


Abstract
Withdrawal is a potent motivator of drug-seeking behavior in human opiate addicts. Paradoxically, opiate withdrawal reduces dopamine release and suppresses behavioral responding in several animal models of addiction. These findings pose critical questions about how a withdrawal state that depresses dopaminergic and behavioral functioning contributes to drug seeking. This study addressed this issue by investigating factors that increase behavioral activity during opiate withdrawal. Initial experiments revealed that the D2-like agonists propylnorapomorphine HCl (NPA; 0.05–0.4 mg/kg, i.p.) and quinpirole (0.2–0.4 mg/kg, s.c.) each produced strong locomotor activating effects during opiate withdrawal that were not apparent in the absence of withdrawal. Concurrent stereotypy ratings indicated that these effects of NPA and quinpirole during withdrawal were not an indirect consequence of changes in the stereotypy-inducing effects of these drugs. Subsequent experiments showed that locomotion was not increased when opiate withdrawal was induced in the presence of the D1-like agonist SKF 38393 (1.0–8.0 mg/kg, i.p.), that the locomotor activation produced by NPA during withdrawal could be attenuated by the D2-like antagonist eticlopride (0.1–0.2 mg/kg, i.p.), and that locomotor activating effects of NPA could be observed when withdrawal was induced by extracting the implanted morphine pellets, but not when the NPA was given after naltrexone antagonism of acute morphine treatment in nondependent rats. These findings indicate that opiate withdrawal regulates the behavioral impact of D2-like receptor stimulation so that locomotion is markedly increased when these receptors are stimulated during periods of withdrawal. This potentiation may be important for facilitating behavioral responses during periods of opiate detoxification.
Abrupt cessation of chronic opiate use results in a well characterized withdrawal syndrome that includes nausea, dysphoria, and anxiety. These consequences of abstinence are sufficiently adverse that opiate abusers will seek more drug to relieve or prevent the symptoms of withdrawal. In fact, the need to self-medicate is thought to be a primary factor contributing to the maintenance of opiate addictions and a major impediment to successful drug detoxification and rehabilitation (Wikler, 1973; Schulteis and Koob, 1996). This effect of withdrawal has prompted detailed study of the physiological mechanisms responsible for the aversive consequences of opiate withdrawal, with the aim of better understanding the biological basis for opiate abuse.
Research into the physiological mechanisms of withdrawal-induced drug seeking has been guided primarily by the tenets of opponent process theory. This theory contends that chronic opiate use gives rise to adaptive opponent processes that counteract initial drug responses and produce tolerance to the effects of the drug (Solomon, 1980; Koob et al., 1989b). During withdrawal, these opponent processes are thought to be expressed as an aversive “drug-rebound” syndrome that motivates continued drug-seeking behaviors. However, this viewpoint also leads to predictions of decreased drug-seeking behavior during opiate withdrawal. According to opponent process theory, the initial positive motivational effects of opiates should show tolerance with repeated drug use, and termination of opiate use should be characterized by a state of anhedonia and avolition. Consistent with this prediction, animal studies have found that opiate withdrawal can decrease locomotor activity (Schulteis et al., 1994), disrupt operant responding for food incentives (Koob et al., 1989a; Schulteis et al., 1994), elevate current thresholds required to initiate responding for brain-stimulation reward (Schaefer and Michael, 1986; Schulteis et al., 1994), and decrease mesolimbic dopamine (DA) release that is critical for appetitively motivated behavior (Pothos et al., 1991;Rossetti et al., 1992). This evidence for reduced behavioral responsiveness during opiate withdrawal raises a paradox with respect to the role that withdrawal plays in stimulating drug-seeking behaviors. Specifically, how does a state that reduces motivated responding increase the propensity of an addict to seek drugs?
It is noteworthy that opiate withdrawal does not uniformly suppress behavioral responses. In fact, it can facilitate aggressive and defensive reactions elicited by aversive stimulation (Davis and Khalsa, 1971; Harris and Aston-Jones, 1993). Such findings suggest that the impact of opiate withdrawal on motivated responding may depend on specific environmental, behavioral, or neuropharmacological conditions. Neuropharmacological factors contributing to increased behavioral activity during opiate withdrawal could include the amount of DA receptor stimulation. Previous studies from this laboratory have shown that D2-like DA receptor stimulation attenuates somatic signs of opiate withdrawal (Harris and Aston-Jones, 1994). DA receptor agonists also have been reported to increase escape-like jumping responses elicited by withdrawal (Schulz and Herz, 1977; Martin and Takemori, 1987; Walters et al., 2000). These findings suggest that DA receptor activation can concurrently reduce the somatic malaise associated with withdrawal and facilitate coordinated motor responses. The experiments presented here assessed the effects of stimulating D1-like and D2-like DA receptors on locomotor activity during morphine withdrawal to determine whether activating one or more of these DA receptor subtypes could indeed increase behavioral responsiveness in rats undergoing withdrawal. Our findings indicate that stimulating D2-like receptors during opiate withdrawal induces marked locomotor activity that is far greater than that displayed in the absence of withdrawal. This supersensitive response to D2-like receptor stimulation may be important for enabling drug-seeking behaviors during withdrawal from chronic opiate use.
