


Abstract
The present study investigated the effects of the novel corticotrophin-releasing factor (CRF)1 receptor antagonist 4-(2-chloro-4-methoxy-5-methylphenyl)-N-[(1S)-2-cyclopropyl-1-(3-fluoro-4-methylphenyl)ethyl]5-methyl-N-(2-propynyl)-1,3-thiazol-2-amine hydrochloride (SSR125543A) in a variety of rodent models of anxiety, including conflict procedures (punished drinking and four-plate), exploration models (elevated plus-maze and light/dark), a fear/anxiety defense test battery, and several procedures based on stress-induced changes in physiological (isolation-induced hyperthermia and tail pinch-induced cortical norepinephrine release) or behavioral (social defeat-induced anxiety, maternal separation-induced vocalization) parameters. Moreover, the effects of SSR125543A were investigated in acute (forced swimming) and chronic (chronic mild stress; CMS) models of depression. SSR125543A and the CRF1receptor antagonist antalarmin displayed limited efficacy in exploration-based anxiety models. In contrast, both compounds produced clear-cut anxiolytic-like activity in models involving inescapable stress, including the conflict procedures, the social defeat-induced anxiety paradigm and the defense test battery (3–30 mg/kg i.p. or p.o.). These effects paralleled those of the anxiolytic diazepam. In addition, SSR125543A and antalarmin antagonized stress-induced hyperthermia, distress vocalization, and cortical norepinephrine release. In the forced swimming test, 30 mg/kg p.o. SSR125543A and 3 to 30 mg/kg p.o. antalarmin produced clear antidepressant-like effects. These latter results were strengthened by the findings from the CMS, which showed that repeated administration of 10 mg/kg i.p. SSR125543A for 30 days improved the degradation of the physical state, the reduction of body weight gain, and anxiety produced by stress. Together, these data indicate that SSR125543A shows good activity in acute and chronic tests of unavoidable stress exposure, suggesting that it may have a potential in the treatment of depression and some forms of anxiety disorders.
Corticotropin-releasing factor (CRF) has been identified as a neuropeptide that plays a central role in the coordination of neuroendocrine, autonomic, and behavioral responses to stress (Vale et al., 1981). It is the main regulator of basal and stress-induced release of the adrenocorticotropic hormone (ACTH) (Stout et al., 1995). Two CRF receptor subtypes, CRF1 and CRF2, with distinct anatomical localization and pharmacology have been identified (Chalmers et al., 1996). CRF1 receptor expression is most abundant in neocortical, cerebellar, and limbic structures, whereas CRF2 receptor expression is generally prominent in subcortical structures. This anatomical information provided a basis for functional hypotheses related to CRF receptor subtypes and suggested that CRF may contribute significantly both to behavioral responses to stress and emotional behavior itself. This idea has been substantiated by numerous studies showing that i.c.v. application of CRF in rodents produces behavioral effects similar to those observed when animals are exposed to stress (for review, seeGriebel, 1999). Studies using CRF transgenic mouse lines overexpressing CRF or knockout mice lacking the CRF1 receptor further emphasized the involvement of CRF in stress-related behaviors, because the former exhibited a behavioral state resembling that produced by anxiety, whereas the latter showed reduced emotionality (for review, see Contarino et al., 1999). On a clinical level, a large body of evidence points to increased cerebrospinal fluid CRF concentrations in drug-free patients with major depression and post-traumatic stress disorder compared with healthy controls (for review, see Kasckow et al., 2001). In this context, it was hypothesized that CRF receptor antagonists may represent novel agents for the treatment of stress-related disorders (Chalmers et al., 1996; Holsboer, 1999).
Several classes of nonpeptide antagonists of CRF1receptors have been identified (for review, see Gutman et al., 2000). A few of them have been studied extensively in experimental models of stress. For example, peripheral administration of the pyrrolopyrimidine derivatives antalarmin, CP-154,526, and R121919 was reported to reduce the effects of acute and repeated stressors on behavior in rodents, and some activity was found in classical models used to screen anxiolytics and antidepressants (for reviews, see Griebel, 1999; Gutman et al., 2000). However, negative results were also reported with some of these procedures. It was suggested that CRF1antagonists may produce positive effects only when the endogenous tone of CRF is high, thereby pointing to a crucial importance of baseline levels of stress when investigating the behavioral actions of these compounds. Recently, the results of the first open-label study examining the effects of R121919 in 20 patients with major depression was published (Zobel et al., 2000). The compound was well tolerated by the patients and did not significantly affect ACTH or cortisol levels at baseline or after a CRF challenge. More importantly, significant reductions in depression and anxiety scores were observed after 30-day treatment with the compound. Although this small open-label study does not provide unequivocal proof, it brings further evidence that a selective CRF1 receptor antagonist may represent a promising alternative to agents currently used for the treatment of anxiety and depressive disorders.
In the present article, we report on the psychopharmacological profile of 4-(2-chloro-4-methoxy-5-methylphenyl)-N-[(1S)-2-cyclopropyl-1-(3-fluoro-4-methylphenyl)ethyl]5-methyl-N-(2-propynyl)-1,3-thiazol-2-amine hydrochloride (SSR125543A), a 2-aminothiazole derivative, which displays high affinities for both native and recombinant human CRF1 receptors (Ki = 1 and 2 nM, respectively), and 1000-fold selectivity for CRF1 compared with CRF2 receptor (Gully et al., 2002). SSR125543A is a potent antagonist at the CRF1 receptor as shown by its ability to inhibit CRF-induced cyclic AMP synthesis in human retinoblastoma Y 79 cells (pA2 = 8.92) and ACTH secretion in mouse pituitary tumor AtT-20 cells (pA2 = 9.63). After in vivo experiments in rats, the compound was found to inhibit CRF- and restraint-stress induced ACTH secretion when given orally. In the present series of experiments, the behavioral effects of SSR125543A were examined using a variety of rodent models of anxiety, including conflict procedures (punished drinking and four-plate), exploration models (elevated plus-maze and light/dark), a fear/anxiety defense test battery, and several test procedures based on stress-induced changes in physiological (isolation-induced hyperthermia and tail pinch-induced cortical norepinephrine release) or behavioral (social defeat-induced anxiety, maternal separation-induced distress vocalizations) parameters. Moreover, the effects of SSR125543A were investigated in acute (forced swimming) and chronic (chronic mild stress; CMS) models used for the characterization of antidepressants. In this latter experiment, the compound was given repeatedly for 30 days. Comparative data for the CRF1 receptor antagonist antalarmin and the anxiolytic and antidepressant diazepam and fluoxetine, respectively, obtained under the same experimental conditions, are also provided. Finally, possible unwanted effects of increasing doses of SSR125543A were examined using motor activity and memory tests.
Materials and Methods
Ethics
All experimental procedures described herein were approved by the Animal Care and Use Committee of Sanofi-Synthelabo Recherche and fully comply with French legislation on research involving animal subjects.
Animals
Male Sprague-Dawley and Wistar rats (Iffa Credo, L'Arbresle, and Charles River, Saint-Aubin-lès-Elbeuf, France), weighing 180 to 330 g at the time of testing, were used in the punished drinking, elevated plus-maze, stress-induced hyperthermia, and forced swimming tests, and in the microdialysis experiment. They were housed in groups of three to eight. Different strains of rats were used to optimize conditions. For example, preliminary data from our laboratory have shown that, unlike Sprague-Dawley rats, Wistar rats from our laboratory animal supplier are poor responders in the elevated plus-maze (i.e., they display weak avoidance responses of the open aversive arms), making it difficult to use them for the screening of anxiolytics. Male CD1 (social defeat stress-induced anxiety, horizontal wire test, rotarod, passive avoidance test, and actimeter), OF1 (mouse defense test battery; MDTB), NMRI (four-plate test), and BALB/c (light/dark test, chronic mild stress procedure) mice weighing 17 to 32 g were supplied by Charles River, Iffa Credo, or Janvier (Le Genest, France). CD1, OF1, and NMRI mice were housed in groups of 20, those used in the MDTB and in the chronic mild stress procedure were housed singly, and BALB/c mice used in the light/dark test were housed in groups of six. Female guinea pigs (Dunkin-Hartley) with four 5-day-old pups were obtained from Harlan (Horst, The Netherlands). Each mother was housed individually with her litter provided with sawdust. Moreover, male Long Evans rats (400–500 g) (Iffa-Credo) were used as threat stimulus in the MDTB. All animals were maintained under standard laboratory conditions (21–23°C; 40–60%relative humidity) and kept on a 12-h light/dark cycle with light onset at 6:00 AM.
