

Abstract
The physiological consequences of activating corticotropin-releasing factor receptor type 2 (CRF2) are not fully understood. The neuroanatomic distribution of this CRF receptor family member is consistent with roles in mediating the actions of CRF and similar ligands on food intake control and integrative aspects of stress-related behaviors. However, CRF2 expression in the adult rat is not influenced by stress, corticosterone (CORT), or food intake. Inimmature rat we have demonstrated striking downregulation of CRF2mRNA in hypothalamic ventromedial nucleus (VMH) after 24 hr of maternal deprivation, a paradigm consisting of both physiological/psychological stress and food deprivation. The current study aimed to distinguish which element or elements of maternal deprivation govern CRF2mRNA expression by isolating the effects of food intake and discrete maternal sensory cues on CRF2mRNA levels in VMH and in reciprocally communicating amygdala nuclei. In maternally deprived pups, CRF2mRNA levels in VMH and basomedial (BMA) and medial (MEA) amygdala nuclei were 62, 72, and 102% of control levels, respectively. Sensory inputs of grooming and handling as well as of the pups’ own suckling activity—but not food intake—fully restored CRF2mRNA expression in VMH. In contrast, all manipulations tended to increase CRF2mRNA levels in BMA of maternally deprived rats, and surrogate grooming increased CRF2mRNA expression significantly above that of nondeprived controls. CRF2mRNA expression was not influenced significantly by plasma adrenocorticotropic hormone (ACTH) and CORT levels. Thus, in the immature rat, (1) CRF2 expression is regulated differentially in hypothalamic and amygdala regions, and (2) CRF2mRNA levels in VMH are governed primarily by maternal or suckling-derived sensory input rather than food intake or peripheral stress hormones. These findings indicate a region-specific regulation of CRF2mRNA, supporting the participation of the receptor in neurochemically defined circuits integrating sensory cues to influence specific behavioral and visceral functions.
- corticotropin-releasing factor
- receptor
- CRF2
- ventromedial hypothalamus
- food intake
- stress
- rat
- neuroendocrine
Corticotropin-releasing factor (CRF) is the key mediator of the neuroendocrine response to stress (Vale et al., 1981). Release of hypothalamic CRF from the paraventricular nucleus (PVN) leads to elevations of plasma levels of the stress hormones adrenocorticotropic hormone (ACTH) and glucocorticoids (Vale et al., 1981; Sawchenko et al., 1993). In the CNS, CRF acts as a neuromodulator in discrete regions (Young et al., 1986; Valentino et al., 1991). In cortical and limbic circuits, CRF mediates the central response to stress and plays a role in anxiety, food intake, learning and memory, and neuronal excitation (Richard, 1993; Tsigos and Chrousos, 1994; Behan et al., 1995; Heim et al., 1997; Baram and Hatalski, 1998). CRF effects are mediated by the activation of G-protein-coupled, membrane-bound receptors (De Souza et al., 1985;Grigoriadis et al., 1993). Two major CRF receptors have been characterized, CRF1 and CRF2 (Chang et al., 1993; Perrin et al., 1993; Lovenberg et al., 1995b), and the latter receptor exists in at least two isoforms (Lovenberg et al., 1995a). The precise functions of these two receptors are not fully understood (Behan et al., 1996). Localization studies in both adult (Chalmers et al., 1995; Lovenberg et al., 1995b) and immature rats (Avishai-Eliner et al., 1996; Eghbal-Ahmadi et al., 1998) demonstrated distinct and mostly nonoverlapping distributions of the two receptors, suggesting functional diversity. In both adult and immature rats CRF2is highly expressed in VMH and in specific amygdala nuclei with reciprocal connections to this hypothalamic region. In VMH, CRF2 is a candidate mediator of the effects of CRF or related ligands on food intake (Spina et al., 1996), as suggested in recent studies that used selective CRF receptor antagonists and antisense oligonucleotides (Smagin et al., 1998). Whether CRF2 also is involved in stress-related functions of CRF has not been resolved.
Understanding the regulation of CRF2 may provide insight into the function of this receptor (Richard et al., 1996; Smagin et al., 1998). Previous studies from our laboratory have documented a striking downregulation of CRF2mRNA levels in VMH of immature rats after a 24 hr maternal separation, a paradigm depriving the pup from food, maternal contact, and maternal licking (considered necessary for the induction of micturition). Thus, maternal deprivation constitutes a complex physiological and psychological stressor as well as a fasting period. The goal of the current study was to distinguish the specific element or elements of the maternal deprivation paradigm responsible for the observed downregulation of CRF2mRNA. In particular, the experimental design dissected out potential regulation of CRF2 expression by stress [as measured by the activation of the hypothalamic–pituitary–adrenal (HPA) axis], by food intake, and by specific sensory cues derived from maternal–pup interactions and from the feeding procedure. CRF2mRNA levels were determined in VMH and in the intercommunicating CRF2mRNA-expressing BMA and MEA amygdala nuclei.
MATERIALS AND METHODS
Animals and tissue preparation. Timed-pregnancy Sprague Dawley rats (Zivic-Miller, Zelienople, PA) were maintained in National Institutes of Health-approved animal facilities on a 12 hr light/dark cycle with access to unlimited lab chow and water. Delivery was verified at 12 hr intervals, and the date of birth was considered day 0. Litters were culled to 12 pups and mixed among experimental groups; thus, effects of experimental manipulations were compared among littermates. Overall, 75 pups of both sexes participated in these experiments. Potential effects of circadian rhythms were addressed by initiating all experiments and by sacrificing animals between 8 and 10 A.M.; cages were undisturbed for 24 hr before experiments. All experimental procedures were approved by the Institutional Animal Care Committee and conformed to National Institutes of Health guidelines.
