


Abstract
Corticotropin-releasing factor (CRF) is one of the principle components of the stress response. The physiological effects of CRF are mediated by two receptor subtypes, CRF1 and CRF2. Recent data obtained with the selective CRF2 antagonist antisauvagine-30 (ASV-30) has begun to suggest that both CRF receptor subtypes may play a role in stress-related behaviors. Exactly how these two receptor subtypes interact to modulate the behavioral and endocrine responses to stress is not clear, however. We have attempted to understand the role of the CRF2 receptor in the behavioral and endocrine responses to stress by comparing the effects of ASV-30 with the mixed CRF1/CRF2 receptor antagonist astressin. Centrally administered ASV-30 reduced anxiety-like behavior in BALB/c mice in three models of anxiety: marble burying [minimal effective dose (MED) = 3 nmol], open field (MED = 3 nmol), and elevated plus maze (MED = 0.1 nmol). ASV-30 did not change locomotor activity or the adrenocorticotropic hormone (ACTH) response to restraint stress. The potent mixed CRF1/CRF2antagonist astressin not only reduced anxiety-like behavior in all three models with equivalent potency but also blunted the ACTH response to restraint stress. Finally, the new selective CRF2receptor agonist urocortin-II produced a dose-dependent increase in anxiety-like behavior in the plus maze test. Therefore, our data suggest that the CRF2 receptor plays a role in the behavioral, but not the hypothalamic-pituitary-adrenal axis, response to stress.
Most anxiety disorders are characterized by the physiological symptoms of arousal, including increased heart rate, blood pressure, and plasma catecholamines (U.S. Department of Health and Human Services, 1999). Central CRF administration mimics many of the above-mentioned symptoms (for review, see Dunn and Berridge, 1990) and produces stress-like behavioral responses such as immobility or activation, depending upon the familiarity of the environment (Britton et al., 1982, 1986). In addition, CRF antagonists attenuate stress-like behaviors in anxiety-inducing paradigms such as the plus maze task, defensive withdrawal, conditioned fear, and open field, suggesting that CRF may play a physiological role in behaviors induced by stressful situations (Britton et al., 1986; Kalin et al., 1988; Menzaghi et al., 1994; Deak et al., 1999; Okuyama et al., 1999; Arborelius et al., 2000). Finally, CRF is elevated in the cerebrospinal fluid of anxiety patients (De Souza and Nemeroff, 1990), suggesting that the CRF system may be hyperactive in anxiety disorders.
The physiological effects of CRF are mediated by two receptor subtypes, CRF1 and CRF2 (Grigoriadis et al., 1996). Although all of the CRF-related peptides bind with similar affinity to the CRF1 receptor, CRF itself is about 30-fold less potent at the CRF2 receptor than urocortin, urotensin, and sauvagine (Donaldson et al., 1996) and greater than 100-fold less potent at the CRF2receptor than the recently discovered urocortin II (Reyes et al., 2001). The two CRF receptor subtypes also show distinct receptor distribution patterns within the rodent limbic system, a region that has been associated with emotional behaviors. CRF1 receptors are primarily localized in the cortical and anterior amygdalar aspects of this circuit, whereas CRF2 receptors have a more subcortical distribution, with the most dense concentrations in the lateral septum, the bed nucleus of the stria terminalis (BNST), the posterior amygdala, and the ventromedial hypothalamus (Chalmers et al., 1995; Rominger et al., 1998; Van Pett et al., 2000; Higelin et al., 2001). In addition, CRF1 receptors are exclusively localized in the anterior and intermediate lobes of the pituitary, whereas CRF2 receptors are found only in the posterior lobe (Chalmers et al., 1995; Van Pett et al., 2000).
Developmental knockout of the CRF1 receptor results in an adult phenotype characterized by a blunted HPA axis response to stress and reduced stress-like behavior (Smith et al., 1998; Contarino et al., 2000). In addition, administration of CRF1-selective small molecule antagonists has been reported to reduce anxiety in a variety of tasks, such as plus maze, defensive withdrawal, and conditioned fear (Deak et al., 1999;Okuyama et al., 1999; Arborelius et al., 2000). These compounds are most effective in models that are based on natural behaviors that are stress-induced, rather than in conditioned anxiety models that are sensitive to benzodiazepines (Griebel et al., 1998). The above-mentioned evidence, along with the receptor subtype localization profile, has led to the hypothesis that the CRF1receptor is the primary mediator for the role of CRF in the physiological response to stress. In contrast, the role of the CRF2 receptor subtype is not well understood. Deletion of the CRF2 receptor gene during development has provided an ambiguous profile, where increased anxiety was observed in some but not all anxiety tasks (Bale et al., 2000; Kishimoto et al., 2000); in males, but not females (Kishimoto et al., 2000); in males and females (Bale et al., 2000); or not at all (Coste et al., 2000). Data obtained using the selective CRF2 antagonist antisauvagine-30 (ASV-30) (Ruhmann et al., 1998) has been more inconsistent. ASV-30 has been reported to be between 100- and 1000-fold selective for the CRF2 receptor, depending upon whether the radiolabeled ligand is sauvagine (Ruhmann et al., 1998) or ASV-30 (Higelin et al., 2001), respectively. Intraseptal administration of ASV-30 has been shown to reduce anxious behavior induced by immobilization stress in the plus maze task or by previous association with footshock in mice (Radulovic et al., 1999). These behavioral data were recently corroborated in rats, where i.c.v. ASV-30 reduced anxious behavior in the plus maze, defensive withdrawal, and a conditioned anxiety paradigm (Takahashi et al., 2001). These recent data suggest that both CRF receptor subtypes may play a role in the behavioral and endocrine responses to environmental stress. We have begun to investigate the relative roles of the two CRF receptors in the response to stress by comparing the efficacy of centrally administered ASV-30 and astressin in three models of anxiety and in the HPA axis response to restraint stress. We also assessed the behavioral effects of the new CRF2-selective agonist Ucn II in one model of anxiety.
