

Abstract
In lower vertebrates, retinal ganglion cells (RGCs) can regenerate their axons and reestablish functional connections after optic nerve injury. We show here that in goldfish RGCs, the effects of several trophic factors converge on a purine-sensitive signaling mechanism that controls axonal outgrowth and the expression of multiple growth-associated proteins. In culture, goldfish RGCs regenerate their axons in response to two molecules secreted by optic nerve glia, axogenesis factor-1 (AF-1) and AF-2, along with ciliary neurotrophic factor. The purine analog 6-thioguanine (6-TG) blocked outgrowth induced by each of these factors. Previous studies in PC12 cells have shown that the effects of 6-TG on neurite outgrowth may be mediated via inhibition of a 47 kDa protein kinase. Growth factor-induced axogenesis in RGCs was accompanied by many of the molecular changes that characterize regenerative growth in vivo, e.g., increased expression of GAP-43 and certain cell surface glycoproteins. 6-TG inhibited all of these changes but not those associated with axotomy per se, e.g., induction of jun family transcription factors, nor did it affect cell survival. Additional studies using RGCs from transgenic zebrafish showed that expression of Tα-1 tubulin is likewise stimulated by AF-1 and blocked by 6-TG. The purine nucleoside inosine had effects opposite to those of 6-TG. Inosine stimulated outgrowth and the characteristic pattern of molecular changes in RGCs and competitively reversed the inhibitory effects of 6-TG. We conclude that axon regeneration and the underlying program of gene expression in goldfish RGCs are mediated via a common, purine-sensitive pathway.
- regeneration
- axon
- retinal ganglion cell
- GAP-43
- E587 antigen
- L1
- neurolin
- DM-GRASP
- reggie-2
- Tα-1 tubulin
- CNTF
- inosine
- optic nerve
- zebrafish
After optic nerve injury, goldfish retinal ganglion cells (gfRGCs) regenerate their axons and reestablish functionally appropriate connections within 1–2 months (Sperry, 1963;Jacobson and Gaze, 1965; Murray, 1976). During this process, gfRGCs undergo dramatic structural and molecular changes (Murray and Forman, 1971; Giulian et al., 1980; Grafstein, 1986). The latter include increased expression of the membrane phosphoprotein GAP-43 (Benowitz et al., 1981, 1983; Skene and Willard, 1981; Heacock and Agranoff, 1982;Perry et al., 1987), certain tubulin isotypes (Burrell et al., 1978;Bormann et al., 1998; Hieber et al., 1998), intermediate filament proteins (Quitschke et al., 1980; Hall et al., 1990; Glasgow et al., 1992), cell surface glycoproteins (Bastmeyer et al., 1990; Vielmetter et al., 1991; Paschke et al., 1992; Ankerhold et al., 1998), transcription factors (Herdegen et al., 1993), and other proteins (Schulte et al., 1997; Lang et al., 1998; Ballestero et al., 1999). These changes could represent downstream consequences of divergent signaling pathways activated by multiple growth factors, or at the other extreme, there could be a “master switch” onto which multiple pathways converge to control a constellation of genes required for axon growth.
Two molecules secreted by goldfish optic nerve glia stimulate gfRGCs to regenerate axons in culture (Schwalb et al., 1995). The more potent of these, axogenesis factor-1 (AF-1), is a small molecule (<1 kDa), whereas AF-2 is a less active, larger polypeptide. Because AF-1 and AF-2 are present in high concentrations in the optic nerve, they may represent physiological effectors of axon regeneration in vivo (Schwalb et al., 1996). Ciliary neurotrophic factor (CNTF) induces rat RGCs to regenerate their axons (Cui et al., 1999; Jo et al., 1999), and one issue examined here was whether CNTF also acts on gfRGCs.
In pheochromocytoma 12 (PC12) cells, NGF and other growth factors induce the rapid activation of a 47–50 kDa serine-threonine kinase (Rowland-Gagne and Greene, 1990). The purine analog 6-thioguanine (6-TG) blocks neurite outgrowth and the activity of this NGF-sensitive (N-) kinase in parallel (Volonte et al., 1989). The effects of 6-TG are highly specific and do not include changes in other signal transduction elements [e.g., Ras and mitogen-activated protein (MAP) kinase], immediate-early gene induction, or changes in cell survival (Volonte et al., 1989; Greene et al., 1990; Batistatou et al., 1992). In gfRGCs, axon regeneration induced by AF-1 is likewise blocked by 6-TG, whereas the purine nucleoside inosine stimulates extensive outgrowth and reverses the inhibitory effects of 6-TG (Benowitz et al., 1998). This suggests that gfRGCs, like PC12 cells, use a purine-sensitive kinase in axon outgrowth. However, gfRGCs can be stimulated to extend axons by several factors, and it is not known whether they all act via the same purine-sensitive mechanism.
Our results show that all agents that stimulate axon outgrowth and the underlying constellation of molecular changes in RGCs act via a common, purine-sensitive pathway.
MATERIALS AND METHODS
Retinal ganglion cell cultures. All procedures were approved by Children's Hospital Animal Care and Use Committee. As described previously (Schwalb et al., 1995), Comet variety goldfish (Mt. Parnell Fisheries, Mt. Parnell, PA) were dark-adapted, anesthetized by hypothermia, and killed by cervical transection. Retinas were dissected under sterile conditions, digested with papain (Worthington, Freehold, NJ) in HEPES-buffered L-15 media (Life Technologies, Gaithersburg, MD) containing l-cysteine (Sigma, St. Louis, MO) at room temperature (RT), and subjected to a series of trituration and sedimentation steps that resulted in a ganglion cell-enriched suspension (Schwartz et al., 1981). For standard bioassays, ∼5 × 104 RGCs in 50 μl were plated in poly-l-lysine (Sigma)-coated wells of a 24-well tissue culture dish (Costar, Cambridge, MA) containing experimental or control samples in medium E, an L-15-based medium supplemented with progesterone, selenium, 3,3′-5-triodo-l-thyronine, apotransferrin, catalase, superoxide dismutase, BSA (type V, with fatty acids), gentamycin, insulin, and HEPES (all from Sigma), with a final volume of 400 μl. Samples were plated in quadruplicate in randomly assigned positions. Each experiment included four wells of a positive control (previously validated AF-1 at 20–30% concentration) and a negative control (medium E in L-15). For immunocytochemistry, ∼5 × 104 RGCs in 25 μl were plated in poly-l-lysine-coated wells of an eight-well glass chamber slide (Lab-Tek II; Nunc, Naperville, IL).