Experimental design. The overall strategy was to test the effect of selective reinstatement of isolated elements of maternal input on CRF2mRNA levels. Table1 provides the experimental groups, animal numbers per group, and mean animal weight gain/loss during the experimental period.
Characteristics of experimental groups and weight changes
Experiment 1 examined the regulation of CRF2mRNA in the immature rat by maternal sensory input. On postnatal day 8 thecontrol group (n = 10) was left untouched in home cages with the dam for the duration of the 24 hr experiment. These animals were sacrificed on the morning of day 9 within 45 sec of entry into the animal facility (Yi and Baram, 1994; Eghbal-Ahmadi et al., 1997; Yan et al., 1998).
The maternally deprived animals (n = 11) were taken to a room away from the dam, kept on a euthermic pad as a group, and maintained on the same light/dark schedule (Avishai-Eliner et al., 1995). This group otherwise was untouched and was sacrificed 24 hr later, on the morning of day 9.
Maternal contact only was provided to a third group (n = 14). This group remained in home cages, but food intake and maternal licking and grooming were prevented. This was achieved by deep anesthesia of the dam with a long-acting barbiturate (phenobarbital) at doses sufficient to eliminate milk ejection (Lincoln et al., 1973) as well as to result in stupor (a total of 129.2 ± 10 mg/kg per 24 hr, given intraperitoneally every 6–8 hr). The absence of milk in the pups’ stomachs was verified at several time points during the experiment, and weight changes were monitored (Table 1).
Maternal licking and grooming are critical elements influencing the neuroendocrine stress response in the developing rat (see Discussion). In addition, the resulting anogenital stimulation induces micturition (Suchecki et al., 1993). Surrogate maternal grooming was provided to the groomed pups (n = 7), which also were kept as a group during the 24 hr deprivation period. Anogenital stroking followed the method described by Suchecki et al., 1993. Briefly, a fine brush moistened with warm water was used every 8–10 hr to stimulate the anogenital area. Stroking resulted in urination within 30–45 sec.
The handled group (n = 10) provided a control for the human contact inevitable in the stroking maneuver. Rats, maternally deprived as a group, were subjected to human handling (without stroking) at the same frequency and for the same duration as the stroking, above. Handling was variably associated with induction of micturition in the pups.
Experiment 2 determined whether downregulation of CRF2mRNA levels after 24 hr maternal deprivation could be reversed by food intake. The procedures for the control and maternally deprived groups are as described in Experiment 1.
The fed, maternally deprived group (n = 11) received a milk-based diet via oral gavage. The diet consisted of diluted Carnation evaporated milk supplemented with corn oil to approximate the composition of rat milk (Messer et al., 1969). The caloric density (172 kcal/100 ml) and intake (7.74 kcal, 430 kcal/kg per day) were calculated to permit normal weight gain (Table 1). Milk was infused for 45 sec/hr during the light period and continuously during dark. Milk volume infused during the 24 hr deprivation period was 4.5 ml. The infusion used a 2.5 mm outer diameter plastic tubing connected to an infusion minipump (KD Scientific, Boston, MA) and involved positioning the tubing in the rat’s mouth to permit suckling. Tubing was inserted into a 6-mm-long cone-shaped plastic tip (Eppendorf) together with a plain gut surgical suture, positioned in the pup’s mouth, and secured by tying the suture thread around the neck. Glue secured the suture and knot in place. The procedure required 1–2 min. Pups were placed in individual containers, were left undisturbed for the duration of the experiment, and were observed to suckle on the plastic tip.
The sham-fed control group for the feeding paradigm, above (n = 12), underwent precisely the same procedure, but milk was not infused. These pups exhibited much more vigorous and consistent suckling than the fed group.
In both experiments all maternally deprived rats were kept euthermic (core temperature, 33–34°C) on a circulated water-based pad, and a 12 hr light/dark cycle was maintained. Experimental animals were weighed at the onset and termination of the experiments. Animals were rapidly decapitated, and trunk blood was collected for measurement of plasma ACTH and corticosterone (CORT) levels, using commercial radioimmunoassay (RIA) kits (INCSTAR, Stillwater, MN, and ICN, Costa Mesa, CA, respectively). Brains were rapidly dissected out onto powdered dry ice as described (Yi and Baram, 1994). Twenty-micrometer-thick coronal brain sections were cut, mounted on gelatin-coated slides, and stored at −80°C (Eghbal-Ahmadi et al., 1998). Because results for the control groups of both experiments did not differ and inter-animal variability was substantial, the data for control groups of these experiments were combined. This is also the case for the maternally deprived group (Table 1).
Probe preparation and in situ hybridization histochemistry (ISH). Preparation and labeling of the CRF2 riboprobe and of the ISH have been described previously (Eghbal-Ahmadi et al., 1997, 1998). Briefly, a plasmid containing the 461 base pair fragment of CRF2 cDNA (kindly provided by Dr. Lovenberg, Neurocrine Bioscience, La Jolla, CA) was linearized with HindIII (Lovenberg et al., 1995b). Radioactive antisense cRNA was synthesized by incorporating [35S]-CTP (New England Nuclear, Boston, MA). The probe was subjected to alkaline hydrolysis and purified by column chromatography [Select-D(RF), 5 Prime→3 Prime, Boulder, CO]. The specific activity of each probe was 1–3 × 106cpm/mg. Probe specificity to CRF2mRNA has been established (Chalmers et al., 1995; Eghbal-Ahmadi et al., 1998).
