

Abstract
Perigeniculate neurons form an interactive sheet of cells that inhibit one another as well as thalamocortical neurons in the dorsal lateral geniculate nucleus (LGNd). The inhibitory influence of the GABAergic neurons of the perigeniculate nucleus (PGN) onto other PGN neurons was examined with intracellular recordings in vitro. Intracellular recordings from PGN neurons during the generation of spindle waves revealed barrages of EPSPs and IPSPs. The excitation of local regions of the PGN with the local application of glutamate resulted in activation of IPSPs in neighboring PGN neurons. These IPSPs displayed an average reversal potential of −77 mV and were blocked by application of bicuculline methiodide or picrotoxin, indicating that they are mediated by GABAA receptors. In the presence of GABAA receptor blockade, the activation of PGN neurons with glutamate could result in slow IPSPs that were mediated by GABAB receptors in a subset (40%) of cells. Similarly, application of specific agonists muscimol and baclofen demonstrated that PGN neurons possess both functional GABAAand GABAB receptors. Examination of the axon arbors of biocytin-filled PGN neurons often revealed the presence of beaded axon collaterals within the PGN, suggesting that this may be an anatomical substrate for PGN to PGN inhibition.
Functionally, activation of inhibition between PGN neurons could result in a shortening or a complete abolition of the low threshold Ca2+ spike or an inhibition of tonic discharge. We suggest that the mutual inhibition between PGN neurons forms a mechanism by which the excitability of these cells is tightly controlled. The activation of a point within the PGN may result in the inhibition of neighboring PGN neurons. This may be reflected in the LGNd as a center of inhibition surrounded by an annulus of disinhibition, thus forming a “center-surround” mechanism for thalamic function.
The thalamic reticular and perigeniculate nuclei both form sheets of interconnected GABAergic neurons that affect the excitability and pattern of activity generated within the thalamus and therefore in nearly all thalamocortical activity (for review, see Steriade and Deschênes, 1984; Jones, 1985). These GABAergic neurons are densely innervated by collaterals from thalamocortical and corticothalamic axons as they pass through the thalamic reticular and perigeniculate nuclei. Perigeniculate neurons innervate thalamocortical cells in laminae A, A1, and perhaps C of the cat and ferret dorsal lateral geniculate nucleus (LGNd) as well as give rise to axonal and dendrodendritic connections to other perigeniculate cells (Ide, 1982; Deschênes et al., 1985; Cucchiaro et al., 1991;Uhlrich et al., 1991; Bal et al., 1995a,b). Although specific roles for perigeniculate and thalamic reticular inhibition of thalamocortical cells in the control and generation of thalamocortical activity during sleep have been demonstrated (Steriade et al., 1985; Mulle et al., 1986; Bal et al., 1995a,b), the role of intraperigeniculate inhibition has been less clear.
Recently, in vitro studies of spindle wave generation in the ferret geniculate slice have suggested that intra-perigeniculate nucleus (PGN) inhibition may play an important role in the pattern and strength of activity generated in these circuits (von Krosigk et al., 1993; Bal et al., 1995a,b). The global block of GABAAreceptors in ferret geniculate slices results in the transformation of normal spindle waves into slow 2–3 Hz synchronized oscillations (Bal et al., 1995a,b) that resemble in some aspects that which occurs in some animal models of generalized absence seizures (Gloor et al., 1990). This transition was proposed to occur, in part, through the disinhibition of PGN cells from one another, thereby allowing these cells to strongly discharge with each cycle of the oscillation. The strong discharge of PGN cells was proposed to give rise to the strong activation of postsynaptic GABAB receptors in thalamocortical cells, which is an essential step in the generation of these abnormal oscillations (Bal et al., 1995a,b).
Previous investigations of GABAergic neurons in the rodent thalamic reticular nucleus reveal that these cells exhibit typical GABAA receptor-mediated increases in Cl− conductance in response to the exogenous application of GABA (McCormick and Prince, 1986) or to the activation of GABAergic axons in the local neuropil with electrical stimulation (Ulrich and Huguenard, 1995, 1996). During the generation of spindle waves, perigeniculate and thalamic reticular neurons receive barrages of EPSPs (Mulle et al., 1986; Bal et al., 1995a,b). These EPSPs arise from burst firing in thalamocortical neurons and the duration of each EPSP barrage is typically shortened and the amplitude decreased by the arrival of IPSPs resulting from the burst firing of neighboring PGN cells (Bal et al., 1995b). Here we have investigated the physiological and functional properties of intra-PGN inhibition and demonstrate that these IPSPs are largely mediated by the activation of GABAAreceptors, although they can also activate GABAB receptors and are functionally important for determining the pattern of discharge within the PGN.
Some of these results have been published previously in abstract form (Sanchez-Vives and McCormick, 1996).
MATERIALS AND METHODS
The methods for slice preparation, drug application, and intracellular recording are given in the accompanying article (Sanchez-Vives and McCormick, 1997) and in other publications (Bal et al., 1995a). Intracellular recording electrodes were formed on a Sutter Instruments P-80 micropipette puller from medium-walled glass (1BF100; WPI) and beveled on a Sutter Instruments beveler. Micropipettes were filled with 1.5–2.0 m potassium acetate and 1.5–2.0% biocytin for intracellular labeling of recorded neurons and had resistances of between 60 and 100 MΩ. Biocytin-filled neurons were visualized through standard avidin–biotin–horseradish peroxidase reaction with diaminobenzidine (Horikawa and Armstrong, 1988). The dendritic and axonal arbors of perigeniculate neurons were examined and photographed with Nomarski optics with 63× and 100× oil immersion lenses. Data are presented as mean ± SD.