Quantitation of neurite outgrowth. Unless noted otherwise, cultures were evaluated after 6 d by an observer blind to the experimental conditions in each well. Neurite outgrowth was assessed in 150 consecutive RGCs per well by the use of inverted phase illumination (400× magnification). RGCs constitute 50–70% of the cells in these cultures, and as verified previously by retrograde labeling, they are readily distinguished by their relatively large size (12–17 μm diameter), round or oval shape, phase-bright appearance, and extension of one to two processes of uniform caliber (Schwartz et al., 1981; Schwalb et al., 1995). Unless noted otherwise, neurite outgrowth is operationally defined as the fraction of cells with axons >5 cell diameters in length averaged over the four replicate wells. Data were normalized by subtracting the level of growth found in negative controls (usually 3–5%) and then dividing by the background-corrected growth of the positive controls (usually in the range of 20–40%). All results are presented ± SEM and are representative of at least three independent experiments. The program used to graph the results (Cricket Graph; Computer Associates) does not show error bars if SEM is <2%.
Factors tested. Inosine (Sigma) and CNTF (Promega, Madison, WI) were purchased commercially. AF-1 and AF-2 were prepared from conditioned medium (CM) containing factors secreted from goldfish optic nerve glia. Optic nerves were cut into 1–2 mm fragments and incubated in saline for 3–4 hr. The resultant CM was filter-sterilized and subjected to centrifugal ultrafiltration using filters with a molecular weight cutoff of 3 kDa (Amicon, Beverly, MA) and then 1 kDa (Filtron). The low-molecular weight fraction that passed through both filters was used as a crude source of AF-1. The neurite-promoting activity in the <1 kDa fraction appears to represent a single molecular species, because only a single peak of activity is detected after further separation by several chromatographic procedures (Schwalb et al., 1995) (N. Irwin, Y. Li, and L. I. Benowitz, unpublished observations). To concentrate AF-1, we lyophilized the <1 kDa fraction of the CM and extracted it with 95% EtOH (16% original volume; 1 hr extraction with frequent vortexing; 0°C). The EtOH-soluble and insoluble fractions were lyophilized, weighed, and bioassayed. The >3 kDa fraction was concentrated 10- to 40-fold, diluted in 25 mm HEPES, pH 7.75, containing 1 mm EDTA, and repeatedly subjected to ultrafiltration to exchange the buffer and eliminate residual AF-1.
Further separation of AF-2. Anion-exchange chromatography was performed on the high-molecular weight fraction of CM using diethylaminoethyl cellulose (DE-52; Whatman, Maidstone, UK). The >3 kDa fraction of CM (5 ml of a 20-fold concentrated sample) was incubated overnight (4°C) with 0.2 ml of preequilibrated DE-52 and transferred to a 10 ml Econo-Column (Bio-Rad, Hercules, CA), and the unbound fraction was collected. Bound proteins were eluted stepwise with increasing concentrations of NaCl in 25 mm HEPES and 1 mm EDTA (1 ml/min). For each fraction, buffer was exchanged for 50 mm NaCl, and the samples were concentrated 10- to 100-fold by ultrafiltration (molecular weight cutoff of 3 kDa). Fractions were bioassayed as described above. Fractions containing neurite-promoting activity were separated by size-exclusion HPLC (Beckman Instruments) using a Biosep SEC-2000 silica column (Phenomenex) at a flow rate of 1 ml/min; the elution profile was monitored spectrophotometrically at 280 nm. Fractions of 1 ml were tested for biological activity.
Antibodies. The following polyclonal sera or monoclonal antibodies (mAb) were used for immunofluorescent staining: The IgG fraction of a polyclonal rabbit serum against recombinant goldfish GAP-43 [provided by Ute Laessing and Christian Leppert and described by Benowitz et al. (1998)] was preabsorbed against Escherichia coli protein to reduce nonspecific binding and then used at a concentration of 1:1000 for immunohistochemistry and 1:2000 for Western blot analysis. A polyclonal serum against c-Jun, which recognizes all three Jun proteins (c-Jun, Jun B, and Jun D) (Kovary and Bravo, 1991), was used at 1:8000 for immunohistochemistry and 1:10,000 for Western blot analysis (antibody 636/3; kindly provided by Thomas Herdegen), and the resulting staining is referred to as Jun-like immunoreactivity (Jun-IR). Also used were E17, a mAb against E587 antigen (E587-Ag), an L1-like cell-adhesion molecule in goldfish (Weiland et al., 1997); N518, a mAb against neurolin, the goldfish homolog of DM-GRASP (Bastmeyer et al., 1995; Leppert et al., 1999); R643 mAb against reggie-2 from goldfish (Schulte et al., 1997;Lang et al., 1998); and a polyclonal serum against goldfish reggie-2 (Lang et al., 1998). Secondary antibodies included the following: Indocarbocyanine (Cy3)-conjugated donkey anti-mouse IgG at 1:1000 (Jackson ImmunoResearch, West Grove, PA) was used to recognize all of the monoclonal antibodies used. Alexa-488-conjugated goat anti-rabbit IgG at 1:1000 (Molecular Probes, Eugene, OR) and HRP-conjugated goat anti-rabbit IgG at 1:10,000 (Jackson ImmunoResearch) were used to recognize rabbit polyclonal antibodies for immunofluorescence and Western blots, respectively.
Immunofluorescence. After 6 d in culture, gfRGCs were rinsed with L-15, fixed 5 min in MeOH (−20°C) followed by 5 min in 2% paraformaldehyde, and then rinsed with PBS. Cells were exposed to both primary antibodies overnight at 4°C in PBS containing 1% BSA and 2% normal goat serum, washed three times with PBS, and incubated for 2 hr at RT with Alexa-488-conjugated goat anti-rabbit IgG and Cy3-conjugated donkey anti-mouse IgG. Cultures were rinsed in PBS and coverslipped in Mowiol (Hoechst) containing n-propylgallate (Sigma) as an antifading agent.
Western blot analysis. gfRGCs were grown in 100 mm dishes in the presence of various trophic factors or inhibitors for 6 d. Cells were collected and centrifuged, and their proteins were separated by SDS-PAGE. Proteins were transferred to polyvinylidene difluoride membranes (Millipore, Bedford, MA), probed with the primary antibody (overnight; 4°C), washed in TBS, and incubated for 1 hr (RT) with an HRP-conjugated secondary antibody followed by an ECL reagent (Amersham Pharmacia Biotech).
