


Abstract
For several decades, it has been suggested that dopamine (DA), especially in nucleus accumbens, mediates the primary reinforcing characteristics of natural stimuli such as food, as well as drugs of abuse. Yet, several fundamental aspects of primary food reinforcement, motivation, and appetite are left intact after interference with accumbens DA transmission. Recent studies have shown that accumbens DA is involved in responsiveness to conditioned stimuli and activational aspects of motivation. In concurrent choice tasks, accumbens DA depletions cause animals to reallocate their choice behavior in the direction of instrumental behaviors that involve less effort. Also, an emerging body of evidence has demonstrated that the effects of accumbens DA depletions on instrumental food-seeking behavior can vary greatly depending upon the task. For example, some schedules of reinforcement are insensitive to the effects of DA depletions, whereas others are highly sensitive (e.g., large fixed ratios). Accumbens DA depletions slow the rate of operant responding, blunt the rate-facilitating effects of moderate-sized ratios, and enhance the rate-suppressing effects of very large ratios (i.e., produce ratio strain). Accumbens DA may be important for enabling rats to overcome behavioral constraints, such as work-related response costs, and may be critical for the behavioral organization and conditioning processes that enable animals to engage in vigorous responses, such as barrier climbing, or to emit large numbers of responses in ratio schedules in the absence of primary reinforcement. The involvement of accumbens DA in activational aspects of motivation has implications for energy-related disorders in psychiatry, as well as aspects of drug-seeking behavior.
The Dopamine (DA)/Reward Hypothesis and the General Anhedonia Model
For several decades, it has been suggested that DA, especially in nucleus accumbens, mediates the primary reinforcing characteristics of natural stimuli such as food (Wise, 1982; Smith, 1995), as well as drugs of abuse (Wise, 1982). The supposition that drugs of abuse act by turning on the brain's natural “reward system” (i.e., the “General Anhedonia Model”; Salamone et al., 1997) has been a dominant theoretical approach for many years. Despite the persistent popularity of this hypothesis, however, there are substantial problems with the notion that accumbens DA mediates reinforcement for natural stimuli such as food (Salamone et al., 1997; Salamone and Correa, 2002). Considerable evidence supports a role for DA systems in various aspects of instrumental behavior, learning, stimulus salience, and responsiveness to the environment, but does not support the notion of a selective involvement of accumbens DA in mediating the primary appetitive motivation processes that underlie positive reinforcement (Salamone and Correa, 2002). DA antagonists and accumbens DA depletions also impair performance on aversively motivated tasks involving avoidance, punishment, place aversion, and taste aversion (Salamone, 1994; Salamone et al., 1997; Killcross et al., 1997; Di Chiara, 2002;Huang and Hsiao, 2002). Accumbens DA transmission is elevated in response to both appetitive and aversive conditions (Salamone, 1994,1996; Salamone et al., 1997; Datla et al., 2002). A detailed examination of the so-called “extinction effect” fails to support the hypothesis that accumbens DA mediates food reward (Salamone, 1986;Salamone et al., 1995, 1997; Salamone and Correa, 2002). Moreover, fundamental aspects of food reinforcement are intact after DA antagonism or accumbens DA depletions (Salamone and Correa, 2002; Baldo et al., 2002). These considerations have led some scientists to question the adequacy of the idea that all drugs of abuse are simply activating the brain's natural reward system (Salamone et al., 1997; Salamone and Correa, 2002).
The present review will discuss some of the problems with the DA hypothesis of reward, but merely as a point of departure for the subsequent consideration of alternative hypotheses. If indeed the field is currently going through some restructuring, then it is critical to consider additional theoretical approaches. Several researchers have emphasized that accumbens DA is involved in various functions related to reinforcement or incentive motivation (Salamone et al., 1997;Berridge and Robinson, 1998; Ikemoto and Panksepp, 1999; Kelley, 1999;Cardinal et al., 2002; Di Chiara, 2002; Salamone and Correa, 2002). The primary focus of the present article is the principle that interference with accumbens DA transmission produces behavioral effects that interact strongly with the response requirements of the task being used (Salamone et al., 1997; Salamone and Correa, 2002). This has led to the suggestion that a major function of accumbens DA is to promote expenditure of effort in instrumental tasks, which is consistent with the long-standing hypothesis that accumbens DA is critically involved in activational aspects of motivation (Salamone, 1988). The discussion below will summarize this literature in detail and will consider its implications for understanding the brain mechanisms involved in expenditure of effort and for aspects of psychiatry and drug abuse.