RESULTS
Perigeniculate neurons inhibit one another through GABAA and GABAB receptors
Intracellular recordings were obtained from 129 perigeniculate neurons. A representative sample of these cells revealed a resting membrane potential of −64 mV (±6; mean ± SD; n= 10) and an apparent input resistance, as measured with 0.2–0.4 nA hyperpolarizing current pulses at rest, of 98 MΩ (±33). During the generation of spindle waves in vitro, PGN neurons often received intermixed EPSP–IPSP barrages (Fig.1A), as reported previously (Bal et al., 1995b). In most PGN cells the PSP barrages were dominated by EPSPs arising from burst discharges of thalamocortical cells, with IPSPs being visible after substantial depolarization from resting membrane potentials (Bal et al., 1995b). However, in many PGN cells (22 of 62), IPSPs were also visible at membrane potentials between −65 and −55 mV during the generation of spindle waves (Fig.1A). As reported previously (Bal et al., 1995b), these IPSPs were typically preceded by a barrage of EPSPs and therefore took the form of EPSP–IPSP sequences (Fig. 1A).

Perigeniculate neurons receive barrages of IPSPs spontaneously and during the generation of spindle waves.A, Intracellular recording from a PGN neuron during the generation of a spindle wave. The IPSPs in this PGN neuron were especially prominent. These IPSPs could appear as purely hyperpolarizing or could be preceded by a barrage of EPSPs at −65 mV. Hyperpolarization of the PGN neuron to −86 mV with the intracellular injection of current resulted in an abolition or reversal of the IPSPs.B, Intracellular recording from a PGN neuron during the spontaneous occurrence of IPSPs at ∼1.5 Hz (Vm = −71 mV). Expansion of these IPSPs reveal that they are composed of four to six individual IPSPs arriving at 180–350 Hz and could be preceded by a barrage of EPSPs.C, Examples of barrages of IPSPs that spontaneously occur in PGN neurons and presumably result from the burst discharge of single PGN neurons.
On occasion, IPSP barrages not only occurred during the generation of spindle waves, but also spontaneously in rhythmic barrages at 1–2 Hz, as if generated by a rhythmically bursting PGN neuron (Amzica et al., 1992; Bal and McCormick, 1993). In contrast to the IPSPs generated during spindle waves, these spontaneous IPSP barrages often lacked any detectable EPSP component (Fig. 1B). Close examination of the spontaneous IPSP barrages revealed that they consist of a high frequency barrage of 2–8 IPSPs of 0.3–2 mV amplitude at 180–350 Hz (Fig. 1B,C), suggesting that these result from the discharge of a single PGN cell.
Intracellular recordings from perigeniculate neurons while glutamate was being applied to local regions of the PGN revealed that activation of restricted regions of this nucleus, typically within 50–250 μm from the recorded PGN cell, resulted in the generation of IPSPs (n = 39) (Fig.2A). These evoked IPSPs often took the form of barrages of IPSPs that varied in amplitude from just detectable (0.2 mV) up to 2 mV in amplitude (Fig.2A). Examination of the inter-IPSP frequency during the generation of these IPSP barrages revealed a steady decrease in frequency from ∼650 to 100 Hz (Figure 2B). Interestingly, this sequence of frequencies is similar to that generated by single PGN neurons in response to the intracellular injection of a depolarizing current pulse in which the PGN cell generated burst, followed by tonic firing (Fig. 2C,D). The similarity of PGN action potential discharge and IPSP generation in response to local glutamate application further supports the hypothesis that these IPSPs are mediated by the local activation of PGN neurons. Activation of these IPSPs while the PGN neuron was depolarized or hyperpolarized to different membrane potentials revealed an average reversal potential of −76.6 mV (±4.1 mV; n = 13) (Fig. 3A,B), which is significantly more depolarized than the reversal potential of GABAA receptor-mediated IPSPs in thalamocortical neurons (−83.3 ± 3.6 mV; n = 9; t = 3.95; p < 0.001) (Sanchez-Vives and McCormick, 1997).

Comparison of interburst frequencies for evoked IPSP barrages and intrinsic burst discharges in single PGN neurons.A, Compound IPSPs induced by glutamate applied locally to the PGN. Individual IPSPs can be distinguished and are indicated byarrows in the expanded trace. B, The initial IPSPs arrive at ∼650 Hz, but this frequency decreases steadily to ∼100 Hz. C, Interspike frequencies in a single PGN neuron generating a low threshold Ca2+spike mediated burst followed by tonic activity in response to the intracellular injection of a depolarizing current pulse.D, Overlap of the frequencies for IPSP and PGN spike generation demonstrating the similarity in these two distributions.

Reversal potential of intra-PGN IPSPs.A, Local application of glutamate (Glu) in the PGN evokes a barrage of IPSPs in a neighboring PGN cell. Each compound IPSP is composed of several presumed unitary IPSPs that are <1 mV in amplitude. This evoked IPSP barrage reverses at approximately −75 mV. B, Graphic representation of the reversal potential of IPSPs evoked in five different PGN cells (each in a different slice). C, Schematic illustration of the recording arrangement and axonal and dendritic connections between PGN cells and other PGN and thalamocortical neurons.