Materials and Methods
Subjects.
Male BALB/c mice (18–20 g) were housed in groups of five in the Neurocrine Biosciences, Inc. (San Diego, CA) vivarium, under a standard 12-h light/dark cycle (7:00 AM/7:00 PM) where the ambient room temperature was maintained at 23–25°C. Mice were allowed food and water ad libitum. All testing was conducted according to the standards set by the Institutional Animal Care and Use Committee at Neurocrine Biosciences, Inc.
Design.
All peptides were administered 30 min before testing. Peptides were administered i.c.v., whereas diazepam and fluoxetine were administered i.p. 60 min before testing. Peptides were always injected in a 5-μl volume, using water as the vehicle. Separate cohorts of mice were used for each peptide or drug, and for each test.
Peptides.
Astressin is a potent mixed CRF1/CRF2 antagonist withKi values in the 1 to 12 nM range for both receptors (Gulyas et al., 1995). ASV-30 is a selective CRF2 antagonist, with reportedKi values for inhibition of radiolabeled sauvagine binding to recombinant mouse CRF2β and rat CRF1receptors of 1.4 and 154 nM, respectively (Ruhmann et al., 1998), and aKd value for radiolabeled ASV-30 binding to recombinant human CRF2α receptors of 0.125 nM with no specific binding detected to the recombinant human CRF1 receptor (Higelin et al., 2001). Urocortin II is a CRF2-selective agonist with aKi value of 0.66 nM for inhibition of radiolabeled sauvagine binding to recombinant mouse CRF2β receptors and >100 nM for the recombinant human CRF1 receptor (Reyes et al., 2001). All of these peptides were synthesized by solid phase methodology with a 4-methylbenzhydrylamine resin (Bachem California, Torrance, CA) on a peptide synthesizer (model 990; Beckman Coulter, Inc., Fullerton, CA). The synthetic method usest-butyloxycarbonyl protection, trifluoroacetic acid deprotection, and hydrogen fluoride cleavage of the finished peptide from the resin anchor. The crude peptide product recovered from the cleavage reaction is purified on a preparative high-performance liquid chromatography system (KP-100; Biotage, Charlottesville, VA) on a C18 cartridge, using a linear gradient of acetonitrile in 0.1% trifluoroacetic acid. The purity of the synthetic product was verified by high-performance liquid chromatography analysis, and its correct structure was confirmed by mass spectrometric analysis on a SCIEX AP1 liquid chromatography/mass spectrometry system (PerkinElmer Instruments, Norwalk, CT).
Marble Burying.
The marble-burying task has been characterized as a model of compulsive behavior (obsessive compulsive disorder) because marble burying is not associated with physical danger and does not habituate upon repeated testing (Njung'e and Handley, 1991a). Furthermore, it is most sensitive to the selective serotonin reuptake inhibitor (SSRI) class of psychotherapeutics, which is currently the medication of choice for obsessive compulsive disorder (Njung'e et al., 1991b; U.S. Department of Health and Human Services, 1999). Mice were placed individually in a small mouse-size Plexiglas cage containing bedding that was 5 cm in depth, along with 25 small marbles arranged in six evenly spaced rows of four, with one marble in the middle. Testing was conducted for a 30-min period under red light and white noise. Mice were initially pretested as described above, and only those that buried at least 12 marbles were used for further testing. On the test day, mice were injected i.c.v. with ASV-30 or astressin (0–10 nmol/5 μl) 30 min before marble exposure. Some groups of mice were also tested with fluoxetine (15–30 mg/kg i.p.) or diazepam (2.5–5.0 mg/kg i.p.). After a 30-min exposure to the marbles, mice were removed and the unburied marbles were counted. Marbles were considered buried if they were at least one-half covered with bedding. A different set of mice prescreened for marble burying was tested 1 week later in the same cages without the presence of marbles to assess any peptide-induced changes in locomotor activity. As in the marble burying, they were injected i.c.v. or i.p. 30 min before testing with either ASV-30, astressin (0–10 nmol/5 μl i.c.v.), diazepam (2.5–5.0 mg/kg i.p.), or fluoxetine (15–30 mg/kg). Locomotor activity was measured for a 30-min period, using a 16-beam photocell array that surrounded each cage (Columbus Instruments, Columbus, OH).