Preserved Aspects of Primary Food Reinforcement and Food Motivation after DA Antagonism or Accumbens DA Depletion: Problems with the General DA/Reward Hypothesis
Animals will self-administer stimulants directly into nucleus accumbens, and there is little disagreement with the idea that interference with accumbens DA transmission has profound effects upon stimulant self-administration (Roberts et al., 1977; Caine and Koob, 1994; Chevrette et al., 2002). Of course, there are disagreements as to the precise interpretation of these findings, the possible role of other transmitters or other brain regions, and the possible substrates for nonstimulant drugs of abuse. Moreover, there are substantial difficulties with the notion that accumbens DA mediates the primary motivating effects of all reinforcers, including natural stimuli such as food. It has been suggested that the hypothesized involvement of accumbens DA in food reinforcement is essentially the linchpin of the general form of the DA reward hypothesis (Salamone and Correa, 2002) because it is supposedly the DA-mediated natural reward system in the brain that is being turned on by drugs of abuse. Nevertheless, there are studies showing that accumbens DA depletions that impair stimulant self-administration have little effect on food-reinforced operant behavior on some schedules (Roberts et al., 1977; Caine and Koob, 1994). Such observations warrant a detailed and critical examination of the hypothesis that accumbens DA mediates the primary reinforcing or motivating effects of natural stimuli such as food. The results of this examination point to severe deficiencies in the general form of the DA hypothesis of reward (Salamone et al., 1997, 1999; Salamone and Correa, 2002).
The suppressive effects of systemic injections of DA antagonists on food-reinforced lever pressing have been attributed to actions on food reward, motivation, or appetite (Wise, 1982; Smith, 1995). Although Berridge, Robinson, and colleagues have argued consistently that accumbens DA does not mediate food-induced hedonic reactions (i.e., “liking”), they have suggested that forebrain DA is involved in aspects of incentive salience (i.e., “wanting”) for food, which includes reinforcer-seeking behavior as well as components of appetite and aspects of food attraction and intake (Berridge and Robinson, 1998;Wyvell and Berridge, 2001). Yet, there are numerous problems with the notion that DA systems directly mediate primary food reinforcement, motivation, or appetite. DA antagonists suppress sucrose intake, which has been claimed to represent part of the “proof” for the DA hypothesis of reward (Smith, 1995). Nevertheless, neuroleptic-induced deficits in sucrose intake are accompanied by impairments in various oral motor functions, including lick duration, lick force, lap volume, lick efficiency, and tongue extension (Fowler and Mortell, 1992; Das and Fowler, 1996; Salamone and Correa, 2002). The effects on sucrose drinking produced by DA antagonists have been interpreted as indicating a reduced effort for obtaining the sucrose (Hsiao and Chen, 1995) and as a lack of sensorimotor responsiveness to a taste stimulus (Muscat and Willner, 1989). Although feeding is impaired by higher doses of DA antagonists, there is little evidence that this reflects a loss of appetite, and several lines of evidence indicate that these deficits are related to motor dysfunctions (for review, see Salamone and Correa, 2002).
If low doses of DA antagonists suppress food-reinforced lever pressing by reducing appetite, then food intake and lever pressing should be suppressed within the same dose range. Yet, DA antagonists suppress food-reinforced lever pressing at doses lower than those that suppress food intake or simple appetitive responses for food (Fibiger et al., 1976; Salamone, 1986; also see work by E. T. Rolls, S. Cooper, J. Horvitz, A. Ettenberg, T. Ljungberg, and others). Several studies have used a concurrent choice task in which rats can lever press for a preferred food (Bioserve pellets) or can approach and consume a less preferred food (lab chow) that is available in the chamber (Salamone et al., 1991; Cousins et al., 1993; Sokolowski et al., 1998; Nowend et al., 2001). Rats that press on a fixed ratio (FR)1 (i.e., one lever press per food pellet) or FR5 schedule typically get most of their food by lever pressing and only consume small amounts of chow. Prefeeding to reduce food motivation suppressed both lever pressing and chow intake (Salamone et al., 1991). In contrast, low-to-moderate doses of DA antagonists produce a very different effect. The DA antagonists cis-flupenthixol, haloperidol, raclopride, SCH 23390, and SKF 83566 all decreased lever pressing for food but substantially increased chow intake (Salamone et al., 1991, 1997, 2002; Koch et al., 2000). The low dose of haloperidol that produced this shift in behavior did not alter food intake or preference in free-feeding choice tests (Salamone et al., 1991). The D1 antagonist SKF 83566 and the D2 antagonist raclopride reduced FR5 lever pressing and substantially increased chow intake, whereas the serotonergic appetite suppressant fenfluramine suppressed both activities (Salamone et al., 2002). These findings demonstrate that interference with DA transmission does not simply reduce appetite. Rats treated with low doses of DA antagonists remain directed toward the acquisition and consumption of food, indicating that fundamental aspects of food reinforcement and primary food motivation remain intact (Salamone et al., 1991; Salamone and Correa, 2002).