Local application of tetrodotoxin (10 μm in micropipette) to block action potential-mediated release of GABA typically abolished glutamate-evoked IPSPs, indicating that they rely on the generation of Na+-dependent action potentials in other PGN cells (Fig. 4A) (n = 4 of 6). In addition, spontaneous or glutamate-evoked IPSPs were blocked or substantially reduced by local application of the GABAA channel blocker picrotoxin (n = 3; 500 μm in micropipette) (Fig.4B) and the GABAA receptor antagonists bicuculline methiodide (n = 11; 200–400 μm in micropipette) (Fig. 4A) and SR 95531 (n = 1; 500 μm in micropipette) (Fig. 4C).

Lateral inhibition in the PGN is mediated largely through the activation of GABAA receptors.A, Local application of glutamate (Glu) in the PGN results in the activation of fast IPSPs followed by barrages of EPSPs associated with the beginning of a spindle wave (complete spindle wave not shown). The block of generation of Na+-dependent action potentials with the local application of tetrodotoxin (10 μm in micropipette) blocks the IPSPs. After recovery of the evoked IPSP, local application of bicuculline (400 μm in micropipette) completely blocks this hyperpolarizing event. The bottom trace inA illustrates that this PGN neuron receives excitatory input from laminae A1. B, Application of glutamate in the PGN results in IPSPs that are completely blocked by local application of picrotoxin (500 μm in micropipette).C, Application of the GABAA receptor antagonist SR95531 (500 μm in micropipette) inhibits IPSPs evoked in a PGN neuron by application of glutamate in the PGN. This neuron generated spontaneous oscillations before and after application of SR95531. D, Schematic illustration of the recording and drug-applying locations. The application of glutamate (glu) in the PGN activated IPSPs in PGN cells as well as initiated spindle waves, which resulted in the occurrence of repetitive barrages of EPSPs and IPSPs in PGN cells. Only the beginning of these spindle waves are illustrated. Results from three different cells are illustrated in A, B, and C.TTX, Tetrodotoxin; Bic, bicuculline;PTX, picrotoxin; SR, SR95531.
In 6 of the 15 PGN cells a slow component evoked by local glutamate application remained after the application of one of these GABAA receptor antagonists. This slow IPSP was blocked by local application of the GABAB receptor antagonist CGP 35348 (2 mm in micropipette) (Fig.5), suggesting that inhibition between perigeniculate cells is mediated by both GABAA and GABAB receptors.
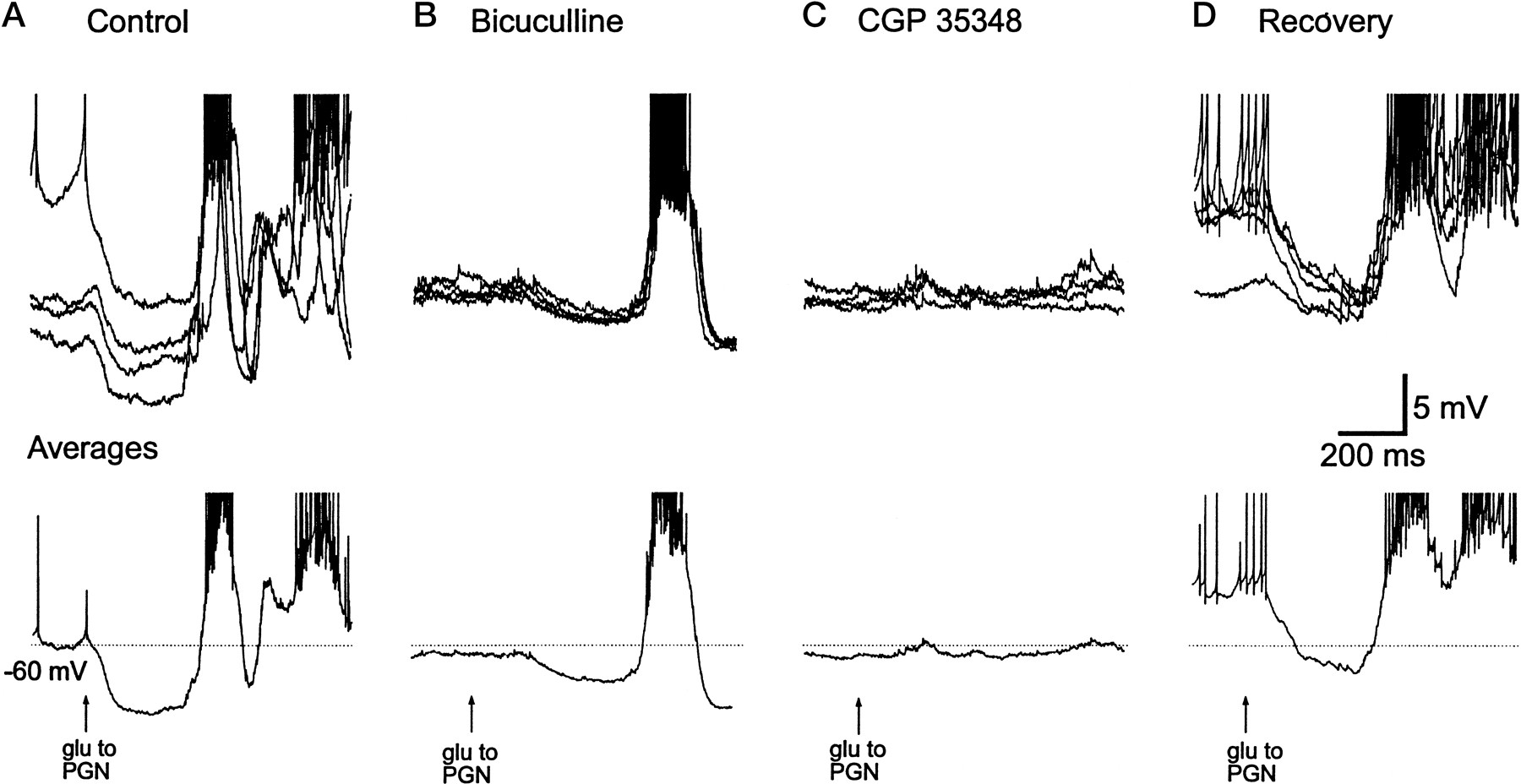
Lateral inhibition in the PGN can activate GABAB receptors. A, Local activation of the PGN with glutamate (glu) evokes IPSPs and a spindle wave in this PGN neuron. B, Local application of bicuculline (400 μm in micropipette) reduces but does not completely block this evoked IPSP. C, Local application of CGP35348 (2 mm in micropipette) blocks the residual IPSP, indicating that it was mediated by GABAB receptors.D, Recovery of the evoked IPSPs after washing out the GABA receptor blockers. In A–D, the top traces show overlapped four to five different applications of glutamate, whereas the bottom traces represent the average. Action potentials have been truncated. The membrane potential of this PGN cell varied between applications in control and recovery owing to the bi-stable nature of PGN cell activity.