Compounds
Compounds were prepared as solutions or suspensions in physiological saline or distilled water containing 0.1% Tween 80 (antalarmin, diazepam, and fluoxetine) or 5% dimethyl sulfoxide and 5% Cremophor EL (SSR125543A). The compounds used were SSR125543A, antalarmin, diazepam, and fluoxetine (synthesized by Sanofi-Synthelabo, Bagneux and Toulouse, France). Compounds administered i.p. and p.o. were given in a constant volume of 5 ml/kg (rats), 2 ml/kg (guinea pig pups), or 20 (mice) ml/kg. The i.p. route was used in some experiments because an oral administration would have increased further the level of stress [i.e., distress vocalizations in guinea pig pups, stress-induced norepinephrine (NE) release in rats, and chronic mild stress in mice] or because it would have interfered with the procedure (i.e., water deprivation in the punished drinking test in rats).
Anxiolytic-Like Activity of SSR125543A
Punished Drinking Test in Rats.
The procedure was a modification of the technique described by Vogel et al. (1971). At the beginning of the experiment, rats, deprived of water but not of food for 48 h before testing, were placed in cages (32 × 25 × 30 cm) with a stainless steel grid floor. Each cage was placed in sound-attenuated boxes that were well ventilated and contained a drinking tube connected to an external 50-ml burette filled with tap water. Trials were started only after the animal's tongue entered in contact with the drinking tube for the first time. An electric shock (0.6 mA/500 ms) was delivered to the tongue after every 20 licks. The number of shocks was recorded automatically during a 5-min period. Data were analyzed with one-way analysis of variance (ANOVA). Subsequent comparisons between treatment groups and control were carried out using Dunnett's t test. Experiments were performed 30 min (antalarmin and diazepam) or 60 min (SSR125543A) after i.p. injection of the compounds. A dose of 2.5 mg/kg i.p. morphine was tested as negative control because this test involves the application of painful stimuli.
Elevated Plus-Maze Test in Rats.
The test apparatus is based on that described by Pellow et al. (1985). All parts of the apparatus were made of dark polyvinylplastic with a black rubber floor. The maze was elevated to a height of 50 cm with two open (50 × 10 cm) and two enclosed arms (50 × 10 × 50 cm), arranged so that the arms of the same type were opposite each other, connected by an open central area (10 × 10 cm). To prevent rats falling off, a rim of Plexiglas (0.5 cm in height) surrounded the perimeter of the open arms. The illumination in the experimental room consisted of one red neon tube fixed on the ceiling, so that experiments were performed under dim light conditions. The light intensity on the central platform was 10 lux. At the beginning of the experiment, rats were placed in the center of the maze, facing one of the enclosed arms, and observed for 4 min. The apparatus was equipped with infrared beams and sensors capable of measuring time spent in open arms, number of open-arm entries, and number of closed-arm entries (defined as entry of all four limbs into an arm of the maze). In addition, rats were observed via video-link by an observer located in an adjacent room. This allowed the recording of a more ethologically orientated measure: 1) attempt: attempt at entry into open arms followed by avoidance responses. This includes stretched attend posture (the rat stretches forward and retracts to original position); and 2) head-dipping: protruding the head over the ledge of an open arm and down toward the floor (this response can occur while the animal's body is in the closed arms, central square, or on open arms). The results were expressed as mean ratio of time spent in open arms to total time spent in both open and closed arms, mean ratio of entries into open arms to total entries into both open and closed arms, mean total number of both closed and open arm entries, mean total number of attempts, and mean total number of head-dips. Data were analyzed with one-way ANOVA. Subsequent comparisons between treatment groups and control were carried out using Dunnett's t test. Experiments were performed 60 min after p.o. administration of SSR125543A, antalarmin, or diazepam.
Stress-Induced Hyperthermia in Rats.
The basal rectal temperature of grouped rats (5–7/cage) was measured with a telethermometer (DM 852; ELLAB Instruments, Copenhagen, Denmark). Animals were then removed from their home cage, and placed individually in a small transparent plastic cage (25 × 15 × 27 cm). Temperature was measured again three times at 15-min intervals. Isolation yielded an enhanced body temperature that putatively reflects a stress-induced anxiogenic response. SSR125543A, diazepam, or antalarmin were administered p.o. or i.p. 60-min before basal temperature measurement. Data were analyzed by two-way ANOVA (time × treatment) with repeated measures, followed by Dunnett'st test.
Stress-Induced Cortical Norepinephrine Release in Rats.
Cortical extracellular NE levels were increased after a 15-min tail pinch in awake rats (Funk and Stewart, 1996). They were measured in 30-μl dialysate samples by a high-performance liquid chromatography system with coulometric detection as previously described (Marco et al., 1998), except for the mobile phase, which contained 7% acetonitryl as organic agent. The analytical system consisted of an electrochemical detector ESA Coulochem II equipped with a model 5014 analytical cell (ESA, Chelmsford, MA). The NE levels in fractional samples were converted to a percentage of the mean value of the 45-min baseline measurements before treatment. Time course effects of tail pinch on NE levels were analyzed by ANOVA with repeated measures, followed by Dunnett's t test analysis. SSR125543A and antalarmin were administered i.p. 30 min before tail pinch. Compound antagonism was evaluated during the tail pinch sampling collection. Statistical analysis was carried out using ANOVA followed by Dunnett'st test.
Light/Dark Test in Mice.
The test apparatus is based on that described by Misslin et al. (1989). It consisted of two polyvinylchloride boxes (20 × 20 × 14 cm) covered with Plexiglas. One of these boxes was darkened. A desk lamp placed 20 cm above the lit box and a neon tube fixed on the ceiling provided the room illumination so that the light intensity in the center of the illuminated box was 1000 lux. An opaque plastic tunnel (5 × 7 × 10 cm) separated the dark box from the illuminated one. At the beginning of the experiment, a mouse was placed in the illuminated box, facing the tunnel. Recording started when the animal entered the tunnel for the first time. The apparatus was equipped with infrared beams and sensors capable of recording the following parameters during a 4-min period: 1) time spent by mice in the lit box; 2) attempt at entry into the lit box followed by avoidance responses (this includes stretched attend posture; the mouse stretches forward and retracts to original position); 3) total number of tunnel crossings; and 4) activity in the lit box. Data were analyzed with one-way ANOVA followed by Dunnett's t test analysis. Experiments were performed 30 min (antalarmin and diazepam) or 60 (SSR125543A and diazepam) min after p.o. administration of the compounds.
Four-Plate Test in Mice.
The test apparatus is based on that described by Boissier et al. (1968). The apparatus consisted of a cage with a floor composed of four rectangular metal plates connected to a device that can generate electric shocks (1 mA, 0.2 s). After a 15-s latency period, the animal is subjected to an electric shock every time it went from one plate to another. The number of punished crossings is recorded during a 1-min test period. Experiments were carried out 60 min after p.o. administration of SSR125543A, antalarmin, or diazepam. In a second experiment, the duration of the anxiolytic-like action of 3 mg/kg p.o. SSR125543A was investigated. Mice were administered with the compound and placed in the apparatus 1, 2, 4, or 6 h later. Each animal was tested once. In a third experiment, possible development of tolerance to the anxiolytic-like activity was investigated after repeated administration of SSR12543. The compound was given orally at doses of 3 or 10 mg/kg, once daily for eight consecutive days. Data were analyzed by one-way ANOVA followed by Dunnett's t test.
Mouse Defense Test Battery.
