

Abstract
The aim of this study was to elucidate electrophysiologically the actions of dopamine and SKF38393, a D1-like dopamine receptor agonist, on the membrane excitability of striatal large aspiny neurons (cholinergic interneurons). Whole-cell and perforated patch-clamp recordings were made of striatal cholinergic neurons in rat brain slice preparations. Bath application of dopamine (1–100 μm) evoked a depolarization/inward current with an increase, a decrease, or no change in membrane conductance in a dose-dependent manner. This effect was antagonized by SCH23390, a D1-like dopamine receptor antagonist. The current–voltage relationships of the dopamine-induced current determined in 23 cells suggested two conductances. In 10 cells the current reversed at −94 mV, approximately equal to the K+ equilibrium potential (EK); in three cells theI–V curves remained parallel, whereas in 10 cells the current reversed at −42 mV, which suggested an involvement of a cation permeable channel. Change in external K+ concentration shifted the reversal potential as expected for Ek in low Na+ solution. The current observed in 2 mm Ba2+-containing solution reversed at −28 mV. These actions of dopamine were mimicked by application of SKF38393 (1–50 μm) or forskolin (10 μm), an adenylyl cyclase activator, and were blocked by SCH23390 (10 μm) or SQ22536 (300 μm), an inhibitor of adenylyl cyclase. These data indicate, first, that dopamine depolarizes the striatal large aspiny neurons by a D1-mediated suppression of resting K+ conductance and an opening of a nonselective cation channel and, second, that both mechanisms are mediated by an adenylyl cyclase-dependent pathway.
- dopamine
- acetylcholine
- striatum
- basal ganglia
- cholinergic neurons
- giant aspiny neurons
- patch clamp
- slice
- electrophysiology
Because clinical studies conducted in the early 1960s indicated that both muscarinic receptor antagonists and dopaminergic receptor agonists ameliorate the symptoms of Parkinson’s disease, it was classically proposed that cholinergic and dopaminergic tones are in balance in the extrapyramidal system (Barbeau, 1962; Lehmann and Langer, 1983). With the advent of chemical agents that act selectively on dopamine (DA) D1 receptors and of brain microdialysis techniques, the acetylcholine (ACh)/DA balance hypothesis has been modified to encompass the finding that activation of DA D1 receptors stimulates the ACh release in the striatum, whereas D2 agonists reduce it (Stoof et al., 1992; Consolo et al., 1993; Di Chiara et al., 1994). This ACh is thought to be supplied primarily by large aspiny neurons (cholinergic interneurons), which account for only 1–2% of the total neuronal population of the rat striatum (Bolam et al., 1984; Phelps et al., 1985). In situ hybridization studies, however, have provided evidence that only a small number of the striatal cholinergic neurons contain D1 receptor mRNAs, whereas the majority of the cells express D2 receptor mRNAs (Le Moine et al., 1991). It is therefore currently hypothesized that although the inhibition of its release by D2 DA receptor activation is direct, the ACh release evoked by D1 DA receptor activation might be indirect through two alternative pathways: excitation of medium-sized spiny neurons containing D1 receptors that release substance P (Ajima et al., 1990; Anderson et al., 1994; Di Chiara et al., 1994) and stimulation of D1 receptors in the thalamus, cerebral cortex, or substantia nigra that in turn elicits an increase in glutamate input to the striatum (Damsma et al., 1991; Consolo et al., 1996a,b; Abercrombie and DeBoer, 1997). Recently, five DA receptor isoforms, grouped into two subfamilies, D1-like (D1 or D1a, and D5 or D1b) and D2-like (D2(short), D2(long), D3, and D4), have been defined so far. Bergson and colleagues (1995) reported that only D5, and not D1, receptor-specific antibodies labeled the striatal cholinergic neurons of rhesus monkey. In addition, a patch-clamp study using a single cell RT-PCR technique (Yan et al., 1997) demonstrated that the majority of the neostriatal cholinergic neurons of the rat (88%) contain D5/D1b receptor mRNAs rather than D1a mRNAs (17%), whereas all the cells express D2 mRNAs. Therefore, direct activation of D5/D1b DA receptors on the striatal cholinergic neurons might contribute to the evoked ACh release. Indeed,Login and Harrison (1996) reported that 50 μm(±)-SKF38393, a D1-like receptor agonist, increased the basal rate of ACh release from dissociated striatal cholinergic cells by 50%, with this action blocked by the D1-like receptor antagonist SCH23390. The present study was conducted to examine the effects of D1-like receptor activation on the membrane excitability of rat striatal giant aspiny neurons to explore the possibility of direct regulation of ACh release by D1-like receptors. We present evidence here that DA and (±)-SKF38393 directly depolarize the cells, which is sufficient to generate action potentials in some cases.
MATERIALS AND METHODS
Slice preparations. Rats were bred under standard animal housing conditions, with a 12 hr light/dark cycle. Food and water were available ad libitum. Experiments were performed on 200-μm-thick sagittal rat brain slices containing the striatum. The ages of the rats (Wistar) ranged from 12- to 20-d-old. The animals were deeply anesthetized with ether and decapitated. The brains were quickly removed and submerged in ice-cold physiological Ringer’s solution oxygenated with a mixture of 95% O2 and 5% CO2. Slices were superfused at 3 ml/min with physiological saline at 33°C. The solution comprised (in mm): NaCl 124, KCl 3, CaCl2 2.4, MgCl2 1.2, NaH2PO4 1, NaHCO3 26, glucose 10, gassed with 95% O2 and 5% CO2.