The possibility that PGN neurons possess GABAB as well as GABAA receptors was further investigated with local application of specific agonists and antagonists. Application of GABA to PGN neurons often resulted in a depolarizing response (Fig.6A). Local application of acetazolamide (200 μm in micropipette) blocked this depolarizing response, suggesting that it is mediated by a GABA-activated bicarbonate conductance (Staley et al., 1995) (Fig.6A). After the application of acetazolamide, the application of GABA to PGN neurons resulted in two distinct phases of hyperpolarization (Fig. 6B). In this cell, the initial, rapid phase exhibited a reversal potential of −83 (Fig.6B). Local application of picrotoxin reduced this rapid hyperpolarizing response and revealed a slow hyperpolarization that reversed at the more negative membrane potential of −100 mV (Fig.6B). In the presence of picrotoxin, increasing the size of GABA applications increased the amplitude and duration of the slow inhibition (Fig. 6C). These results suggest that PGN neurons possess both GABAA and GABAB receptors. This hypothesis was tested further with the local application of the specific GABAA receptor agonist muscimol and the GABAB receptor agonist baclofen.

Perigeniculate neurons possess both GABAA and GABAB receptors. Local application of GABA to a perigeniculate cell results in the generation of a depolarizing response. Local application of acetazolamide (200 μm in micropipette) blocks the depolarizing response and reveals an underlying hyperpolarization that has distinct rapid and slow phases. B, The rapid phase reverses at approximately −83 mV and is blocked by picrotoxin. After the application of picrotoxin, the slow phase remains and exhibits a reversal potential of −95 mV. C, In the presence of picrotoxin, slowly increasing the application of GABA results in progressive increases in the amplitude of the slow hyperpolarization.
The application of muscimol (n = 4; 100 μm in micropipette) at membrane potentials of −60 to −70 mV could evoke depolarizing responses in PGN neurons (Fig.7A). Local application of acetazolamide (200 μm in micropipette) blocked these depolarizing responses and revealed muscimol-induced hyperpolarizing responses (Fig. 7A), which were completely blocked by local application of picrotoxin (not shown). Local application of the GABAB receptor agonist baclofen (200 μm in micropipette) to PGN neurons resulted in a prolonged hyperpolarization, an inhibition of action potential discharge, and an increase in apparent input conductance (n = 4) (Fig.7B). Local application of CGP35348 (2 mm in micropipette) reversed these effects (n = 2), indicating that they were mediated through activation of GABAB receptors (Fig. 7B).

Actions of the specific agonists muscimol and baclofen on PGN cells. A, Repeated, local application of the GABAA receptor agonist muscimol to a PGN cell evokes depolarizing responses at −72 mV. The local application of acetazolamide (200 μm in micropipette) results in an abolition of these depolarizing responses and the appearance of hyperpolarizing responses to muscimol. B, Local application of the GABAB receptor agonist baclofen (200 μm in micropipette) results in a prolonged inhibition and increase in membrane conductance. Local application of CGP35348 (2 mm in micropipette) results in a reversal of these effects.
Presynaptic GABAB receptors inhibit thalamocortical EPSPs
Previous studies have demonstrated that glutamatergic transmission may be reduced through the activation of presynaptic GABAB receptors (Thompson et al., 1993; Emri et al., 1996; Isaacson and Hille, 1997). To test the possibility that thalamocortical EPSPs in the ferret PGN may be regulated by GABAB receptors, we activated EPSPs or EPSCs in PGN neurons through the local application of glutamate in the A1-lamina of the LGNd (Fig. 8C). In addition, to reduce the confounding influences of postsynaptic increases in K+ conductance in response to activation of GABAB receptors, these experiments were performed with microelectrodes containing 2 m CsAc and 25 mmQX-314. Under these conditions, the activation of GABABreceptors with local application of baclofen (100–200 μmin micropipette) in the PGN resulted in an average reduction of EPSC amplitude by 40.3% (±6.2%; n = 7) (Fig.8A,B). The local application of CGP 35348 resulted in an immediate reversal of these inhibitory influences of GABAB receptor activation (Fig. 8A,B). Recording of multiple unit activity near the site of glutamate application did not reveal a detectable decrease in activated discharge (n = 4), indicating that these reductions in EPSP/EPSC amplitude were unlikely to be the result of inhibition of thalamocortical neurons by the local application of baclofen in the PGN (Fig. 8A,B).

