

Abstract
A disconnection procedure was used to test whether the hippocampus and anterior thalamic nuclei form functional components of the same spatial memory system. Unilateral excitotoxic lesions were placed in the anterior thalamic (AT) nuclei and hippocampus (HPC) in either the same (AT-HPC Ipsi group) or contralateral (AT-HPC Contra group) hemispheres of rats. The behavioral effects of these combined lesions were compared in several spatial memory tasks sensitive to bilateral hippocampal lesions. In all of the tasks tested, T-maze alternation, radial arm maze, and Morris water maze, those animals with lesions placed in the contralateral hemispheres were more impaired than those animals with lesions in the same hemisphere. These results provide direct support for the notion that the performance of tasks that require spatial memory rely on the operation of the anterior thalamus and hippocampus within an integrated neural network.
Pathology in the hippocampus (HPC) is sufficient to induce many of the memory deficits associated with temporal lobe amnesia (Scoville and Milner, 1957;Squire and Zola Morgan, 1991). From this, it is known that the integrity of the hippocampus is vital for normal episodic memory. There is not the same consensus over the key sites responsible for the memory deficits in diencephalic amnesia, but recent evidence strongly implicates the anterior thalamic nuclei (Aggleton and Brown, 1999;Harding et al., 2000; Van der Werf et al., 2000). Because the hippocampus and the anterior thalamic (AT) nuclei are anatomically interlinked, it is possible that they form part of the same mnemonic system, a notion first proposed by Delay and Brion (1969). Although clinical data have not provided a definitive test of this proposal, it is possible to examine this relationship in animals using disconnection procedures. The present study provides a direct test of the importance of these interactions for one class of memory, spatial memory.
Lesion studies have confirmed that the rat hippocampal formation is vital for the processing of spatial memory information in a variety of learning tasks. These include nonmatching-to-place in the T-maze, foraging in the radial arm maze, navigation in the Morris water maze, and delayed nonmatching-to-position in an automated test chamber (Morris et al., 1982; Aggleton et al., 1986, 1990, 1992;Jarrard, 1993). The same tasks are also disrupted by lesions of the anterior thalamic nuclei, suggesting that this region also plays a vital role in spatial memory function (Aggleton and Sahgal, 1993; Byatt and Dalrymple-Alford, 1996; Warburton et al., 1997; Warburton and Aggleton, 1999). These functional similarities are supported by the anatomical links between the two regions because the hippocampus projects to the anterior thalamic nuclei both directly and indirectly via the mammillary bodies (Swanson et al., 1987). The anterior thalamic nuclei, in turn, project directly to the hippocampal formation (Shibata, 1993) via the cingulum bundle and indirectly via the retrosplenial cortices (Mufson and Pandya, 1984; Van Groen and Wyss, 1995). Thus, lesions in two reciprocally connected regions result in superficially similar effects.
To determine whether the hippocampus and anterior thalamic nuclei function in an interdependent manner, animals were first prepared with unilateral lesions of both the hippocampus and the anterior thalamic nuclei. These animals were then tested on a battery of behavioral tasks that are highly sensitive to bilateral hippocampal lesions. In one group, the unilateral lesions were in contralateral hemispheres and in the other they were in the same hemisphere. If the two regions are functionally interdependent, then those animals with crossed lesions should be considerably more impaired. It was predicted that the presence of anterior thalamic damage in the same hemisphere would have little or no additional effect if the two regions are working in concert. A third group of animals with just unilateral hippocampal lesions were also included.
MATERIALS AND METHODS
Subjects
The subjects were male rats of the pigmented DA strain (Bantin and Kingman, Hull, UK). Throughout the period of the experiment, the animals were housed in pairs under diurnal conditions (14/10 hr light/dark cycle). At the start of testing, the animals were aged 4 months and weighed between 215 and 230 gm. All experiments were conducted in accordance with the United Kingdom Animals (Scientific Procedures) Act of 1986.
Surgery
Before surgery, all animals were deeply anesthetized by intraperitoneal injection (60 mg/kg) of pentobarbitone sodium (Sagatal) and then placed in a stereotaxic head holder (David Kopf Instruments, Tujunga, CA) with the nose bar at +5.0. The scalp was then cut and retracted to expose the skull. Craniotomies were then made directly above the target regions, and the dura were cut to expose the cortex.