Open Field Behaviors.
Thirty minutes after injection of ASV-30, astressin (0–10 nmol/5 μl i.c.v.), or diazepam (0.25–5 mg/kg i.p.) mice were placed into a clear Plexiglas box (50 × 50 × 22 cm) surrounded by an array of photocell beams (Accuscan, Columbus, OH). A lamp directed on the center of the field provided a light level of 120 lux in the center of the arena. Testing was conducted during the light cycle in a room with constant white noise. Each animal was placed in the center of the arena to initiate the 10-min testing session. Time spent and distance traveled in the center and the margins of the Plexiglas arena were recorded as photobeam breaks in 10-min bins.
Elevated Plus Maze.
Mice were injected with ASV-30, astressin (0–10 nmol i.c.v.), or diazepam (0.5–2.5 mg/kg i.p.) 30 min before placement on the plus maze. Some mice were also treated with the recently discovered CRF2-selective agonist Ucn II (0.03–3 nmol i.c.v.) 30 min before plus maze testing. The plus maze was constructed of black Plexiglas and consisted of two open arms (30 × 5 cm) and two enclosed arms of the same dimensions with walls 30 cm in height. The arms extended from a central platform (5 × 5 cm). Ambient fluorescent light provided a light level of approximately 180 lux on the open arms. Testing was conducted during the light cycle. Mice were habituated to the experimental room 1 h before testing, and initially placed on the center platform facing an open arm. A camera placed above the maze recorded any behaviors for a 5-min period. Variables recorded from the resulting videotape included time spent in the center of the maze or the open and closed arms, along with the number of entries into open or closed arms. The percentage of time spent in the open arms and the number of entries into the open arms relative to total time and entries suggest the degree of anxiety (Pellow et al., 1985; File, 2001). In contrast, the number of closed arm entries has been suggested as a measure of activity, rather than anxiety (File, 2001), although this interpretation could be complicated by gender and whether the mouse is albino or pigmented (Cohen et al., 2001; File, 2001).
Functional Specificity of ASV-30: Effects on Duration and Recovery of Restraint Stress-Induced ACTH Release.
Two of the reports describing the phenotype resulting from CRF2 receptor deletion showed that the ACTH response to restraint stress was higher in the knockout mice after 2 min of restraint stress than in wild types (Bale et al., 2000; Coste et al., 2000). One of the reports also showed that the ACTH response to restraint stress in the CRF2 knockout mice was not normalized at the same rate as in wild-type controls, suggesting that the CRF2 receptor could serve to modulate the physiological adaptation to stress (Coste et al., 2000). To address this issue in adult mice, we injected ASV-30 (1 or 10 nmol) or astressin (1 nmol i.c.v.) 30 min before 2, 10, or 30 min of restraint stress. In addition, some animals were treated with ASV-30 (1 or 10 nmol) or astressin (1 nmol) 30 min before the 30-min restraint stress and blood was collected 10, 30, and 60 min after the cessation of restraint stress. ACTH assays were conducted with the resulting plasma using the radioimmunoassay kit (Nicols Institute Diagnostics, San Juan Capistrano, CA).
Statistics.
Dose-response relationships were initially tested for statistical significance using one-way ANOVA with Fisher's least significant difference used as the post hoc test for means comparisons if the main effects were significant. The minimal effective dose (MED) was defined as the lowest dose to reach statistical significance with the α value set at p < 0.05. In cases where dose-response relationships were assessed over time, a mixed design, repeated measures ANOVA (dose × time) was used for the analysis. Statistically significant interactions for these mixed design ANOVAs were simplified using Fisher's least significant difference test. The ACTH responses to restraint stress at different time points were analyzed using a between-groups ANOVA design because different mice were used for each time point.
Results
Marble Burying.