Materials and Methods
Subjects
The subjects were male Sprague-Dawley rats (Taconic Farms Inc., Germantown, NY) weighing from 230 to 270 g. All rats were housed in pairs in a 21°C humidity-controlled Association for the Assessment and Accreditation of Laboratory Animal Care-approved animal care facility with food and water available ad libitum. The rats were acclimatized to the colony room for 5 days before the start of experimentation, and they were briefly handled on three occasions during this period. Behavioral testing was conducted during the light phase of the diurnal cycle for all but the quinpirole and eticlopride studies, which were performed during the dark phase. Our preliminary studies indicate that the diurnal cycle does not influence the ability of propylnorapomorphine HCl (NPA) to stimulate locomotor activity during opiate withdrawal. All procedures were approved by an Institutional Animal Care and Use Committee in accordance with National Institutes of Health guidelines.
Apparatus
Locomotor activity and stereotypy were measured in eight Plexiglas enclosures (40 × 40 × 40 cm; San Diego Instruments Inc., San Diego, CA), each equipped with 16 pairs of photocell sources and detectors stationed 3.5 cm above the floor and 3.5 cm apart around the perimeter. The photocell sources and detectors were connected via an interface to an IBM 486 computer with PAS software (San Diego Instruments Inc.) that recorded photocell beam interruptions. The enclosures were contained within a small, dimly lit room with white noise (70 dB) present to mask extraneous sounds.
Procedures
Induction of Dependence.
After acclimation to the colony room, the rats were anesthetized with halothane, and a 1-cm incision was made between the scapulae. A small cavity was then created under the skin and two 75-mg morphine base pellets (National Institute on Drug Abuse Drug Supply, Research Triangle Park, NC) were inserted. The incision was then closed with wound clips. This treatment has been found to elevate plasma morphine levels and induce tolerance and dependence that reaches maximal levels within 3 days, and lasts for at least 12 days after the initial implantation (Gold et al., 1994). Nondependent control rats were treated similarly except that placebo pellets instead of morphine pellets were implanted s.c. A small number of rats (approximately 5%) succumbed to overdose after implantation of the morphine pellets, resulting in unequal nvalues for some of the experiments.
Influence of Naltrexone-Precipitated Morphine Withdrawal on DA Agonist Actions.
Three separate experiments were conducted to assess the effects of naltrexone-precipitated morphine withdrawal on behavioral activity induced by DA receptor agonists. In one experiment, morphine-dependent and placebo-control animals were subdivided into groups (n = 8 or 9/group) that were tested for locomotor activity and stereotypy 3 days after pellet implantation. Five minutes before testing, half of the rats from each implant group received an i.p. injection of naltrexone (1.0 mg/kg) whereas the remaining rats were given saline. These treatments were followed 5 min later by an injection of one of four doses of the D2-like DA receptor agonist, NPA (0.05, 0.1, 0.2, or 0.4 mg/kg, i.p.) or its vehicle (0.001 N HCl). The rats were then placed immediately into the activity enclosures, where horizontal locomotion and stereotypy responses were measured for the next 60 min. Horizontal locomotion was measured continuously by a computer that recorded the number of photobeam breaks occurring during the hour-long test session. Stereotypy measures were obtained by observing each rat for 5 s every 4 min throughout the session and recording the predominant behavior displayed. The behaviors recorded included lying, standing still, grooming, locomoting, rearing, exploratory sniffing, repetitive licking, repetitive sniffing, and repetitive paw placement. The rats were considered to be engaged in stereotypy when the predominant behavior during a 5-s observation period involved repeated licking or sniffing of a surface within a restricted location, or repetitive sequences of forepaw placements along the floor or walls of the test chamber. Observational measures obtained in each experiment of this study were performed by the same individual. Although this individual was not deliberately made blind to the treatment conditions, the complexity of the counterbalanced experimental design prevented accurate tracking of the group assignments for each animal by this individual.
A second experiment examined the effects of naltrexone-precipitated morphine withdrawal on behavioral activity induced by the D2/D3 receptor agonist quinpirole. Three days after pellet implantation, morphine-dependent and placebo-control animals were subdivided into groups (n = 7 or 8/group) that received a s.c. injection of quinpirole (0.2 or 0.4 mg/kg) or vehicle (0.001 N HCl) 10 min before the test session. Ten min later, half of the rats in each dosage group were given an i.p. injection of 1.0 mg/kg naltrexone whereas the remaining animals received saline. All rats were then placed immediately into the test enclosures where horizontal locomotion and stereotypy were measured for the next 60 min.
The third experiment examined the effects of naltrexone-precipitated morphine withdrawal on behavioral activity after injections of the D1-like agonist SKF 38393. Three days after pellet implantation, morphine-dependent and placebo-control animals were subdivided into groups (n = 6/group) that first received an i.p. injection of 1.0 mg/kg naltrexone or saline. Five min later, these rats were given an i.p. injection of SKF 38393 (1.0, 2.0, 4.0, or 8.0 mg/kg) or vehicle (sterile water) and placed immediately into the test enclosures for 60 min.
Effects of a D2-Like Antagonist on Withdrawal-Induced Potentiation of NPA-Elicited Activity.