Fundamental aspects of food motivation are preserved after local interference with accumbens DA transmission (Salamone et al., 1993a,1997; Cousins et al., 1993; Ikemoto and Panksepp, 1999; Kelley, 1999). Intra-accumbens infusions of doses of DA antagonists that impaired locomotor activity, runway performance, and conditioned reinforcement failed to impair sucrose consumption (Ikemoto and Panksepp, 1999; Baldo et al., 2002). Although forebrain DA depletion severely impairs feeding, considerable evidence indicates that this effect is dependent upon DA depletions in ventrolateral neostriatum and is related to orofacial and forepaw motor deficits and sensorimotor impairments that result from DA depletions in this region (Jicha and Salamone, 1991;Salamone et al., 1993a). Accumbens DA depletions do not impair feeding upon lab chow (Koob et al., 1978; Salamone et al., 1993a; Kelley, 1999). Moreover, detailed analyses failed to find any effect of accumbens DA depletions on food intake, feeding rate, food handling, or time spent feeding (Salamone et al., 1993a). Because time allocation has been viewed as a critical behavioral marker of reinforcement value, these results suggest that accumbens DA depletions do not blunt food reinforcement. As described above, studies with a concurrent choice task demonstrated that low doses of DA antagonists decreased lever pressing and increased chow intake. This effect is not produced by local depletion of ventrolateral neostriatal DA, which instead results in severe motor impairments that decrease both behaviors (Cousins et al., 1993). Rather, decreases in lever pressing and increases in chow intake result from accumbens DA depletions, as well as from intra-accumbens DA antagonism (Salamone et al., 1991; Cousins et al., 1993; Cousins and Salamone, 1994; Koch et al., 2000; Nowend et al., 2001). The shift from lever pressing to chow intake occurs if either a D1 or D2 family antagonist is used (Salamone et al., 1991; Nowend et al., 2001) and occurs with injections into either medial core, lateral core, or medial shell subregions of the accumbens (Salamone et al., 1991; Nowend et al., 2001). Thus, although lever pressing is decreased by interference with accumbens DA transmission, DA-depleted rats show a compensatory reallocation of behavior and select a new path to an alternative food source (i.e., concurrently available chow). This provides further evidence that important aspects of the primary or unconditioned reinforcing properties of food are intact after interference with accumbens DA transmission. In addition, these results have been interpreted as showing that aspects of incentive salience (i.e., wanting) are impaired by accumbens DA depletions (e.g., tendency to work for food), whereas other aspects of wanting are relatively intact after accumbens DA depletions (e.g., tendency to consume food, aspects of appetite; see Salamone and Correa, 2002).
In view of the hypothesized reward functions of accumbens DA and the robust effects of accumbens DA depletions on stimulant-reinforced behavior, one of the startling findings to emerge has been the relative inability of accumbens DA depletions to impair performance on some schedules of reinforcement. Accumbens DA depletions that severely impaired amphetamine-reinforced behavior failed to disrupt food-reinforced responding on a variable ratio 2.5 schedule of reinforcement (Roberts et al., 1977). Fixed-interval (FI) 30-s responding (i.e., the first response after a 30-s interval is reinforced) was only mildly affected by accumbens DA depletions (Cousins et al., 1999). Accumbens DA depletions did not significantly affect variable-interval (VI) 30-s responding (Sokolowski and Salamone, 1998; Correa et al., 2002). Several studies have shown that food-reinforced FR1 performance is relatively insensitive to the effects of accumbens DA depletions (McCullough et al., 1993; Salamone et al., 1995; Aberman and Salamone, 1999). The fact that positively reinforced behavior on some schedules is not impaired by accumbens DA depletion or is affected only marginally suggests that maintenance of positively reinforced responding per se is not the key process that is impaired by these depletions. The FR1 is a simple schedule of positive reinforcement that is highly sensitive to extinction and to reinforcer devaluations such as prefeeding (Salamone et al., 1995; Aberman and Salamone, 1999); however, this schedule is relatively insensitive to accumbens DA depletions.
Schedule Dependence of Effects of Accumbens DA Depletions on Operant Performance.
Although some schedules of reinforcement are relatively insensitive to accumbens DA depletions, impairments in food-reinforced lever pressing were observed when schedules that generate high rates of responding or involve larger ratio requirements were used (Salamone et al., 1991, 1993b). These observations led to the suggestion that accumbens DA depletions make animals more sensitive to work requirements on instrumental tasks (Salamone et al., 1991, 1994,1997). One way of varying work requirements on operant tasks is to vary the size of the ratio (i.e., the number of responses required for each reinforcer). Aberman and Salamone (1999) studied various ratio requirements (i.e., FR1, 4, 16, and 64) and observed that the effects of accumbens DA depletions were highly schedule-dependent. Accumbens DA depletions did not impair FR1 responding and produced only a mild and transient effect on FR4 responding. Lever pressing on the FR16 schedule was reduced by DA depletions, whereas the most sensitive schedule in this experiment was the FR64, which showed catastrophic effects. DA depleted rats on the FR64 schedule showed “ratio strain”, i.e., their responding was not merely reduced in rate but rather was suppressed so severely that many of them essentially ceased pressing. Several factors could be responsible for the differences in sensitivity to DA depletion shown under these diverse schedule conditions. The FR64 schedule has a relatively low reinforcement density compared with the FR1 (i.e., approximately a 5- to 7-fold difference in total food obtained). In addition, the FR64 schedule generates a high rate of baseline responding (i.e., 2500–3000 responses/30 min), whereas the FR1 generates a relatively low rate (i.e., 250–350 responses/30 min). A recent experiment (Salamone et al., 2001) was designed to investigate the relevance of such factors. The effects of accumbens DA depletions were investigated using six schedules: FR5, 20, 50, 100, 200, and 300. In the first three schedules, the reinforcement consisted of one food pellet per ratio completed. To maintain the same programmed reinforcement density in the remaining schedules, the reinforcement per ratio completed was increased to two pellets for FR100, four pellets for FR200, and six pellets for FR300. Rats were trained before surgery and were able to maintain high levels of responding on all schedules. After surgery, rats with accumbens DA depletions exhibited behavioral deficits that were highly dependent upon the ratio requirement. There were small and transient effects of DA depletion on FR5 performance, but as the ratio value got larger, the impairment became greater. On the FR20 and 50 schedules, response rates were reduced by DA depletions. Responding on the FR200 and 300 schedules was severely impaired, and on the last day of FR300 testing, no DA-depleted rats completed a single ratio. Thus, accumbens DA depletions again were shown to enhance ratio strain, making rats extremely sensitive to high-ratio requirements (Salamone et al., 2001). Baseline levels of responding and molar obtained reinforcement densities under control conditions were approximately the same across the FR50 to FR300 schedules. Therefore, the induction of ratio strain by DA depletions was relatively independent of the baseline rate of responding and the overall density of food reinforcement. The results of these studies demonstrate that accumbens DA depletions alter the relation between ratio requirement and response output, with two major consequences; these depletions slow the maximal response rate and induce ratio strain (see Fig. 1).

