/S1734-1140(09)70136-7]
Salat, K., Librowski, T., Moniczewski, A., et al., 2012a. Analgesic, antioedematous and antioxidant activity of γ-butyrolactone derivatives in rodents. Behav. Pharmacol., 23(4):407–416. [doi:10.1097/FBP.0b013e3283566042]
Salat, K., Moniczewski, A., Salat, R., et al., 2012b. Analgesic, anticonvulsant and antioxidant activities of 3-[4-(3-trifluoromethyl-phenyl)-piperazin-1-yl]-dihydrofuran-2-one dihydrochloride in mice. Pharmacol. Biochem. Behav., 101(1):138–147. [doi:10.1016/j.pbb.2011.12.014]
Sałat, K., Gawlik, K., Witalis, J., et al., 2013a. Evaluation of antinociceptive and antioxidant properties of 3-[4-(3-trifluoromethyl-phenyl)-piperazin-1-yl]-dihydrofuran-2-one in mice. Naunyn Schmiedeberg’s Arch. Pharmacol., 386(6):493–505. [doi:10.1007/s00210-013-0847-2]
Sałat, K., Moniczewski, A., Librowski, T., 2013b. Transient receptor potential channels-emerging novel drug targets for the treatment of pain. Curr. Med. Chem., 20(11): 1409–1436. [doi:10.2174/09298673113209990107]
Sandkühler, J., 2009. Models and mechanisms of hyperalgesia and allodynia. Physiol. Rev., 89(2):707–758. [doi:10.1152/physrev.00025.2008]
Santos, A.R., Calixto, J.B., 1997. Ruthenium red and capsazepine antinociceptive effect in formalin and capsaicin models of pain in mice. Neurosci. Lett., 235(1–2):73–76. [doi:10.1016/S0304-3940(97)00722-2]
Sawynok, J., Liu, X.J., 2003. The formalin test: characteristics and usefulness of the model. Rev. Analg., 7(2):145–163. [doi:10.3727/000000003783992982]
Walker, K.M., Urban, L., Medhurst, S.J., et al., 2003. The VR1 antagonist capsazepine reverses mechanical hyperalgesia in models of inflammatory and neuropathic pain. J. Pharmacol. Exp. Ther., 304(1):56–62. [doi:10.1124/jpet.102.042010]
Westlund, K.N., Kochuko, M.Y., Lu, Y., et al., 2010. Impact of central and peripheral TRPV1 and ROS levels on proinflammatory mediators and nociceptive behavior. Mol. Pain, 6:46. [doi:10.1186/1744-8069-6-46]
Xiao, W.H., Zheng, H., Bennett, G.J., 2012. Characterization of oxaliplatin-induced chronic painful peripheral neuropathy in the rat and comparison with the neuropathy induced by paclitaxel. Neuroscience, 203:194–206. [doi:10.1016/j.neuroscience.2011.12.023]
Zhao, M., Isami, K., Nakamura, S., et al., 2012. Acute cold hypersensitivity characteristically induced by oxaliplatin is caused by the enhanced responsiveness of TRPA1 in mice. Mol. Pain, 8:55. [doi:10.1186/1744-8069-8-55]
Author information
Authors and Affiliations
Corresponding author
Additional information
Project supported by the National Science Centre Grant (No. DEC-2012/05/B/NZ7/02705), Poland
ORCID: Kinga SAŁAT, http://orcid.org/0000-0003-0614-5393
Rights and permissions
About this article

Cite this article
Sałat, K., Filipek, B. Antinociceptive activity of transient receptor potential channel TRPV1, TRPA1, and TRPM8 antagonists in neurogenic and neuropathic pain models in mice. J. Zhejiang Univ. Sci. B 16, 167–178 (2015). https://doi.org/10.1631/jzus.B1400189
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1631/jzus.B1400189
Key words
- Allyl isothiocyanate
- Capsaicin
- Formalin
- Neurogenic pain
- Transient receptor potential channels
- Paclitaxel-induced sensory neuropathy