The test was conducted in an oval runway as described previously (Griebel et al., 1997). Procedure: 1) Pretest: 3-min familiarization period. Sixty minutes after p.o. administration of SSR125543A, antalarmin, or diazepam, subjects were placed into the runway for a 3-min familiarization period, in which line crossings were recorded. 2) The rat avoidance test. Immediately after the 3-min familiarization period, the experimenter introduced a hand-held dead rat (killed by CO2 inhalation just before the beginning of the experiment) five times at one end of the runway and brought up to the subject at a speed of approximately 0.5 m/s. Approach was terminated when contact with the subject was made or the subject ran away from the approaching rat. If the subject fled, avoidance distance (the distance from the rat to the subject at the point of flight) was recorded. 3) Chase/flight test. The rat was then brought up to the subject at a speed of approximately 2 m/s. A constant distance of 2 m separated the rat and the subject when the former was introduced in the runway. The following parameters were recorded: number of stops (pause in movement), orientations (subject stops then orients the head toward the rat) and chase speed (measured when the subject is running straight). The rat was removed after the chase was completed. 4) Straight alley. By the closing of two doors (60 cm distant from each other), the runway was then converted to a straight alley in which the subject was constrained. The rat was introduced in one end of the straight alley. During 30 s, the number of approaches/withdrawals (subject must move more than 20 cm forward from the closed door then return to it) and immobility time were recorded. After this session, it was removed from the straight alley area. 5) Forced contact. Finally, the experimenter brought the rat up to contact the subject in the straight alley. Approaches were directed quickly (within 1 s) to the subject's head. For each such contact, upright postures and bites by the subjects were noted. 6) Post-test: Contextual defense. Immediately after the forced contact test, the rat was removed and the doors were opened. Escape attempts were recorded during a 3-min session. Data were analyzed by one-way ANOVA, followed by Dunnett's t test.
Social Defeat Stress-Induced Anxiety in Elevated Plus-Maze in Mice.
Social defeat was used as stressor before exposure to the elevated plus-maze. This stressor has been shown to produce significant anxiogenic-like activity without any physical signs of distress. The procedure was a modification of the technique described by Miczek (1979). A naı̈ve mouse was placed in the cage of a resident male aggressor, which was selected for high levels of aggression (Simiand et al., 1993). Social agonistic offensive and defensive behaviors were interrupted by the experimenter and the intruder removed from the area when it displayed a submissive posture after being attacked. Thereafter, the intruder was returned to the resident cage for 60 min and placed in a cylindric wire mesh enclosure to avoid physical contact or injury. At the end of the interaction period, the intruder mouse was placed onto the central platform of the elevated plus-maze during a 5-min period (Lister, 1987). To increase slightly open arm exploration, a rim (1 cm in height) surrounded the perimeter of these arms. Time spent in open arms was recorded. The results were expressed as mean ratio of time spent in open arms to total time spent in both open and closed arms. The compounds were administered i.p. or p.o. 15 min (antalarmin), 30 min (diazepam), or 60 min (SSR125543A) before social defeat. Data were analyzed by a single factor ANOVA or with the nonparametric Kruskal-Wallis test. Subsequent comparisons between treatment groups and control were carried out using Dunnett'st test procedure or the nonparametric Mann-WhitneyU test with α adjustment of Holm, respectively.
Maternal Separation-Induced Distress Vocalizations in Guinea Pig Pups.
The procedure was adapted from that described by Molewijk et al. (1996). Briefly, from day 9 of age, pups entered at the most, three pretest sessions (with 2-day intervals) consisting of 5 min of isolation in a sound-attenuated cage equipped with white noise and white illumination, and the duration of their vocalizations were recorded by the experimenter. Immediately after the 5-min isolation, the subjects were returned to their mothers and littermates. Pups emitting vocalization during at least 120 s entered subsequent compound experimentation. Each pup was tested with vehicle and two dose levels of a compound with a washout period of 3 days between each treatment. SSR125543A and antalarmin were given i.p., respectively, 30 min and 3 h before the experiment, which lasted 5 min. A repeated measures ANOVA was used with an appropriate covariance structure for vocalization duration analysis. Dunnett's t test analysis was subsequently applied to determine which dose was significantly different from vehicle.
Antidepressant-Like Activity of SSR125543A
Forced Swimming Test in Rats.
The procedure was a modification of the technique described by Porsolt et al. (1977). Animals were placed in individual glass cylinders containing water. Two swimming sessions were conducted (an initial 15-min pretest followed 24 h later by a 5-min test). The total duration of immobility was measured for a 5-min period. The animal was judged to be immobile whenever it remained floating passively in the water. SSR125543A, fluoxetine, and antalarmin were administered p.o. twice (15 min after the first session on day 1, and 60 min before session 2 on day 2). Data were analyzed by one-way ANOVA followed by Dunnett's ttest.
CMS.
This test is based on the procedure originally designed by Willner et al. (1992) for rats, and recently adapted for mice byKopp et al. (1999). The CMS protocol consists of the sequential application of a variety of mild stressors, including restraint, forced swimming, water deprivation, and pairing with another stressed animal, each for a period of between 2 and 24 h, in a schedule that lasts for 3 weeks, and is repeated thereafter (Table1). Reduction of body weight gain has been observed in chronically stressed rodents, thereby providing a valid measure of a depressant symptom. Parallels between human depression and chronically stressed animals have also been drawn on the reduction of the efficiency with which even the smallest tasks (e.g., washing and dressing in the morning) are accomplished in depressed patients, leading to the inability to maintain minimal personal hygiene, and the decrease in grooming behavior seen in stressed animals. In this latter case there is a degradation of the physical state of the coat, consisting of a loss of fur and dirty fur. Based on these observations, we measured body weight and physical state about once a week over the entire CMS period. Results were analyzed by a two-way ANOVA (treatment × week) with repeated measures followed by Newman-Keuls post hoc test. At the end of the 10-week CMS period, mice were tested in the elevated plus-maze and the light/dark tests (for details, see above) to assess the impact of CMS on anxiety levels. These behaviors were evaluated because individuals with a major depressive episode frequently present with symptoms of anxiety (DSM-IV, 1994). In the elevated plus-maze test, anxiety was evaluated by measuring the number of mice that entered the open arms, whereas in the light/dark test, it was assessed by measuring activity in the illuminated box. In the former, results were expressed as total number of mice that entered the open arms and analyzed by a chi square independence (χ2) test, whereas in the latter, data were analyzed by a Student's t test. The administration of 10 mg/kg SSR125543A started 4 weeks after the beginning of the CMS. Animals were treated i.p. once a day until all experiments were completed (30 days).
Chronic mild stress schedule
Evaluation of Potential Side Effects of SSR125543A
Effects on Spontaneous Locomotor Activity in Mice: Actimeter.
Testing was conducted in square, clear Plexiglas boxes (22 × 27 × 10 cm) equipped with infrared beams and sensors and placed in sound-attenuated cupboards. Horizontal locomotor activity was quantified as total number of beams crossed during a 20-min period. Sixty minutes after p.o. administration of vehicle or various doses of SSR125543A, subjects were placed individually in the center of the apparatus. ED50 values were calculated by probit analysis.
Effects on Muscle Tone in Mice: Horizontal Wire Test.
It consisted of individually taking mice by the tail and allowing them to grasp a horizontally strung wire (20 cm above the bench level, 2 mm in diameter, 15 cm in length) with their forepaws. Inability to grasp the wire with the forepaws or inability to actively grasp the wire within 5 s with at least one hindpaw is measured. ED50 values were calculated by probit analysis. Experiments were performed 60 or 120 min after oral administration of SSR125543A.
Effects on Motor Coordination in Mice: Rotarod Test.
The apparatus consisted of a plastic cylinder (4 cm in diameter) turning at 4 turns/min. Sixty or 120 min after oral administration of SSR125543A, mice were placed on the turning rotarod. The occurrence of fall from the rotarod was noted during the 2-min period that followed. ED50 values were calculated by probit analysis.
Effects on Learning and Memory in Mice: Passive Avoidance Test.