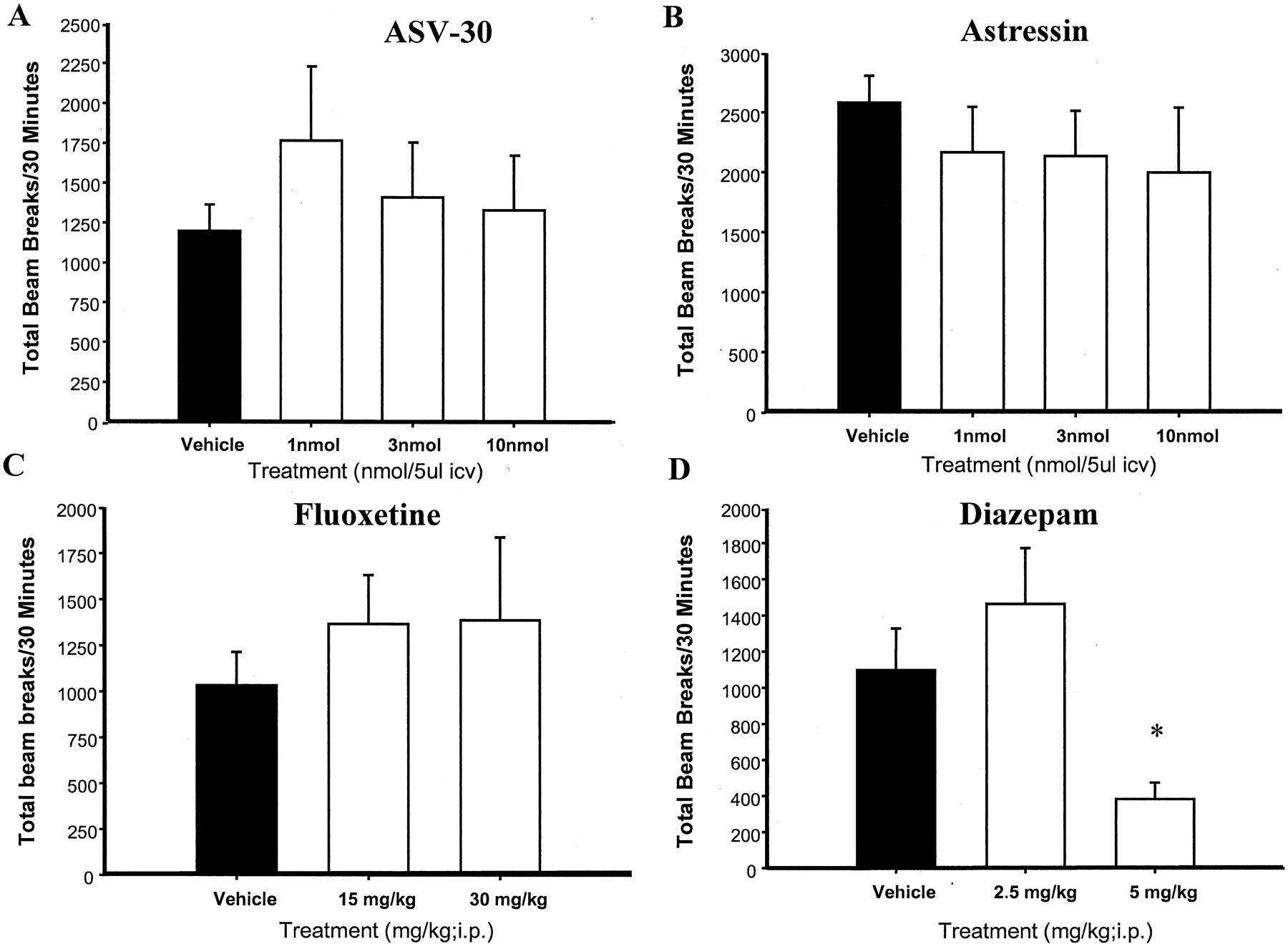
ASV-30 significantly reduced marble burying [F(5,78) = 11.7; p < 0.0001], with the MED at 3 nmol (p < 0.002 versus vehicle; Fig.1A). In contrast, ASV-30 did not significantly alter locomotor activity in the same environment [F(3,1015) = 0.48; p < 0.69] (Fig.2A). Astressin also reduced marble burying [F(4,46) = 3.56; p < 0.01] (Fig. 1B) without having significant effects on locomotor activity [F(4,1334) = 0.39; p < 0.81] (Fig.2B). The MED for astressin was also 3 nmol. Similar to ASV-30 and astressin, fluoxetine significantly reduced marble burying [F(2,31) = 10.25; p < 0.0004] (Fig.1C) without affecting locomotor activity [F(2,754) = 0.42; p < 0.66] (Fig. 2C). Finally, diazepam reduced marble burying [F(2,25) = 4.89; p < 0.016] (Fig. 1D), but also produced a significant reduction in locomotor activity [F(2,897) = 117; p< 0.0001] (Fig. 2D). As can be observed in Fig. 1, astressin, ASV-30, fluoxetine, and diazepam all reduced marble burying to a similar degree.