Rats in the disconnection groups received unilateral lesions of the AT nucleus combined with a lesion of the hippocampus. For approximately one-half of the animals, the contralateral group, the AT and HPC lesions were placed in opposite hemispheres (AT-HPC Contra). For most of the remaining animals, the ipsilateral group, unilateral AT and HPC lesions were made in the same hemisphere (AT-HPC Ipsi). Because of the close proximity of the anterior thalamus and the dorsal hippocampus, the AT-HPC Ispi group contained a set of animals (n = 4) with a unilateral hippocampal lesion combined with a small amount of excitotoxic damage in the contralateral dorsal hippocampus, but no anterior thalamic damage (HPC+). This group was used to test for any additional contralateral hippocampal damage that may have occurred in the AT-HPC Contra group, after the lesion of the anterior thalamic nuclei. This group also provided comparisons between the effects of a unilateral hippocampal lesion and a unilateral hippocampal with ipsilateral anterior thalamic nuclei lesion.
Excitotoxic lesions of the anterodorsal (AD), anteroventral (AV), and anteromedial (AM) thalamic nuclei were made by injecting 0.09m NMDA (Sigma, Poole, UK) dissolved in phosphate buffer, pH 7.2, through a 1 μl Hamilton syringe into three sites in the hemisphere. Each injection was made gradually over a 5 min period, and the needle was left in situ for an additional 5 min before being withdrawn. The anteroposterior (AP) and lateral (LAT) stereotaxic coordinates relative to ear-bar zero, with the incisor bar set at +5.0 to the horizontal plane, are shown in Table1. The dorsoventral (DV) coordinates were calculated relative to the top of the cortex.
Lesion coordinates for the anterior thalamic nuclei
Lesions of the hippocampus were made using 0.06 m NMDA dissolved in phosphate buffer, pH 7.2, injected into 12 sites into the hippocampus using a procedure similar to that described by Jarrard (1989). Each injection was made through a 1 μl Hamilton syringe gradually over a 3 min period, and the needle was left in situ for an additional 3 min before being withdrawn. The AP and LAT stereotaxic coordinates were calculated relative to bregma, and the DV coordinates were calculated relative to the top of the cortex. The incisor bar was set at +5.0 to the horizontal plane. The coordinates used and the amount of neurotoxin injected are shown in Table2.
Lesion coordinates for the hippocampus
In the unilateral HPC lesion control animals (HPC+), the contralateral hippocampal damage was made by injecting 0.01 μl of NMDA into the hemisphere opposite to the unilateral hippocampal lesion at the following coordinates: AP, −2.0; LAT, ±1.6; DV, −3.1. Sham control lesions (SHAM group; n = 12) were made using the same preoperative procedure as described above, but in these cases, after the craniotomy, the injection needle was lowered to the level of the anterior thalamic nuclei bilaterally and then removed.
At the completion of surgery, the skin was sutured and an antibiotic powder (Acramide; Dales Pharmaceuticals, Skipton, UK) was applied. All animals then received 5 ml of glucose saline (subcutaneously) containing etamiphylline (Millophyline; 35 mg/kg, s.c.; Arnold's, Romford, UK), a cardiac stimulator. Postoperative care also included systemic analgesia (Temgesic; Reckett and Colman, Hull, UK).
Apparatus
Spatial forced alternation in the T-maze. All testing for the forced alternation task was performed in a modifiable T-maze. The floors of the maze were 10 cm wide and made of wood, and the walls were 17 cm high and made of clear Perspex. The stem was 70 cm long with a guillotine door located 25 cm from the end of the stem, so a start area was created. The crosspiece was 140 cm long, and at each end there was a food well 2 cm in diameter and 0.75 cm deep. The entire maze was supported by two stands 94 cm high. Illumination was provided by a fluorescent light suspended 164 cm above the apparatus.
Morris water maze. The water maze used in this experiment was a 2 m diameter white fiberglass pool, 60 cm high and mounted 58 cm above the floor. The pool was situated in a room that contained posters on the wall. Other distal cues included a curtain used to conceal the experimenter. Illumination was provided by four floor-mounted spotlights (500 W) placed in each corner of the room. The water in the maze was made opaque by the addition of 2 l of milk. An escape platform was placed 2 cm beneath the water surface and kept in a constant position in the pool during the acquisition trials. The temperature of the water was 25°C at the beginning of each testing period. The swim paths of the rats were tracked using a video camera suspended directly above the pool, and all sessions were recorded on videotape. Data were collected and analyzed on-line using an HVS image analyzer (HVS Image, Hampton, UK) connected to an Archimedes RISC computer (Acorn) using Watermaze software (Edinburgh University, Edinburgh, UK).