Whole-cell and perforated patch-clamp recordings. Whole-cell recordings were made with glass pipettes (3–4 MΩ) that contained (in mm): potassium methylsulfate 120, KCl 6, NaCl 6, EGTA 0.6, HEPES 10, MgCl2 2, ATP 4, GTP 0.3, biocytin 20, pH 7.2. Tetrodotoxin (TTX) was added to the external solution at 0.5 μm when required. For ion substitution experiments, NaCl was replaced with choline chloride in low-sodium external solutions. When CdCl2 was added at 200 μm, NaH2PO4 was omitted. To isolate the nonselective cation current, tetraethylammonium chloride (TEA-Cl) 30 mm, 4-aminopyridine (4-AP) 10 mm, CsCl 2 mm, MgCl2 3.6 mm, no Ca2+, and TTX 0.5 μm were added to the external solution, and cesium-methanesulfonate was substituted for potassium methylsulfate in the pipette solution. Membrane potentials and currents were recorded with an EPC-7 amplifier (List). Series resistance was partially compensated for by the amplifier. The given membrane potentials have been corrected for the junction potential between the pipette solution (potassium methylsulfate) and the bath solution before making a gigaseal (usually the pipette 7 mV negative to the bath). Membrane conductance was measured from the slope of the chord in −10 mV step pulses.
Perforated patch recordings using nystatin were also made to confirm the results obtained by the whole-cell recordings. Nystatin (5 mg) was dissolved in a methanol solution containingN-methyl-d-glucamine (Sigma, St. Louis, MO) (0.1m) and methanesulfonate (0.1 m) adjusted to pH 7.0 immediately before use. After the nystatin solution (50–100 μl) was dried by nitrogen gas, the internal solution was added and shaken well at a final concentration of 250–500 μg/ml.
Electrophysiological identification of the neostriatal neurons. Neurons in the neostriatum were visualized using a 40× water immersion objective. Particular attention was paid to preferentially recording the largest neurons in a slice. At the beginning of the recording, a set of depolarizing and hyperpolarizing step current pulses were routinely given in the current-clamp mode to identify the neuronal types. Cells whose resting membrane potentials were over −50 mV were discarded. Electrophysiological criteria for identification of the neostriatal neurons of the rat have been described in detail previously (Kawaguchi, 1992, 1993). Briefly, the cells that had a resting potential of approximately −60 mV and displayed long-lasting afterhyperpolarizations and strong time-dependent hyperpolarizing rectification were classified as long-lasting afterhyperpolarization (LA) cells. Even when recordings were made with a Cs+-containing pipette, the voltage traces obtained immediately after the rupture of the patch membranes usually displayed, very briefly (a few seconds), firing patterns similar to those obtained with K+-patch pipettes, which suggested the neuronal types. Immunohistochemical evidence has indicated that LA cells are large aspiny cholinergic interneurons (Kawaguchi, 1992, 1993).
Histochemical procedures. To identify the recorded cells morphologically, 20 mm biocytin was included in the pipette solution to fill them by diffusion (Horikawa and Armstrong, 1988). Slices (200 μm) containing biocytin-filled cells were fixed by immersion in 4% paraformaldehyde and 0.2% picric acid in 0.1m phosphate buffer (PB) overnight in at 4°C, rinsed in PB for 30 min, and incubated in PB containing 0.5% H2O2 for 30 min to suppress endogenous peroxidase activity. They were then incubated in 10 and 20% sucrose for 30 min and 1 hr, respectively, frozen, and stored in a freezer until histochemistry was performed. The slices, without resectioning, were then washed with Tris-buffered saline (TBS) containing 0.5% Triton X-100 and avidin–biotin peroxidase complex (Vector Laboratories, Burlingame, CA) at a dilution of 1:100 for 4–6 hr at room temperature. After rinsing they were reacted with 3, 3′-diaminobenzidine tetrahydrochloride (DAB; 0.05%) and H2O2 (0.003%) in TBS, post-fixed in 0.1% osmium tetroxide in 0.1 m PB for 5 min, and mounted on slides.
Drugs. DA and other agents such as CsCl, CdCl2, TTX, and nifedipine were applied by changing the solution superfusing the slice to one that contained the drug. Time taken to reach the neurons was usually <1 min. Drugs used were DA, (±)-sulpiride (Sigma), (±)-SKF38393 hydrochloride, (−)-quinpirole hydrochloride, R(+)-SCH23390 hydrochloride, (+)-bromocriptine methanesulfonate (RBI, Natick, MA), tetrodotoxin (Sankyo), 4-aminopyridine (Sigma), tetraethylammonium chloride (Nakarai tesque), nifedipine (Sigma), 9-(tetrahydro-2-furyl) adenine (SQ22536, Sigma), and 1,2-bis(2-aminophenoxy)ethane-N,N,N′,N′-tetra-acetic acid (BAPTA, Sigma). DA was dissolved in the external solution containing 0.1% L(+)-ascorbic acid or 50 μm sodium metabisulfite immediately before use to prevent its oxidative degradation. l(+)-ascorbic acid or sodium metabisulfite alone did not exert any effects on the excitability of the neurons. All the experiments were performed in a dark room to slow the light-induced degradation of some drugs.
Results are expressed as mean ± SD values, and comparisons between groups were made using the Student’s t test.
RESULTS
Physiological and morphological identification of the large aspiny neurons
The results presented here were obtained with a total of 186 rat neostriatal neurons identified as large aspiny cholinergic interneurons (LA cells). For this neuron sample, resting membrane potential was −60.1 ± 5.5 mV (n = 42), and membrane input conductance was 3.5 ± 1.7 nS (n = 41). Injection of depolarizing current pulses elicited action potentials (amplitude 66.0 ± 11.6 mV, width at half-amplitude 1.6 ± 0.3 msec) that were followed by a longer-duration and larger-amplitude afterhyperpolarization (−16.5 ± 8.2 mV; n = 40) than the other classes of neurons (Fig.1A). During hyperpolarizing current pulses, they showed prominent sag, which was demonstrated to be caused by activation of hyperpolarization-activated cation currents (Ih) (Jiang and North, 1991; Kawaguchi, 1993). Subsequent staining with biocytin revealed large somata with aspiny or sparsely spiny dendrites.