The apparatus consisted of a black and white two-compartment box separated by a “guillotine” door. The white compartment was small (10 × 10 × 12 cm), lit by a 100-W bulb and with plastic floor. The black compartment was large (22.5 × 16 × 12 cm) with a stainless grid floor connected to a constant current shock generator. Sixty minutes after oral treatment with SSR125543A, mice were placed in the small light compartment. Entrance into the dark box usually occurred within 30 s and was punished by an electrick footshock (0.75 mA, 2 s) (trial 1). The mouse was then returned to its home cage. Twenty-four hours later (trial 2) the mouse was again placed in the light compartment and allowed to explore the box. Animals staying in the illuminated box for more than 60 s were considered as remembering the task. Retention was thus quantified by the percentage of animals avoiding the dark compartment. Comparisons between control and treated groups were performed using Fisher's test.
Results
Anxiolytic-Like Activity of SSR125543A
Punished Drinking Test in Rats.
SSR125543A [F(4,95) = 3.87, P < 0.01] and antalarmin [F(4,65) = 5.58, P < 0.001] significantly increased punished responding at 20 and 10 mg/kg i.p., respectively, whereas diazepam produced similar effects at 3 mg/kg i.p. (Fig. 1). Morphine, at 2.5 mg/kg i.p., did not significantly modify the behavior of animals in this test (T = 1.7, P = 0.11; data not shown).

Effects of diazepam (▪), antalarmin (▧), and SSR125543A (▨) in the punished drinking conflict test in rats. Data represent mean ± S.E.M. ★, P < 0.05 (Dunnett's t test). n = 14 to 20.
Elevated Plus-Maze Test in Rats.
SSR125543A failed to modify significantly both spatiotemporal measures of anxiety and head-dips, but it significantly decreased the number of aborted attempts at entry into open arms followed by avoidance responses over the entire dose range (3–30 mg/kg p.o.) [F(3,23) = 7.58,P < 0.001] (Table 2). Antalarmin increased significantly the percentage of time spent in open arms at 30 mg/kg p.o. [F(3,24) = 3.61,P < 0.05] and the number of head-dips over the edge of the open arms from 10 mg/kg [F(3,24) = 7.79,P < 0.001], but it failed to affect significantly the percentage of entries made into open arms and attempts. Diazepam modified all anxiety-related measures. The compound significantly increased both the percentage of time spent [F(3,28) = 9.46, P < 0.001] and the percentage of entries made [F(3,28) = 6.12, P < 0.01] into open arms. With respect to the ethologically derived measures, it significantly reduced attempts [F(3,28) = 24.36,P < 0.001] and increased head-dips [F(3,28) = 9.26, P < 0.001]. Finally, none of the compounds significantly modified the number of total arm entries, a reliable measure of motor activity.
Effects of SSR125543A, antalarmin, and diazepam in the elevated plus-maze test in rats
Stress-Induced Hyperthermia in Rats.
All the compounds reduced the rise in body temperature after isolation stress (Fig.2). These effects reached statistical significance at all three time periods for SSR125543A [F(3,168) = 348.85, P < 0.001] at 3 and 10 m/kg p.o., antalarmin [F(3,36) = 57.13,P < 0.001] at 30 mg/kg i.p., and diazepam [F(3,36) = 54.87, P < 0.001] at 4 mg/kg p.o.

Effects of SSR125543A, antalarmin, and diazepam on isolation stress-induced hyperthermia in rats. Data represent mean ± S.E.M. ★, P < 0.05 (Dunnett'st test). n = 7 to 17.
Stress-Induced Cortical Norepinephrine Release in Rats.
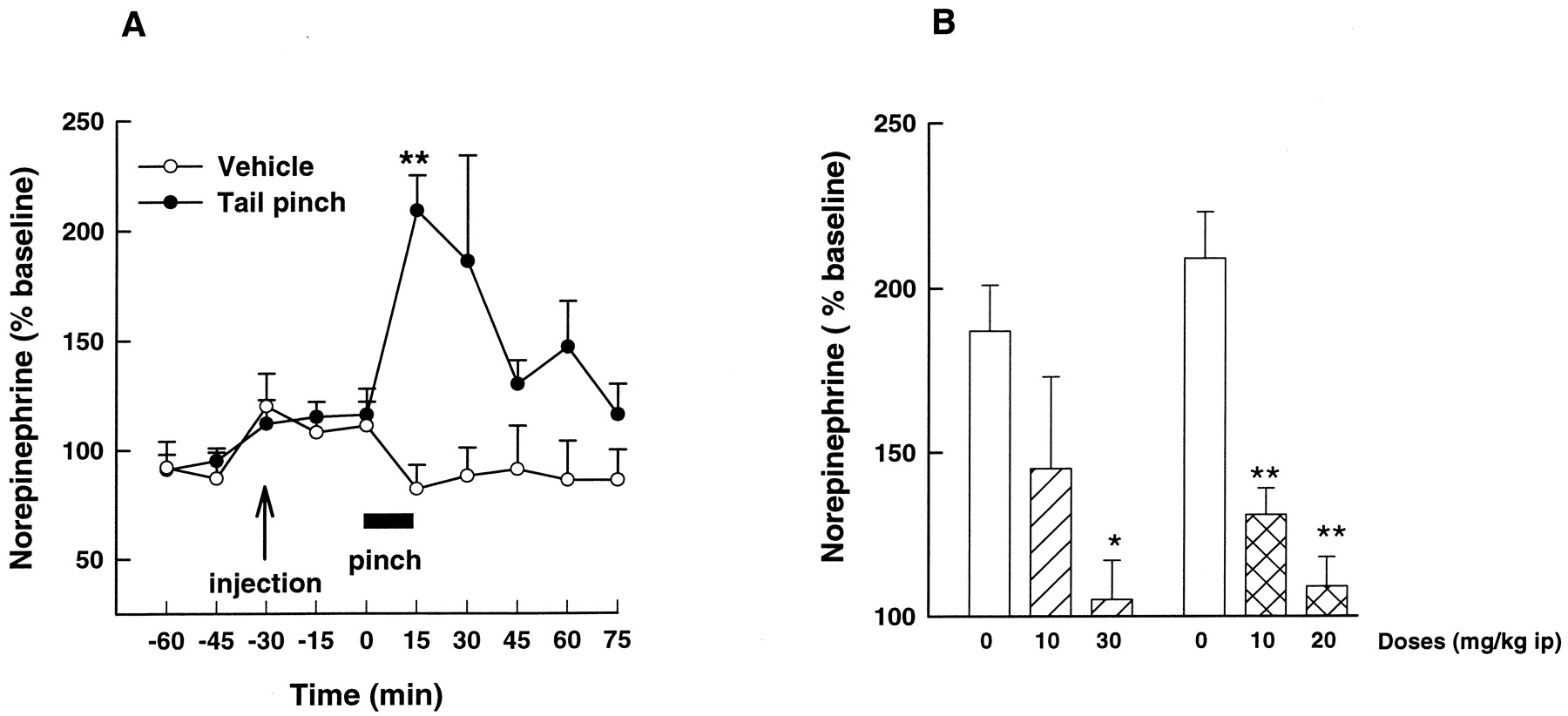
In awake rats, a 15-min tail pinch induced a marked and transient increase in NE release in the prefrontal cortex (109 ± 16 and 87 ±15% of stimulation over baseline level, F(1,9) = 11.74,P < 0.01, n = 5 for each control group). NE release returned to basal levels 45 min after cessation of the stimulation [repeated ANOVA: F(5,54) = 4.97,P < 0.001] (Fig. 3A). SSR125543A (10 and 20 mg/kg i.p.) administered 180 min before tail pinch, significantly [F(2,12) = 23.88,P < 0.001) reduced the evoked NE release (Fig. 3B). Similarly, the administration of the CRF receptor antagonist antalarmin (10–30 mg/kg i.p.) 30 min before tail pinch reduced [F(2,15) = 4.39, P < 0.05] the enhancing effect of tail pinch on cortical NE release. Under these conditions, basal extracellular NE levels in the prefrontal cortex were not affected by SSR125543A or antalarmin.

A, effect of 15 min tail pinch on prefrontal cortex NE release. The changes in NE levels are expressed as a percentage of the mean value of the three basal samples collected at −60, −45, and −30 min before the tail pinch. Each data point represents mean ± S.E.M of five to six animals, P < 0.01 compared with control group by ANOVA with repeated measures followed by Dunnett's t test. B, reversal of tail pinch-induced NE release by SSR125543A and antalarmin given i.p. 180 min (
 ) and 30 min (▨) before pinch, respectively. ■, vehicle. Each data point represents mean ± S.E.M of NE release during tail pinch from five to seven animals. ★, P < 0.05, ★★P < 0.01 compared with respective control group by ANOVA followed by Dunnett's t test.