A, effect of accumbens DA depletions on FR1, 4, 16, and 64 responding, from Aberman and Salamone (1999). B, effects of accumbens DA depletions [6-hydroxydopamine (6-OHDA)] on FR5, 20, 50, 100, 200, and 300 from Salamone et al. (2001). In each study, responding was suppressed by accumbens DA depletions in a schedule-dependent fashion. In both cases, depletion of DA in nucleus accumbens reduced the maximal rate of responding (i.e., – – –; ceiling). In addition, responding on higher ratios showed “ratio strain”; i.e., responding was suppressed so severely that many animals essentially ceased responding (arrow).
Although baseline response rate was not a critical factor for inducing ratio strain in the Salamone et al. (2001) experiment, the response slowing on some schedules that is seen after accumbens DA depletions appears to be highly related to baseline response rate. Over the last few years, a number of schedules have been investigated, including various fixed and progressive ratios, FI 30-s, VI 30-s, and tandem VI/FR schedules (Salamone et al., 1993b; Aberman and Salamone, 1999;Cousins et al., 1999; Correa et al., 2002). Across these schedules, there is a linear relation between baseline rate of responding under control conditions and the degree of suppression produced by accumbens DA depletions, such that the deficit is greater with schedules that generate increased response rates (Fig.2; see review by Salamone et al., 1999). Another important aspect of the response slowing produced by accumbens DA depletions is that it is characterized by alterations in the inter-response time distribution (Salamone et al., 1993b; Sokolowski and Salamone, 1998; Salamone et al., 1999). DA depletions decrease the relative number of fast inter-response times and increase the relative number of pauses in responding.

Scatterplot showing the relation between baseline or control rates of responding on various interval and ratio schedules of reinforcement and the extent to which accumbens DA depletions suppress responding (expressed as a percentage of control responding) on that schedule. Unmarked data points and least-squares regression line are from Salamone et al. (1999). Additional data points are added for the tandem VI 30-s/FR1 and VI 30-s/FR5 schedules from Correa et al. (2002). All these studies were conducted with the same operant boxes, using the same type of lever. Caull and Brindle (2001) have shown similar effects with the effects of DA antagonists.
Economic and Ergonomic Approaches to the Study of Accumbens DA Function.
As described above, interference with accumbens DA transmission does not appear to blunt food-related hedonic reactions (Berridge and Robinson, 1998), nor does it suppress appetite (Salamone et al., 2002). Instead, it appears to produce “anergia” or “psychomotor slowing” (Salamone et al., 1994, 1997, 1999); rats with accumbens DA depletions are generally less active and are less likely to work for significant stimuli such as food. This summary certainly does not capture all the manifestations of interference with DA transmission (e.g., see discussions of accumbens DA, learning, and information processing in Ikemoto and Panksepp, 1999; Kelley, 1999;Cardinal et al., 2002; Datla et al., 2002; see also electrophysiology studies by A. Grace, P. O'Donnell, C. Pennartz, T. Nicola, and others). Nevertheless, it is a useful synthesis of a large portion of the work in this area and summarizes at least one of the major functions performed by accumbens DA. Accumbens DA depletions suppress spontaneous and amphetamine-induced locomotor activity (Koob et al., 1978; Robbins et al., 1983; Cousins et al., 1993; Correa et al., 2002). Schedule-induced activities, such as locomotion, licking, and excessive drinking, also are reduced by accumbens DA depletions (Robbins et al., 1983; McCullough and Salamone, 1992). Accumbens DA release is enhanced during scheduled food presentation in a manner that is correlated with the degree of locomotor activity (McCullough and Salamone, 1992), and DA release during lever pressing is increased in a manner that is correlated with response rate (McCullough et al., 1993; Salamone et al., 1994; Sokolowski et al., 1998; Cousins et al., 1999), but not with total amount of food received (McCullough and Salamone, 1992;Salamone et al., 1994; Sokolowski et al., 1998). Based upon these observations and the evidence described above, it is reasonable to suggest that accumbens DA plays a major role as an activator, or invigorator, of various behaviors observed in the context of instrumental conditioning.