Effects of DA on neostriatal large aspiny neurons.A, Membrane properties in response to constant-current pulses applied intracellularly. The resting membrane potentials (∼ −60 mV) and input resistances (∼430 MΩ), as well as the long-duration, large-amplitude afterhyperpolarization and the prominent sag during hyperpolarizing current pulses all fit well with the physiological properties of large aspiny neurons (LA cells).B, Dose–response relation for DA in neostriatal large aspiny cells. The holding potential was −60 mV. DA was bath-applied with 50 μm sodium metabisulfite or 0.1% ascorbic acid to prevent its oxidative degradation. Vertical linesrepresent SD values. Numbers in parentheses refer to numbers of tested cells. C, DA depolarization of a striatal large aspiny neuron. The external solution containedl(+)-ascorbic acid (0.1%). DA (50 μm), bath-applied at 3 ml/min, caused a transient depolarization and a train of action potentials. Haloperidol (100 μm) completely blocked the DA response. D, The effects of DA were analyzed under a whole-cell voltage clamp (holding potential −60 mV).Da, Suppression of the 100 μm DA-induced current by pretreatment with SCH23390 (10 μm), an antagonist of the D1-like DA receptor. TTX was present at 0.5 μm in the external solution. Note that the amplitude of DA-induced current gradually increased during washout of the antagonist. Db, SCH23390 (10 μm)) alone evoked an outward shift of the holding current. Application of DA (100 μm) elicited a small inward current. TTX was 0.5 μm. Membrane conductances were monitored periodically with hyperpolarizing voltage steps of 10 mV. Holding potential was −60 mV. Calibration bars apply to both a andb.
Dopamine depolarized the LA cells by activation of D1-like DA receptor in a dose-dependent manner
Figure 1C shows the effects of DA on the excitability of a striatal large aspiny neuron. The whole-cell patch technique was used under a current clamp without TTX. DA was bath-applied in external solution containing 0.1% l(+)-ascorbic acid or 50 μm sodium metabisulfite. In all cells tested (8/8 cells), application of DA (1–100 μm) elicited a slow transient depolarization (9.4 ± 5.0 mV). This was sufficient to initiate a train of action potentials in three cells. The depolarization was completely blocked by haloperidol (1 μm), a nonselective DA receptor antagonist.
The effects of DA were analyzed under whole-cell voltage clamp (holding potential −60 mV) with TTX (0.5 μm) in the external solution (Fig. 1D). DA invariably induced an inward current, and it was suppressed by application of SCH23390 (10 μm), an antagonist of the D1-like DA receptor. Combined administration of DA and SCH23390 evoked an outward current (7/19 cells, 12.0 ± 8.6 pA), no change (1/19 cells), or a small inward current (11/19 cells, −12.3 ± 9.3 pA), so that the average amplitude of the DA-induced current (−60.6 ± 32.5 pA,n = 17) was significantly larger than that obtained with SCH23390, an antagonist of the D1-like DA receptor (−2.7 ± 14.6 pA; n = 19; p < 0.01) (see Fig. 6). As shown in Figure 1Da, combined application of DA (100 μm) plus SCH23390 (10 μm) produced almost no appreciable change in the holding current (+0.7 pA) in this cell. However, application of DA (100 μm) alone produced a small decrease in membrane conductance (from 6.9 to 6.8 nS) along with an inward shift of the holding current (−23.7 pA). The suppression of the DA-induced current by the antagonist was gradually relieved during washing, reaching −35.0 pA at the end of the experiment. In all cells tested (n = 6), the DA-induced current in the presence of SCH23390 (−8.35 ± 17.4 pA) was significantly smaller than that obtained 6 min after washout of the antagonist (−23.2 ± 11.8 pA;p < 0.01). Notably, application of SCH23390 itself elicited an outward shift of the holding current in 12 of 19 cells as exemplified in Figure 1Db. The average SCH23390-induced current was 11.5 ± 15.0 pA (n = 19). This indicates that some cells might be continuously slightly depolarized by D1-like receptor activation, possibly by spontaneous release of DA from terminals in the vicinity. These results suggest that the majority of the DA-induced current is mediated by D1-like DA receptor activation.
Overall, this DA-induced inward current was obtained in all of the striatal LA cells tested (43 cells), and the effect was concentration-dependent. A dose–response curve for the DA-induced inward current is shown in Figure 1B. The EC50 value was determined to be ∼15 μm. The DA-induced current reached steady-state values with a delay of ∼1 min without showing any desensitization. Usually the current stayed constant during DA application and longer treatment (>1 min), often preventing return of the holding current to the pretreatment level (Fig. 2B). The DA-induced inward current persisted even in perfusion medium containing 200 μm Cd2+, 0 mmCa2+, 3.6 mm Mg2+, and 0.5 μm TTX (n = 5), suggesting mediation by a direct effect of DA on recorded cells (data not shown; also see Fig. 5). In a small population of the cells (3/36 cells) that were mostly recorded with the nystatin method, the inward current was preceded by a small transient outward current as shown in Figure4B. This was not studied further in detail.