For ISH, sections spanning the coronal levels of the VMH and the amygdaloid nuclei that were studied were brought to room temperature, air-dried, and fixed in fresh 4% buffered paraformaldehyde for 20 min, followed by dehydration and rehydration through graded ethanols (Baram and Lerner, 1991). Sections were exposed to 0.25% acetic anhydride in 0.1 m triethanolamine, pH 8, for 8 min and were dehydrated through graded ethanols. Sections were air-dried and prehybridized for 1 hr at 55°C in a humidity chamber and then were hybridized overnight at 55°C with 1 × 106 cpm of35S-labeled ribonucleotide probe. After hybridization, the sections were washed in 2× SSC for 5 min at room temperature (1× SSC denotes 0.15 m NaCl and 15 mm trisodium citrate buffer, pH 7) and were digested with RNase (200 μg/ml RNase A; Calbiochem, LaJolla, CA) for 30 min at 37°C. Sections underwent successive washes (at 55°C) in 2× SSC and 1× SSC for 5 min, 0.25× SSC for 30 min, and in 0.1× and 0.03× SSC for 1 hr each, followed by dehydration through 100% ethanol. Sections were apposed to film (Hyperfilm β-Max, Amersham, Arlington Heights, IL) for 6–10 d, and representative ones also were dipped in NTB2 nuclear emulsion (Eastman Kodak, Rochester, NY) and exposed for 3–4 weeks.
Acquisition and quantitative analysis of CRF2mRNA ISH signal. Semiquantitative analysis was performed after ISH as previously described (Eghbal-Ahmadi et al., 1997, 1998; Hatalski et al., 1998). Briefly, digitized images of each brain section were acquired with a StudioStar scanner (AGFA, resolution 1200 × 1200 dots per inch) and analyzed by the ImageTool software program (version 1.25; University of Texas Health Science Center, San Antonio, TX). Densities were calibrated by using 14C standards and are expressed in nCi/gm after being corrected for background by subtracting the density of the hybridization signal over the dorsomedial hypothalamus immediately above VMH, a region devoid of CRF2mRNA (Chalmers et al., 1995; Eghbal-Ahmadi et al., 1998). For balanced comparison among the different experimental groups, four sections containing the core of the nucleus and demonstrating the highest optical density values were used from each brain for each region. The significance (p < 0.05, or as indicated) of observed quantitative differences among experimental groups was evaluated with one-way ANOVA or the unpaired Student’st test (with Welch’s correction for unequal variance, if appropriate), as indicated in the text.
RESULTS
CRF2mRNA expression in VMH is regulated by specific sensory input
As shown in Figure 1, CRF2mRNA levels in VMH of maternally deprived rat pups (180 ± 19.6 nCi/gm) were significantly lower than those of the control group (290 ± 40.3 nCi/gm; p = 0.017; Student’s t test). The presence of maternal contact, without food or active maternal cues, did not influence CRF2mRNA expression significantly; the contact group CRF2mRNA levels (215 ± 26 nCi/gm) were not statistically different from those of maternally deprived rats (p = 0.27) and tended to be lower than those of nondeprived controls (p = 0.1). In contrast,active surrogate maternal cues resulted in striking upregulation of CRF2 gene expression in VMH (Figs. 1,2). Anogenital stroking of maternally deprived rats—as well as just handling them—restored VMH CRF2mRNA expression to levels comparable to those of the nondeprived control group: 304 ± 60.3 and 286 ± 28.0 nCi/gm, respectively. These values were significantly higher than VMH–CRF2mRNA levels in maternally deprived rats (p = 0.02 and 0.003 when comparing the stroked and handled groups, respectively, with the maternally deprived group). Figure 2, showing dark-field photomicrographs of matched coronal sections at VMH level from control, from maternally deprived, and from stroked, maternally deprived rats, demonstrates the restoration of CRF2mRNA signal intensity in VMH of the deprived rats by surrogate maternal grooming.

CRF2mRNA expression in the VMH of immature rats subjected to selective physiological manipulations. A quantitative analysis of signal over VMH was achieved after ISH of matched coronal brain sections. Control, Animals were kept undisturbed in home cages; Deprived, pups were separated from the mother for 24 hr; Contact, pups were permitted contact with an anesthetized dam; Dep/Groom, maternally deprived animals were offered surrogate maternal grooming;Dep/HNDL, a deprived group was handled by a human as a control for the grooming; Dep/Fed, animals were gavage-fed an appropriate volume and calories of a milk-based diet;Dep/Sham-F, maternally deprived animals were undergoing the feeding procedure, but not receiving milk. *p< 0.05, significant difference from control; ♦ indicates significant difference from the maternally deprived group value. Values are expressed as means ± SEM. Group sizes (n) are found in Table 1.

Effect of physiological manipulations on CRF2mRNA levels in VMH of immature rats. Dark-field photomicrographs of coronal brain sections at the level of the VMH (encircled by the dotted line) after ISH that used a riboprobe directed against CRF2mRNA. Shown are a control rat (A), a rat after a 24 hr maternal deprivation (B), and a maternally deprived rat subjected to surrogate grooming (C). For orientation purposes the position of the third ventricle is indicated by a vertical dashed line. Note the localization of CRF2mRNA signal primarily over the dorsomedial portion of VMH. Scale bar, 70 μm.