The idea that activation and effort are vital components of instrumental behavior or foraging is not a recent invention. This has been a persistent theme in psychology for several decades (Salamone, 1988; Salamone and Correa, 2002; and references cited therein). For example, in 1964, Cofer and Appley (referenced in Salamone and Correa, 2002) hypothesized the existence of an “anticipation-invigoration mechanism” triggered by conditioned stimuli, which invigorated instrumental behavior. This suggestion seems particularly prescient in view of the modern concepts that accumbens DA is involved in behavioral activation and responsiveness to conditioned stimuli (Salamone, 1986,1988; McCullough and Salamone, 1992; Salamone et al., 1997; Ikemoto and Panksepp, 1999; Wyvell and Berridge, 2001; Cardinal et al., 2002; Lopez and Ettenberg, 2002) and that accumbens DA transmission is activated by arousing stimuli, conditioned stimuli that predict reward, and during the instrumental phase of behavior in anticipation of reinforcer delivery (Richardson and Gratton, 1996; Salamone, 1996;Sokolowski et al., 1998; Datla et al., 2002; Horvitz, 2002; Kiyatkin, 2002). Pavlovian and instrumental associative factors, motivational processes, and work requirements of the schedule all serve to regulate instrumental behavior (Salamone, 1992; Salamone et al., 1997;Aberman and Salamone, 1999; Cardinal et al., 2002). Instrumental behavior can be viewed as a behavioral adaptation to the constraints imposed by schedule contingencies. For several years, “economic models” have described how instrumental behavior represents a constant interaction between the tendency to obtain a commodity (i.e., reinforcer) and the response costs involved in procurement (e.g., Hursh et al., 1988). DA antagonism and accumbens DA depletions alter the outcome of cost/benefit analyses in behavior, change work-related elasticity of demand for food, and reduce the tendency to overcome work-related response costs attached to instrumental contingencies (Salamone et al., 1991, 1997, 2001, 2002; Cousins et al., 1993; Cousins and Salamone, 1994; Hsiao and Chen, 1995; Aberman and Salamone, 1999;Neill et al., 2002).
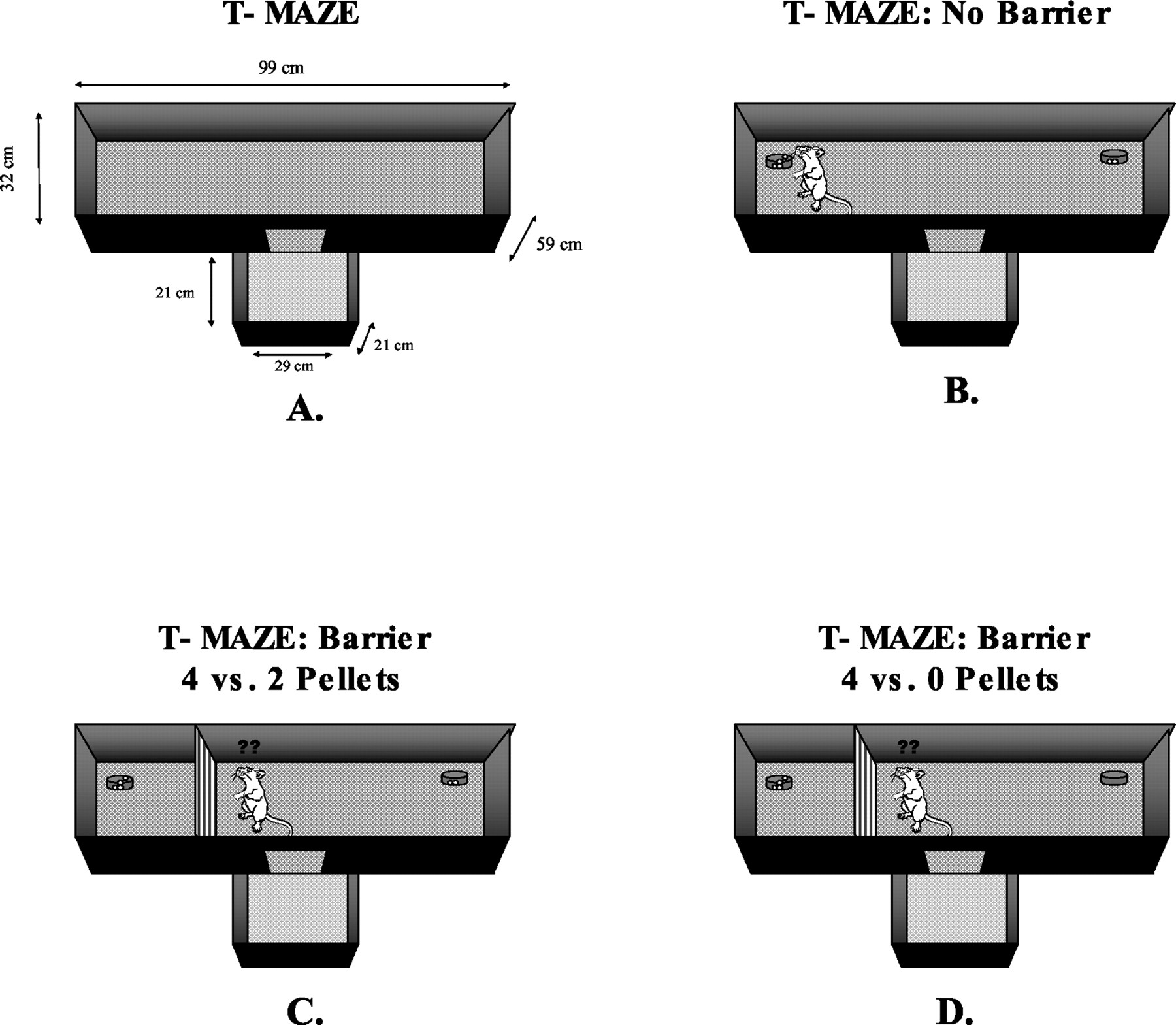
A T-maze task was developed to assess the effects of accumbens DA depletions on response choice based upon task difficulty (Salamone et al., 1994; Cousins et al., 1996; Fig. 3). The two choice arms of the maze had different reinforcement densities (e.g., four versus two food pellets, or four versus zero), and under some conditions, a 44-cm barrier was placed in the arm with the higher density of food reinforcement to vary task difficulty. When no barrier was present, rats chose the arm with the high reinforcement density, and neither haloperidol nor accumbens DA depletion altered their response choice (Salamone et al., 1994). When the arm with the barrier contained four pellets, but the other arm contained no pellets, rats with accumbens DA depletions were very slow, but still chose the high-density arm, climbed the barrier, and consumed the pellets (Cousins et al., 1996). Accumbens DA depletions dramatically altered choice behavior when the high-density arm (four pellets) had the barrier present and the arm without the barrier contained an alternative food source (two pellets). In this case, DA-depleted rats showed a decreased choice for the high-density arm and an increased choice for the low-density arm (Salamone et al., 1994; Cousins et al., 1996). Thus, accumbens DA depletions or DA antagonism can alter maze choice behavior based upon the response requirement (Salamone et al., 1994; Cousins et al., 1996), lead to a reallocation of behavior away from lever pressing and toward chow intake in the concurrent choice task (Salamone et al., 1991; Cousins et al., 1993; Koch et al., 2000; Nowend et al., 2001), and enhance ratio strain in operant schedules (Aberman and Salamone, 1999; Salamone et al., 2001). These diverse studies demonstrate that accumbens DA is important for enabling rats to overcome behavioral constraints such as work-related procurement costs.