) and 30 min (▨) before pinch, respectively. ■, vehicle. Each data point represents mean ± S.E.M of NE release during tail pinch from five to seven animals. ★, P < 0.05, ★★P < 0.01 compared with respective control group by ANOVA followed by Dunnett's t test.
Light/Dark Test in Mice.
Neither SSR125543A nor antalarmin (1–30 mg/kg i.p.) significantly modified the behavior of mice in this procedure. This was in contrast to diazepam, which affected all anxiety-related measures in both experiments (Table3). It increased significantly the time spent [SSR125543A and antalarmin experiments: F(5,80) = 15.54, P < 0.001 and F(5,74) = 13.38, P < 0.001, respectively] and activity [SSR125543A and antalarmin experiments: F(5,80) = 17.51, P < 0.001 and F(5,74) = 6.77,P < 0.001, respectively] in the illuminated box, and increased the number of tunnel crossings between both boxes [SSR125543A experiment only: F(5,80) = 15.31,P < 0.001]. Moreover, diazepam significantly decreased the number of aborted attempts at entry into the lit box [SSR125543A and antalarmin experiments: F(5,80) = 2.7,P < 0.05 and F(5,74) = 6.3,P < 0.001].
Effects of SSR125543A, antalarmin, and diazepam in the light/dark test in mice
Four-Plate Test in Mice.
In the acute experiments, all the compounds increased the number of punished crossings [SSR125543A: χ2 = 27.24, P < 0.01; antalarmin: T = 4.38, P < 0.01; diazepam:F(3,46) = 36.04, P < 0.01] (Table4). Post hoc analysis revealed that these effects reached statistical significance with SSR125543A from 1 mg/kg p.o., antalarmin at 30 mg/kg p.o., and diazepam over the entire dose range (1–4 mg/kg p.o.). When SSR125543A was given repeatedly at 3 and 10 mg/kg p.o. for 8 days, the anxiolytic-like activity was still present [F(2,52) = 6.6, P < 0.01]. Finally, the time course of the anxiolytic-like action of SSR125543A at 3 mg/kg p.o. indicated that effects lasted up to 4 h [F(4,55) = 4.72, P < 0.01].
Effects of acute and repeated (once/day/8 days) treatments with SSR125543A in the four-plate test in mice, and time course of the acute effects
Mouse Defense Test Battery.
When given orally, SSR125543A and diazepam, but not antalarmin significantly modified both flight measures [SSR125543A: avoidance distance: F(3,23) = 10.41, P < 0.001; avoidance frequency:F(3,26) = 6.1, P < 0.01; diazepam: avoidance distance: F(4,42) = 8.62, P< 0.001; avoidance frequency: F(4,50) = 18.66,P < 0.001] (Table 5). Whereas SSR125543A decreased avoidance distance at 30 mg/kg and avoidance frequency from 10 mg/kg, diazepam decreased both parameters from 3 mg/kg. When mice were chased by the rat, only diazepam significantly decreased flight speed at 3 and 10 mg/kg [F(4,50) = 4.84, P < 0.01]. Furthermore, antalarmin and diazepam, but not SSR125543A significantly decreased risk assessment behaviors during the chase test [antalarmin: stops: F(4,33) = 5.31, P < 0.01; orientations: F(4,33) = 3.72, P < 0.05; diazepam: stops: F(4,50) = 43.98,P < 0.001; orientations: F(4,50) = 12.47, P < 0.001]. However, only diazepam was able to modify this behavior in the straight alley situation [approach/withdrawal: F(4,50) = 14.55,P < 0.001; immobility; F(4,50) = 5.1,P < 0.01]. Upon forced contact with the rat, all compounds significantly decreased defensive threat and attack reactions [SSR125543A: upright postures: F(3,26) = 4.1,P < 0.05; bites: F(3,26) = 19.48,P < 0.001; antalarmin: upright postures:F(4,33) = 3.35, P < 0.05; bites:F(4,33) = 6.36, P < 0.001; diazepam: upright postures: F(4,50) = 34.96, P < 0.001; bites: F(4,50) = 39.31, P < 0.001] (Fig. 4). Whereas SSR125543A and antalarmin decreased upright postures at 10 and 30 mg/kg, diazepam produced such an effect from 1 mg/kg. Defensive biting was reduced by the two CRF1 receptor antagonists at all doses, and by diazepam from 1 mg/kg. After the removal of the rat from the runway cage, only diazepam significantly reduced the number of escape attempts at 1 mg/kg [F(4,50) = 34.45,P < 0.001]. Finally, none of the compound treatment significantly altered activity before (line crossings) exposure to the rat (Table 5).
Effects of SSR125543A, antalarmin, and diazepam on several behavioral responses displayed by Swiss mice before (line crossings), during (chase speed, immobility time, flight, and risk assessment) and after (contextual defense) exposure to a Long Evans rat in the mouse defense test battery

Effects of SSR125543A, antalarmin, and diazepam on defensive threat and attack reactions upon forced contact with a hand-held dead Long Evans rat in the mouse defense test battery. Data represent mean ± S.E.M. ★, P < 0.05 (Dunnett's t test). n = 7 to 8.
Social Defeat Stress-Induced Anxiety in Elevated Plus-Maze in Mice.
Statistical analysis indicated significant effects in all compound-treatment groups: SSR125543A [F(3,36)=25.56,P < 0.01]; antalarmin [χ2 = 17.5, P < 0.01]; and diazepam [F(2,22) = 8.46, P < 0.01]. Post hoc analysis revealed that social defeat had a significant impact on the performance of mice exposed to the elevated plus-maze as was shown by the dramatic decrease in the percentage of time spent in open arms in stressed animals compared with control mice (Fig.5). This anxiogenic-like effect of stress was significantly antagonized by oral pretreatment with SSR125543A at 10 and 30 mg/kg, antalarmin at 30 mg/kg, and diazepam at 2 mg/kg.

Effects of SSR125543A (▨), antalarmin (▧), and diazepam (
 ) on social defeat-induced anxiogenic-like behavior in the elevated plus-maze test in mice. Data represent mean ± S.E.M. ★,P < 0.05 versus nonstressed animals (■); †,P < 0.05 versus vehicle-treated stressed mice (▪); Newman-Keuls test. n = 8 to 10.
) on social defeat-induced anxiogenic-like behavior in the elevated plus-maze test in mice. Data represent mean ± S.E.M. ★,P < 0.05 versus nonstressed animals (■); †,P < 0.05 versus vehicle-treated stressed mice (▪); Newman-Keuls test. n = 8 to 10.
Maternal Separation-Induced Distress Vocalizations in Guinea Pig Pups.
There was a significant effect in both the SSR125543A [F(3,48) = 8.56, P < 0.001] and the antalarmin [F(3,57) = 13.58, P < 0.001] group. Further analysis showed that SSR125543A at 10 mg/kg i.p. and antalarmin at 3 and 30 mg/kg i.p. significantly reduced distress calls of guinea pig pups separated from their mother (Table6).
Effects of SSR125543A and antalarmin on separation-induced distress vocalizations in guinea pig pups
Antidepressant-Like Activity of SSR125543A
Forced Swimming Test in Rats.