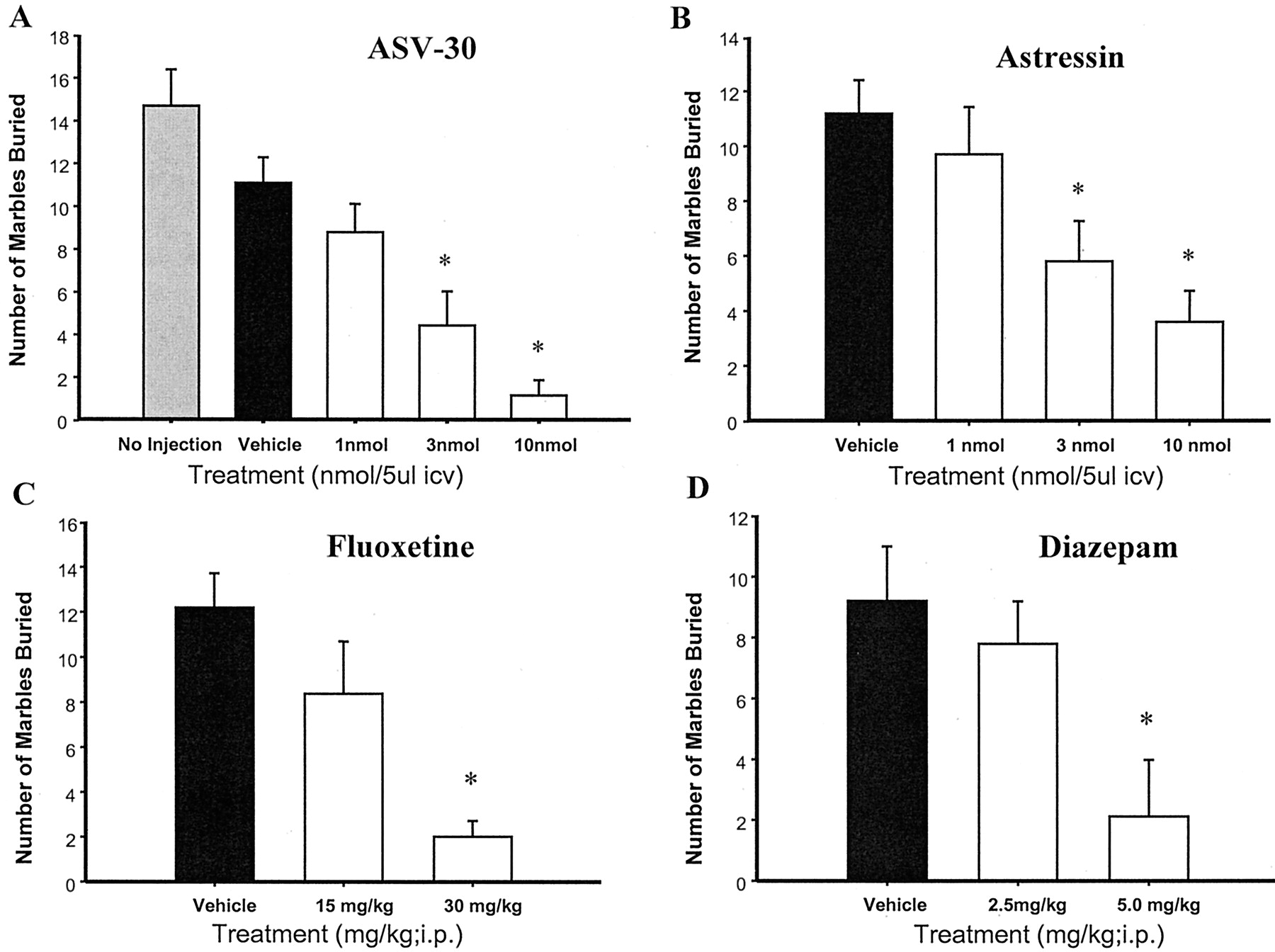
Effects of ASV-30 (A) and astressin (B) on marble burying in comparison with fluoxetine (C) and diazepam (D). All mice were equally distributed between treatment groups based upon the number of marbles buried during a pretreatment screening test. Animals that buried 12 or fewer marbles were not included in the study. Each graph shows the average number of marbles buried 30 min after i.c.v. injection of ASV-30 or astressin or after i.p. injection of fluoxetine or diazepam. There were 9 to 19 mice/group in these studies. ∗,p values < 0.02 to 0.0001 versus vehicle.

Effects of ASV-30 (A) and astressin (B) on locomotor activity in comparison with fluoxetine (C) and diazepam (D). Mice were prescreened for marble burying and distributed across treatment groups as described in the legend to Fig. 1 and then tested for locomotor activity instead of marble burying 30 min after ASV-30, astressin, fluoxetine, or diazepam injection. Locomotor activity was measured as total beam breaks during the 30-min test period. There were 9 to 10 mice in each group. ∗,p < 0.0001 versus vehicle.
Open Field Behaviors.