This experiment determined whether the enhanced locomotor activity produced by NPA during morphine withdrawal could be blocked by the D2-like antagonist eticlopride. Rats were implanted with morphine or placebo pellets and then tested for locomotor activity 3 days later. For this test, the rats were first injected with eticlopride (0.1 or 0.2 mg/kg, i.p.) or saline. All rats were then given an injection of NPA (0.2 mg/kg, i.p.) 25 min later, and these injections were followed 5 min thereafter by an i.p. injection of 1.0 mg/kg naltrexone or saline. The rats (n = 7 or 8/group) were then placed immediately into the test enclosures where horizontal locomotion was measured for 60 min.
DA Agonist Actions during Withdrawal Induced by Cessation of Morphine Treatment.
The rats in this experiment received s.c. implants of either morphine or placebo pellets as described above. Three days later, the rats were briefly anesthetized and the pellets were extracted to initiate withdrawal. Different groups of rats were then tested 1, 2, 4, or 8 days after removal of the pellets to assess the effects of NPA on locomotion at different stages of morphine withdrawal. On each test day, two groups of morphine-dependent rats and two placebo-control groups (n = 7 or 8/group) were weighed, and the rats were placed individually into clear plastic cages (45 × 24 × 20 cm), where they were observed continuously over a 30-min period for somatic signs of opiate withdrawal. Each occurrence of wet dog shakes, teeth chatter, writhing, eye twitching, jumping, and diarrhea during this period was recorded. In addition, the rats were checked every 10 min for evidence of ptosis, rhinorrhea, lacrimation, and vocalization on touch. The rats were then injected with either 0.2 mg/kg NPA or vehicle (0.001 N HCl) and placed immediately into the activity enclosures, where locomotor activity and stereotypy were measured for 60 min.
Effects of Acute Morphine Antagonism on DA Agonist Actions.
A final experiment measured locomotor activity in rats (n = 8/group) when NPA was coadministered with naltrexone after acute exposure to morphine. Drug-naive rats were given an i.p. injection of 10 mg/kg morphine or saline. Each animal was subsequently given either 1.0 mg/kg naltrexone or saline 25 min later. The rats then received 0.2 mg/kg NPA or its vehicle 5 min thereafter, and they were placed immediately into the activity enclosures, where locomotor activity and stereotypy were recorded for 60 min.
Drugs
Each morphine pellet contained 75 mg of morphine alkaloid base, 68.5 mg of microcrystalline cellulose, 1.5 mg of magnesium stearate, and 2.5 mg of colloidal silicon dioxide. Placebo pellets were similarly formulated but contained no morphine. R(−)-NPA and (−)-quinpirole HCl were dissolved in 0.001 N HCl. Naltrexone HCl, morphine sulfate, and S(−)-eticlopride HCl were dissolved in 0.9% sodium chloride solution, and (±)-SKF 38393 HCl was dissolved in distilled water. The DA agonists and naltrexone were administered 0 or 5 min before the start of testing so that they would reach maximum effectiveness during the course of the test sessions. The eticlopride and acute morphine injections were given 30 min before the start of the sessions so that they would reach peak effectiveness before the onset of the DA agonist and naltrexone effects. Morphine and placebo pellets were implanted s.c. (2 pellets/rat), and all other compounds were injected as solutions in a volume of 1 ml/kg. Morphine pellets, placebo pellets, and morphine sulfate were generously donated by the National Institute on Drug Abuse Drug Supply (Research Triangle Park, NC), whereas all other compounds were purchased from Research Biochemicals International (Natick, MA).
Statistical Analyses
The measure of locomotor activity used for all analyses was the total number of activity counts (i.e., photobeam breaks) recorded within a 60-min session. The measure of stereotypy used was the number of 5-s observation periods in which stereotyped responses (i.e., repetitive licking, repetitive sniffing, or repetitive paw placement) were recorded. Locomotor and stereotypy scores from placebo- and morphine-implanted rats in the precipitated withdrawal experiments were analyzed separately using two-way between groups ANOVAs with antagonist treatment (i.e., naltrexone or saline), and DA agonist or antagonist dose as factors. Similar two-way ANOVAs were used to analyze locomotor activity in placebo- and morphine-implanted rats after pellet removal, with NPA dose and test day serving as factors. Somatic withdrawal signs and body weights measured on each test day after pellet removal also were subjected to two-way ANOVAs, with pellet implant and test day serving as factors. The data for groups treated with NPA or its vehicle were combined for these analyses, as somatic signs and body weights were recorded before drug treatments on each test day. Locomotor activity scores measured in saline- and morphine-treated rats in the acute antagonism experiment were analyzed using two-way between groups ANOVAs with antagonist injection and NPA dose serving as factors. Significant effects obtained from the ANOVAs were subjected to further post hoc analysis using Fisher's least significant difference (LSD) test. Differences were considered significant at P < .05.
Results
Influence of Naltrexone-Precipitated Morphine Withdrawal on DA Agonist Actions.