Concurrent choice T-Maze procedure used in Salamone et al. (1994) and Cousins et al. (1996). A, dimensions of the T-Maze. B to D, various procedures that have been used. B, four pellets in one arm, two pellets in the other, with no barrier. C, four pellets in one arm, two pellets in the other, with a 44-cm barrier blocking the arm with the higher density of food. D, four pellets in one arm, no pellets in the other, with a 44-cm barrier blocking the arm with food reinforcement. See the text for results of accumbens DA depletions. This procedure recently has been used to assess the effects of prefrontal cortex lesions as well (M. Walton and colleagues, unpublished manuscript).
The Significance of Ratio, Force, and Time Requirements as Factors That Modulate Sensitivity to the Effects of Accumbens DA Depletions
Although it is clear that the effects of accumbens DA depletions interact with work contingencies such as ratio requirements, the specific aspects of work that make animals sensitive to DA depletion remain somewhat uncertain. A detailed examination of the literature suggests that rats with accumbens DA depletions are sensitive to a combination of factors, which can be present in some tasks but not others. For example, accumbens DA depletions make rats differentially sensitive to ratio requirements such as FR1 versus FR5, but DA-depleted rats do not appear to show heightened sensitivity to force requirements, such as the addition of weights on the lever up to 96 g (S. M. Weber, J. D. Salamone, unpublished observations). As noted above, the response slowing produced by accumbens DA depletions across a large group of schedules is directly related to the baseline rate of responding. Nevertheless, baseline rate of responding cannot be the only factor that determines sensitivity to DA depletion because very large ratio schedules are highly sensitive to the effects of accumbens DA depletion, even if those high ratios do not result in increased baseline response rates (Salamone et al., 2001). Schedules with very large ratios (i.e., FR200, 300), which generate large numbers of responses over long periods of time in the absence of primary reinforcement, are particularly sensitive to the effects of accumbens DA depletions (Salamone et al., 2001). Consistent with this observation, it was noted that rats with accumbens cell body lesions showed reduced preference for delayed reinforcement, which may indicate that the accumbens is necessary for sustaining responding during delayed or intermittent reinforcement (Cardinal et al., 2001, 2002). Yet, it does not appear as though intermittence of reinforcement per se is the primary factor determining sensitivity to accumbens DA depletions. If intermittence (i.e., periods of time or large numbers of responses without reinforcement) were the key factor, then interval schedules, which have programmed periods of time with no reinforcement, should be particularly sensitive to the effects of accumbens DA depletions. In fact, FI 30- or 60-s performance was shown to be only marginally affected by accumbens DA depletions (Robbins et al., 1983;Cousins et al., 1999). Two recent studies reported that accumbens DA depletions did not impair performance on a VI 30-s schedule (Sokolowski and Salamone, 1998; Correa and Salamone, 2002).