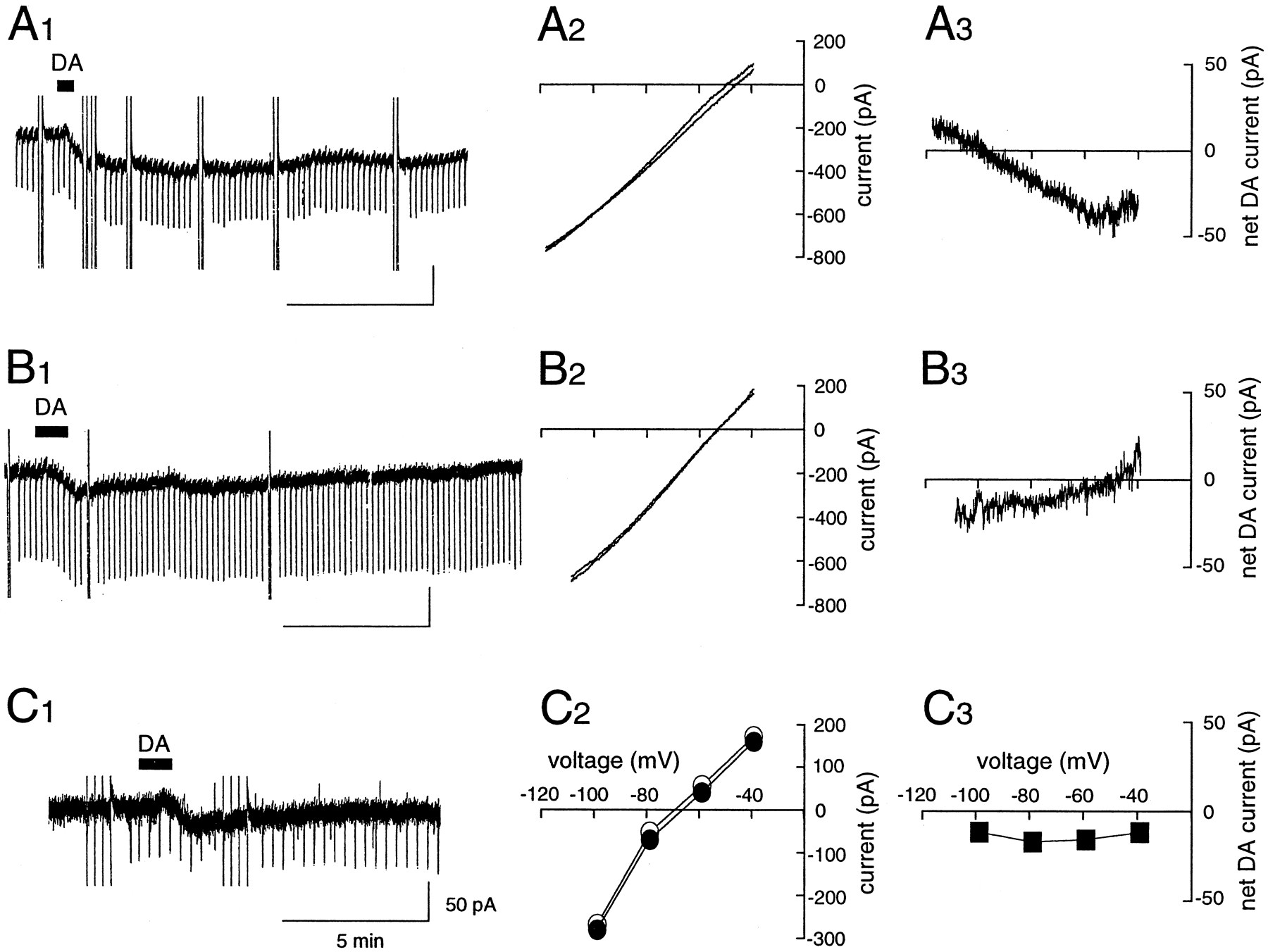
Two conductances mediate the DA-induced inward current. Shown are typical examples expressing a reversal potential close to the potassium equilibrium potential (EK, −100.6 mV) (A), or a reversal close to −40 mV (B), or no reversal potential within the tested voltage range (C). Voltage ramps (10 mV/sec) were used to construct I–V curves forA and B. Data forI–V plots in C were taken at the end of the voltage pulse. C2 , open circles indicate before DA application; filled circles indicate after DA application. There was no significant difference between measurements taken at the beginning and at the end of the voltage pulse. A3, B3 , and C3 indicate the net DA-induced currents, calculated as differences between I–Vplots obtained before and during the peak of the DA responses.
Two separate conductances are involved in the DA responses in LA cells
Detailed comparison of the current–voltage relationships of the DA-induced current suggested that at least two separate conductances mediated the DA response. In 10 of 23 cells, the response to DA reversed at −93.7 ± 15.7 mV, approximately equal to the estimated K+ equilibrium potential (EK, −100.6 mV), with a slight reduction in membrane conductance from 7.4 ± 2.1 to 7.0 ± 1.7 nS, as shown in Figure 2A. The net current was −35.5 ± 18.4 pA. In contrast, 10 of 23 cells showed an increase in membrane conductance from 5.9 ± 3.4 to 8.3 ± 3.6 nS. These cells showed a linear current–voltage relation with a positive slope and a reversal potential of −42.1 ± 10.1 mV (Fig.2B). The amplitude of the net current was −48.0 ± 47.1 pA. In the remaining three cells, the DA-induced inward current was associated with almost no change in membrane conductance from 4.5 ± 1.3 to 4.5 ± 0.8 nS (Fig. 2C). The net current was −22.6 pA and the I–V relations did not cross within the voltage range tested (−120 to −40 mV).
Because these results suggested an involvement of multiple separate conductances in the generation of DA responses, cells were first tested in perfusion medium containing low Na+ (27 mm) and 0.5 μm TTX to maximize the contribution of a K+ current component to the DA response (Fig. 3). The reversal potentials of the DA-induced current in this particular experiment were −88.5 ± 17.8, −56.1 ± 19.5, and −45.0 ± 16.3 mV with external K+ concentrations of 3, 9, and 16 mm, respectively. The values were rather more positive than estimated with the Nernst equation, probably because they contained effects of Na+-permeable conductance and insufficient space clamping. However, the reversal potentials shifted to less negative values in solutions of elevated K+as expected and gave a Nernst slope of −53.3 ± 5.4 mV per 10-fold change in K+ concentration. These results indicate that K+ channels were closed during flow of the DA-mediated inward current.

One of the inward currents caused by DA is K+ dependent. A Nernst plot of reversal potentials against three K+ concentrations. Na+ concentration in the external solutions was reduced to 27 mm to maximize the K+component. TTX was present at 0.5 μm. Thecontinuous line plots the mean values of the reversal potentials obtained at each concentration. The dashed line was calculated from the Nernst equation for a K+-selective electrode. Numbers in parentheses refer to numbers of tested cells.