As can be observed in Fig.3, ASV-30 significantly increased the percentage of time [F(4,64) = 3.4; p< 0.01] spent in the center of the arena. Astressin also increased the proportion of time in the center of the arena but only as a statistical trend [F(4,64) = 2.06; p< 0.09]. Similarly, diazepam increased the percentage of time spent in the center of the arena [F(3,36) = 2.9;p < 0.049], with the maximal effect at 0.5 mg/kg (p < 0.008). The percentage of distance traveled in the center of the arena in relation to the entire area was affected in a similar manner by ASV-30, with the MED at 3 nmol. Astressin and diazepam also increased the proportion of distance traveled in the center of the arena, but the overall F values for distance did not reach statistical significance for either astressin [F(4,64) = 2.09; p < 0.09] or diazepam [F(3,36) = 2.42; p < 0.08] (Data for percentage of distance are not shown).

Effects of ASV-30 (A) and astressin (B) on the percentage of time spent in the center of an open field in comparison with diazepam (C). Time spent in either the center or the margins of the arena was measured using defined photocell arrays interfaced to software by Columbus Instruments. There were 10 to 20 mice in each group, representing a compilation of three experiments for each peptide. ∗, p values < 0.039 to 0.0078 versus vehicle.
Plus Maze.
ASV-30 (Fig.4A), astressin (Fig. 4B), and diazepam (Fig. 4C) all significantly increased the percentage of time spent in the open arms [F(5,141) = 2.5; p < 0.03], [F(5,74) = 2.4; p < 0.04], and [F(3,36) = 6.0; p < 0.002], respectively. Percentage of time in open arms was calculated as the proportion of time spent in the open arms in relation to time spent in the closed arms and the center of the maze. The MED values for ASV-30, astressin, and diazepam were 0.1 nmol (p < 0.036), 0.1 nmol (p < 0.048), and 0.5 mg/kg (p < 0.01), respectively. The percentage of open arm entries was altered in a similar manner by ASV-30 and astressin, but the overall Fvalues for this variable did not reach statistical significance for either peptide [F(5,141) = 2.05; p < 0.07] and [F(5,74) = 1.8; p < 0.11], respectively. Percentage of open arm entries for diazepam was, however, significantly increased [F(3,36) = 4.6;p < 0.008], with the MED at 0.5 mg/kg (p < 0.02). Percentage of open arm entries is calculated as the proportion of entries into the open arms relative to total arm entries. Ucn II significantly reduced percentage of time spent in the open arms of the plus maze ([F(4,94) = 3.42; p < 0.01] (Fig. 4D) and produced a nonsignificant trend toward reducing the percentage of open arm entries [F(4,94) = 2.3; p < 0.06] (data not shown). The M.E.D for the effects of Ucn II on percentage of open arm time was 0.03 nmol. Ucn II did not alter the number of closed arm entries [F(4,94) = 0.54; p < 0.7].

Effects of ASV-30 (A) and astressin (B) on the percentage of time spent in the open arms of the plus maze in comparison with diazepam (C). Percentage of time was calculated as time spent in the open arms divided by time spent in the closed arms and the center of the maze. D, percentage of time spent on the open arms of the plus maze 30 min after i.c.v. injection of Ucn II. There were 10 to 20 mice in each group, representing a compilation of three experiments for each peptide. ∗, p values < 0.04 to 0.0002 versus vehicle.
ACTH Response to Restraint Stress.
As shown in Fig.5, ACTH levels increased along with i.c.v. injection and restraint stress duration, with 10 and 30 min of restraint stress inducing significantly higher ACTH levels than no restraint stress [F(15,118) = 19.5; p< 0.0001] (post hoc p values < 0.008–0.0001 versus no restraint stress). Furthermore, ACTH levels did not return to baseline levels until 30 min after the cessation of stress (p values < 0.07–0.46 versus no stress 30 and 60 min after stress cessation). Astressin (1 nmol) significantly suppressed the stress-induced rise in ACTH at all stress durations and during the first 10 min of recovery (p values < 0.03–0.0001 versus vehicle). In contrast, ASV-30 did not alter the response to restraint stress at any duration at either dose, and did not alter the rate of recovery. ASV-30, however, did produce a slight attenuation of the ACTH response to i.c.v. injection itself at the 1-nmol dose (p < 0.05 versus vehicle); however, this effect did not increase with the higher dose of ASV-30.

Effects of ASV-30 and astressin on the ACTH response to restraint stress of different durations (2, 10, and 30 min) and the recovery from 30 min of restraint stress (10, 30, and 60 min after the cessation of restraint). Two types of comparator groups were included: a group that experienced no injection or restraint and a group that received i.c.v. injection of vehicle or peptides but no restraint stress. Peptides were administered 30 min before the onset of restraint at a dose of 1 to 10 nmol/5 μl i.c.v. There were six to nine mice in each group. ∗, p values < 0.008 to 0.0001 versus no injection; #, p values < 0.05 to 0.0001 versus vehicle (restraint stress); ■, vehicle; ▪, 1 nmol of astressin; ▧, 1 nmol of ASV-30; ▨, 10 nmol of ASV-30.