Correa et al. (2002) used VI schedules of reinforcement to investigate the role of intermittence as a determinant of sensitivity to accumbens DA depletions. Two VI 30-s schedules were used, each with different response requirements added to the interval requirement. For one of the schedules, the rats were reinforced for the first response after the interval elapsed (VI 30 s). For the other schedule, an additional work requirement was attached by requiring rats to make five responses after the interval to receive reinforcement (tandem VI 30/FR5). Attachment of the additional work requirement led to greater response rates during presurgical training. The effects of DA depletion on responding were highly schedule-dependent (Correa et al., 2002). DA depletions had no significant effect on lever pressing under the condition with a low-response requirement (VI 30-s), but these depletions substantially disrupted responding on the schedule with the higher work requirement (VI 30/FR5). These results indicate that intermittence of reinforcement in itself, at least within the time intervals tested, is not the major determinant of the response slowing produced by accumbens DA depletions on some operant schedules. It is possible that several factors, such as ratio requirements, intermittence, and reliance on conditioned stimuli to elicit and sustain responding in the absence of primary reinforcement, combine to make some schedules sensitive to the effects of accumbens DA depletions. Indeed, one of the patterns that has emerged recently is that there is something special about ratio requirements, which makes responding particularly sensitive to accumbens DA depletions. It is possible that the activating effects of conditioned stimuli have different characteristics in interval and ratio schedules. With ratio schedules, there is an important feedback relation between response rate and reinforcement rate, and during ratio performance the responses themselves are the best predictors of reinforcement. Another important factor may be that, for low-to-moderate value ratio schedules, increasing the ratio value tends to increase response rate. The higher response rate in the VI 30/FR5 schedule can be seen as an adaptation to the constraint of having an additional ratio requirement attached to the schedule (Correa et al., 2002). Rats increase reinforcement density in schedules with ratio components (including the VI 30/FR5) by increasing response output, and one of the effects of accumbens DA depletions may be to reduce the activating (i.e., rate-enhancing) effects of ratio requirements. Summarizing across a number of experiments, it appears as though accumbens DA depletions slow the local rate of responding, blunt the rate-facilitating effects of moderate sized ratios, and enhance the rate-suppressing effects of very large ratios (i.e., produce ratio strain).
Nucleus Accumbens DA Facilitates Sustained Effort over Time for Reinforcer-Seeking Behavior: Implications for Psychology and Psychiatry
It is clear that work-related response costs are an important factor in determining which operant schedules or instrumental tasks are sensitive to the effects of accumbens DA depletions. Yet, one should not assume that force requirements or caloric expenditure, in the most direct sense, are the primary factors that make work more difficult in DA-depleted animals. Various forms of work, including schedules of reinforcement, not only have force requirements, but they also have skill requirements, temporal components, and even cognitive aspects, which powerfully influence the perceived difficulty of a task. We still do not fully understand which particular aspects of effort make some tasks relatively sensitive to dopaminergic manipulations, and research needs to continue on this important issue. Nevertheless, we can state with reasonable support that the effects of accumbens DA depletions differ depending upon the response requirements of the task. A reasonable working hypothesis, based upon the review above, is that interference with accumbens DA impairs the exertion of sustained effort over time (Salamone et al., 2001; Neill et al., 2002; Salamone and Correa, 2002). Interference with accumbens DA appears to blunt the activating effects of conditioned stimuli (e.g., explicit cues, contextual, or temporal stimuli) that elicit and sustain instrumental responding, as well as schedule-induced behaviors, in the absence of primary reinforcement. Indeed, rats with accumbens DA depletions are highly dependent upon primary reinforcement and have great difficulty sustaining ratio performance over time (Salamone et al., 2001). Ratio schedules are particularly sensitive to the reduction in behavioral activation produced by accumbens DA depletions. DA depletions may reduce the response-facilitating impact of the ratio requirement, may alter the feedback relation between responding and reinforcement (i.e., reduced ratio responding lowers reinforcement rate, which reduces behavioral activation, which in turn suppresses responding further), or may enhance the impact of the perceived work load stemming from the ratio requirement. Such work-related factors, combined with reduced impact of conditioned stimuli, may render ratio schedules highly sensitive to the effects of interference with accumbens DA, and some of these factors may lead to the behavioral reallocation observed in operant and T-maze concurrent choice tasks.