Figure 1 shows the effects of the D2-like agonists NPA and quinpirole, and the D1-like agonist SKF 38393, on locomotor activity in morphine-dependent and nondependent rats given naltrexone or saline. These experiments revealed that the D2-like agonists NPA (0.1, 0.2, and 0.4 mg/kg) and quinpirole (0.2 and 0.4 mg/kg) markedly increased locomotion in dependent rats undergoing naltrexone-precipitated withdrawal (Fig. 1, A and B). This effect was selective for the withdrawal state, as the D2-like agonists produced either no effect (NPA) or only a moderate increase (quinpirole) in locomotor activity when given to nondependent rats in combination with either saline or naltrexone, and they significantly inhibited locomotor activity when given to morphine-dependent rats in the absence of naltrexone. In contrast to these effects of D2-like agonists, treatments with the D1-like agonist SKF 38393 did not alter locomotor activity relative to vehicle-treated controls under any of the experimental conditions examined (Fig. 1C).

Effects of NPA (A), quinpirole (B), and SKF 38393 (C) on locomotor activity in nondependent (PLA) and opiate-dependent (MOR) rats given saline (SAL) or naltrexone (NTX) before a 60-min test session. Asterisks indicate groups in which locomotor activity differed significantly from levels observed in the respective vehicle control groups (Fisher's LSD test, P < .05;n = 6 to 9/group).
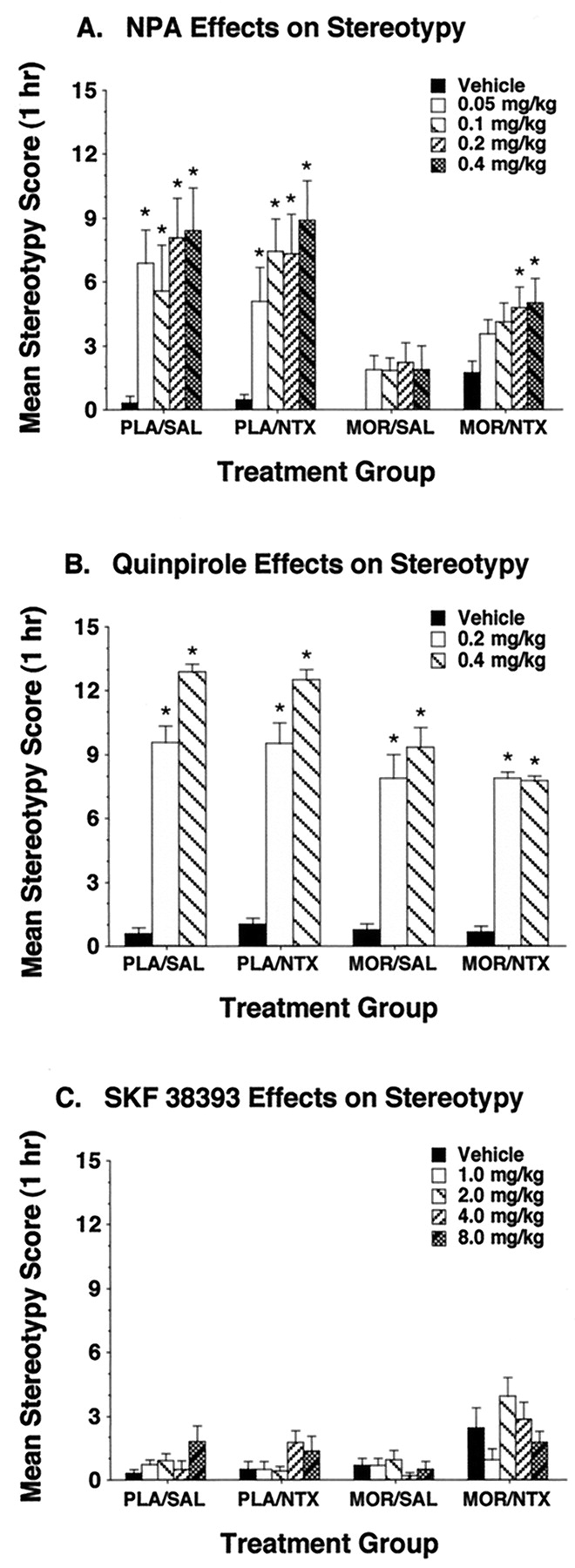
Stereotypy measures obtained during each drug test showed a somewhat different pattern of variation across experimental conditions than was observed with locomotor activity, indicating that the effects of withdrawal on locomotion were dissociable from concurrent changes in stereotypy. Thus, NPA consistently induced high levels of stereotypy when given to nondependent rats that were treated with either naltrexone or saline before testing (Fig.2A). This stereotypy was absent in morphine-dependent rats that were not subjected to withdrawal, and they were partially reinstated in a dose-dependent manner in dependent rats undergoing precipitated withdrawal. Quinpirole also induced strong stereotypy in nondependent rats given either saline or naltrexone pretreatments (Fig. 2B). However, this stereotypy was only slightly attenuated in morphine-dependent rats, and the levels of this behavior were not altered by the induction of withdrawal. Injections of SKF 38393 did not induce stereotypy under any of the experimental conditions in this study (Fig. 2C).