Radial arm maze. The radial maze was constructed from a wooden center piece measuring 34 cm in diameter, to which were attached eight identical arms (90 cm long and 10 cm wide). The walls of the center piece and arms were made of Perspex. The height of the walls was 15 cm, and the height of the wall surrounding the center piece was 29 cm. Eight guillotine doors, also made out of Perspex, separated the arms from the center piece and were opened using a pulley system attached to a circular piece of wood above the center piece to which the doors were attached. The floor of the apparatus was painted white. The room was lit using fluorescent overhead lights and contained several prominent cues, such as posters and the door. The doors were operated by the experimenter, who sat in the same location on every occasion. At the end of each arm was a hidden food well 2 cm in diameter and 0.5 cm deep.
Behavioral procedures
Spatial forced alternation in the T-maze. Two weeks after surgery, all animals were food deprived to 85% of their free-feeding body weight by restricting their daily food intake to ∼15 gm of laboratory diet (Harlan Teklad, Bicester, UK). Each animal was then given several days of pretraining in the T-maze apparatus to ensure that the rat would run reliably down the stem and arms of the maze to find food pellets in the food wells. After this, the experiment proper began.
Each trial consisted of two stages. At the start of the trial, three food pellets (45 mg; Campden Instruments, Loughborough, UK) were placed in each food well, and a metal barrier was placed at the neck of the T-maze so that one arm was closed off. As a consequence, the animal was forced to enter a preselected arm on each “sample run” and then allowed to eat the food there. The animal was then picked up and confined in the start box for a delay of 10 sec, during which the metal barrier was removed. The door to the start box was then opened, and the animal was allowed a free choice between the two arms of the T-maze. On this “choice run,” the criteria for selecting an arm consisted of the rat placing a back foot in one of the arms. No retracing was permitted. If the rat had alternated, i.e., had entered the arm not visited previously on the sample run, it was allowed to eat the food reward before being returned to its cage. If the other arm was chosen, i.e., the same arm as visited on the sample run, the rat was confined to that arm for ∼10 sec and then returned to its cage. Rats were tested in groups of three or four with each rat having one trial in turn, so that the intertrial interval was ∼4 min. The animals received six trials per day for a total of 15 sessions.
Morris water maze. After completion of T-maze alternation testing, animals were returned to a free-feeding regimen, and 5 d later water maze acquisition training began. Eight start positions (north, south, east, west, northeast, northwest, southeast, and southwest) were allocated relative to the experimenter. Each rat was placed in the pool facing the wall at one of the eight start locations, a different start location being used for every trial within a session. The rat was required to swim to an underwater platform that was positioned in the same quadrant throughout the acquisition sessions, although different rats were trained to swim to different quadrants. Both start positions and platform position were counterbalanced between the groups.
Animals were initially tested with four trials per day on each of 10 consecutive days. Each acquisition trial was terminated either when the animal located the hidden escape platform or after 120 sec had elapsed. If the rat located the hidden platform, it was allowed to remain on the platform for 30 sec. After an intertrial interval of 2 min, the rat was then placed in the pool at the second start location and so on for four trials. If the rat failed to find the platform after 120 sec, it was placed on the platform and allowed to remain on it for 30 sec.
After these 10 sessions, animals received a spatial probe trial on day 11. All rats were first given one normal acquisition trial in which the platform was located in the training quadrant as usual. Then the platform was removed from the maze, and the swim path and distance swum in each of the four quadrants was recorded over 60 sec. For this probe trial, each rat was placed at a start position directly opposite to where the platform had been located. For example, if the platform had always been located in the southwest quadrant, the rat was now placed in the water, facing the wall, at the northeast position.
Radial arm maze. Animals were again placed on a restricted diet until they had achieved 85% of their free-feeding weight, and 15 d after the water maze probe trial, the radial arm maze training began. All animals received two habituation sessions (5 min) in the radial arm maze, during which time each rat was placed in the center of the maze and allowed to move freely around the maze until either it had eaten all available reward pellets or 5 min had elapsed.