Discussion
ASV-30 reduced anxiety-like behavior to the same extent as astressin in several animal models. Similar to the SSRI fluoxetine, both peptides reduced marble burying without altering locomotor activity. Marble burying has been characterized as a model of compulsive behavior because animals bury marbles in familiar settings that have not been paired with aversive stimuli, display the behavior regardless of satiety condition, and continue to show the behavior even after repeated exposures (Njung'e et al., 1999a). Marble burying cannot be characterized, however, as a traditional anxiety task because it is not sensitive to doses of benzodiazepine agents that do not disrupt motor activity. Thus, the biological substrate for this behavior may be different from that of more traditional anxiety paradigms, such as shock-based, conflict models of anxiety. Interestingly, CRF1 receptor antagonists have also been shown to be more sensitive to “natural stress”-induced models of anxiety than to the traditional conflict paradigms (Griebel et al., 1998).
Marble-burying behavior is sensitive to the effects of SSRIs, which are effective anticompulsive therapeutics (Njung'e et al., 1999b; U.S. Department of Health and Human Services, 1999). Interestingly, CRF2 receptors are quite densely localized in the dorsal raphe nucleus (Chalmers et al., 1995), and there is evidence that CRF-immunoreactive neurons are closely associated with tryptophan hydroxylase staining neurons in the raphe (Lowry et al., 2000). Intraraphe CRF has been shown to reduce serotonin levels in the lateral septum and striatum (Price and Lucki, 2001) and to inhibit firing of dorsal raphe neurons at low doses (Kirby et al., 2000) in vivo. In addition, a very recent article demonstrated that local injection of the mixed CRF1/CRF2 antagonistd-Phe CRF (12–41) into the dorsal raphe dose dependently inhibited learned helpless behavior, whereas CRF itself enhanced learned helplessness (Hammack et al., 2002). Moreover, intraraphe Ucn II and ASV-30 reproduced the effects of CRF and d-Phe CRF (12–41), respectively, indicating that CRF2receptors are specifically involved (Maier et al., 2001). Thus, it is possible that CRF antagonists could reduce anxious or compulsive behavior by attenuating stress-induced inhibition of serotonergic activity.
As observed in the marble-burying task, ASV-30 and astressin had equivalent potency in the open-field paradigm. MED values for the open field paradigm were the same for both peptides (3 nmol), and efficacy at the MED was similar to that of the 0.5-mg/kg dose of diazepam. The dose-response function for the plus maze task, however, was quite different from either the marble-burying or open field tests. The MED for both peptides in the plus maze was much lower, at 0.1 nmol, and for ASV-30, seemed to be an all-or-none effect, rather than a stepwise or inverted U dose-response function. In contrast, diazepam produced a simple stepwise dose-response function, with an MED similar to that for the open field test (0.5 mg/kg). The basis for these differences in the dose-response functions of both peptides across anxiety tasks is not clear, possibly reflecting variability between cohorts of mice or issues with the quantitative nature of the variables in each task.
In general, however, our mouse data are in agreement with the rat data from Takahashi et al. (2001) where ASV-30 reduced anxiety-like behavior in several models of anxiety. Furthermore, Takahashi et al., (2001)also observed differences in ASV-30 potency in the plus maze task compared with defensive withdrawal (which could be viewed as a version of the open field task). Similar to Takahashi et al. (2001), our data are not consistent with Kishimoto et al. (2000), who showed that i.c.v. administration of a dose equivalent to our minimal effective dose (0.1 nmol) of ASV-30 increased anxious behavior in C57BL/6J mice using the plus maze test. One explanation for the discrepancy could be mouse strain. We used BALB/c mice, which was the strain used by another group that demonstrated anxiolytic efficacy for intraseptal ASV-30 using conditioned fear and stress-enhanced anxiety-like behavior on the plus maze task as the measures of anxiety (Radulovic et al., 1999). There is evidence that some mouse strains are more sensitive to the mixed CRF1/CRF2 receptor antagonist α-helical CRF (9–41) than others (Conti et al., 1994).
The ASV-30 data from our group, Takahashi et al. (2001), and Radulovic et al. (1999) suggest a role for the CRF2receptor in anxiety. However, it could be argued that ASV-30 exerted its anxiolytic effects partially through antagonism of CRF1 receptors because ASV-30 does inhibit sauvagine binding in CRF1-expressing cells to a certain extent (Ki > 150 nM). This explanation is doubtful because ASV-30 and astressin were equipotent as anxiolytics, even though ASV-30 is at least 100-fold less potent than astressin as an inhibitor of CRF1receptor binding. Furthermore, ASV-30 did not alter either the magnitude or the recovery rate of the ACTH response to restraint stress, a function that is presumed to be exclusively attributed to CRF1 receptor activation. In contrast, astressin had extremely marked effects on the magnitude and the recovery of this HPA response to stress when tested in parallel with ASV-30. Taken together, these data do suggest that the anxiolytic effects of ASV-30 were not mediated by the CRF1 receptor.