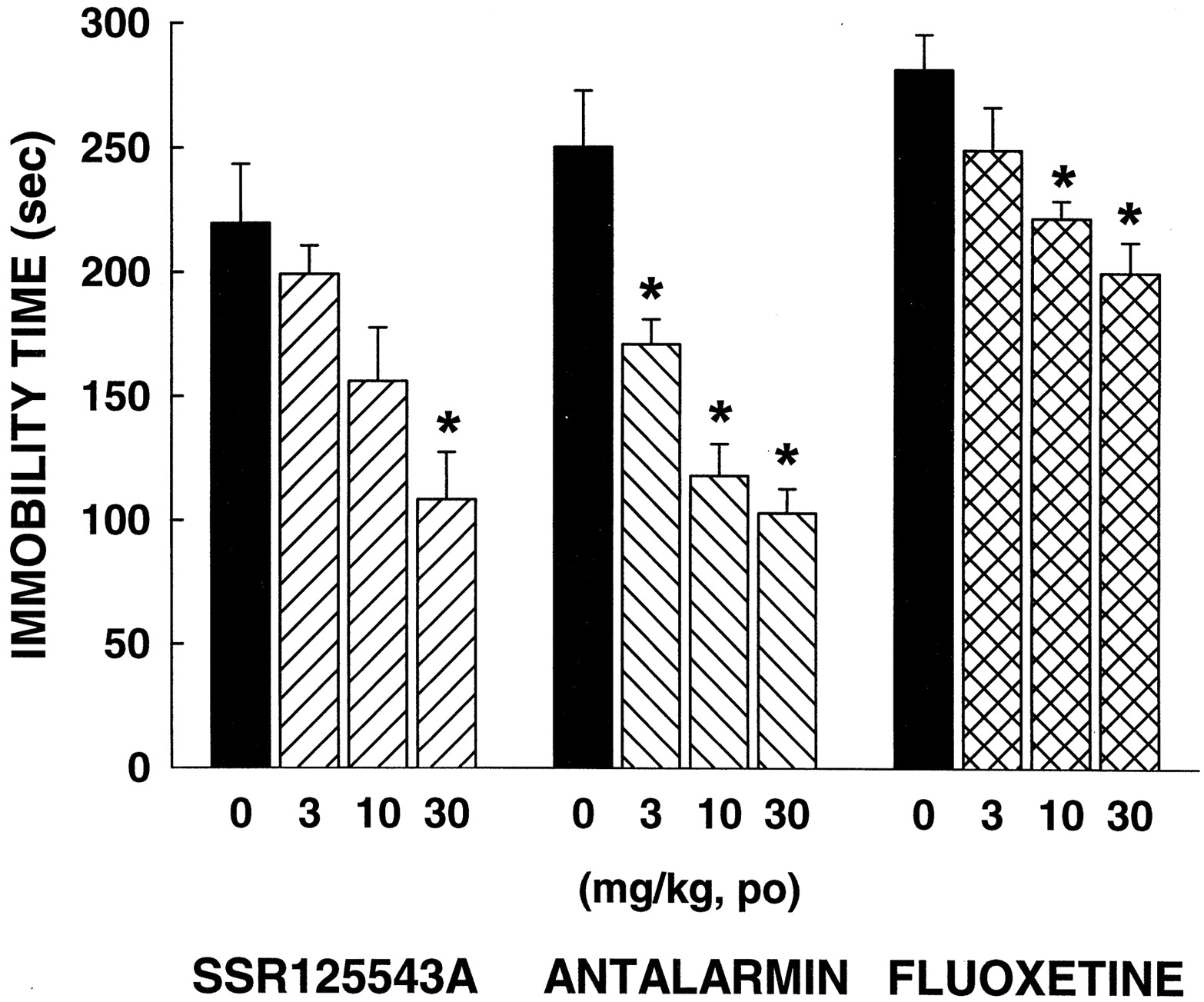
ANOVA indicated significant effects with SSR125543A [F(3,24) = 6.42,P < 0.01], antalarmin [F(3,23) = 20.13, P < 0.001], and fluoxetine [F(3,23) = 6.88, P < 0.01]. Dunnett's analysis showed that SSR125543A significantly decreased immobility time at 30 mg/kg p.o., whereas antalarmin and fluoxetine produced such effects at 10 and 30 mg/kg p.o. (Fig.6).
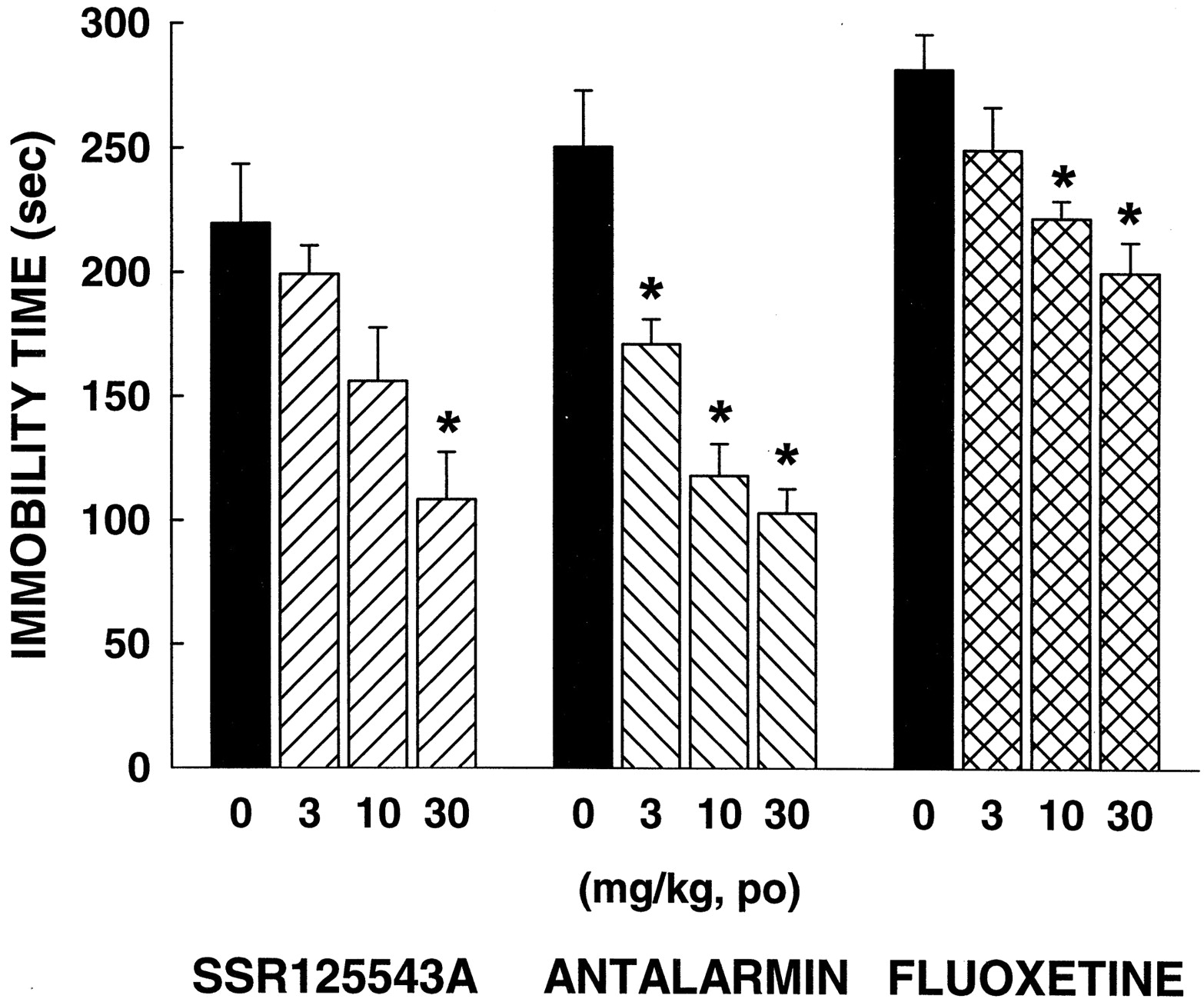
Effects of SSR125543A, antalarmin, and fluoxetine in the forced swimming test in rats. Data represent mean ± S.E.M. ★, P < 0.05 (Dunnett's t test).n = 6 to 7.
Chronic Mild Stress.