CRF2mRNA expression is regulated differentially in BMA and MEA amygdala nuclei
CRF2mRNA expression in BMA was influenced significantly by the experimental manipulations (p < 0.0024; ANOVA). Although eliminating or providing active maternal cues influenced pups’ CRF2mRNA levels in BMA in a parallel trend to that found in VMH, the magnitude of these effects differed in these two CRF2-expressing regions (Figs. 3,4). Thus, after complete deprivation of pups from both active and passive maternal cues, BMA–CRF2mRNA levels were 57 ± 8.9 nCi/gm as compared with 78 ± 9.0 nCi/gm in nondeprived controls, not a significant reduction (p = 0.11). In contrast, providing recurrent active sensory input to deprived pups strikingly increased CRF2mRNA expression in BMA; grooming (via stroking) the pups resulted in CRF2mRNA levels of 134 ± 23.4 nCi/gm, significantly higher not only from those of the deprived group (p = 0.0007) but also from levels of “normally” stimulated, i.e., nondeprived controls (p = 0.04). The less intense active stimulation generated by the handling of maternally deprived pups led to BMA–CRF2mRNA levels of 104 ± 13.4 nCi/gm, which were significantly higher than those of deprived (p = 0.004), but not of control, groups (p = 0.11). Finally, passive contact with an anesthetized dam resulted in modest changes in CRF2mRNA expression in BMA; the levels (86 ± 12.1 nCi/gm) tended to be higher than those of deprived rats (p = 0.06) and did not differ appreciably from those of controls (p = 0.61).

CRF2mRNA expression in BMA of immature rats subjected to selective physiological manipulations. Quantitative analysis of signal over the BMA was achieved after ISH of matched coronal brain sections. Control, Animals kept undisturbed in home cages; Deprived, pups separated from the dam for 24 hr; Contact, pups permitted contact with an anesthetized dam; Dep/Groom, maternally deprived animals offered surrogate grooming; Dep/Hndl, a deprived group handled by a human as a control for the grooming;Dep/Fed, animals gavage-fed an appropriate volume and calories of a milk-based diet; Dep/Sham-F, maternally deprived animals undergoing the feeding procedure, but not receiving milk. *p < 0.05, significant difference from control; ♦p < 0.05, significant difference from the maternally deprived group; •p = 11. Values are expressed as means ± SEM. Group size is found in Table1.

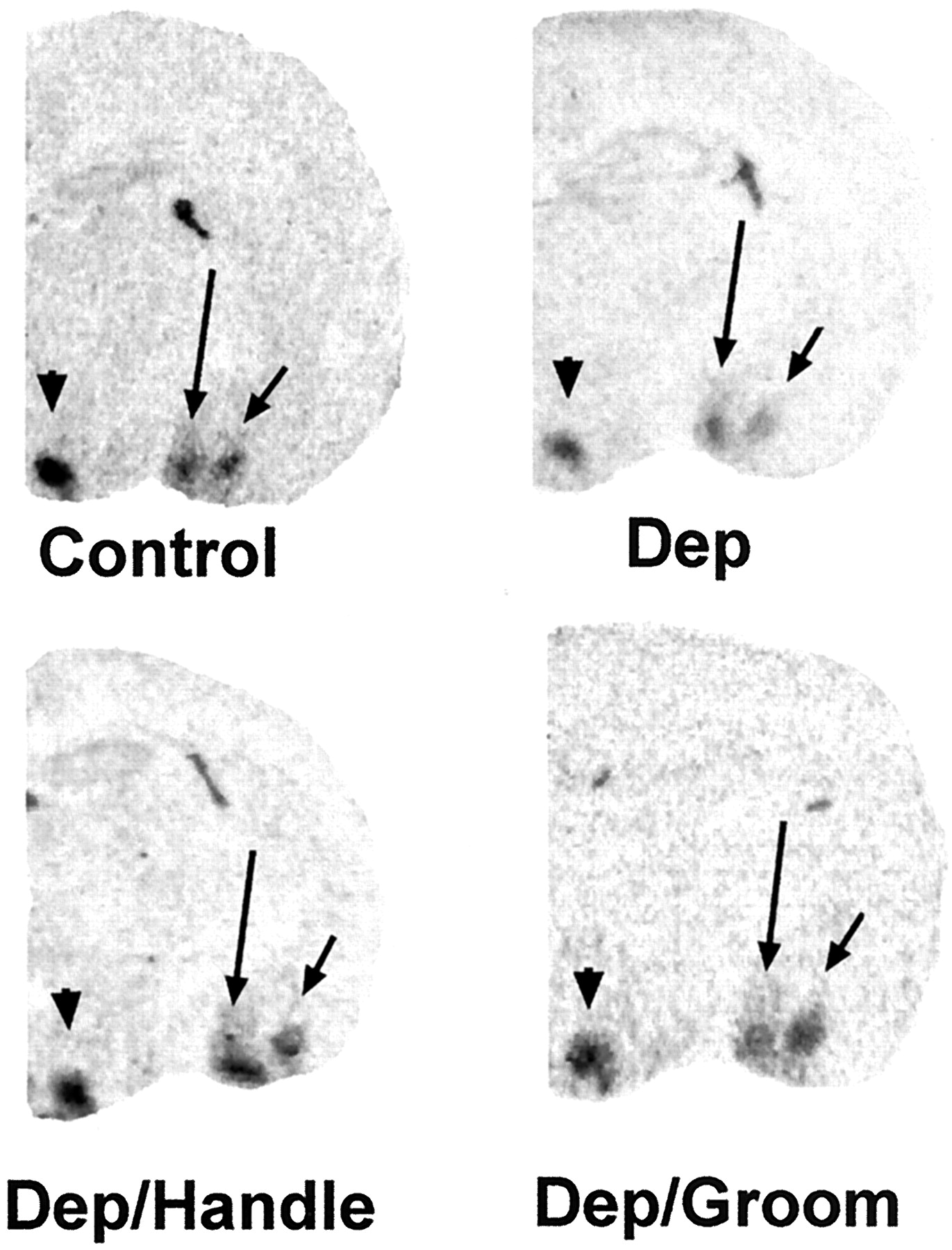
Differential regulation of CRF2mRNA expression in specific hypothalamic and amygdala regions by discrete sensory input. Shown are photomicrographs of coronal brain sections at the level of VMH (arrowhead), MEA (long arrow), and BMA (short arrow). Sections were subjected to ISH for CRF2mRNA. Decreased signal over VMH and BMA is apparent in sections from a maternally deprived rat (Dep) as compared with control levels. Surrogate grooming (Dep/Groom) or handling by a human (Dep/Handle) enhanced the CRF2mRNA signal over VMH and BMA. No effect of any of the experimental manipulations on CRF2mRNA expression in MEA is evident (see also Fig.5).