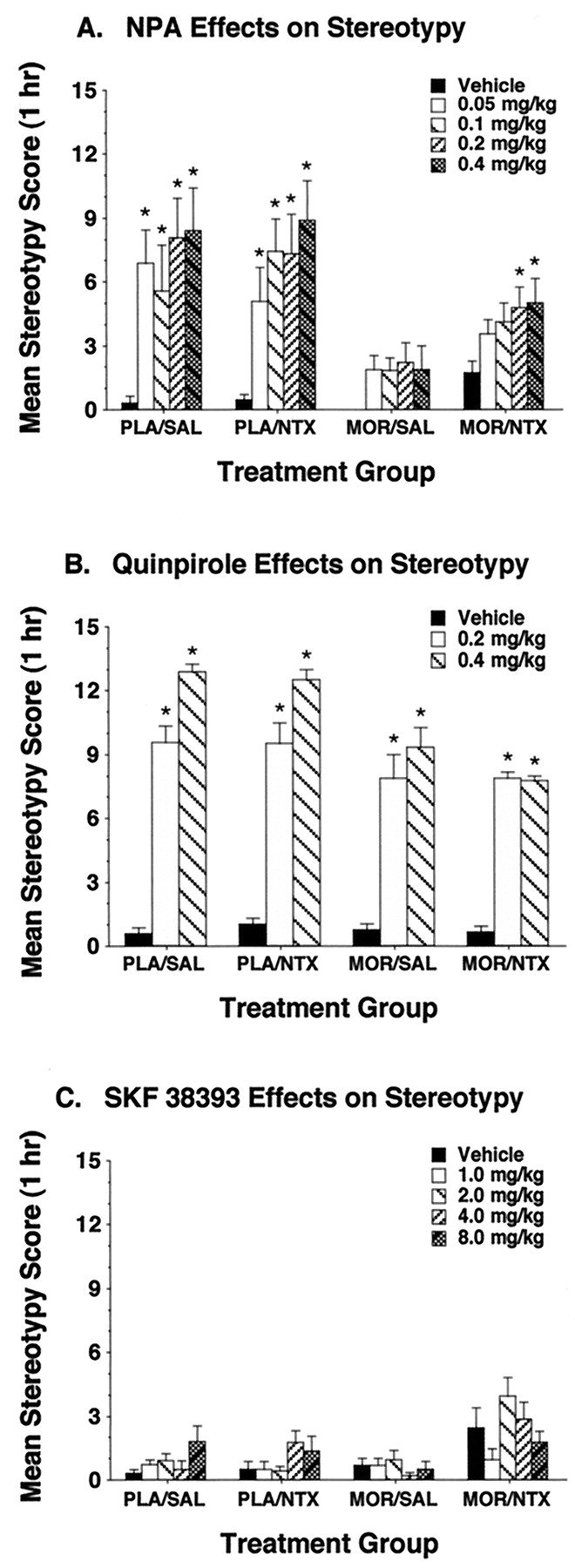
Effects of NPA (A), quinpirole (B), and SKF 38393 (C) on stereotypy in nondependent (PLA) and opiate-dependent (MOR) rats given saline (SAL) or naltrexone (NTX) before a 60-min test session. Asterisks indicate groups in which stereotypy differed significantly from levels observed in the respective vehicle control groups (Fisher's LSD test, P < .05;n = 6 to 9/group).
Although the rats in these experiments were tested in a novel environment, the novelty of the test situation was not a critical factor influencing the effects of opiate withdrawal on the locomotor effects of the D2-like agonists. This was confirmed in an additional experiment, wherein morphine-dependent rats given four prior exposures to the test environment displayed significant hyperactivity (3536 counts in 1 h; n = 5) during tests with 0.2 mg/kg NPA and naltrexone relative to activity measured during habituation sessions (mean counts/1-h session = 1734, 1167, 934, and 824 for sessions 1–4). The magnitude of this hyperactivity response was similar to that produced by this dose of NPA in morphine-withdrawn rats in Fig. 1A. Other findings from our laboratory further indicate that the locomotor activating effects of NPA during morphine withdrawal do not decline when rats are tested repeatedly in the same environment (J. P. Druhan, C. L. Walters, and G. Aston-Jones, unpublished observations).
Effects of D2-Like Antagonists on Withdrawal-Induced Potentiation of NPA-Elicited Activity.
Figure3 shows the effects of the D2-like antagonist eticlopride on locomotor activity in NPA-treated rats subjected to naltrexone-precipitated withdrawal or control conditions. Consistent with the results described above, NPA produced a strong increase in locomotor activity when given to morphine-dependent rats undergoing naltrexone-precipitated withdrawal in the absence of eticlopride. This locomotor-activating effect of NPA was blocked in a dose-dependent manner by injections of eticlopride. Eticlopride also reduced locomotion in morphine-dependent rats that were not subjected to withdrawal, but it did not significantly affect locomotor activity in any of the nondependent groups.

Effects of the D2-like antagonist eticlopride on NPA-induced locomotor activity during withdrawal. All of the rats in this experiment received an injection of 0.2 mg/kg NPA before testing. Asterisks indicate groups in which locomotor activity differed significantly from levels observed in the respective vehicle control groups (Fisher's LSD test, P < .05;n = 7 or 8/group).
DA Agonist Actions during Withdrawal Induced by Cessation of Morphine Treatment.