Figure 4 shows an example in which the same cell was exposed first to 100 μm DA in a low Na+ solution and then in normal Ringer’s solution. It showed linear current–voltage relations within a voltage range tested with a reversal potential near the equilibrium potential of K+ channel in a low Na+ solution. In the control solution (151 mm Na+), however, the I–V relations did not cross within the voltage range between −120 and −60 mV. Importantly, the net current in this cell was −33.2 pA in the low Na+solution and −95.8 pA in the control solution. On average, the DA-induced inward current at the holding potential of −60 mV in the low Na+ solution was −27.6 ± 14.6 pA (n = 7), which was significantly smaller than that in the control solution (−60.6 ± 32.5 pA; n = 17;p < 0.05) (see Fig. 6). These results suggest that at least in some cells, activation of Na+-permeable cation conductance is involved in the DA-induced current, together with a block of resting K+ conductance.

Lowering external Na+ ion concentration reduces the amplitude of the DA-induced inward current in a cell. Inward currents caused by DA (100 μm) in 27 mm (A1 ) and 151 mm(B1 ) [Na+]o, respectively. Voltage ramps (−125 to −60 mV, 9.3 mV/sec) were applied before (1, 2) and during (3, 4) the inward shift of the holding current. A2 ,B2 , Two superimposedI–V plots, one before (1, 3) and the other during (2, 4) the peak DA response. A3 , B3 ,I–V plots of the net DA-induced currents. Note that the reversal potential was close toEK in 27 mm[Na+]o, but no reversal potential was obtained within the voltage range tested in 151 mm [Na+]o. In a small population of the cells, DA elicited an early outward current followed by an inward current as observed in B1 .
To characterize further the role of the cation conductances, barium ions (2 mm) were included in the external solution to block K+ conductance. Barium ions are known to inhibit various K+ channels such as the delayed rectifier, inward rectifier, leak K+, and M-channels. In all cells (n = 8), switching to the Ba2+-containing medium evoked an inward shift of the holding current of −108.4 ± 48.6 pA with a reduction of membrane conductance from 10.6 ± 5.0 to 5.1 ± 1.8 nS. The reversal potential of the Ba2+-induced current was −86.2 ± 6.4 mV, suggesting that the K+channels were indeed blocked. Application of DA (100 μm) under these conditions induced a further shift of the holding current (−16.8 ± 9.2 pA; n = 8) (see Fig. 6) accompanied by an increase in membrane conductance from 4.7 ± 1.5 nS to 5.1 ± 1.7 nS in six of eight cells tested (Fig.5A). The current–voltage relationships of the net DA-induced inward current showed a slightly positive curve within a voltage range between −110 and −35 mV and a negative curve in more depolarized regions (−35 to +20 mV). To test the possibility that the latter component might be attributable to opening of voltage-sensitive Ca2+ channels, 200 μm Cd2+ and 5 μmnifedipine were added to the external solution. Cs+(2 mm) was also included to block the hyperpolarization-induced cation channel (Ih). In 10 of 11 cells tested, DA (100 μm) evoked a small inward current (−21.7 ± 7.3 pA;n = 10) (Fig. 6) with a small increase in membrane conductance from 4.9 ± 1.8 nS to 5.1 ± 2.1 nS in Ba2+-containing solution. TheI–V curves obtained before and during DA exposure again crossed or almost crossed at potentials around −40 mV in 10 cells (Fig. 5Ba). Also, as the membrane potential was further depolarized to +20 mV, these cells showed a decrease in membrane conductance, indicating that this component might be not caused by activation of voltage-dependent Ca2+channels. Rather, it might be caused by either the voltage dependency of the channel or simply by a space clamp error. In one cell, the net DA current reversed the sign at −28 mV (Fig. 5Bb), suggesting that cations permeated nonselectively through the channel. Similar findings were obtained when the recording was made with a Cs+-containing patch pipette in the presence of TTX (n = 13; data not shown).

The Ba2+-resistant component of the DA-induced current was evoked by DA (100 μm) in a solution containing 2 mm Ba2+(A) and a solution containing Ba2+ (2 mm), Cs+ (2 mm), and Cd2+ (200 μm) (B). A2, Ba2 , Bb2 , Two superimposed I–V plots, one before and the other during the peak DA response.A3 , The net DA-induced current showed an atypical I–V relation. Addition of Cd2+ did not eliminate the net DA inward current seen at more depolarized potentials than −35 mV (Ba3 ). In another cell the current reversed the sign at −28 mV (Bb3 ).

Summary histograms representing mean ± SD of the amplitude of DA-induced inward currents. SDs are shown withbars. Numbers in parentheses refer to numbers of tested cells. From the left, the current induced by DA (100 μm) as a control (open bar), that with DA plus SCH 23390 (10 μm) (filled bar) in saline with TTX (0.5 μm), that with DA alone in a solution containing 27 mm Na+ (gray bar), that in a solution containing Ba2+, that in a solution containing Ba2+, Cs+, Cd2+, and nifedipine (5 μm), that recorded with a Cs+-filled pipette in a saline, and that in a low Na+ (27 mm) solution. Comparisons were made with the Student’s t test against the control group of neurons tested in the saline solution (*p < 0.05; ** p < 0.01).
The Na+ permeability was also tested with a Cs+-containing patch pipette by lowering the external Na+ concentration. Overall, the DA-induced inward current in 26 mm Na+ was −10.7 ± 13.2 pA (n = 5), whereas that under high Na+ conditions was −34.7 ± 18.1 pA (n = 15) (Fig. 6), suggesting that a TTX-insensitive Na+-permeable conductance may contribute partly to the inward current. A similar cation conductance has been reported for neurons of the prefrontal cortex of the rat (Shi et al., 1997) and characterized as a nonspecific mechanism in that it was not sensitive to classical DA agonists and antagonists. However, application of (±)-SKF38393 (5 μm), an agonist of D1-like DA receptor, elicited a small DA-induced inward current in Ba2+-containing solution in one of four cells tested, suggesting that this possibility is unlikely in this cell.