Presynaptic GABAB receptors regulate the amplitude of excitation from thalamocortical onto PGN neurons.A, EPSPs recorded in a PGN cell evoked by local application of glutamate (glu) in lamina A1 (Vm = −75 mV). The local application of baclofen (200 μm in micropipette) results in a 41% reduction in the amplitude of these evoked EPSPs. In contrast, multiple unit activity recorded at the site of glutamate application is unaffected by the application of baclofen in the PGN. The local application of CGP 34348 (2 mm in micropipette) reverses this action of baclofen. B, EPSCs recorded in a PGN cell and induced by glutamate application in A1 lamina (Vholding = −65 mV). In the top traces, four to five superimposed responses of different glutamate applications are shown. The middle traces are the average of the ones above. The simultaneous extracellular recordings of the glutamate activated area of the LGNd are shown in thelowest traces. Local application of baclofen results in a reduction of EPSC amplitude, an effect that is reversed by the local application of CGP 35348. C, Schematic illustration of the recording arrangement used for A andB. All the recordings shown in this figure were performed with 2 m cesium acetate and 25 mm QX 314 in the intracellular electrode to block the activation of outward currents in response to baclofen.
Intra-PGN inhibition controls the duration of burst discharges
Local application of glutamate in lamina A1 typically activated a barrage of EPSPs in PGN neurons (Fig.9A,B). These EPSPs could then activate a low threshold Ca2+ spike and burst of 1–10 action potentials. Sometimes these EPSPs were followed by disynaptic IPSPs as a result of indirect activation of PGN cells (Fig.9A). Similarly, local application of glutamate in the PGN could result in direct depolarization of the PGN cell, which was followed by a brief hyperpolarizing phase (Fig. 9C,D). The local or bath application of GABAA receptor antagonists bicuculline (n = 10; 400 μm in micropipette), penicillin-g (n = 2; 100 mmin micropipette), picrotoxin (n = 7, 500 μm in micropipette, 20–50 μm in bath), or SR 95531 (n = 4, 500 μm in micropipette, 50 μm in bath) to the PGN resulted in a marked prolongation of the bursts of action potentials evoked indirectly through activation of thalamocortical cells (Fig. 9B) as well as those evoked with local application of glutamate (Fig.9C,D). These results indicate that the activation of GABAA receptors in the PGN strongly influences the duration and intensity of burst discharges in PGN neurons.

Lateral inhibition in the PGN controls burst firing in these cells. A, Application of glutamate in lamina A1 while recording intracellularly from a PGN cell results in a barrage of EPSPs that is interrupted by a burst of IPSPs in the PGN neuron. At −61 mV, the PGN neuron is excited and inhibited by the barrage of PSPs. At −73 mV, the PSP barrage initiates a low threshold Ca2+ spike and burst of action potentials. At −79 mV, both the EPSPs and IPSPs are clearly visible. At −92 mV, the IPSPs are abolished or reversed. The bottom trace is an intracellular recording of a thalamocortical neuron in the region of local glutamate application in lamina A1. B, Local application of glutamate (glu) in lamina A1 results in a barrage of EPSPs that activate a low threshold Ca2+ spike in this PGN neuron (Vm = −78 mV). The local application of bicuculline to this cell results in a marked prolongation of the burst of action potentials. C, D, In two other cells, application of glutamate in the PGN induces direct activation, but also activation of the neighboring cells, which induces a brief period of inhibition (double arrows) and the generation of another burst of action potentials. Local application of either bicuculline (C) (400 μm in micropipette;Vm = −76 mV) or penicillin g (D) (100 mm in micropipette;Vm = −73 mV) results in an abolition of the brief period of inhibition, indicating that it was mediated through the activation of GABAA receptors.
The possibility that lateral inhibition within the PGN may control the amplitude and duration of low threshold Ca2+ spikes in PGN cells was examined by activating IPSP barrages in PGN neurons that are actively involved in generating low threshold Ca2+ spikes (Fig.10). Pressure-pulse application of glutamate in the PGN activated a barrage of IPSPs in the recorded PGN cell (Fig. 10A). Intracellular injection of a hyperpolarizing current pulse resulted in a typical rebound low threshold Ca2+ spike (Fig. 10B). Activating the barrage of IPSPs from the PGN to this neuron at different times during the generation of the low threshold Ca2+ spike demonstrated that these IPSPs can control the amplitude and duration of the rebound Ca2+ spike (Fig. 10C). Activation of the IPSPs on the falling phase of the low threshold Ca2+ spike resulted in a shortening of the duration of this event (Fig. 10C). As the IPSPs and the Ca2+ spike overlapped more and more in time, the amplitude of the low threshold Ca2+ spike was markedly diminished, until it was almost completely inhibited by the PGN-generated IPSPs (Fig. 10C, bottom trace).

The activation of IPSPs in PGN cells can control the amplitude and duration of low threshold Ca2+spikes in PGN neurons. A, The local application of glutamate (glu) in the PGN evokes a barrage of IPSPs in this PGN cell after the injection of a hyperpolarizing current pulse. This cell was recorded with a microelectrode containing 50 mm QX-314 in 1.2 m potassium acetate to block the generation of action potentials. In addition, CNQX (10 μm) was included in the bath to block the activation of EPSPs (Vm = −70 mV). B, Increasing the amplitude of the current pulse results in the generation of a rebound low threshold Ca2+ spike.C, The timing between the injection of the hyperpolarizing current pulse and the activation of the IPSPs is varied. Shortening the delay between the onset of the low threshold Ca2+ spike and the activation of the IPSPs to increase the overlap between these two events resulted in a marked reduction in the amplitude and duration of the rebound Ca2+ spike. The full amplitude of the hyperpolarizing electrotonic response to the current pulse is not shown for illustrative purposes. D, Schematic diagram illustrating recording arrangement.