Quantitation of protein expression. Cultures were viewed under 400× magnification, alternating between phase-contrast and epifluorescent illumination. Fluorescence was assessed in at least 200 consecutively encountered RGCs per well. Results are reported as the average of two to four independent experiments. To correlate protein expression with axon growth, we recorded fluorescence levels for RGCs with axons ≤2 cell diameters in length (− axons) and for RGCs with axons >2 cell diameters in length (+ axons). Fluorescence was graded as undetectable (−), light (+/−), moderate (+), or intense (++). To subdivide RGCs into antigen expressing and nonexpressing cells, we pooled the numbers of − and +/− cells into one group and the numbers of + and ++ cells into another. Counting was done without knowledge of the experimental conditions in each well.
Tα-1 tubulin expression: generation of a1TIpEGFP transgenic zebrafish. Zebrafish embryos at the one to four cell stage were injected with the −1696a1TIpEGFP expression vector that contains 1696 bp of 5′-flanking DNA of the goldfish α1 tubulin gene along with exon 1, intron 1 and the first codon of exon 2 (amino acid 2 begins exon 2) fused in-frame with the enhanced green fluorescent protein (EGFP) sequence. Expression of this construct faithfully reproduces the pattern of expression of Tα-1 tubulin during axon development and regeneration (Hieber et al., 1998). At 48 hr after injection, embryos were screened for EGFP expression by the use of epifluorescence microscopy. Positive embryos were raised to sexual maturity (∼6 months) and then bred with wild-type fish; progeny were screened for EGFP expression. Approximately 400 embryos were screened before discarding an injected fish. Mature injected fish that produced EGFP-positive progeny were saved as transgenic founders. EGFP-positive progeny were raised to sexual maturity and bred with wild-type fish to produce F2 progeny. F2 progeny were similarly screened for EGFP expression at 48 hr, and positive fish were saved. To prepare cultures, transgenic zebrafish were dark-adapted, anesthetized, and killed, and their retinas were dissected and dissociated as described above. Culture conditions were the same as those used for gfRGCs. After 5 d in culture, dissociated zebrafish RGCs (zfRGCs) were examined under phase and fluorescent microscopy. A minimum of 150 consecutive zfRGCs per well were evaluated for axon growth (>2 cell diameters) and for expression of Tα-1.EGFP. Analyses were done by an investigator blind to the experimental conditions and were performed in four replicate wells for each experiment.
RESULTS
Several growth factors stimulate axogenesis in goldfish RGCs
As described previously, the principal neurite-promoting activity in optic nerve conditioned medium (ON CM) is a small hydrophilic molecule (<1 kDa) designated AF-1 (Schwalb et al., 1995, 1996). AF-1 is soluble in 95% ethanol (Fig.1a). This property enables us to separate it from >97% of the inorganic components of the conditioned medium, which remain insoluble. AF-1 stimulates maximal growth when tested at 20–30% of its original concentration in ON CM (Fig. 1b).

Goldfish RGCs extend neurites in response to several trophic agents. a, AF-1 is a potent neurite-promoting factor that is present in the low-molecular weight fraction of ON CM. In a typical experiment, AF-1 induces 30–40% of cultured RGCs to extend axons ≥5 cell diameters in length by 6 d. The net growth in these cells, obtained by subtracting growth in negative control cells grown in defined media (∼4%), is used to normalize the data in each experiment. AF-1 can be extracted and concentrated in 95% ethanol (EtOH extract).b, AF-1 induces maximal growth at concentrations ≥20%.c, The high-molecular weight fraction from ON CM contains a neurite-promoting factor that binds to an anion-exchange column (DE-52) at pH 7.75 and elutes with 0.2–0.3 m NaCl.d, At concentrations >0.5×, partially purified AF-2 induces a maximal response equal to ∼35% of that of AF-1 (solid line). However, by the use of a lower criterion for growth (neurites > 2 cell diameters in length), AF-2 is seen to affect many more cells (dashed line).e, Recombinant rat CNTF, at concentrations ≥2 ng/ml, induces 50–70% the level of growth seen with AF-1.dia., Diameters; pct., percent.
On the basis of size-exclusion and ion-exchange chromatography, we reported previously that goldfish ON CM also contains a positively charged, 12 kDa axogenesis factor we called AF-2 and noted that some preparations additionally contained an axon-promoting factor of ∼70 kDa Mr (Schwalb et al., 1995). The present study used a protocol similar to that used previously to separate AF-2 but included EDTA in the buffers to inhibit Ca2+-dependent proteases. Under these conditions, most of the axon-promoting activity in the >3 kDa fraction behaved as a 70 kDa acidic protein. At pH 7.75, the neurite-promoting activity in the >3 kDa fraction bound to an anion-exchange column and was eluted with 0.2–0.3 m NaCl (Fig.1c); levels of a 12 kDa axon-promoting factor that failed to bind to this column were correspondingly diminished. On a size-exclusion column, the biologically active fractions from the ion-exchange column eluted at 6–7 min, similar to the elution time of bovine serum albumin (Mr = 68 kDa; data not shown). Our interpretation of these results is that the 12 kDa species described previously represents a partially degraded form of the 70 kDa protein. Even when concentrated severalfold, AF-2 stimulated considerably less outgrowth than did AF-1 (Fig. 1d). However, by the use of a less stringent criterion for growth (axons > 2 cell diameters), AF-2 induced ∼60% the level of outgrowth of AF-1 after 6 d (data not shown).
Recombinant human CNTF had failed to stimulate outgrowth from goldfish RGCs in a previous study (Schwalb et al., 1995). However, because of the marked effects of CNTF on mammalian RGCs (Meyer-Franke et al., 1995; Cui et al., 1999; Jo et al., 1999) and the known evolutionary divergence between the rat and human sequences (Lam et al., 1991), we reexamined the role of CNTF on goldfish RGCs using a recombinant rat protein (rrCNTF; Promega). At concentrations ≥2 ng/ml, rrCNTF induced 50–80% the level of growth stimulated by AF-1 (Fig. 1e). For reasons that remain unknown, although several preparations of rrCNTF repeatedly gave the type of outgrowth shown here, other batches had no activity.