Removal of the morphine pellets after dependence induction resulted in body weight reductions and spontaneous somatic signs of withdrawal that were evident on days 1, 2, and 4 after pellet removal (Table 1). Rats subjected to this morphine withdrawal procedure also showed a progressive increase in locomotor responses to NPA, with increases reaching statistical significance on the 4th day after pellet removal (Fig.4A). The ability of NPA to stimulate locomotor activity then declined by the 8th day after removal, so that the increased locomotion was no longer significant. Importantly, locomotor activity levels of nondependent rats that had placebo pellets implanted and then removed were not affected by NPA (Fig. 4B), indicating that the stimulant effects of NPA in dependent rats were related to the cessation of prolonged morphine exposure.
Evidence of spontaneous withdrawal after pellet removal in morphine-dependent rats

Effects of NPA or vehicle injections on locomotor activity in dependent (A) and nondependent (B) rats tested 1, 2, 4, or 8 days after pellet removal. Asterisks indicate groups in which locomotor activity differed significantly from levels observed in the respective vehicle control group (Fisher's LSD test,P < .05; n = 7 or 8/group).
As with naltrexone-precipitated withdrawal, the increased activation produced with NPA could not be attributed to decreased stereotypy. Although stereotypy levels were initially reduced 1 day after pellet removal (consistent with the desensitized response of morphine-dependent rats in Fig. 1B), this behavioral response increased steadily during the 4-day period when sensitized locomotor responses to NPA were emerging (data not shown).
Effects of Acute Morphine Antagonism on DA Agonist Actions.
Figure 5 shows the effects of NPA on locomotor activity in rats given naltrexone or saline after an acute injection of 10.0 mg/kg morphine or its vehicle. NPA did not affect locomotor activity in control animals that received neither morphine nor naltrexone, and it decreased activity when given either with naltrexone in the absence of morphine, or with morphine in the absence of naltrexone. Importantly, NPA failed to stimulate activity when given in combination with acute morphine and naltrexone, indicating that the ability of NPA to increase locomotion during periods of reduced opiate receptor stimulation depended on prolonged exposure to morphine.
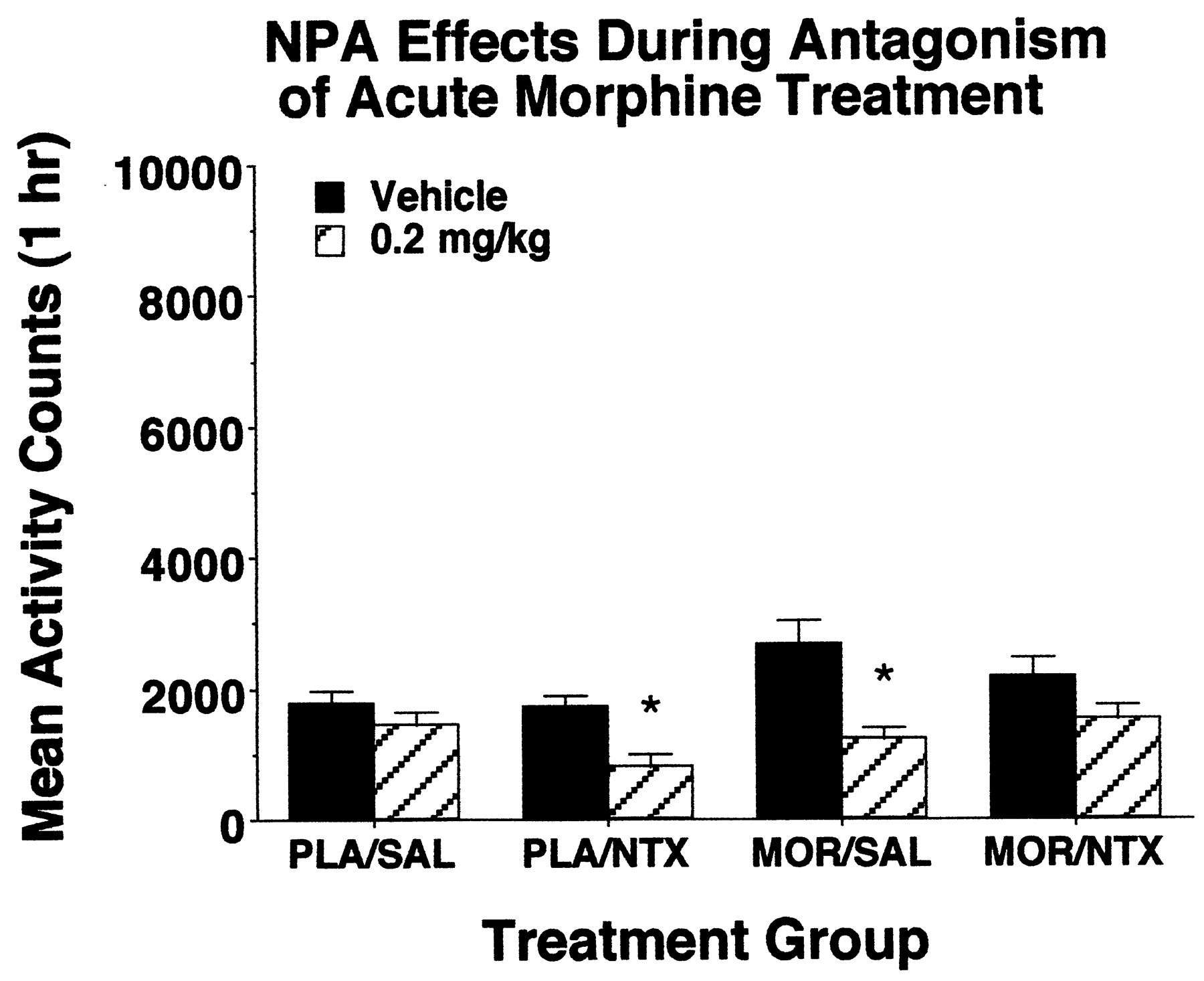
Effects of NPA or vehicle injections on locomotor activity in nondependent rats that received acute injections of saline or 10.0 mg/kg morphine followed 25 min later by saline or 1.0 mg/kg naltrexone. Asterisks indicate groups in which locomotor activity differed significantly from levels observed in the respective vehicle control groups (Fisher's LSD test, P < .05;n = 8/group).