Perigeniculate neurons give rise to axon collaterals within the PGN
Visualization of the axon arbors of PGN neurons with intracellular injection of biocytin revealed a dense innervation of the LGNd (laminae A, A1, and C) as well as, in several cases (18 of 32 cells), axon collaterals within the borders of the PGN (Figs.11, 12). The innervation of LGNd often was not uniform, in that one of the laminae (A, A1, or C) was more densely innervated than the other. Indeed, in many instances there appeared to be laminations within the A-laminae, perhaps corresponding to the “ON” and “OFF” zones of the ferret LGNd (Stryker and Zahs, 1983) (not shown). In the dorsal-ventral plane of the sagittal slices, individual PGN fibers typically covered a distance of between 153 and 666 μm (average of 350 ± 120 μm) in the A-laminae (n = 31) (Fig. 11A,B). Within the PGN, axon collaterals were typically observed to travel 100–650 μm in the dorsal-ventral plane before entering the LGNd (n= 8) (Fig. 12A). Of 18 filled PGN cells with axon collaterals within the PGN, 14 gave rise to collaterals that stayed within the dendritic arbor of the filled cell, whereas the axon collaterals of the remaining four extended beyond this region. The intra-PGN axon collaterals exhibited frequent boutons, approximately one every 2–30 μm, thereby giving rise to many putative en passant synaptic contacts (Figs.11D–G; 12B,E,F). Individual “boutons” were typically ∼1 μm in diameter and ranged from 0.5 to 2.2 μm. Examination of PGN axon collaterals within the A-laminae, or within the interlaminar regions, revealed similar characteristics concerning both the size of the boutons and the distance between them (Figs. 11, 12C,H). The number of boutons within the PGN varied from a few to several hundred. The collateral shown in Figure 12A has 20 boutons, whereas the axons shown in Figure 11A–C gave rise to several hundred putative synaptic contacts within the PGN.

Axon collaterals from perigeniculate cells densely innervate both the LGNd and the PGN. A, Dark-field photomicrograph of a section that contains the axonal innervation made by a single PGN cell filled with biocytin. The axonal arbor generates a column ∼300 μm in diameter through laminae A and A1. The cell body is not in this section. The top white arrowhead illustrates the location of the higher power photomicrograph in D, whereas the bottom white arrowhead corresponds to that in E.B, Dark-field photomicrograph of the axonal arbor from a different PGN neuron. Again, the axonal arbor appears as a column ∼250 μm in diameter throughout the PGN and A-laminae. In this cell, there appears to be an increased density of boutons in the interlaminar zone as we have described previously (Sanchez-Vives et al., 1996). Thetop white arrowhead corresponds to the location of the higher power photomicrograph in F, whereas the lower is that illustrated in G. C, Higher magnification of the cell shown in B. The beaded axonal arbors are present both in the LGNd and PGN and are similar in appearance. D, Higher power photomicrograph with Nomarski optics of boutons in the PGN from the cell inA. E, Synaptic boutons in lamina A from the cell in A. F, Synaptic boutons in the PGN for the cell in B and C.G, Synaptic boutons in lamina A for the cell inB and C. Histological sections were 60–100 μm thick.

Perigeniculate cells give rise to axon collaterals and putative synaptic boutons within the PGN.A, Biocytin-filled PGN cell. The axon gives rise to a collateral at 160 μm from the soma (arrow labeled B).B, Higher magnification of the axon collateral in the PGN. The arrow is pointing to the same location as inA. Approximately 20 boutons are contained on this collateral. Two higher power photographs of the indicated regions of the axon collateral are illustrated. C, Higher power photograph of the axonal innervation of lamina A by the cell shown inA. Note that most of the putative synaptic contacts are made en passant. D, A biocytin-filled PGN cell, the cell body of which is located on the border with lamina A. This cell gives rise to two axon collaterals in the PGN: one at 122 μm (labeledE) and another at 420 μm (labeled G). Additional collaterals in the PGN were also seen in other sections (not shown). The arrows marked with E, F, G, and H correspond to the same locations pointed out by the corresponding arrows in E, F, G, andH at higher power. E, This collateral in the PGN forms a string of boutons and extends over 245 μm.F, Another collateral in the PGN. G, Higher power view of a few boutons formed by the collateral in the more dorsal portion of the section, as indicated in D.H, Higher power photomicrograph of the boutons in lamina A. These boutons appear similar to those in the PGN. OR, Optic radiation; PGN, perigeniculate nucleus.
DISCUSSION
Previous studies of the GABAergic neurons of the thalamic reticular nucleus have demonstrated that they exhibit functional GABAA receptors that when activated result in increases in membrane Cl− conductances (McCormick and Prince, 1986; Bal et al., 1995b; Ulrich and Huguenard, 1995). In addition, recent studies by Ulrich and Huguenard (1996) have demonstrated that activation of the local neuropil in the rodent thalamic reticular nucleus with electrical stimulation can activate GABAA and occasionally GABAB receptor-mediated IPSPs. Presumably these IPSPs arise from the activation of axon collaterals from other thalamic reticular neurons; however, because these investigators used electrical stimulation to evoke these events, it is also possible that they originate from axons of extrathalamic sources, such as the substantia nigra reticulata or the basal forebrain and other forebrain structures (Jourdain et al., 1989; Pare et al., 1990; Asanuma, 1994). Indeed, recently Pinault et al. (1995, 1997) suggested, on the basis of morphological studies, that axonal synaptic interactions between thalamic reticular cells in the adult rat are rare.