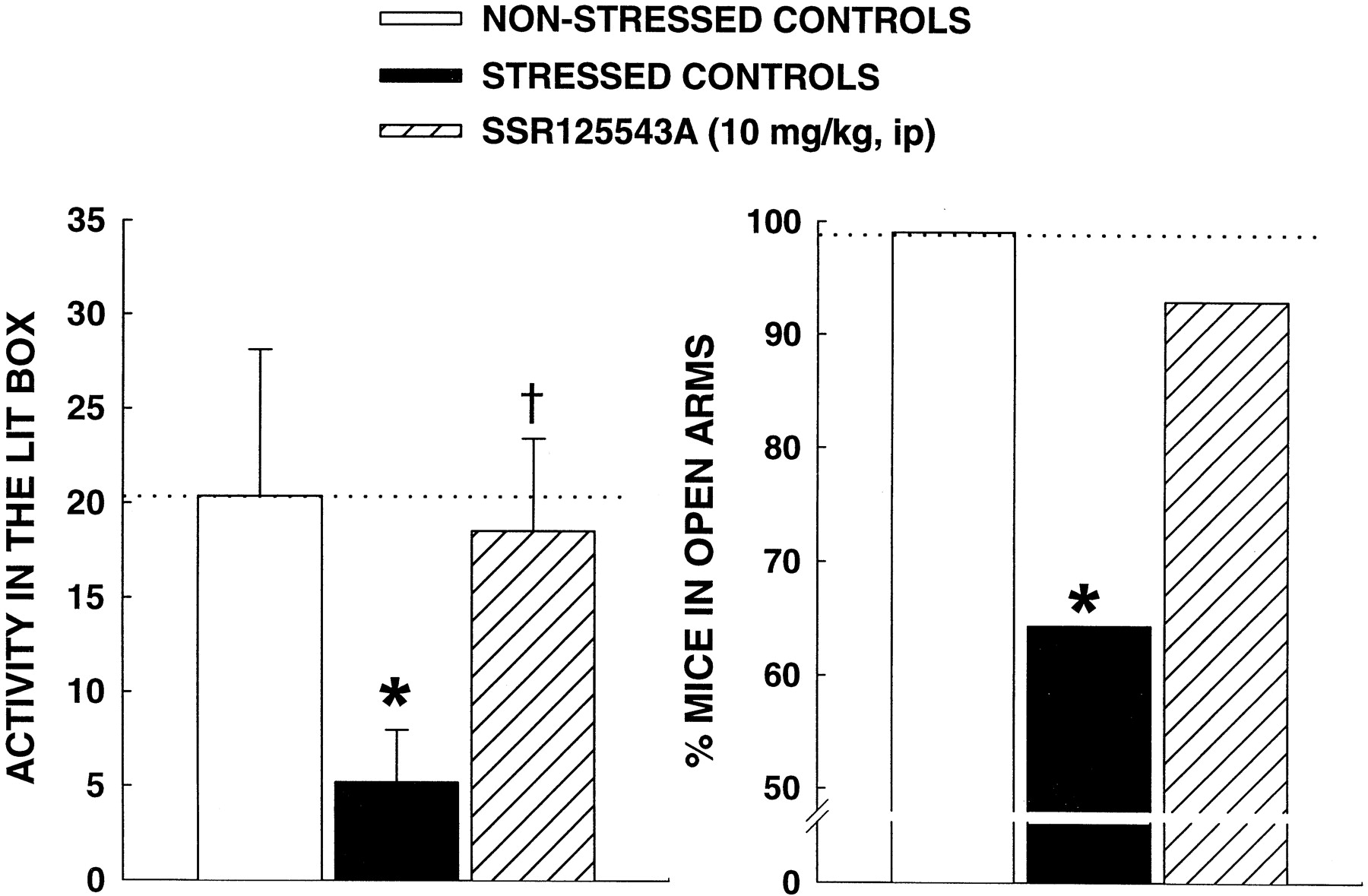
Two-way ANOVA with repeated measures showed a significant effect for weight [F(12,282) = 3.83, P < 0.01] and for physical state [F(16,376) = 4.05, P < 0.001]. Further analysis indicated a significant less body weight gain in vehicle-treated stressed animals from week 5 to the end of the experiment (week 10) (Fig. 7, top). In stressed mice treated with SSR125543A (10 mg/kg i.p.) from week 6, there was also a significant reduction in body weight gain before the treatment was initiated (i.e., at weeks 5 and 6). However, two weeks after the beginning of the treatment, SSR125543A significantly improved the reduction of body weight gain compared with stressed controls, an effect that lasted until the end of the CMS procedure (Fig. 7, top). There was a significant degradation of the physical state of the coat of mice due to stress. In the vehicle-treated group, the effect lasted until the end of the 10-week CMS (except at week 4). In contrast, the degradation of the physical state of the animal's coat was significantly improved by SSR125543A after 2 weeks of treatment, an effect that lasted until the CMS was over (Fig. 7, bottom). Moreover, chronically stressed mice showed reduced activity in the illuminated box of the light/dark apparatus (T = −1.9, P < 0.05) and made significantly less entries into the open arms of the elevated plus-maze (χ2 = 6.84,P < 0.05) compared with nonstressed controls. The anxiogenic-like effect of stress in the former was reversed significantly by SSR125543A (P = 0.03, versus stressed controls), whereas a strong trend for an antagonism of the effect was observed in the elevated plus-maze (P = 0.1, versus stressed controls) (Fig. 8).

Effects of repeated administration of SSR125543A on the reduction of body weight gain (top) and on the degradation of the physical state of the coat of animals (bottom) in chronically stressed (CMS) mice. Data represent mean ± S.E.M. ★,P < 0.05 versus vehicle-treated stressed animals (●); †, P < 0.05 versus nonstressed mice (▪); Newman-Keuls test. n = 15 to 18.

Effects of repeated administration of SSR125543A for 30 days on chronic mild stress-induced anxiogenic-like behavior in the elevated plus-maze (right) and light/dark (left) tests in mice. Data represent mean ± S.E.M. ★, P < 0.05 versus nonstressed animals (■); †, P < 0.05 versus vehicle-treated stressed mice (▪), Student's t test or χ2. n = 10 to 15.
Evaluation of Potential Side Effects of SSR125543A (Table7)
Effects on Spontaneous Locomotor Activity in Mice: Actimeter.
After oral administration of increasing doses of SSR125543A (3–60 mg/kg), spontaneous locomotor activity was not significantly affected over the 20-min recording period (data not shown).
Summary of the pharmacological properties of SSR125543A in rodents
Effects on Muscle Tone in Mice: Horizontal Wire Test.
Results showed that none of the mice treated orally with SSR125543A up to 100 mg/kg failed to pull themselves up and put their hindlimbs on the wire within 5 s at any time postadministration (60 or 120 min) (data not shown).
Effects on Motor Coordination in Mice: Rotarod Test.
Results indicated that all the animals administered with an oral dose of SSR125543A up to 100 mg/kg were able to maintain themselves on the turning rod 60 or 120 min after administration of the compound (data not shown).
Effects on Learning and Memory in Mice: Passive Avoidance Test.
In trial 2, nine of 10 vehicle-treated mice that were placed into the lit box did not enter the black box over the 60-s test period, thus remembering the electric shock received the previous day. Animals treated orally with SSR125543A at 10, 30, and 100 mg/kg displayed comparable performance because 90, 80, and 90% of them, respectively, did not leave the illuminated box.
Discussion
The results of the present study revealed that the selective CRF1 receptor antagonist SSR125543A displayed a behavioral profile in rodents that is consistent with an anxiolytic-like action. In the punished drinking conflict procedure in rats and in the four-plate test in mice, SSR125543A and antalarmin produced an increase in rates of responding suppressed by punishment. The absence of significant modifications in rates of unpunished crossings in the four-plate procedure (data not shown), taken together with the findings from activity tests (Table 7), indicate that the anxiolytic-like effects were observed at doses that did not impair motor activity. Although in the four-plate test the increase in punished responding with the CRF1 compounds was somewhat smaller than that produced by diazepam, suggesting weaker anxiolytic-like activity, in the punished drinking test, the magnitude of the anticonflict effects was similar to that of benzodiazepine. Although in a previous study we failed to detect any anticonflict activity of the CRF1 receptor antagonist CP-154,526 (Griebel et al., 1998), the present positive effects of SSR125543A and antalarmin agree with other studies that reported anticonflict activity of CRF1 antagonists (Brocco et al., 1998), thereby indicating that such models are suitable for the screening of these compounds. It is unlikely that the positive effects of CRF1 receptor antagonists in punished-based anxiety models are due to decreased sensitivity to electric shocks because there is as yet no evidence that these compounds have analgesic effects. Moreover, compounds that are endowed with potent analgesic properties, such as morphine, are inactive in conflict tests (Treit, 1985; present data). Two additional experiments performed in the four-plate test showed that the anxiolytic-like activity of SSR125543A (3–10 mg/kg p.o.) lasted for up to 4 h and was still present after repeated administration of the compound for 8 days. This latter result indicates that no tolerance to the anxiolytic-like activity of SSR125543A develops, which is in agreement with a previous finding showing a lack of tolerance to the anxiolytic-like effects of another CRF1 antagonist, CP-154,526, in the defensive withdrawal paradigm in rats (Arborelius et al., 2000).