On the subsequent test trials, all eight arms were baited with two food pellets (45 mg; Campden Instruments). At the beginning of a trial, the rat was placed in the center of the maze with the doors to the arms closed, all doors were then opened, and the rat was allowed to choose an arm. When the rat had consumed the food pellets, it returned to the center of the maze, and the arm doors were closed, confining the rat to the center of the maze for a 10 sec interval. The doors were then reopened, and the rat was allowed to visit another arm. If an incorrect choice was made, i.e., an arm chosen that had been visited previously, the animal was confined to that arm for a 5 sec interval without reward before being allowed to return to the center. The test session ended once the rat had visited all eight arms or a maximum of 10 min had elapsed. The rats were tested for 18 d.
RESULTS
Histology
From the histological analysis, the final group numbers were as follows: SHAM, n = 12; AT-HPC Contra, n= 8; AT-HPC Ipsi, n = 5; HPC+, n = 4.
AT-HPC Contra
Animals in which there was evidence of bilateral thalamic damage or in which there was insufficient damage in either the anterior thalamic nuclei or the hippocampus were excluded before the data analysis. The cases with the largest and smallest combined lesions are shown in Figure 1. In the resulting eight cases, the anterior thalamic lesions were almost complete as the AD and AM nuclei had primarily disappeared; however, there was some sparing of the AV nuclei in two cases. In two cases, there was additional damage in the rostral medial dorsal thalamus, and in one case, there was additional damage in the lateral dorsal thalamus. In six cases, there was a very restricted zone of neurotoxic damage to granule cells in the medial (outer) blade of the dentate gyrus (Swanson et al., 1987) immediately above the anterior thalamic nuclei (Fig. 1). In the remaining two cases, there was no additional damage to the hippocampus in the hemisphere containing the anterior thalamic lesion.
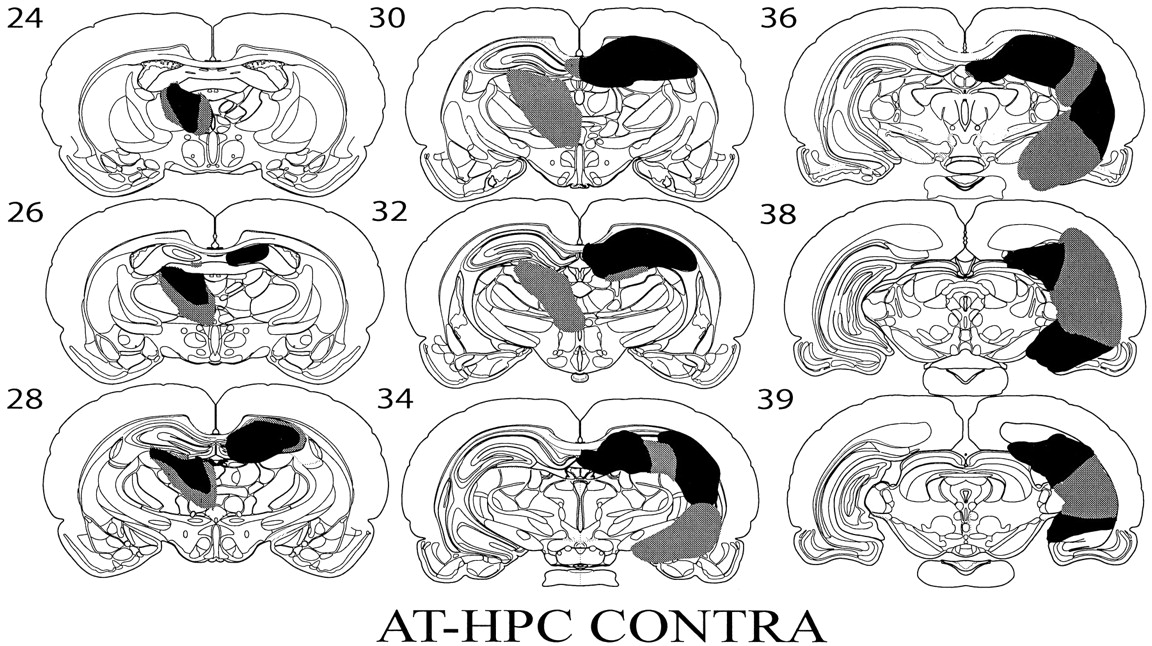
Diagrammatic reconstructions showing the cases with the largest (gray) and smallest (black) lesions in the AT-HPC Contra group. Thenumbers refer to the plate numbers from Swanson (1998).