Effects of a D5/D1 DA receptor agonist on striatal LA cells
The effects of (±)-SKF38393 were tested in 108 striatal LA cells to allow comparison with the DA-induced membrane depolarization. Four cells were also analyzed for the effects of a mixture of DA (100 μm) plus sulpiride (10 μm), a D2-like receptor antagonist, but the results were similar to those obtained with (±)-SKF38393 (data not shown). In the current clamp mode, (±)-SKF38393 (10 μm) depolarized 10 of 11 cells (9.2 ± 4.9 mV; n = 10) and elicited action potentials in four cells (Fig.7A). (±)-SKF38393 (1–50 μm) evoked an inward current in the whole-cell clamp configuration in a dose-dependent manner. The amplitude clamped at the resting membrane potential (−60 mV) was −17.3 ± 31.3 pA (n = 12), −19.1 ± 30.2 pA (n = 8), −41.7 ± 52.1 pA (n = 40), and −53.1 ± 39.4 pA (n = 6) at 1, 3, 10, and 50 μm, respectively. The current–voltage relations were also similar to those with DA. The I–V curves of the net SKF38393-induced current of 10 of 14 cells reversed the sign at −88.26 ± 24.7 mV with a negative slope conductance (Fig.7B). On the other hand, those of the remaining four cells did not cross the voltage axis within the voltage range tested (−120 to −40 mV) (Fig. 7C). Also, the I–Vcurve of the net outward current elicited by administration of SCH23390 was just the opposite of the SKF38393-induced current (data not shown). These results suggest that, as in the DA case, SKF38393 evokes an inward current by two separate ionic mechanisms: blockade of K+ conductance and opening of nonselective cation conductances.

Effects of the D1-like agonist, SKF38393, on striatal LA neurons. A, A whole-cell current-clamp recording with a resting membrane potential of −65 mV illustrates a slowly rising, prolonged, and reversible membrane depolarization with action potentials occurring during the peak of the response. B1 , C1 , Voltage-clamp traces (holding potential −60 mV) recorded from another LA cell in saline containing TTX (0.5 μm) illustrate a slow inward current induced by SKF38393. Two superimposedI–V plots before (open circles in B2 and 1 inC2 ) and during the peak of the response (filled circles in B2 and2 in C2 ) show a reversal potential close to EK(B2 ) and no reversal potential within the tested voltage range (C2 ). B3 ,C3 , I–V plots of the net SKF38393-induced currents (filled triangles in B3 and 2–1in C3 ).
Forskolin mimics and occludes the (±)-SKF38393-induced inward current
Because D1-class DA receptors are known to positively couple with G-proteins and an adenylyl cyclase-cAMP cascade, the effects of (±)-SKF38393 on the membrane excitability of LA cells might also be mediated by this signaling pathway. To test this possibility, we bath-applied forskolin, a lipophilic adenylyl cyclase activator, to the cells at 10 μm (Fig.8A). A large inward current was evoked in all cells tested (−98.1 ± 82.3 pA;n = 13), whereas dideoxyforskolin, an inactive enantiomer of the forskolin, caused no response at 10 μm(1.2 ± 2.2 pA; n = 3). The averageI–V curve of the net forskolin-induced current was qualitatively similar to those obtained with DA and (±)-SKF38393 (data not shown). The current decreased with membrane depolarization, but no reversal of polarity was seen within the tested voltage range. Furthermore, it was found that the effects of (±)-SKF38393 were no longer evident after the forskolin current reached a steady state (Fig.8B). These results suggest that adenylyl cyclase transduces D1-like DA receptor-mediated effects in striatal LA cells.

Forskolin, an activator of adenylyl cyclase, mimics and occludes the effects of SKF38393 on the striatal LA cells.A, Dideoxyforskolin (10 μm,stippled bar), an inactive forskolin analog, evoked no response, whereas forskolin (10 μm, filled bar) caused a slow and reversible inward current. Holding potential was −60 mV. B, Application of SKF38393 (filled bars) resulted in a slow inward current (left trace). Fifteen minutes after washout of SKF38393 (10 μm), treatment with forskolin (10 μm,stippled bar) elicited a large slow inward current and occluded the effect of the second application of SKF38393.
SQ22536, an adenylyl cyclase inhibitor, reduces D5/D1 DA receptor-mediated inward currents
If adenylyl cyclase mediates the effects of (±)-SKF38393, its inhibition should reduce the actions of (±)-SKF38393. In fact, application of 9-(tetrahydro-2-furyl) adenine (SQ22536), an agent that is known to decrease the activity of adenylyl cyclase, reduced the amplitude of the (±)-SKF38393-induced inward current. Neostriatal slices were preincubated in an oxygenated saline containing SQ22536 (300 μm) for 30 min to 6 hr. Control slices were also taken from the same animals and incubated in the saline in the same manner. Recordings were made alternately. The amplitudes of the (±)-SKF38393-induced currents in SQ22536-treated cells were −9.6 ± 13.9 pA (n = 5), −8.9 ± 13.9 pA (n = 5), and −10.9 ± 14.2 pA (n= 6) at 3, 10, and 50 μm, respectively. In contrast, those in control cells were −3.1 ± 6.0 pA (n = 6), −38.9 ± 65.0 pA (n = 6), and −53.1 ± 39.4 pA (n = 6; p < 0.05) at 3, 10, and 50 μm, respectively. These results provide further evidence that activation of adenylyl cyclase is required to elicit the actions of (±)-SKF38393.