Growth factor combinations
If the various trophic factors stimulate growth via a common pathway, their combined effects should be additive at low concentrations but should plateau at the level achieved with saturating doses of one factor alone. On the other hand, if they stimulate outgrowth via different pathways, their combined effects at saturating concentrations might exceed those of any one factor alone. The experiments shown in Figure 2 suggest that the various factors stimulate growth via a common pathway. At suboptimal concentrations (Fig. 2a), AF-1 (12%) stimulated ∼30% the level of growth seen in the positive control, AF-2 (0.25×) had no measurable effect, and CNTF (1 ng/ml) stimulated 18% the level of outgrowth seen in the positive control. When combined, these concentrations of AF-1 plus AF-2 or of AF-1 plus CNTF showed additive effects, inducing more growth than either factor alone (*p ≤ 0.05 for AF-1 + AF-2 compared with either AF-1 or AF-2 alone; *p < 0.05 for AF-1 + CNTF vs CNTF alone; p = 0.06 for AF-1 + CNTF vs AF-1 alone; allp values based on two-tailed t tests). CNTF + AF-2 did not show additive effects. At higher concentrations (Fig.2b), there was no additivity; addition of either AF-2 (at 1.5×) or CNTF (20 ng/ml) to a high concentration of AF-1 (27.5%) did not increase growth beyond the level achieved with AF-1 alone. Likewise, outgrowth induced by high concentrations of AF-2 plus CNTF was not significantly higher than that with CNTF alone.

Partially additive effects of AF-1, AF-2, and CNTF. a, At low concentrations, the effects of the three growth factors are additive. The three bars on theleft indicate the levels of outgrowth induced by low concentrations of AF-1 (12%), AF-2 (25%), or CNTF (1 ng/ml) alone. When combined at these concentrations, AF-1 + AF-2 had additive effects, as did AF-1 + CNTF (*p < 0.05, comparing the effect of the two factors combined with the growth induced by either one alone). b, At higher concentrations, each factor alone induced a characteristic maximum response; when any two were combined, their effects did not exceed those of the more potent factor alone. ns, Growth not significantly different from the level stimulated by the more potent factor alone.
6-TG inhibits the effects of all three growth factors
In PC12 cells, the purine analog 6-TG inhibits neurite outgrowth and N-kinase activity in parallel (Volonte et al., 1989; Volonte and Greene, 1992). In gfRGCs, 20 μm 6-TG caused outgrowth induced by AF-1, AF-2, and CNTF to decline below baseline levels (Fig.3a); survival was unaffected (Fig. 3b).

The purine analog 6-TG blocks neurite outgrowth induced by all three growth factors. a, Lightly shaded bars indicate growth induced by saturating concentrations of AF-1, AF-2, or CNTF. In the presence of 6-TG (20 μm), all growth was reduced below baseline (darkly shaded bars). b, 6-TG does not diminish cell survival (shown as the number of viable gfRGCs per 400× microscope field, averaged over 20–30 fields per well and 4 wells per condition).
Inosine acts competitively with 6-TG
Unlike polypeptide growth factors, inosine induces axon outgrowth in gfRGCs via a direct intracellular mechanism (Benowitz et al., 1998). As shown in Figure 4, inosine induces somewhat less growth than does AF-1 throughout the first 6 d in culture. By 12 d, both stimulated ∼80% of the cells to extend axons ≥5 cell diameters in length (data not shown). Despite the greater potency of AF-1, inosine is much more effective than is AF-1 in overcoming the inhibitory effects of 6-TG. Whereas high concentrations of AF-1 were unable to reverse the inhibitory effects of 10 μm 6-TG (Fig. 4b), increasing concentrations of inosine fully overcame this blockade (Fig. 4c). Thus, inosine and 6-TG act competitively.
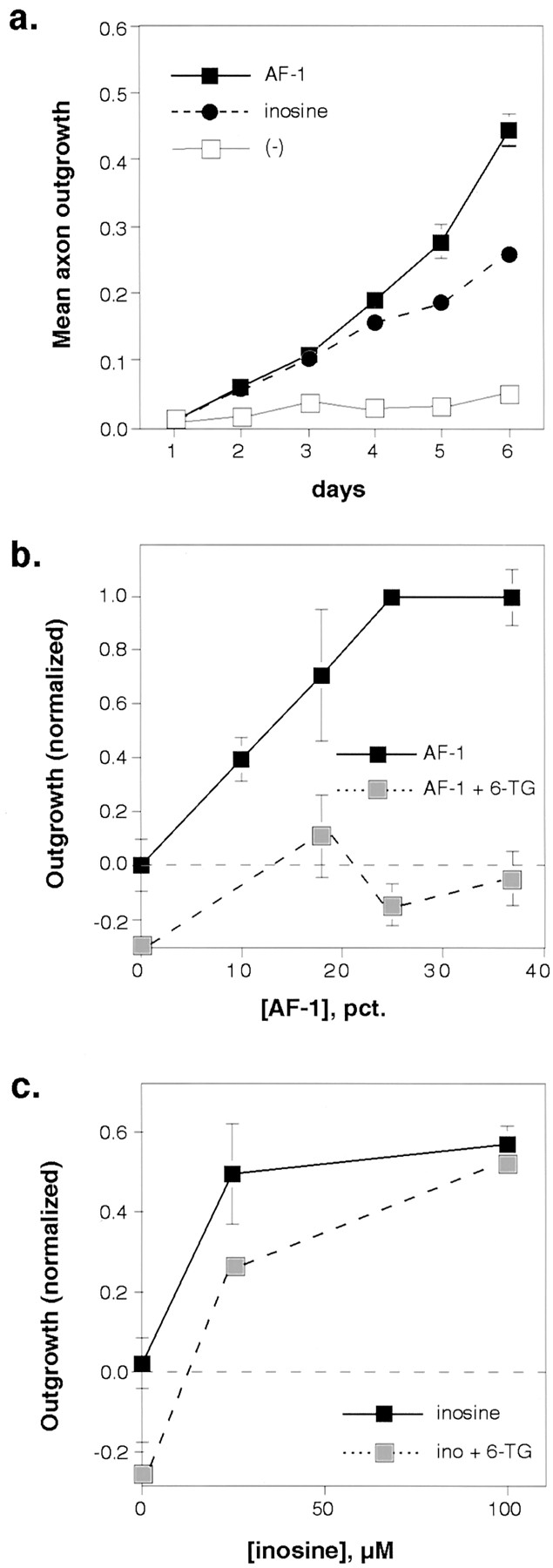
Contrasting effects of AF-1 and inosine.a, Over the first 6 d in culture, AF-1 (30% concentration; filled squares) stimulated somewhat more vigorous outgrowth than did inosine (100 μm;filled circles; ED50 = 11.4 μm). b, 6-TG and AF-1 behave noncompetitively. The inhibitory effects of 10 μm 6-TG on outgrowth (lightly shaded squares) were not overcome with increasing concentrations of AF-1. c, Inosine and 6-TG act competitively. Increasing concentrations of inosine reversed the inhibitory effects of 6-TG (lightly shaded squares).Ino, Inosine.