Our results clearly demonstrate, with local activation of perigeniculate GABAergic cells with glutamate, that these neurons exert potent inhibitory influences on one another, typically through the activation of GABAA receptors but also occasionally through the activation of GABAB receptors. As in thalamocortical and many other cell types, the activation of GABAAreceptors results in hyperpolarizing responses through increases in membrane Cl− conductance, whereas the activation of GABAB receptors results in a slower IPSP mediated through an increase in K+ conductance. In addition, PGN neurons may also exhibit depolarizing responses to GABA and GABAA receptor agonists, presumably owing to GABAA receptor-mediated increases in bicarbonate conductance (Staley et al., 1995), because this effect was blocked by acetazolamide. The synaptic activation of depolarizing GABAA receptor-mediated IPSPs in PGN neurons has not yet been demonstrated.
Previous morphological and ultrastructural examination of neurons in the cat PGN has demonstrated both axon collaterals as well as dendrodendritic synaptic contacts that presumably provide the physical substrate for the inter-PGN inhibitory influences that we have demonstrated here (Ide, 1982; Uhlrich et al., 1991). Similarly, we observed in 56% of filled PGN cells axon collaterals within the PGN. In the rodent thalamic reticular nucleus, axon collaterals and interthalamic reticular neuron inhibitory influences have been observed in young animals (Spreafico et al., 1988; Cox et al., 1996; Ulrich and Huguenard, 1996). However, a recent morphological and ultrastructural study of thalamic reticular neurons in the adult rat have failed to observe local axon collaterals forming synaptic contacts with other thalamic reticular neurons, although dendrodendritic synapses are observed (Pinault et al., 1997). These results suggest that the precise physical substrate of the PGN to PGN cell inhibition demonstrated here may require closer examination; it will be particularly important to determine the postsynaptic targets of intra-PGN axon collaterals.
Interestingly, in situ hybridization studies of the localization of the mRNA for different GABAA subunits in the rat and monkey thalamus have revealed heterogeneous distributions of each of these, with the sensory relay nuclei exhibiting prominent levels of α1/β2,3/γ2subunits and the thalamic reticular nucleus exhibiting γ2, and perhaps α3, subunits only (Fritschy and Möhler, 1995; Huntsman et al., 1996). These results suggest that the GABAA receptors in thalamic reticular/perigeniculate nuclei are different from those on thalamocortical neurons. GABAA receptor-mediated IPSCs in thalamic reticular cells have a more prolonged duration than those in thalamocortical cells (Ulrich and Huguenard, 1995), which is also seen on the single-channel level as more prolonged duration of single-channel open times in thalamic reticular neurons (Kang et al., 1996). The relationship between these properties of GABAergic inhibition and the functional expression of the various GABAergic receptor subunits remain to be explored.
In our studies (Sanchez-Vives and McCormick, 1996, 1997) and others (Ulrich and Huguenard, 1996), the most notable difference between PGN to thalamocortical inhibition and PGN to PGN inhibition is in the degree to which these pathways activate GABABreceptor-mediated IPSPs. Strong activation of perigeniculate or thalamic reticular nuclei typically activates GABABreceptor-mediated IPSPs in thalamocortical cells (Huguenard and Prince, 1994; Warren and Jones, 1994; Bal et al., 1995a,b), whereas these are less common and more difficult to evoke in PGN and thalamic reticular cells (Ulrich and Huguenard, 1996; present study). Perigeniculate- or thalamic reticular-evoked GABAA receptor-mediated IPSPs in thalamocortical neurons do seem to have a more negative reversal potential than those found in PGN neurons (average of −83 vs −77 mV), although this may be attributable to varying setpoints for Cl− homeostasis in the two cell types (Ulrich and Huguenard, 1997).
Functional consequences of intra-PGN inhibition
Functionally, intra-PGN inhibition strongly controls the excitability and pattern of activity generated by PGN cells and thus the pattern of activity generated throughout thalamocortical circuits. Perigeniculate and thalamic reticular cells generate action potentials in two distinct firing modes: single spike activity and low threshold Ca2+ spike-mediated high-frequency bursts (Mulle et al., 1986; Avanzini et al., 1989; Bal and McCormick, 1993; Contreras et al., 1993). The activation of PGN-mediated IPSPs in PGN cells was capable of inhibiting not only single-spike activity but also determining whether or not the postsynaptic PGN cell generated a low threshold Ca2+ spike and a burst of action potentials. The ability of PGN to PGN inhibition to determine the amplitude and duration of low threshold Ca2+ spikes and bursts of action potentials occurring in these cells has important functional consequences on the generation of normal and abnormal thalamocortical activity.
During periods of slow wave sleep, thalamocortical circuits generate rhythmic oscillatory activity in the frequency range of delta waves (0.5–4.0 Hz), spindle waves (6–14 Hz), and even slower rhythms (0.1–0.5 Hz) (Steriade et al., 1993, 1994). During the generation of spindle waves in the ferret LGNd in vitro, PGN neurons receive barrages of EPSPs from burst firing in thalamocortical neurons (Bal et al., 1995b). These barrages of EPSPs typically activate low threshold Ca2+ spikes and bursts of 2–10 action potentials in PGN neurons and therefore result in the activation of IPSPs in neighboring PGN neurons. The activation of intra-PGN IPSPs controls the amplitude and duration of the thalamocortical-induced EPSP barrages as well as the intensity and duration of burst discharges generated by PGN cells (Bal et al., 1995b; present study). In contrast to those in thalamocortical cells, we have not observed barrages of IPSPs in GABAergic PGN neurons to result in the generation of rebound low threshold Ca2+ spikes.