Discussion
In this study, doses of the D2-like agonists NPA or quinpirole that had little or no effect on locomotor activity in nondependent rats greatly enhanced locomotion in morphine-dependent rats undergoing naltrexone-precipitated withdrawal. These findings indicate that opiate withdrawal potentiates locomotor activating effects produced by D2-like receptor stimulation. Importantly, enhanced locomotor responses to NPA were observed when withdrawal was induced by removal of the morphine pellets. This result, and the lack of potentiated responses to NPA after naltrexone antagonism of an acute morphine injection, indicated that the enhanced locomotor response during naltrexone-precipitated withdrawal was not due to pharmacokinetic interactions between the D2-like agonists, morphine, and naltrexone. Rather, processes initiated by opiate withdrawal appeared to alter the functional impact of D2-like receptor stimulation to produce more behavioral activation than occurred in the absence of withdrawal. This potentiating effect of withdrawal was selective for stimulation of D2-like DA receptors, as it was not observed after injections of the D1-like agonist SKF 38393.
The enhanced locomotor response to NPA during withdrawal was blocked by the D2-like receptor antagonist eticlopride at doses that did not affect locomotion in nondependent rats given NPA. This result confirmed the importance of D2-like receptor stimulation in mediating the activity-inducing effects of NPA during withdrawal. Eticlopride also reduced locomotor activity in morphine-dependent rats given NPA in the absence of withdrawal, suggesting that these rats were more susceptible to the motor suppressant effects of D2-like antagonists. However, the pattern and magnitude of dose-response effects observed in the presence and absence of withdrawal differed, suggesting that the blockade of NPA-induced locomotion during withdrawal was not due to nonspecific inhibitory effects of eticlopride on behavior. It is more likely that eticlopride prevented the stimulant effects of NPA by blocking the actions of this agonist at D2-like receptor sites, although future studies using additional DA antagonists and increased dose ranges are necessary to confirm this conclusion.
NPA and quinpirole inhibited locomotor activity when given to dependent rats in the absence of withdrawal. This effect depended on concurrent opioid receptor stimulation, as it did not occur when NPA or quinpirole were administered to placebo-implanted rats. However, the inhibitory effects were not dependent on adaptations induced by chronic morphine because they could be observed after acute morphine injections. Although the specific neuropharmacological mechanism underlying these effects could not be discerned, they likely involved the stimulation of D2-like autoreceptors. These autoreceptors are sensitive to low doses of DA agonists, and the decreased neuronal firing and DA release produced by stimulation of these receptors would be expected to reduce general activity. Surprisingly, the locomotor inhibition produced by NPA in morphine-dependent rats was not reversed by eticlopride. Instead, the D2-like antagonist produced further reductions in activity. This effect of eticlopride may have resulted from a blockade of postsynaptic D2-like receptors at the doses used. Like autoreceptor stimulation, postsynaptic receptor blockade interferes with DA transmission in forebrain regions. If morphine-treated animals were more susceptible to response decrements produced by decreased DA transmission, then reductions in locomotor activity would be predicted regardless of whether transmission was decreased by stimulation of autoreceptors or by antagonism of postsynaptic D2-like receptors.
At high doses, DA agonists elicit repetitive stereotyped movements that interfere with generalized activity and forward locomotion. Accordingly, one might wonder whether the decreased activity induced by NPA and quinpirole in dependent, nonwithdrawn rats was due to increased stereotypy, and whether the hyperactivity during opiate withdrawal reflected a reversal of this stereotypy and consequent unmasking of the locomotor stimulant effects of the D2-like agonists. This explanation is not tenable, however, as stereotypy levels were actually lower than normal when dependent rats were given NPA or quinpirole in the absence of withdrawal. Moreover, stereotypy induced by NPA increased relative to these reduced levels during withdrawal, and stereotypy produced by quinpirole was not affected. It is further noteworthy that opiate withdrawal can potentiate other D2 agonist-induced responses that are less affected by changes in stereotypy, such as penile erection and yawning (Reddy et al., 1993; Piepponen et al., 1996).