Purinergic control of gene expression
The expression of GAP-43, E587-Ag, and neurolin all paralleled axon outgrowth. After 6 d in culture with defined media alone, ∼15% of RGCs showed a moderate level of GAP-43 expression (Figs.5b,6a,b), and only 1% showed intense staining (data not shown). AF-1 and inosine (100 μm) increased the overall number of RGCs with appreciable GAP-43 staining (Figs. 5e,h, 6a,b) and increased the number of cells with intense staining 10-fold (data not shown). 6-TG (10 μm) suppressed GAP-43 expression below baseline (Fig. 5k), whereas inosine competitively reversed this inhibition (Fig. 5n).

Expression of growth-associated proteins is controlled via a purine-sensitive mechanism. Dissociated gfRGCs were treated with either medium alone (a–c), AF-1 (d–f), inosine (g–i), AF-1 plus 6-TG (j–l), or AF-1 plus 6-TG and inosine (m–o). a–c, A small number of RGCs maintained in medium alone expressed GAP-43 (b,arrowhead) or neurolin (c,arrow); few untreated cells expressed both;a shows the same cells under phase optics.d–f, When stimulated with AF-1, most RGCs that extended axons expressed GAP-43 (e); some (arrow) but not all (solid arrowhead) of these also expressed E587-Ag; d shows the same cells under phase optics. g–i, Inosine treatment results in the same pattern of GAP-43 expression seen with AF-1 (h); neurolin was coexpressed in many (arrows) but not all (arrowheads) of the GAP-43-expressing RGCs; open arrowheads point to cells with neither protein. g shows the same cells under phase optics. j–l, In the presence of AF-1 + 6-TG, axon outgrowth and the expression of GAP-43 (k) and E587-Ag (l) declined below baseline; open arrowheads show cells expressing neither protein; solid arrowheads show cells expressing GAP-43 but not E587-Ag;j shows the same cells under phase optics.m–o, Inosine restored GAP-43 (n) and E587-Ag (o) expression in the presence of 6-TG back to the levels seen in AF-1-treated cultures;arrows show cells coexpressing both proteins;solid arrowheads show RGCs expressing GAP-43 but not E587-Ag; open arrowheads show cells expressing neither protein; m shows the same cells under phase optics. Scale bar, 40 μm.
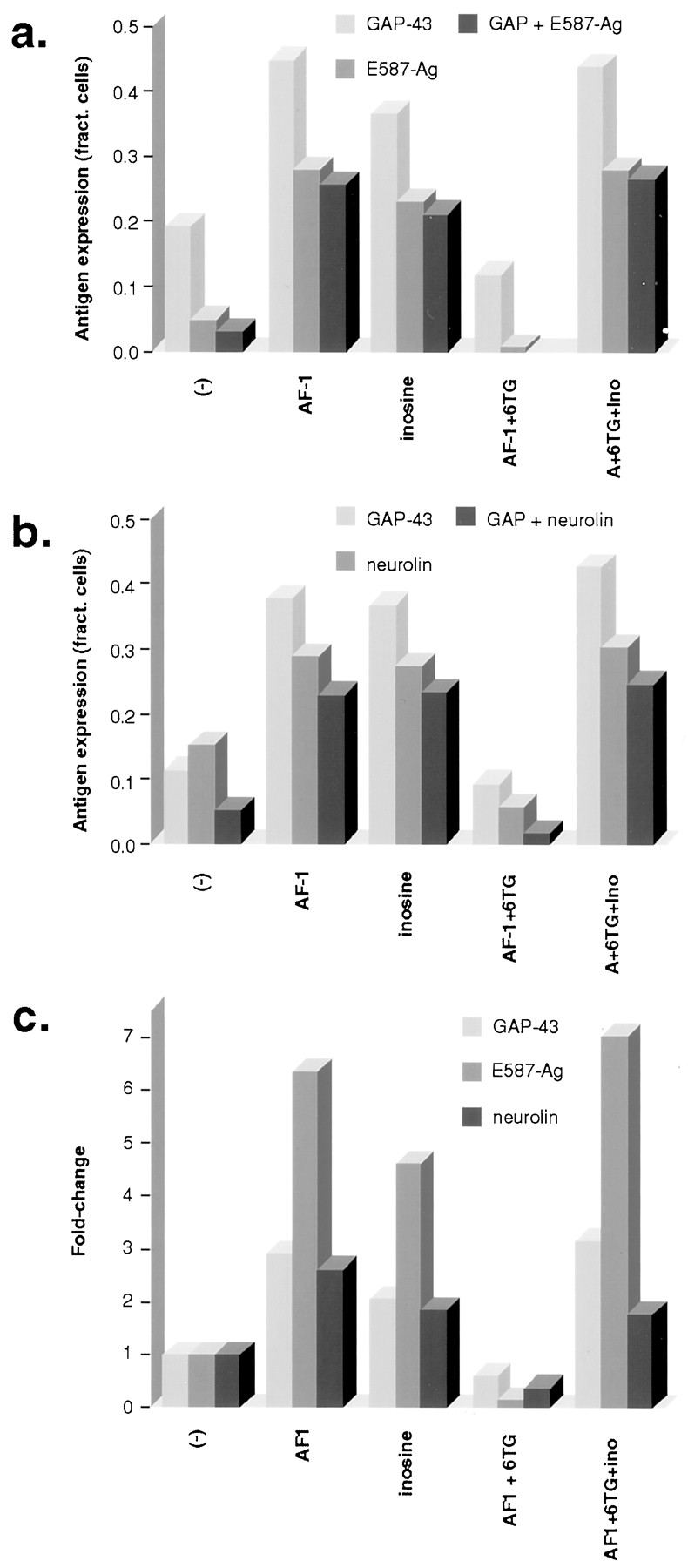
Coexpression of growth-associated proteins: quantitation. a, AF-1 and inosine increased the number of gfRGCs with detectable levels of GAP-43 and E587-Ag. 6-TG reduced expression below baseline (AF-1 + 6TG), whereas inosine restored expression of both (A + 6TG + Ino). Across all conditions, GAP-43 was detected in more cells than was E587-Ag, whereas most cells that expressed E587-Ag also expressed GAP-43.b, Neurolin expression followed a pattern similar to that of GAP-43 and E587-Ag, i.e., induction by AF-1 and inosine, suppression by 6-TG, and restoration with inosine. Under most conditions, neurolin was detected in approximately the same percentage of cells as was GAP-43. c, The fold-increase in protein expression is shown. Of the three proteins studied, E587-Ag expression showed the highest degree of regulation under the different experimental conditions. Results represent the averages of two independent experiments; in each of these, 150 consecutive RGCs were visualized per well, and the results were averaged across four wells. Counting was performed by an investigator blind to the experimental conditions.