After the block of GABAA receptors, thalamocortical-induced barrages of EPSPs in PGN cells become larger in amplitude, and PGN neurons generate high-frequency discharges of up to 60 action potentials per burst (Bal et al., 1995b). This increase in discharge of PGN neurons has important network consequences because it results in the strong activation of GABAB postsynaptic receptors on thalamocortical cells (Bal et al., 1995a,b; Kim et al., 1997;Sanchez-Vives and McCormick, 1997), resulting in slow and prolonged (∼300 msec) IPSPs in these neurons. The end result of this prolongation of the GABAergic IPSPs in thalamocortical cells, in conjunction with the block of GABAA receptors, is that the spindle waves are abolished and replaced by a “paroxysmal” slow oscillation in which nearly every thalamocortical and PGN neuron discharges synchronously every 250–500 msec (2–4 Hz) (Bal et al., 1995a,b). This transition, which depends critically on prolonged burst firing in PGN cells, may explain the critical involvement of GABAB receptors in the generation of generalized spike and wave seizures in the rodent (Hosford et al., 1992; Marescaux et al., 1992; Snead, 1995). Interestingly, in one rodent model of generalized epilepsy, thalamic reticular neurons have been found to express unusually large concentrations of the low threshold Ca2+ current (Tsakiridou et al., 1995), although at an earlier age than the behavioral expression of the seizures (Marescaux et al., 1992).
Given that PGN neurons inhibit one another through axon and perhaps dendrodendritic connections, the PGN may be thought of as an interactive sheet of GABAergic neurons with local inhibitory interactions. Presumably, the activation of a particular point of neurons within this network will result in the inhibition of neurons in an annulus surrounding the activated point. Because perigeniculate neurons appear to innervate only thalamocortical cells in the ferret LGNd (Bal et al., 1995a,b), this pattern of activity in the PGN may then be represented in the A-laminae as a center of inhibition surrounded by disinhibited thalamocortical cells. Similarly, if a local group of PGN cells were to be inhibited, for example from the activation of cholinergic fibers (Lee and McCormick, 1995) or extrathalamic sources of GABAergic innervation (Jourdain et al., 1989;Pare et al., 1990; Asanuma, 1994), then there would be expected to be a center of decreased excitability adjacent to a surround of increased excitability in this nucleus. This pattern of activity would then be reflected in the LGNd as a localized region of increased excitability surrounded by a region of decreased excitability (see also Steriade, 1991). In this manner, the local inhibitory influences in the PGN may set-up a “center-surround” organization for the modulation of thalamocortical excitability in the LGNd. However, as demonstrated with the interaction between EPSP and IPSP barrages in PGN neurons during the generation of spindle waves, the influence of monosynaptic connections must be interpreted in the context of the activity of all of the different patterns of synaptic inputs impinging on each cell. For example, the activation of a given portion of the PGN may not result in a decrease in discharge rate of neighboring PGN cells if these cells are also activated simultaneously by the same or other inputs. Indeed, one possibility is that the PGN to PGN connection serves to influence the timing as well as the probability of action potential generation.
One characteristic of PGN and thalamic reticular cells is their long dendritic trees (Scheibel and Scheibel, 1966; Uhlrich et al., 1991) and their relatively large receptive fields (Uhlrich et al., 1991; Jones and Sillito, 1994). Investigations of the receptive field properties of perigeniculate neurons in the anesthetized cat indicate that they typically are binocular, although dominated by one eye, have large receptive fields in comparison to thalamocortical cells, and exhibit both ON and OFF visual responses (Sanderson, 1971; So and Shapley, 1981; Xue et al., 1988; Uhlrich et al., 1991). Presumably these properties allow PGN neurons to contribute to binocular and long-range inhibition in the LGNd (Eysel et al., 1986). We propose that intra-PGN inhibition may operate to functionally limit the dendritic length of PGN cells such that localized activation of inhibitory inputs may block or reduce the activation of these neurons from localized branches of these neurons. Similarly, the activation of ascending cholinergic systems, which can activate K+ conductances in PGN cells (Lee and McCormick, 1995), could also limit the ability of distal excitatory inputs to generate action potentials in these neurons. Thus, in the awake, behaving animal, the effective dendritic length of perigeniculate cells is likely to be controlled in a dynamical manner, depending on the activity of neighboring perigeniculate cells as well as ascending inputs from the brainstem. The investigation of these dynamic properties of PGN function are likely to yield a better understanding of the role of intra-PGN inhibition in normal and abnormal thalamocortical function.
Note added in proof: A recent study has also demonstrated that inhibition between GABAergic thalamic reticular neurons may be capable of strongly controlling the duration of low threshold Ca2+ spikes in these cells [Ulrich D, Huguenard JR (1997) GABAA-receptor mediated rebound burst firing and burst shunting in thalamus. J Neurophysiol 78:1748–1751].
Footnotes
This research was supported by grants from National Institutes of Health and the Klingenstein Fund. M.V.S.-V. was a fellow from NATO and the Epilepsy Foundation of America. We thank Uhnoh Kim for helpful discussions.
Additional information about these and related findings may be obtained at http://info.med.yale.edu/neurobio/mccormick/mccormick.html.
Correspondence should be addressed to David A. McCormick, 333 Cedar Street, Section of Neurobiology, Yale University School of Medicine, New Haven, CT 06510.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}