In contrast to robust alterations of CRF2mRNA expression induced by maternal input manipulations in VMH and BMA, CRF2mRNA signal over MEA was not affected significantly by treatment (F = 0.76; p = 0.6, ANOVA). As is evident from Figures 4 and 5, CRF2mRNA levels were remarkably independent of maternal or surrogate sensory cues, ranging from 114 ± 16.4 nCi/gm in deprived–handled rats to 149 ± 23.3 nCi/gm in the maternally deprived group. CRF2mRNA signal over MEA of control pups was at least as high as that in BMA (146 ± 20.6 vs 78 ± 9.0 nCi/gm) and well above expression levels in most other amygdala nuclei (Eghbal-Ahmadi et al., 1998), excluding potential difficulty in detecting the signal as a source of apparent homogeneity of CRF2mRNA expression in MEA.

CRF2mRNA expression in MEA of immature rats subjected to physiological manipulations. Quantitative analysis of signal over MEA was achieved after ISH of matched coronal brain sections. Group name abbreviations are as in Figure 1. No significant differences among values were observed (p > 0.05, ANOVA). Values are expressed as means ± SEM. The animal number for each group is found in Table 1.
Influence of food intake and the feeding procedure on CRF2mRNA expression
Administration of milk-based formula at the caloric density and volumes described earlier was effective in permitting normal growth. The gavage-fed group gained an average 10.8% of body weight during the experiment, whereas the sham-fed group lost 7.5% of body weight (Table1). These values are similar to the control group’s weight gain (16.1%) and to the maternally deprived group’s weight loss (5.3%), respectively (Table 1).
CRF2mRNA levels in VMH of maternally deprived gavage-fed rats (209 ± 22.7 nCi/gm) were intermediate between that of maternally deprived and control groups (see Fig. 1). Interestingly, CRF2mRNA levels in VMH of the sham-fed group (247 ± 26.9 nCi/gm) were significantly higher than those of the deprived group (p = 0.02) and indistinguishable from those of controls (p = 0.84). As mentioned above, the sham-fed pups—but not those fed—were observed to engage in persistent and vigorous suckling on the plastic tip. The specific influence of food intake per se and of the feeding procedure on CRF2mRNA levels in BMA was less conclusive (see Fig. 3). Because of the relatively modest effect of 24 hr maternal deprivation on transcript levels, sham-fed rats (79 ± 8.3 nCi/gm) did not differ from either control or maternally deprived groups (p = 0.92 and 0.07 vs control and deprived groups, respectively). Food intake led to a significant elevation of CRF2mRNA expression (84 ± 7.8 nCi/gm) compared with the maternally deprived group (p = 0.03). As described above for the other manipulations, neither food intake nor the feeding procedure influenced CRF2mRNA levels in MEA significantly, compared with either control or maternally deprived pups (see Fig. 5).
Plasma ACTH and CORT levels after maternal deprivation and selective reintroduction of sensory cues, feeding procedure, and food intake
The expected changes in peripheral stress hormones were observed after maternal deprivation; ACTH levels did not differ among groups (Fig. 6A), whereas plasma CORT in the maternally deprived group was elevated when compared with the control group (4.6 ± 0.7 vs 0.97 ± 0.14 μg/dl;p < 0.001). Neither reinstatement of maternal contact nor human stroking or handling influenced plasma ACTH or CORT to any significant degree as compared with levels observed with maternal deprivation per se (Fig. 6B). Interestingly, plasma CORT levels in rats subjected to the feeding procedure—regardless of actual food intake—were elevated significantly above those of both the control and the maternally deprived groups (Fig.6B).

Plasma stress hormone levels in immature rats subjected to maternal deprivation, and selective reinstitution of maternal sensory cues and feeding. A, ACTH plasma levels did not differ among groups. B, Plasma CORT levels of all maternally deprived groups were elevated significantly as compared with controls (*p < 0.01). In addition, CORT levels in groups subjected to the feeding procedure were significantly higher than those of both control and maternally deprived groups (♦p < 0.01). See Figure 1 for abbreviations.
DISCUSSION
A previous study from this laboratory reported a robust (62%) reduction of CRF2mRNA levels in VMH of immature rats after a 24 hr maternal deprivation, providing the first demonstration of the physiological regulation of the expression of this receptor in the CNS (Eghbal-Ahmadi et al., 1997). The current study investigated the mechanism or mechanisms responsible for this downregulation of CRF2mRNA expression as a means for trying to better understand the functional role or roles of the receptor. In addition, the current study examined the regulation of CRF2mRNA abundance in intercommunicating and CRF2mRNAexpressing amygdala nuclei.