On Western blots (see Fig. 8), the level of GAP-43 induction by AF-1 or inosine appears to be considerably larger than that seen by cell counting. One likely reason for this is the nonlinearity of the ECL signal. Another is that the cell counts in the negative controls almost exclusively reflect cells with moderate levels of staining and do not indicate the marked increase in cellular staining that occurs in response to AF-1 or inosine. On similar Western blots, CNTF and AF-2 were also seen to induce GAP-43 expression, although to a lesser extent than did AF-1 (data not shown).
E587-Ag, an L1-like cell adhesion molecule in goldfish, was detected at moderate levels in <5% of unstimulated RGCs and was upregulated by AF-1 (Fig. 5f) and by inosine (Fig. 6a). 6-TG suppressed E587-Ag expression below baseline levels (Fig.5l), whereas inosine restored its expression (Fig.6a). Across all experimental conditions, almost all cells that expressed E587-Ag also expressed GAP-43; however, there were some cells in which only GAP-43 was detected (Figs. 5f,6a). Neurolin, the goldfish homolog of DM-GRASP, was detected at moderate levels in ∼13% of unstimulated RGCs (Figs.5c, 6b). AF-1 and inosine increased the overall number of RGCs showing neurolin staining (Figs. 5i,6b) and increased the number with intense staining from essentially none to 10% (data not shown). As with the other antigens, 6-TG reduced neurolin expression below baseline levels even in the presence of AF-1, whereas inosine competitively restored its expression (Fig. 6b). Most cells that expressed GAP-43 also expressed neurolin and vice versa (Figs. 5b,c,h,i, 6b).
Of the three proteins studied here, E587-Ag expression showed the strongest degree of regulation. Whereas AF-1 and inosine increased the number of dissociated RGCs expressing moderate-to-strong levels of GAP-43 or neurolin twofold to threefold, they caused E587-Ag expression to increase fivefold to sevenfold relative to baseline. Conversely, 6-TG caused E587-Ag expression to decline somewhat more than either GAP-43 or neurolin expression (Fig. 6c).
Molecular changes not controlled via purine-sensitive mechanisms
Jun family transcription factors are undetectable in intact gfRGCs and are induced after axotomy (Herdegen et al., 1993). Unlike the proteins described above, Jun-IR remained high in dissociated gfRGCs irrespective of whether cells grew axons; Jun-IR levels were unaffected by AF-1 or purinergic manipulations (Fig.7e,j,o). Reggie-2, another protein expressed by gfRGCs after axotomy, was likewise expressed in cell bodies irrespective of whether they were stimulated to extend axons (Fig. 7c,h,m).

Jun and reggie-2 expression are unaffected by AF-1 or 6-TG, although reggie-2 localization is altered. Dissociated gfRGCs were treated with either control medium alone (a–e), AF-1 (f–j), or 6-TG (k–o). All RGC somata showed reggie-2 (arrowheads; c, h, m) and Jun-IR (e, j, o), although not necessarily GAP-43 expression (arrowheads; b, g, l). a, d,f, i, k,and n show the same cells under phase optics. In the presence of 6-TG, RGC somata were always reggie-2 positive (m), even when their neurites contained GAP-43 but no reggie-2 (arrows; l, m). Scale bar, 30 μm.
The absence of regulation for jun proteins and reggie is confirmed on Western blots. In conformity with the immunohistochemical results, the expression of jun proteins and reggie is neither upregulated by AF-1 or inosine nor diminished by 6-TG (Fig. 8). Thus, expression of the latter proteins is related to axotomy per se and not to whether cells extend axons.

Western blot analysis of protein expression. In agreement with the cell counting, Jun family transcription factors (Mr = 48 and 45 kDa) were elevated in all dissociated gfRGCs and were not affected by AF-1 or by purinergic treatments. A similar lack of regulation was seen for reggie. In contrast, GAP-43 levels were strongly regulated by AF-1 and by purinergic manipulations. For each antigen, all lanesare from the same blot and exposure time but were repositioned to maintain the same sequence as in the other figures. A, AF-1; i, inosine; 6, 6-TG. Molecular weight markers are indicated on the left.
Relationship of gene expression to axon growth
Axon growth and GAP-43 expression were highly correlated under all experimental conditions. Even under conditions in which very few cells had axons (e.g., negative controls or in the presence of 6-TG), GAP-43 could be detected in ∼50% of the cells that had axons (Fig.9a). AF-1 and inosine greatly increased the number of cells extending axons, yet the fraction of cells with axons that expressed GAP-43 increased only slightly (to 60–70%). Across all conditions, cells with axons >2 cell diameters in length were approximately three times more likely to show GAP-43 staining than were cells without axons. Like GAP-43, E587-Ag was found in many more cells with axons than in those without axons (Fig.9b). However, the fraction of axon-bearing cells that expressed E587-Ag changed markedly under different experimental conditions. The few cells that extended axons in control cultures or in cultures treated with 6-TG showed little E587-Ag expression. AF-1 and inosine increased the fraction of neurons expressing E587-Ag, and this change was seen in cells both with and without axons (Fig.9b). Neurolin followed a pattern of expression that was intermediate between those of GAP-43 and E587-Ag (Fig. 9c). Finally, although the expression of reggie was not regulated by growth factors or 6-TG, its disposition in the axon was altered under different experimental conditions; whereas only ∼12% of the axons that formed in the absence of growth factors had little reggie-2 along their length, ∼35% of those that grew in the presence of AF-1 or inosine showed reggie-2 immunostaining (Fig. 9d).

Relationship of protein expression to axon growth.a, Across all conditions, gfRGCs with axons tended to express GAP-43 (darkly shaded bars), whereas the vast majority of cells without axons did not (lightly shaded bars). Data represent means from two to four independent experiments; error bars represent SEM or range, depending on the number of independent experiments. b, Unlike GAP-43, the association of E587-Ag with axon-bearing cells was enhanced by growth factors and diminished by 6-TG. E587-Ag was generally detected in fewer neurons than was GAP-43, but when expressed, it tended to be associated more with axon-bearing RGCs than with cells without axons.c, Neurolin expression followed a pattern intermediate between that of GAP-43 and E587-Ag. d, Reggie-2 was expressed in the cell bodies of all RGCs in culture (lightly shaded bars). However, its localization in axons varied under different stimulation conditions (darkly shaded bars).fract., Fraction of.