Of course, accumbens DA has a functional significance beyond the performance of ratio schedules or maze tasks in rats. It is likely that the behavioral activation functions of accumbens DA have important implications for a variety of different fields. The notions that organisms make decisions based upon cost/benefit analyses and that work requirements shape choice behavior are important ideas in several fields, including ethology, economics, and industrial psychology. Although anhedonia often is stressed as a symptom of depression, it should also be emphasized that anergia, or psychomotor slowing, is a critical aspect of depression in many people (Stahl, 2002). Lack of energy is the depressive symptom that is most strongly correlated with the lack of the social function shown by depressed patients and is closely related to various work-related impairments such as days in bed, days of lost work, and low work productivity. In addition, fatigue and disinterest in activities are some of the best predictors of lack of remission with antidepressant drug treatment (Stahl, 2002). Considerable evidence in the psychiatry literature suggests that DA systems play an important role in psychomotor slowing (Willner, 1983;Salamone et al., 1999; Stahl, 2002). In addition, there are people who do not meet the diagnostic criteria for depression, yet they do have a motivational disturbance that has variously been referred to as psychomotor slowing, anergia, or apathy (Campbell and Duffy, 1997), which may involve DA systems.
Whether one agrees or disagrees with the notion that accumbens DA mediates the hedonic or reinforcing effects of food, stimulants, or other drugs of abuse, it is clear that accumbens DA plays a role in drug self-administration. DA systems may provide a substrate for some of the rewarding actions of stimulants but may play a different role for other substances. Although Wise (1982) has been a major proponent of the DA hypothesis of reward, he also has emphasized the role of DA systems in “motivational arousal” and responsiveness to conditioned incentive stimuli (Wise, 1982), which are positions that are similar to those presented above. Several drugs of abuse are psychomotor stimulants, and a number of other drugs of abuse that are “sedatives” also have stimulant properties at low doses, which may mean that motor stimulant effects bear an important relation to features of drug abuse (Wise and Bozarth, 1987). Ethanol is an example of a sedative-hypnotic drug that has stimulant effects at low doses (Correa et al., 1999), and this action may be related to the tendency to work for ethanol during self-administration (Nadal et al., 2002). Recently, effort in drug-seeking or “drug pursuit” behaviors has received greater emphasis as an important factor in the self-administration of a number of drugs, including ethanol (Czachowski et al., 2001), cocaine (Olmstead et al., 2000), and amphetamine (Vezina et al., 2002). Compulsiveness, persistence, and effort in drug-seeking behavior are classic features of drug abuse in humans (Koob et al., 1998). Although it may no longer be tenable to suggest that drugs of abuse are simply turning on the brain's natural reward system (Salamone and Correa, 2002), it clearly is the case that accumbens DA participates in the brain circuitry that regulates vital components of instrumental behavior and motivation (Cardinal et al., 2002; Correa and Salamone, 2002; Di Chiara, 2002; see discussion of the “motive circuit” in Pierce and Kalivas, 1997). This involvement may manifest itself in various aspects of drug abuse (Di Chiara, 2002), including drug seeking behavior, cue-related relapse (Cardinal et al., 2002), and sensitization to components of incentive motivation with repeated drug usage (Pierce and Kalivas, 1997; Berridge and Robinson, 1998).
Acknowledgments
Many thanks to the numerous researchers who have made contributions to the relevant literature (e.g., P. Killeen, G. H. Collier, J. E. R. Staddon, and others) but who could not be cited directly because of space considerations.
Footnotes
-
↵1 Present address: Àrea de Psicobiologia, Campus de Riu Sec, Universitat Jaume I, 12079 Castelló, Spain.
-
Much of the research described in this article was supported by a series of grants to J.S. by the National Science Foundation of the United States.
-
DOI: 10.1124/jpet.102.035063
- Abbreviations:
- DA
- dopamine
- FR
- fixed ratio
- SCH 23390
- R-(+)-7-chloro-8-hydroxy-3-methyl-1-phenyl-2,3,4,5-tetrahydro-1H-3-benzazepine
- VI
- variable interval
- FI
- fixed interval
- SKF 83566
- (±)-7-bromo-1-(fluoresceinyl thioureido)phenyl-8-hydroxy-3-methyl-2,3,4,5,-tetrahydro-1H-benzazepine
- Received October 8, 2002.
- Accepted December 10, 2002.
- The American Society for Pharmacology and Experimental Therapeutics
References
In this issue





{kind=link}
{kind=link}
{kind=link}
Jump to section
- Article
- Abstract
- The Dopamine (DA)/Reward Hypothesis and the General Anhedonia Model
- Preserved Aspects of Primary Food Reinforcement and Food Motivation after DA Antagonism or Accumbens DA Depletion: Problems with the General DA/Reward Hypothesis
- The Significance of Ratio, Force, and Time Requirements as Factors That Modulate Sensitivity to the Effects of Accumbens DA Depletions
- Nucleus Accumbens DA Facilitates Sustained Effort over Time for Reinforcer-Seeking Behavior: Implications for Psychology and Psychiatry
- Acknowledgments
- Footnotes
- References
- Figures & Data
- Info & Metrics
- eLetters