The major findings of the current study were (1) CRF2mRNA levels in VMH were restored to control values by sensory cues mimickingactive maternal input (licking and grooming), but not bypassive maternal contact; (2) in VMH, food intake in quantities and intake pattern approximating “normal” suckling resulted in CRF2mRNA levels intermediate between maternally deprived and control rats; however, sham-feeding, leading to vigorous persistent suckling, restored CRF2mRNA expression to control levels; (3) CRF2mRNA expression in BMA was highly upregulated by active sensory input simulating maternal grooming and was less influenced by maternal deprivation than expression in VMH; (4) Expression of CRF2mRNA in MEA was not influenced by any alteration of the physiological parameters tested. These findings also indicate that CRF2mRNA is regulated differentially in VMH, BMA, and MEA, three interconnected regions of a hypothalamic–limbic–autonomic circuit.
High expression of CRF2mRNA has been demonstrated in VMH of the adult and developing rat (Lovenberg et al., 1995b; Eghbal-Ahmadi et al., 1998), but the functions of CRF2 in this region are not fully understood. VMH has been documented to be an important participant in the regulation of food intake and energy balance (Rohner-Jeanrenaud, 1995) and in the interactions of these with the HPA axis and circadian rhythm (Dallman, 1984; Choi et al., 1996). Several recent lines of evidence have implicated VMH–CRF2 in the regulation of food intake; Smagin et al. (1998) found that reducing CRF2mRNA levels in VMH by using the antisense oligonucleotide strategy diminished the anorexic effects of both CRF and urocortin. In addition, the authors showed that selectively blocking the other member of the CRF receptor family, CRF1, did not alter the anorexia induced by CRF, suggesting that this action was mediated by CRF2. A second group of investigators (Richard et al., 1996) studied CRF2mRNA in VMH of genetically obese rats. The authors demonstrated a reduction of VMH–CRF2mRNA levels in these rats when compared with lean controls after an overnight fast. The implications of these findings vis-á-vis potential functions of CRF2 in food intake and/or energy balance have not been resolved fully (Rohner-Jeanrenaud et al., 1989; Rohner-Jeanrenaud, 1995).
Modulation of CRF2mRNA expression by experimental manipulations that approximate physiological conditions may provide an important tool for understanding the function of this receptor in specific brain regions during discrete developmental ages and time frames. It is recognized, however, that the functional significance of changes in CRF2mRNA levels requires further examination, because additional post-transcriptional regulatory steps may lead to an incomplete correlation between mRNA and receptor–protein levels (and ligand binding). Previous work from this laboratory demonstrated high and selective expression of CRF2mRNA in VMH of the developing rat starting in utero (Eghbal-Ahmadi et al., 1998). In addition, transcript levels remained high and relatively constant throughout development, suggesting continuous and important functions for the CRF2 protein. Indeed, using the strategy of profound perturbation of the physiological milieu of the immature rat—24 hr of maternal deprivation—robust downregulation of CRF2mRNA levels in VMH was demonstrated (Eghbal-Ahmadi et al., 1997). Maternal deprivation is a composite of several interventions; food intake and the associated active suckling by the immature rat, passive maternal contact, and active licking and grooming by the dam are all eliminated. The role of each of these components of maternal input in influencing CRF2mRNA levels in VMH and in reciprocally connected CRF2-expressing amygdala regions of the immature rat has been the focus of the current study.
As shown in Figures 1 and 2, maternal deprivation resulted in a significant downregulation of CRF2mRNA expression in VMH. Selective reintroduction of the elements of maternal input suggested that the regulation of CRF2mRNA expression in this region is complex. Surrogate maternal licking and grooming enhanced CRF2mRNA levels of maternally deprived animals significantly, resulting in a control-like expression pattern. The maneuver of anogenital stroking mimics maternal grooming, considered necessary to induce micturition in the neonatal rat (Suchecki et al., 1993). Because this maneuver inevitably involves handling by a human, a control group was handled at the same intervals and for the same duration, but without anogenital stimulation. Interestingly, both manipulations increased CRF2mRNA levels in VMH of maternally deprived rats to control levels. Several possible explanations may account for this finding; first, sensory input either from the anogenital region or from human handling may alter CRF2mRNA expression via converging pathways, as discussed below. Alternatively, because handling frequently led to urination, neuronal signals resulting from the micturition process itself—rather than the instigating sensory stimuli—may govern CRF2mRNA expression.
Supporting the important influence of sensory input on CRF2mRNA expression in VMH was the striking effect of the sham-feeding procedure. This procedure, in which plastic tips were placed in the pups’ mouths, stimulated suckling. Although pups receiving food suckled periodically, the sham-fed group was distinguished by frequent and vigorous suckling. Thus, the orosensory input generated by the pups’ own actions may have participated in upregulating CRF2mRNA levels in VMH.
The effect of actual food intake on CRF2mRNA levels in VMH of maternally deprived rats was surprising. In VMH, reinstitution of food intake led to intermediate levels of CRF2mRNA, between those of maternally deprived and control rats. The caloric density and total volume of the provided diet approximated the daily requirement of 8-d-old rats and resulted in the expected weight gain, suggesting an appropriate energy intake (Table 1). Core temperatures of the artificially fed pups were in the range documented for littermates immediately on removal from home cages, indicating that the experimental conditions provided a euthermic environment without excessive heat loss. Therefore, the lack of complete restitution of CRF2mRNA expression in VMH of artificially fed maternally deprived rats was not attributable to inadequacy of actual food intake and suggested that other factors influenced CRF2mRNA levels under these conditions.