Tα-1 tubulin expression
Retinal ganglion cells were cultured from transgenic zebrafish harboring an EGFP gene fused to exon 2 of Tα-1 tubulin and driven by the Tα-1 promoter. As in gfRGCs, axon outgrowth in transgenic zebrafish RGCs (tg zfRGCs) was regulated by AF-1 and by purinergic manipulations (Fig.10a,c). Tα-1.EGFP expression paralleled outgrowth (Fig. 10b,d). In control cultures, EGFP could be detected in ∼8% of tg zfRGCs after 5 d. AF-1 caused the number of zfRGCs with detectable levels of the reporter to increase nearly threefold (Fig. 10b,d), which paralleled the threefold to fourfold increase in the fraction of cells with axons ≥2 cell diameters in length (from ∼10 to 40%). Inosine induced a twofold increase in the number of cells expressing the reporter. The effects of AF-1 were inhibited by 6-TG, whereas inosine competitively restored transgene expression (Fig. 10d). Like GAP-43, Tα-1.EGFP expression had a striking relationship to axon growth. That is, across all conditions, EGFP was detected in only ∼5% of cells without axons but was seen in 25–40% of cells bearing axons (Fig.10e). This percentage was lowest in cells treated with 6-TG, presumably because the population of RGCs with axons in this condition had, on the average, lower levels of outgrowth than did the others. Most cells that expressed Tα-1.EGFP also expressed GAP-43 (Fig.10f,g). Thus, Tα-1 expression parallels axon growth and is regulated by a purine-sensitive mechanism similar to that controlling GAP-43, E587-Ag, and neurolin expression.

Regulation of Tα-1 tubulin expression. tg zfRGCs harbored a reporter construct expressing a Tα-1.EGFP fusion protein driven by the Tα-1 promoter. a, b, tg zfRGCs extended axons (a) and exhibited reporter activity (b) in response to AF-1. Filled arrowheads show cells with axon growth and reporter expression;open arrowheads show a cell with little outgrowth and low levels of transgene expression; arrows point to axons. c, Axon outgrowth in tg zfRGCs followed the same pattern observed in gfRGCs, i.e., induction by AF-1 and inosine, inhibition by 6-TG, and restoration by inosine (***p < 0.001, comparing growth with that in negative controls). d, Expression of Tα-1 is regulated via a purine-sensitive mechanism and parallels axon outgrowth (***p < 0.001, **p < 0.01, comparing reporter expression with negative controls).e, Relationship of Tα-1.EGFP expression to axon growth is shown. Across all conditions, 30–40% of RGCs with axons expressed appreciable levels of the reporter, whereas cells without axons generally did not. f, g, Coexpression of GAP-43 (red staining; f) and Tα-1.EGFP (g) is shown. The white asteriskdenotes a growth cone.
DISCUSSION
Our results show that three different factors, AF-1, AF-2, and CNTF, stimulate gfRGCs to regenerate their axons in culture. Axon regeneration was accompanied by increased expression of several well characterized growth-associated proteins, including GAP-43, Tα-1, and two Ig superfamily cell adhesion molecules. The purine analog 6-TG suppressed all of these changes, whereas the purine nucleoside inosine acted in an opposite manner, stimulating expression of these proteins and competitively reversing the inhibitory effects of 6-TG. Purinergic manipulations did not affect cell survival or molecular changes associated with axotomy per se.
Different growth factors stimulate axogenesis via a common pathway
AF-1 and AF-2 are secreted by optic nerve glia and may therefore contribute to gfRGC axon regeneration in vivo. AF-1 is a highly potent, small, hydrophilic molecule. AF-2, which induces relatively lower levels of outgrowth in culture, was described previously as a basic, 12 kDa polypeptide (Schwalb et al., 1995). However, when prepared in the presence of a Ca2+ chelator, AF-2 behaved like an acidic, 70 kDa protein, presumably a precursor of the 12 kDa species. In addition to these, CNTF stimulated axon outgrowth in gfRGCs. This is consistent with the known axogenic effects of CNTF on mammalian RGCs (Mey and Thanos, 1993; Cui et al., 1999; Jo et al., 1999). CNTF mRNA and protein are abundant in mammalian optic nerve (Stockli et al., 1991). This raises the question of whether AF-2 is a goldfish homolog of CNTF, yet this seems unlikely because (1) AF-2 and CNTF have greatly different sizes (Stockli et al., 1989) and (2) CNTF induced stronger outgrowth than did AF-2. In addition to AF-1 and AF-2, goldfish optic nerve has also been reported to secrete factors that stimulate axon regeneration in mammalian neurons (Schwartz et al., 1985; Caday et al., 1989).
The signal transduction pathways underlying axon outgrowth in gfRGCs are not well characterized. In culture, the effects of AF-1 appear to be at least partly independent of MAP kinase-1 or kinase-2 (ERK-1, ERK-2) and phosphatidylinositol-3 (PI-3) kinase. Blocking either of these signaling pathways leaves outgrowth unimpaired, whereas blocking both only partially inhibits outgrowth (Benowitz et al., 1998). CNTF acts via a multimeric receptor complex containing CNTF receptor-α, leukocyte inhibitory factor receptor-β, and gp130 to activate a Jak-STAT transduction pathway (Stahl et al., 1994; Symes et al., 1997). Nothing is known about AF-2 signaling. Despite the different levels of growth induced by the three factors, and the possibility that each may initially act via different receptors and signaling pathways, their effects on axon outgrowth appear to converge on a 6-TG-sensitive mechanism. Mixing experiments provide additional evidence that the three factors stimulate growth via a final common pathway. Whereas low concentrations of AF-1, AF-2, and CNTF induced outgrowth in an additive manner, their effects saturated at the level of growth induced by the more potent factor alone.
In PC12 cells, NGF binding to TrkA activates several well characterized signal transduction cascades, e.g., those involving MAP kinases and PI-3 kinase (Traverse et al., 1992; Yao and Cooper, 1995; Creedon et al., 1996; Klesse et al., 1999). Via mechanisms that are not yet known, NGF also rapidly activates N-kinase, a 47–50 kDa serine-threonine kinase (Blenis and Erikson, 1986; Rowland-Gagne and Greene, 1990). N-kinase is implicated in neuritic differentiation because of the fact that 6-TG selectively inhibits outgrowth and N-kinase activity in parallel, while not affecting cell survival or the activity of other signaling molecules (Greene et al., 1990; Batistatou et al., 1992). On the basis of these findings, it is likely that the inhibitory effects of 6-TG on gene expression and axon outgrowth in gfRGCs likewise involve N-kinase inhibition.