The data obtained by ourselves and others with ASV-30, which indicate an anxiolytic effect of CRF2 receptor antagonism, are also complimented by recent antisense studies. Ho et al. (2001)have shown that central administration of CRF2-specific antisense oligonucleotides reduced anxiety in the conditioned freezing task, paralleled by a drop in CRF2 receptor number by 70% in animals showing reduced anxiety-like behavior. In addition, central administration of the CRF2 receptor-selective agonist Ucn II dose dependently increased anxiety-like behavior in the plus maze task, reducing both percentage of time and entries into open arms. Although it could be argued that these “anxiogenic” effects of Ucn II could be related to reduced locomotor activity, the number of closed entries was not altered by the peptide. All of the above-mentioned evidence is consistent with the idea that antagonism of CRF2 receptors acts to attenuate anxiety or stress-like behaviors.
The hypothesis that antagonism of CRF2 receptors acts to reduce anxiety is not consistent with the overall profile of increased sensitivity to stress suggested by the phenotype of the CRF2 knockout mice. It could be argued that the phenotype of the CRF2 knockout mice was the result of developmental compensation for the deletion of the CRF2 receptor. Indeed, one group reported elevations in Ucn and CRF in the Edinger-Westphal nucleus and the central nucleus of the amygdala (Bale et al., 2000), and suggested that these changes could account for the increased sensitivity to stress observed in these mice. Because the HPA axis response to stress seemed normal to hypersensitive, it is possible that heightened CRF1-mediated HPA activation could at least partially explain the complex behavioral phenotype of these mice.
Exactly how central CRF1 and CRF2 receptors interact to mediate behavioral responses to stress is not well understood. Although the CRF1 receptor seems to be primarily involved in the HPA axis response to stress, its involvement in the extra-pituitary aspects of arousal is not clear. For example, although the CRF1-selective antagonist antalarmin did inhibit i.c.v. CRF-induced blood pressure elevation (Briscoe et al., 2000), another CRF1-selective antagonist, CP154,526, only partially attenuated the effects of i.c.v. CRF on heart rate and plasma catecholamines (Nijsen et al., 2000). Furthermore, CRF2 receptors are quite densely distributed in brain areas associated with the central control of autonomic nervous system function, such as the ventromedial hypothalamus, the lateral septal area, the BNST, and the posterior amgydala (Miyazawa et al., 1988; Saito et al., 1989; Chalmers et al., 1995; Wilkinson and Pittman, 1995). Interestingly, microinjection of CRF into the BNST increases heart rate and cardiac PQ interval, whereas microinjection of the mixed CRF antagonist α-helical CRF (9–41) attenuates conditioned freezing and the stress-induced changes in cardiac PQ interval associated with freezing (Nijsen et al., 2001). The above-mentioned evidence could suggest that central CRF2 and CRF1 receptors may act to integrate the autonomic nervous system with the HPA axis, respectively, under stressful conditions. Future studies with selective CRF2receptor agonists and antagonists, such as Ucn II and ASV-30 should clarify the roles of the two receptor subtypes in the stress response.
In summary, ASV-30 reduced anxiety-like behavior in three models of anxiety (marble burying, open field, and plus maze) without producing significant changes in locomotor activity or altering the ACTH response to restraint stress. Furthermore, the effects of ASV-30 were equivalent to those of the potent mixed CRF1/CRF2 antagonist astressin, and between 50 and 100% of the magnitude of non-CRF agents, such as fluoxetine and diazepam. In addition, the potent and selective CRF2 agonist Ucn II dose dependently increased anxiety-like behavior. Taken together, these data argue that central CRF2 receptors may play a role in the behavioral, but not the endocrine response to stress.
Acknowledgments
We are grateful to Dr. George Koob for helpful comments in the preparation of this manuscript.
Footnotes
-
↵1 Present address: Department of Metabolic Research, Bristol-Meyers Squibb, 311 Pennington-Rocky Hill Rd., Pennington, NJ 08543.
-
This work was supported in part by a Small Business Innovation Research Grant 1 R43 MH65106-01 from the National Institute of Mental Health.
- Abbreviations:
- CRF
- corticotropin-releasing factor
- BNST
- bed nucleus of the stria terminalis
- HPA
- hypothalamic-pituitary adrenal
- ASV-30
- antisauvagine-30
- Ucn II
- urocortin II
- SSRI
- selective serotonin reuptake inhibitor
- ACTH
- adrenocorticotropic hormone
- ANOVA
- analysis of variance
- MED
- minimal effective dose
- CP154,526
- butyl-[2,5-dimethyl-7-(2,4,6-trimethylphenyl)-7H-pyrrolo[2,3-d]pyrimidin-4-yl]ethylamine
- Received January 10, 2002.
- Accepted March 1, 2002.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}