NPA did not stimulate locomotor activity after antagonism of an acute morphine injection, indicating that the enhanced locomotor response to D2-like agonists during withdrawal depended on prolonged exposure to morphine. This suggests that sustained opioid receptor stimulation induces functional adaptations that alter the response of central nervous system systems to concurrent changes in activity at opioid and D2-like receptors. These adaptations probably do not involve an up-regulation of D2 receptors by chronic morphine, as these receptors are often desensitized or unchanged after repeated morphine treatments (Navarro et al., 1992; Reddy et al., 1993; Nestby et al., 1995). The adaptations also do not appear to be related to the long-term behavioral sensitization observed in rats after intermittent injections of opiate or psychomotor stimulant compounds. This latter form of sensitization can be detected up to 1 year after the suspension of chronic drug treatments, whereas the potentiation of D2-like agonist effects by opiate withdrawal was transient and no longer significant 8 days after pellet removal (more recent experiments indicate that the potentiation is completely absent 21 days after pellet removal). There is, however, another form of pharmacologically induced sensitization that may involve mechanisms similar to those underlying the potentiation of D2-like agonist effects during withdrawal. Locomotor activity elicited by infusions of μ-opioid agonists into the nucleus accumbens (NAc) is markedly potentiated after neurotoxic lesions of the mesolimbic DA system or chronic treatments with D2-like or mixed D1-like/D2-like DA receptor antagonists (Kalivas and Bronson, 1985; Stinus et al., 1986). It appears that opioid and D2-like receptor-regulated processes are closely linked such that chronic changes in one system can profoundly affect the functioning of the other.
Recent studies in our laboratory indicate that enhanced locomotor responses to D2-like agonists during opiate withdrawal depend on interactions between opioid and D2-like receptor systems within the NAc (Walters et al., 1998). However, it is unlikely that these interactions involve a joint regulation of common intracellular transduction mechanisms, as opiate withdrawal produces effects on cellular activity and signal transduction that are opposite in direction from those induced by D2-like receptor activation. For example, opiate withdrawal increases cAMP production, neuronal activity, and Fos-related antigen expression within the NAc (Duman et al., 1988;Terwilliger et al., 1991; Rasmussen et al., 1995; Walters et al., 2000), whereas stimulation of D2-like receptors inhibits these processes within the NAc (White and Wang, 1986;Schoffelmeer et al., 1987; Walters et al., 2000). Instead, opiate withdrawal and D2-like agonists may act on different populations of NAc cells that have opposing influences on “downstream” processes. In situ hybridization studies indicate that μ-opioid receptors are localized on cells that project preferentially to the ventral tegmental area, whereas D2-like receptors are found on cells that project almost exclusively to the ventral pallidum and influence ventral tegmental neurons only indirectly by way of their connections with pallido-tegmental projections (Lu et al., 1998; Georges et al., 1999). This differential localization of μ-opioid and D2-like receptors within direct and indirect pathways is similar to the situation described for the descending striatonigral pathways, wherein μ-regulated direct and D2-regulated indirect projections are believed to have opposite influences on the functional output of the basal ganglia (Steiner and Gerfen, 1998). If a similar situation holds true for accumbens efferents, then opposing effects of opiate withdrawal and D2-like receptor stimulation on neuronal activity within the NAc could translate into synergistic actions within the ventral tegmentum.
The finding that opiate withdrawal potentiates behavioral activity produced by D2-like receptor activation has important implications for current models of how opiate withdrawal stimulates drug-seeking behavior. It is generally believed that opiate withdrawal promotes drug seeking by inducing an aversive subjective state that addicts are motivated to alleviate through continued drug use. This aversive state is dependent on increased noradrenaline release within the bed nucleus of the stria terminalis (Delfs et al., 2000). The present results suggest that opiate withdrawal might also facilitate drug seeking by potentiating behavioral responses to D2-like receptor stimulation. Although DA levels within the NAc are reduced during opiate withdrawal, the enhanced sensitivity to D2-like receptor stimulation could result in augmented behavioral responses to stressors or drug-related stimuli that increase DA release (Abercrombie et al., 1989; Kalivas and Duffy, 1995; DiCiano et al., 1998). This role for altered responsiveness to D2-like receptor stimulation in drug seeking would be consistent with recent findings showing that quinpirole reinstates extinguished heroin self-administration responses when given 4 days after access to heroin reinforcement has been terminated, but not 21 days after termination of reinforcement when dependence would have subsided (De Vries et al., 1999). It is also noteworthy that stimulation of accumbens D2-like receptors attenuates somatic signs of opiate withdrawal (Harris and Aston-Jones, 1994). This reduction in somatic malaise could also contribute to the facilitation of drug-seeking responses by decreasing impediments to behavioral responding imposed by the withdrawal state. An important goal for future research on opiate abuse will be to determine how these dopaminergic actions interact with other aspects of withdrawal to regulate drug-seeking responses during periods of abstinence.
Acknowledgment
We thank Dr. Jill Delfs for helpful comments on the manuscript.
Footnotes
-
Send reprint requests to: Dr. J.P. Druhan, Veterans Administration Medical Center, Mail Code 151, University and Woodland Aves., Philadelphia, PA 19104. E-mail:druhan{at}mail.med.upenn.edu
-
↵1 This research was supported by Public Health Service Grant DA10088 from the National Institute on Drug Abuse.
- Abbreviations:
- DA
- dopamine
- NAc
- nucleus accumbens
- NPA
- propylnorapomorphine HCl
- LSD
- least significant difference
- Received February 15, 2000.
- Accepted April 20, 2000.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}