BAPTA has no effects on D5/D1 DA receptor-mediated inward currents
Dopamine D1-like receptors have been observed to increase the production of inositol triphosphates and mobilize intracellular Ca2+ in oocytes from Xenopus laevis with messenger RNA isolated from rat striatum (Mahan et al., 1990). In addition, activation of D1-like receptors has been reported to be associated with the facilitation of L-type Ca2+ channels, an effect mediated through a cAMP-dependent protein kinase in chromaffin cells (Artalejo et al., 1990) and in a subset of rat neostriatal neurons (Surmeier et al., 1995). Therefore, to test whether a rise in intracellular Ca2+ mediated the effects of D1-like receptor stimulation in the cells, BAPTA, an agent that is known to chelate cytosolic Ca2+ ions, was included in patch pipettes at 10 or 20 mm, and the actions of (±)-SKF38393 (10 μm) were compared with those of forskolin (10 μm), and substance P (0.5 μm) as a control, which is known to elicit an inward current in LA cells (Aosaki and Kawaguchi, 1996). As shown in Figure 9, our data clearly showed no signs of suppression of the (±)-SKF38393-induced currents evoked by (±)-SKF38393 in all cells tested [−100.7 ± 78.5 pA at 10 mm BAPTA (n = 5), −56.7 ± 32.2 pA at 20 mmBAPTA (n = 5)]. These results suggest that a rise in cytosolic Ca2+ ions is not associated with the observed actions of (±)-SKF38393 in LA cells.

Slow inward currents evoked by SKF38393 (10 μm), substance P (0.5 μm), and forskolin (10 μm) are unaffected by intracellular application of BAPTA (20 mm), a potent Ca2+chelator.
It is thought that activation of D2 DA receptor inhibits ACh release in the striatum. We therefore examined the actions of two kinds of D2-like DA receptor agonists, such as quinpirole (1–20 μm) and bromocriptine (10 μm), to study whether the D2-like DA receptor activation causes any effect on the membrane excitability of striatal cholinergic neurons. Unexpectedly, however, quinpirole elicited mixed results such as an inward current (7/17 cells), no response (3/17 cells), an inward current followed by an outward current (1/17 cells), and an outward current (6/17 cells), whereas bromocriptine caused no response in all cells tested (four cells). Moreover, combined application of quinpirole and sulpiride produced inconsistent results (data no shown). These results imply that the site of action of D2-like agents might be primarily at the presynaptic axon terminals of the LA cells. Further investigation is needed on this matter.
DISCUSSION
The present study demonstrates that DA induces a membrane depolarization/inward current in striatal cholinergic interneurons via activation of postsynaptic D1-like DA receptors. Another novel finding is that a depolarization/inward current evoked by D1 class receptor activation is caused by closing a resting K+ conductance and opening a nonselective cation conductance via an adenylyl cyclase-cAMP-dependent pathway. These results support the idea that D1-class receptors located on the neostriatal cholinergic interneurons might participate in enhancement of membrane excitability, thereby facilitating ACh release.
D1-class DA receptors (D1, D5) are known to activate adenylyl cyclase, whereas D2-like receptors (D2, D3, D4) inhibit, or are not coupled to, adenylyl cyclase. Our conclusion that DA acts on D1-like receptors on the LA neurons is based on the following observations. First, the effect of DA was blocked by the D1-like antagonist SCH23390 and mimicked by the D1-like agonist SKF38393 or DA plus sulpiride, a D2-like antagonist. Second, SCH23390 itself elicited an outward current with an opposite I–V relation to that obtained with DA or SKF38393 (data not shown). Third, our finding that forskolin, which stimulates adenylyl cyclase activity and increases cAMP levels, mimicked the effects of DA and SKF38393 and the inactive congener 1,9-dideoxyforskolin had no effects, provides further support for the involvement of D1-like receptors. Requirement of adenylyl cyclase activation for expression of the effects of SKF38393 was also confirmed by the fact that SQ22536, which decreases the adenylyl cyclase activity, reduced the SKF38393-induced current. In addition, D2-like agonists such as quinpirole and bromocriptine elicited inconsistent effects on the membrane excitability, suggesting that the site of action of the D2-like agents might be primarily at the presynaptic axon terminals of the striatal cholinergic interneurons.
The present study showed that membrane depolarization might involve at least two ionic mechanisms: blockade of resting K+conductance and activation of nonselective cationic conductances. The former appears to predominate in striatal LA cells, because the Ba2+-resistant component of the DA-induced current was only around −20 pA, whereas the control DA current was approximately −60 pA (Fig. 6). Reduction of Na+concentration in the external solution revealed that (1) the current was decreased with hyperpolarizing potentials and reversed at a potential close to the estimated EK: (2) this current was associated with a decrease in membrane conductance; and (3) the reversal potential shifted to less negative potentials in solutions of elevated K+, as expected for K+ conductances. A comparable enhancement of cell excitability, mediated by suppression of a K+conductance, has been suggested previously from experiments with cultured substantia nigra pars reticulata neurons (Kim et al., 1995), in which a D1 receptor agonist reduced an inwardly rectifying K+ conductance. However, this mechanism cannot apply to the data presented here because theI–V relationships did not show any anomalous rectification in the striatal LA cells and these have no detectable inward rectifier (Jiang and North, 1991; Aosaki and Kawaguchi, 1996).