Several aspects of the feeding procedure may have influenced CRF2mRNA expression. For example, the differential suckling activity between sham-fed and truly fed groups was noted above. In addition, the procedure resulted in significantly higher plasma CORT levels in both feeding groups as compared with control and maternally deprived rats (Fig. 6B), suggesting that the procedure provided additional stress. However, plasma ACTH levels in these groups was not elevated when compared with either control or maternally deprived pups [and are consistent with those reported by others who used the same RIA kit (Suchecki et al., 1993; Van Oers et al., 1998)]. This fact suggests that the complex feeding procedure increased adrenal sensitivity to CORT secretagogues in an additive manner to the established effect of maternal deprivation itself (for review, see Suchecki et al., 1993). Although food intake has been shown by the same authors to diminish adrenal responsiveness to stress, plasma CORT levels did not differ in our hands between the truly fed and sham-fed groups, suggesting that any effect of food intake may have been overridden.
In addition, rats subjected to the feeding procedure were kept in individual containers, whereas members of each of the other experimental groups were kept together. Nevertheless, it is considered unlikely that individual housing contributed to the increased plasma CORT levels of the artificially fed and sham-fed groups. Previous work from this laboratory (Avishai-Eliner et al., 1995) has shown that morning plasma CORT levels of maternally deprived 9-d-old rats kept in individual containers (6.1 ± 1.1 μg/dl) were not higher than those of group-deprived rats (7.5 ± 1.0 μg/dl) and were close to those observed in the current experiments (5.1 ± 0.9 μg/dl). In fact, in the earlier study, individually deprived rats had a smaller peak plasma CORT level after an additional stressor than did group-deprived littermates (13.9 ± 1.3 vs 19.4 ± 1.2 μg/dl, respectively). These values are consistent with CORT levels observed for pups subjected to the feeding procedures in the current studies (13.8 ± 2.8 and 17.1 ± 2.2 μg/dl for fed and sham-fed groups, respectively). This is best interpreted to indicate an additive effect of the feeding procedure and of maternal deprivation stress on adrenal sensitivity.
Indeed, the stress-related element of the maternal deprivation paradigm and of the feeding procedures may account for the significant variability of CRF2mRNA expression among members of the same experimental group. Individual variability in responses to subacute and chronic stress have been well established in the developing rat (Gilles et al., 1996) and human (Lewis, 1992) and have been shown to emerge on the exposure of individuals to a modulation of maternal–pup interactions (Gilles et al., 1996). This fact may have contributed to the apparent partial effects of maternal deprivation and selective restitution of maternal input in the current experiments, as described for VMH above and for BMA below. However, no correlation between plasma levels of peripheral stress hormones and CRF2mRNA expression in VMH, BMA, and MEA was evident in the current experiments. This finding is consistent with that of Makino et al. (1997), who failed to alter CRF2mRNA expression in PVN by either injection stress or CORT administration. A similar lack of stress effect on CRF2mRNA levels in VMH has been documented by Richard in both lean and obese rats (Richard et al., 1996).
In the current study, CRF2mRNA expression was influenced by maternal deprivation and by selective restoration of sensory cues and food not only in VMH but also in reciprocally interconnected regions rich in CRF2mRNA (Canteras et al., 1994; Lovenberg et al., 1995b; Eghbal-Ahmadi et al., 1998). In BMA the most striking influence on CRF2mRNA expression was the upregulation—above both deprived and control levels—induced by anogenital stroking (see Fig.3). These findings suggest that a more specific sensory stimulus—i.e., stroking—or micturition-related anogenital input may influence CRF2mRNA levels in BMA, as compared with the less selective somatosensory stimuli sufficient to upregulate CRF2mRNA expression in VMH. Supporting the regional specificity of CRF2mRNA expression, levels of this transcript in MEA did not vary significantly with any of the manipulations or conditions examined in the present studies. Thus, the factors influencing CRF2mRNA levels in MEA of the immature rat remain to be explored.
VMH, BMA, and MEA form a reciprocally interconnected series of links in an important integrative circuit (Fig.7). BMA interconnects with VMH (Canteras et al., 1994; Petrovich et al., 1996) and projects heavily to central amygdala nucleus (CEA), a key integrating region of the stress response and a major efferent source for brainstem and hypothalamic autonomic nuclei (Pitkanen et al., 1997; Swanson and Petrovich, 1998). Petrovich et al. (1996) suggested a role for BMA–hypothalamic pathways for both gustatory and other sensory processing, proposing a BMA–CEA–neuroendocrine information flow for gustatory input and a BMA–VMH–neuroendocrine information flow involved with more general unimodal or polymodal sensory input. Thus, in the context of the current studies, CRF2 in BMA and VMH may participate in the transmission and integration of suckling, feeding, and other sensory–tactile inputs to amygdaloid motivational and autonomic control areas (e.g., CEA).

Schematic of a proposed neurochemically defined circuit, using CRF or similar ligand and neurotransmitter and CRF2 as the receptor. Quantitative changes in CRF2mRNA expression may lead to altered neurotransmission in this circuit, which is involved in the transduction and integration of somatosensory, gustatory, and visceral signals and the neuroendocrine stress response. Blue frames indicate CRF2mRNA expression. Red shading over a region indicates the presence of CRF-expressing neurons. Red arrows denote established CRF-containing pathways.CEA, Central amygdaloid nucleus–bed nucleus of the stria terminalis continuum; NTS, nucleus of the solitary tract; PBN, parabrachial nucleus; SCN, suprachiasmatic hypothalamic nucleus.