The purine nucleoside inosine induces outgrowth via a direct intracellular mechanism of action (Benowitz et al., 1998). As shown here, inosine stimulated ∼60% the level of outgrowth stimulated by AF-1 and induced a similar pattern of molecular changes. Because inosine acts as a competitor to 6-TG, we have hypothesized that inosine stimulates axon outgrowth by acting as an agonist at the same site that is blocked by 6-TG, i.e., N-kinase.
Regulation of gene expression
One of the most striking correlates of axon regeneration in vivo is the upregulation of GAP-43 expression (Benowitz et al., 1981; Skene and Willard, 1981; Benowitz and Lewis, 1983). GAP-43 is a phosphoprotein that is linked to the submembrane cytoskeleton (Meiri and Gordon-Weeks, 1990) and that plays a role in the structural remodeling of axonal endings (Benowitz and Routtenberg, 1997). In the mouse retinofugal pathway, GAP-43 is crucial for axon pathfinding (Strittmatter et al., 1995; Kruger et al., 1998; Sretavan and Kruger, 1998), perhaps via an involvement in L1-mediated guidance mechanisms (Meiri et al., 1998). Across all experimental conditions, including even control cultures or neurons treated with 6-TG, most neurons that had an axon expressed GAP-43. AF-1, AF-2, CNTF, and inosine induced GAP-43 expression and axon growth in parallel. Thus, irrespective of overall levels of outgrowth, GAP-43 is intimately associated with growing axons, in which it is concentrated along the axonal membrane, particularly in varicosities from which filopodia emerge.
E587-Ag and neurolin are members of the Ig superfamily of cell adhesion/recognition molecules. Both are upregulated in gfRGCs during axon regeneration in vivo (Vielmetter et al., 1991; Paschke et al., 1992; Laessing and Stuermer, 1996). E587-Ag is structurally similar to L1 (Giordano et al., 1997) and contributes to the efficacy of axon elongation and fasciculation within the optic nerve (Bastmeyer et al., 1995; Walsh et al., 1997). Of the proteins studied here, E587-Ag expression showed the strongest induction by growth factors and inhibition by 6-TG. E587-Ag was generally found in RGCs extending axons when cells were stimulated by AF-1 or inosine. However, unlike GAP-43, E587-Ag was not found in the few RGCs that extended axons in control cultures or in the presence of 6-TG. Neurolin is a homolog of mammalian DM-GRASP. Like E587-Ag, neurolin appears to be involved in RGC axon guidance rather than in growth per se (Ott et al., 1998; Leppert et al., 1999). Its regulation by purines and the relationship of its expression to axon growth were intermediate between the patterns seen for GAP-43 and E587-Ag.
Reggie proteins are localized in microdomains on the cytoplasmic face of the plasma membrane and are induced in RGCs after optic nerve lesions in vivo (Schulte et al., 1997; Lang et al., 1998). The association of reggie-1 and reggie-2 microdomains with activated glycosylphosphatidylinositol (GPI)-linked CAMs (Lang et al., 1998) and with the tyrosine kinase fyn (Stuermer et al., 2000) suggests that reggie proteins participate in axon growth in a context-dependent manner, which may involve GPI-linked CAMs. Because reggie-1 and reggie-2 are invariably coexpressed, we have restricted our studies to reggie-2. Our results indicate that whereas the basal induction of reggie-2 in axotomized gfRGC somata was unaffected by AF-1 or 6-TG, thedistribution of reggie-2 in axons changed under different experimental conditions. Thus, whereas reggie-2 was detected in ∼12% of axons that grew in negative control cultures (or in cultures treated with 6-TG), it appeared in approximately three times as many axons when cells were stimulated by AF-1 or inosine.
Tα-1 is a neuron-specific isoform of tubulin that is induced when axons are growing (Hieber et al., 1998). Via the use of a reporter construct, we showed here that Tα-1 expression is regulated via the same purine-sensitive pathway that controls other growth-associated proteins in gfRGCs. These studies open the way for future genetic analyses using transgenic zebrafish to elucidate the signal transduction pathways that control axon outgrowth.
Axotomy induces expression of Jun family transcription factors irrespective of whether neurons regenerate their axons (Herdegen et al., 1993; Hull and Bahr, 1994). In conformity with this, our cellular immunostaining and Western blots showed that the expression of Jun proteins was high in control RGCs in culture and was neither upregulated by AF-1 nor downregulated by 6-TG. The proteins visualized here are tentatively identified as c-jun, jun-B, and jun-D based on the molecular weights of the mammalian proteins (Kovary and Bravo, 1991). The absence of Jun regulation by 6-TG is consistent with the observation that 6-TG does not diminish NGF induction of c-jun, zif-268, or c-fos in PC12 cells (Batistatou et al., 1992).
Summary and conclusions
Axon regeneration in gfRGCs is marked by a constellation of molecular changes and can be stimulated by at least three growth factors. All of these changes appear to be mediated via a purine-sensitive pathway. Previous studies in PC12 cells show that 6-TG selectively blocks the activity of N-kinase and neurite differentiation in parallel, and we therefore hypothesize that N-kinase is a critical component of the transduction pathway controlling axon outgrowth and the underlying pattern of molecular changes in gfRGCs. In agreement with previous studies, we find that this pathway does not control cell survival or the expression of several immediate-early genes. Our laboratory has now isolated the purine-sensitive kinase, and as predicted, it is distinct from other signal transduction kinases implicated in growth factor signaling in neurons (L. I. Benowitz, Y. Li, J. O'Toole, G.-f. Hu, and N. Irwin, unpublished observations). The wider significance of the purine-sensitive pathway for controlling axon growth has been demonstrated in the rat spinal cord, where inosine induces massive axon growth after injury to the corticospinal tract (Benowitz et al., 1999).
Footnotes
This work was supported by the National Institutes of Health Grant EY 05690 to L.B., Boston Life Sciences, Inc., the Boston Neurosurgical Foundation, Bundesministerium für Bildung, Wissenschaft und Forschung to C.S., Fonds der Chemischen Industrie to C.S., and the Shelley and Steven Einhorn Research Fund of The Glaucoma Foundation to D.G. We wish to acknowledge the preliminary studies of Amir Taghinia, William Schloss, and Eli Diamond on gene expression and John Mush for managing the zebrafish facility in D.G.'s laboratory.
B.P. and R.T. contributed equally to this work.
Correspondence should be addressed to Dr. Larry I. Benowitz, Laboratories for Neuroscience Research in Neurosurgery, Children's Hospital, Boston, MA 02115. E-mail: benowitz{at}a1.tch.harvard.edu.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}