Our data revealed an additional Ba2+-resistant component, considered to be a nonselective cation conductance because (1) the DA-induced current showed a linear I–Vrelation with a positive slope conductance and a reversal potential of around −40 mV in 10 of 23 cells and the Ba2+-resistant current reversed at −28 mV in one cell, and (2) the current component was associated with an increase in membrane conductance. However, in 10 of 11 cells the component displayed an atypical I–V relationship: between −35 and +20 mV, the current remained inward and showed a negative slope conductance. This cannot be attributed to activation of voltage-dependent Ca2+ channels because it was observed even in the presence of Cd2+ in the external solution. Therefore, this might be attributable to a space clamp error or to its own voltage-dependent property, because activation of the similar nonselective cation conductance by DA has been reported in LB-cluster neurons of Aplysia (Matsumoto et al., 1988). In this latter case, D1 receptor stimulation produced an increase in permeability, mainly to Na+, to elicit a cAMP-dependent inward current. This depolarizing response was also associated with negative slope conductance at more depolarized potentials. In addition, a similar atypical I–Vrelationship was reported for the ionic mechanism of an inward current elicited by muscarinic receptor activation (Haj-Dahmane and Andrade, 1996) and a DA-induced inward current (Shi et al., 1997) in prefrontal cortical neurons. It is noteworthy, however, that, unlike the striatal LA neurons, the DA-induced inward current observed in the prefrontal neurons was not sensitive to any of the classical DA agonists and antagonists (Shi et al., 1997). The mechanism proposed here (block of resting K+ conductance and activation of nonselective cationic conductances) has been proposed for other neurotransmitters (Benson et al., 1988; Larkman and Kelly, 1992;Guérineau et al., 1994, 1995; Dong et al., 1996; Kolaj et al., 1997). Taking into consideration recent studies showing that the striatal cholinergic interneurons possess D5 and, to a lesser extent, D1 receptors (Bergson et al., 1995; Yan et al., 1997), the specific findings described here, compared with those reported for actions of conventional D1 receptors, might stem from the unique properties of D5 receptors.
Recent microdialysis studies have consistently indicated that application of DA, whether in vivo or in vitro, increases net ACh release and that this is mediated by activation of D1-like receptors (Ajima et al., 1990; Bertorelli and Consolo, 1990; Consolo et al., 1992; Zocchi and Pert, 1993). However, there are several possible mechanisms of D1-like receptor-mediated control of ACh release. First, several in vivo studies have provided evidence of the importance of glutamate released from extrastriatal pathways such as in the parafascicular nucleus of the thalamus, cerebral cortex, and substantia nigra pars reticulata (Damsma et al., 1991; Consolo et al., 1996a,b; Abercrombie and DeBoer, 1997). Second, it has been demonstrated that local release of substance P from the terminals of medium spiny neurons, which contain D1-like receptors, increases the ACh release (Arenas et al., 1991; Guevara Guzman et al., 1993; Anderson et al., 1994; Steinberg et al., 1995; Khan et al., 1996). Finally, direct activation of the D1-like receptors on the cholinergic neurons might directly facilitate the ACh release (Di Chiara et al., 1994), as suggested in this paper. However, in vitro studies using striatal slice preparations similar to ours have generated conflicting results. Although SKF38393 stimulated K+-evoked ACh release in neostriatal slice preparations at concentrations of 1 μm (Stoof and Kebabian, 1982) and 10 μm (Gorell and Czarnecki, 1986), it was found to be without effect, up to 100 μm, byScatton (1982). Electrically evoked release of ACh has also been found to be reduced by D1-like antagonists (Consolo et al., 1987) or remain unchanged by D1-like selective agents (Dolezal et al., 1992; Tedford et al., 1992). These discrepancies might be attributable to differences in the experimental conditions applied and difficulties in controlling various related factors. In this regard, the studies of Login and colleagues (1995, 1996) are noteworthy. They developed a dissociated striatal neuron preparation to monitor fractional ACh efflux, with isolation of the activity of cholinergic neurons per se, and found that 50 μm (±)-SKF38393 increased the basal rate of release by 50% and that this was inhibited by a D1-like antagonist. Our observations, therefore, provide further support for their conclusion that ACh is secreted via direct activation of D1-like receptors on cholinergic cells and provide an explanation of how it might occur. The question now is whether this mechanism is feasible under physiological conditions.
Evidence has accumulated that most D1 and D5 DA receptors are located outside synaptic contacts formed by dopaminergic terminals (Bergson et al., 1995; Hersch et al., 1995; Caillé et al., 1996), suggesting that a nonsynaptic dopaminergic neuromodulation predominates in the striatum. Recent electrochemical measurements of DA efflux from synaptic clefts have shown that DA released by bursts of action potentials diffuses up to 12 μm from release sites reaching a homogeneous DA concentration of 0.2 to 1 μm in the extrasynaptic extracellular space before elimination by reuptake (Garris et al., 1994; Gonon, 1997). Because it is known that DA binds to D5/D1b receptors with an affinity that is highest among DA receptor subtypes (for example, 3- to 10-fold higher than that of DA binding to D1 receptors) (Seeman and Van Tol, 1994), this concentration range would be high enough to depolarize striatal cholinergic neurons. These are characterized by shallow resting membrane potentials (approximately −60 mV) and slow irregular but tonic (3–10 Hz) spontaneous activity, and therefore small (0.5–5 mV) EPSPs could easily trigger their action potentials (Wilson et al., 1990). Furthermore, activation of D1-like receptors has been reported to potentiate the NMDA component in striatal neurons (Cepeda et al., 1993), which might further stimulate ACh release, because an NMDA component of the EPSPs is already fully operative at rest in these neurons (Kawaguchi, 1992). Taking all the available data together, we propose that DA directly activates D1-like receptors (mainly D5), thus depolarizing the striatal cholinergic neurons by blocking resting K+ and opening nonselective cation conductances and making them ready to respond to EPSPs driven by thalamic, cortical, and nigral input.
Footnotes
This work was supported by the Frontier Research Program and Grants-in-Aid 07458217 (B) and 07279250 for Scientific Research in the Priority Area of “Functional Development of Neural Circuits” of the Ministry of Education, Science, Sports and Culture of Japan. We thank Ms. Naoko Wada for technical assistance.
Correspondence should be addressed to Dr. Toshihiko Aosaki, Laboratory for Neural Circuits, Bio-Mimetic Control Research Center, The Institute of Physical and Chemical Research (RIKEN), 2271–130, Anagahora, Shimoshidami, Moriyama-ku, Nagoya, Aichi 463–0003, Japan.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}