Indeed, an interesting characteristic of the selective distribution of CRF2 expression in the rodent brain is the presence of CRF2-containing neurons and CRF terminals in regions involved in the micturition reflex arc and its regulation by stress and circadian rhythms (Fig. 7). Previous data from this laboratory showed high CRF2mRNA levels in thalamic paraventricular nucleus (PVT) of the adult (Eghbal-Ahmadi et al., 1998). CRF2mRNA has been observed in PVT of the immature rat, but low transcript levels and the neuroanatomic features of PVT prohibited an ISH-based quantitative analysis of effects of experimental manipulations tested in the current study on levels of PVT–CRF2mRNA in the developing rat. PVT receives monosynaptic neuronal input from Barrington’s nucleus in the pontine tegmentum, the brainstem micturition center (Otake and Nakamura, 1995) (Fig. 7). Axon collaterals of these same neurons project to the spinal cord micturition center at L6 (Otake and Nakamura, 1996). Importantly, a substantial subset of neurons projecting from Barrington’s nucleus to PVT contains CRF and may influence the activity of PVT via CRF2 receptors. [CRF2mRNA is highly expressed in Barrington’s nucleus, and the expression of the peptide in this nucleus is regulated by stress (Imaki et al., 1991).] PVT projects to a number of regions involved in circadian rhythms, such as the suprachiasmatic nucleus and VMH (Moga et al., 1995). PVT also projects heavily to CEA (Moga et al., 1995). Recently, PVT has been implicated in modulating stress-induced activation of the HPA axis, possibly via inhibiting CEA (Bhatnagar and Dallman, 1998). Interestingly, CEA sends a robust projection of CRF-expressing neurons to PVT (Otake and Nakamura, 1995), perhaps exerting their effects via the activation of CRF2 receptors found in this region. Finally, PVT receives afferents from MEA, a nucleus integrating chemosensory stimuli relayed by genital somatosensory inputs (Canteras et al., 1995). Again, a subset of MEA neurons express CRF, but it has not been determined whether these are the source of afferents to PVT (Canteras et al., 1995).
Based on the results of the current studies, combined with the neuroanatomic information presented here, it is proposed that CRF2 expressed in VMH, BMA, and MEA participates in a neurochemically defined neuronal circuit that uses CRF or related ligands as neurotransmitters and functions via CRF2receptor activation, as shown in the schematic (Fig. 7). Information flow from somatosensory input reaches MEA and BMA and is relayed to “defensive” medial hypothalamic centers (Swanson and Petrovich, 1998) and to CEA for further integration. CEA, the central integrator of autonomic regulation, also receives visceral input from the nucleus of the solitary tract via the CRF-containing parabrachial nucleus (Swanson et al., 1983; Jia et al., 1994) and modulates central components of the HPA axis, further influenced by input from PVT (Bhatnagar and Dallman, 1998). Reciprocal interactions among VMH, PVT, and CEA as well as projections outside the circuit (e.g., to suprachiasmatic nucleus) enable the integration of this information within the context of circadian rhythms as well as further segregation and sorting of signals (Swanson and Petrovich, 1998). Circuit output to behavioral and visceral effectors probably does not involve CRF2-expressing neurons.
The studies reported here probed the regulation of CRF2mRNA expression in the neonatal rat, and it is conceded that CRF2mRNA regulation may differ in the adult. CRF2mRNA expression in most brain regions, including VMH and BMA, is relatively constant throughout postnatal development, suggesting persistent and age-independent functions for this receptor (Eghbal-Ahmadi et al., 1998). This contrasts with the developmental profile of CRF1mRNA, which shows dramatic quantitative variations with age (Avishai-Eliner et al., 1996). However, it is recognized that several components of rat neuroendocrine stress response are under maternal regulation during the developmental period studied here (Stanton et al., 1988; Levine et al., 1992; Rosenfeld et al., 1992; Suchecki et al., 1993), so that CRF2mRNA expression may be regulated by maternally derived cues to a larger extent during this formative and “plastic” epoch of neuronal circuit development than during adulthood. For example, maternal presence is widely considered to account for low plasma CORT and relatively low-magnitude hormonal responses to stress during the 4–15 postnatal days (Stanton et al., 1987). Maternal deprivation enhances basal and stress-induced CORT levels, as found also in the current study. However, elevated plasma CORT levels in the absence of increased ACTH levels after maternal deprivation are consistent with maternal suppression of adrenal sensitivity to ACTH. Indeed, discrete aspects of maternal input have been shown to influence the sensitivity of specific elements of the developing neuroendocrine stress response to noxious or hormonal stimuli (Suchecki et al., 1993). Contact with an anesthetized dam eliminated an enhanced CORT response to subsequent stress in 12-d-old rat pups (Stanton et al., 1987), whereas feeding maternally deprived pups suppressed both basal and stress-induced plasma CORT secretion induced by maternal deprivation (Suchecki et al., 1993). In addition, evidence for the powerful modulatory effects of maternal anogenital grooming (stimulating micturition) on ACTH and CORT responses has been documented (Suchecki et al., 1993). The current study suggests that CRF2 may participate in neurochemically defined circuits integrating maternal (and orosensory) cues, micturition, and reduction of the HPA axis sensitivity to stress.
In summary, CRF2mRNA expression in select hypothalamic and amygdala regions of the neonatal rat is influenced differentially by sensory cues provided by the dam and those related to feeding as well as by food intake. These findings may suggest that, at least during the developmental period studied, CRF2 may function in integrating these inputs into neuronal somatovisceral loops. In addition, the differential regulation of CRF2mRNA expression in the regions studied indicates that the functions of CRF2 may be region- and/or circuit-specific.
Footnotes
This work was supported by National Institutes of Health Grants NS 28912 and HD 34975. Critical comments by Drs. Christine Gall and James Fallon are appreciated.
Correspondence should be addressed to Dr. Tallie Z. Baram, Departments of Pediatrics and Anatomy and Neurobiology, Medical Science I, 4475, University of California at Irvine, Irvine, CA 92697-4475.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}