Results from exploration-based models of anxiety showed that SSR125543A and antalarmin produced weak anxiolytic-like activity in the elevated plus-maze test in rats and were completely inactive in the light/dark test in mice. In the former model, SSR125543A only significantly decreased risk assessment (i.e., attempts), and antalarmin weakly but significantly increased one of two spatiotemporal measures of anxiety and directed exploration. Negative or weak effects of CRF1 receptor antagonists in anxiety models based on spontaneous exploratory activity are not uncommon (Lundkvist et al., 1996; Griebel et al., 1998; Okuyama et al., 1999; He et al., 2000). It has been suggested that CRF1 receptor antagonists may produce anxiolytic-like effects only in stressed animals when CRF levels are increased (Menzaghi et al., 1994). To illustrate this idea, several studies have shown that CRF receptor antagonists, including nonpeptide (i.e., CRA1000, CRA1001) and peptide (α-helical CRF9–41, astressin) ligands produced anxiolytic-like effects in the elevated plus-maze or the light/dark test only when animals had been stressed by exposure to conspecific aggression or by forced swimming (Heinrichs et al., 1992; Menzaghi et al., 1994; Okuyama et al., 1999; Spina et al., 2000). Moreover, findings from Adamec et al. (1991) have revealed that repeated handling altered the anxiolytic-like effects of α-helical CRF9–41 in the elevated plus-maze. Consequently, we investigated the effects of SSR125543A and antalarmin in stressed animals. Introduction of a socially naive male mouse into the home territory of a resident counterpart results in agonistic interactions, leading to the rapid social defeat of the intruder. Exposure to the aggressive resident has been shown to produce a stress-response profile consisting of neuroendocrine activation and coping behaviors such as submission (Heinrichs et al., 1992). Subsequent exposure of defeated animals to the elevated plus-maze resulted in a dramatic reduction of exploration of open arms, which is indicative of increased levels of anxiety. Our results showed that SSR125543A and antalarmin administered before social conflict reversed the heightened emotionality produced by the resident exposure stressor in this test. This action paralleled that of an anxiolytic dose of diazepam.
The findings obtained in the MDTB further support the idea that baseline levels of stress are of crucial importance when investigating the behavioral actions of CRF1 receptor antagonists. Although diazepam affected all defensive responses, thereby showing clear anxiolytic-like activity, SSR125543A and antalarmin had weak or no effects in the phases where escape from the oncoming rat was possible (i.e., rat avoidance and chase/flight tests) or when the threat stimulus was removed from the test arena (i.e., contextual defense). In contrast, both compounds displayed clear-cut effects upon contact with the rat as was shown by the marked action on defensive upright posture and biting. These effects are unrelated to motor impairment because activity measures recorded before and during confrontation with the rat were not significantly altered by the compounds. Unlike the other phases of the MDTB, the forced contact test has been suggested to be particularly stressful for animals because they have no possibility to escape and confrontation with the threat stimulus is unavoidable. Thus, assuming that the CRF system contributes significantly to the stress responses displayed by mice in this test battery, we would expect a CRF antagonist to attenuate these reactions.
The antistress profile of SSR125543A and antalarmin was confirmed in three additional experiments. Both compounds counteracted the increase in body temperature after isolation stress, as did diazepam. In addition, they clearly reduced distress vocalizations in guinea pig pups, although for some undetermined reasons the effects of antalarmin were not dose-dependent. This latter finding is in agreement with a previous study showing that antalarmin reduced distress vocalizations in rat pups (Coverdale et al., 1998). Furthermore, the compounds prevented tail pinch stress-induced increase in cortical NE release. This latter finding supports further the idea that stress-induced activation of the prefrontal NE system may be under the control of endogenously released CRF (Steinberg et al., 2000).
The idea that nonpeptide small molecule CRF1receptor antagonists may represent novel treatment for depressive disorders is substantiated by a large body of animal and clinical data (for reviews, see Holsboer, 1999; O'Brien et al., 2001). The most compelling evidence comes from the finding that the CRF1 receptor antagonist R121919 reduced depressive symptomatology in a small open-label study involving 20 patients with a major depressive episode (Zobel et al., 2000). To investigate potential antidepressant-like effects of SSR125543A and antalarmin, we used the forced swimming test, a classical acute model of depression (Porsolt et al., 1977). In addition, the effects of repeated administration of SSR125543A were investigated in the chronic mild stress model (Willner et al., 1992). Results from the forced swimming test showed that SSR125543A and antalarmin produced dose-dependent antidepressant-like activity. The antidepressant potential of SSR125543A was confirmed in the chronic mild stress procedure. SSR125543A antagonized significantly the reduction of body weight gain seen in chronically stressed mice after 2 weeks of treatment. Moreover, the compound improved the degradation of the physical state of the coat of stressed animals. These findings suggest that SSR125543A normalized grooming and feeding behaviors, two activities impaired by chronic stress. Importantly, SSR125543A has no effect on food intake or body weight in normal mice (Arnone, personal communication), indicating that the compound-induced increase in body weight in stressed animals is not due to a direct action of SSR125543A on feeding. Chronic mild stress caused the appearance of an “anxious” profile as was evidenced by the findings from the elevated plus-maze and the light/dark tests. These behavioral changes were not seen in animals treated with SSR125543A for 30 days, indicating that the compound was able to prevent the stress-induced increase in anxiety levels. Based on the high predictive validity of the chronic mild stress model, indicating that only clinical effective antidepressants are active in this test (Papp et al., 1996), these findings clearly suggest that SSR125543A has antidepressant-like properties. A question that arises from these results is whether the onset of antidepressant-like action of SSR125543A may be shorter than that of currently used antidepressants. Preliminary data from our laboratory with a slightly modified version of the chronic mild stress indicate that the prototypical antidepressant fluoxetine was active after 2 weeks of treatment, as was SSR125543A. However, additional experiments are required before any definite conclusion can be drawn on the onset of the antidepressant-like action of SSR125543A.
In conclusion, this study demonstrates that the potent, selective nonpeptide CRF1 receptor antagonist SSR125543A is able to reduce anxiety- and depressive-related responses in several animal models. Although the compound has limited or no efficacy in classical models of anxiety, it shows good activity in acute and chronic tests of unavoidable stress exposure. Importantly, the side effect profile of SSR125543A compares favorably with that of currently used anxiolytic agents because the compound was devoid of effects in tests of activity and memory up to 100 mg/kg, a dose much higher than those producing anxiolytic- and antidepressant-like effects. Together, these data suggest that SSR125543A may have a potential in the treatment of depression and some forms of anxiety disorders.
Acknowledgments
The expert technical assistance of Carmen Aliaga, Georgette Gout, Monique Lhermitte, and Nicolas Moindrot is greatly appreciated. The automation of most procedures used in this study was carried out by Bernard Kleinberg to whom we are thankful.
Footnotes
- Abbreviations:
- CRF
- corticotropin-releasing factor
- ACTH
- adrenocorticotropic hormone
- MDTB
- mouse defense test battery
- CMS
- chronic mild stress
- NE
- norepinephrine ANOVA, analysis of variance
- CP-154,526
- butyl-[2,5-dimethyl-7-(2,4,6-trimethylphenyl)-7H-pyrrolo[2,3-d]pyrimidin-4-yl]-ethylamine
- R121919
- 3-[6-(dimethylamino)-4-methyl-pyrid-3-yl]-2,5-dimethyl-N,N-dipropyl-pyrazolo[2,3-a]pyrimidin-7-amine
- Received October 11, 2001.
- Accepted December 31, 2001.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}