In all cases, there was extensive damage to the hippocampus both dorsally and ventrally. (The term hippocampus is used to refer to the dentate gyrus, CA1–CA4, and subiculum). In five cases, there was near total cell loss in the dorsal hippocampus, whereas in two cases, there was some sparing of the dorsal lateral tip of the hippocampus, and in one case, there was some sparing of the dorsal CA3 field. In seven cases, the majority of the ventral hippocampus was damaged with only the most ventral tip of the hippocampus spared. In one case, most of the ventral hippocampus was damaged, but there was some sparing of the CA3 field.
AT-HPC Ipsi
Of the five rats with a combined ipsilateral anterior thalamic and hippocampal lesion, four rats had substantial damage in both the anterior thalamic nuclei and the ipsilateral hippocampus. The fifth case had sustained significant damage in the hippocampus and in the AD nucleus; however, much of the AV and AM thalamic nuclei were spared. In all animals, the amount of damage to the hippocampus was extensive both dorsally and ventrally (Fig. 2). In three cases, the only area of the dorsal hippocampus that was spared was the most lateral tip, whereas in the other two cases, there was some sparing of the CA1 field. In all cases, >80% of the ventral hippocampus had gone, with only the most ventral tip spared in all cases and some sparing of the dorsal tip in one case. The extent of hippocampal and anterior thalamic damage was comparable with that in the cases with contralateral lesions. The cases with the largest and smallest combined lesions are shown in Figure 2.

Diagrammatic reconstructions showing the cases with the largest (gray) and smallest (black) lesions in the AT-HPC Ipsi group. Thenumbers refer to the plate numbers from Swanson (1998).
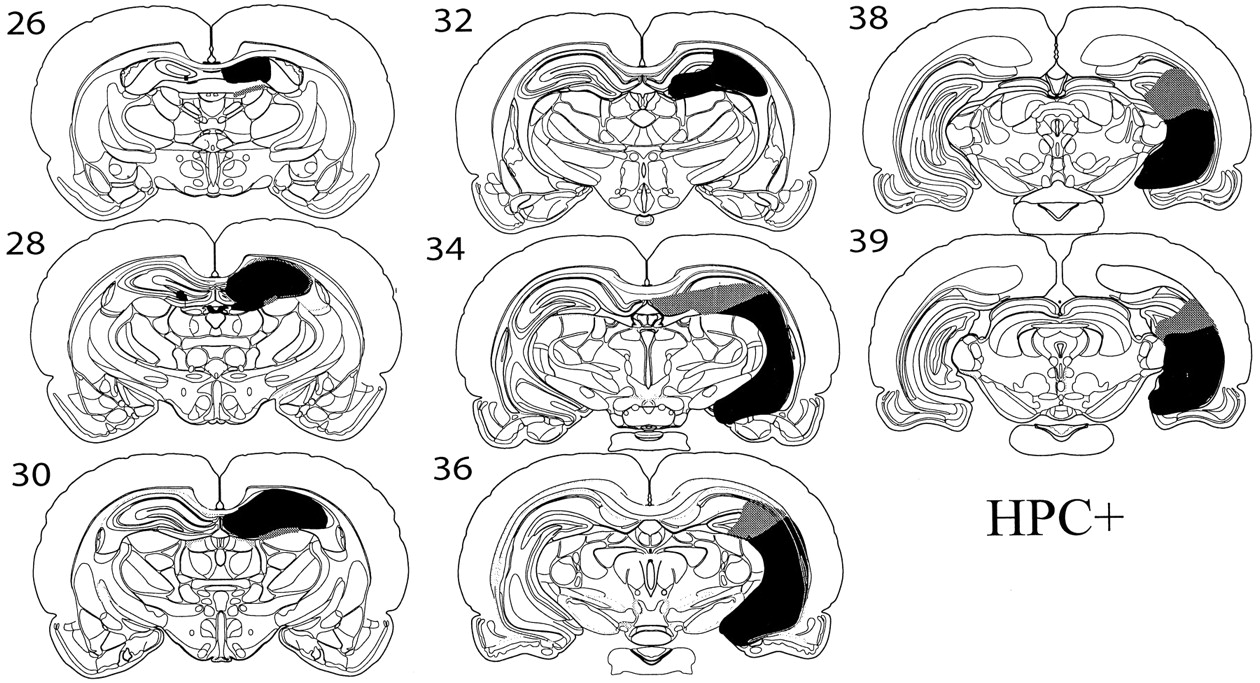
HPC+
This group comprised a set of four animals with a unilateral hippocampal lesion combined with a small amount of excitotoxic damage in the contralateral dorsal hippocampus but no anterior thalamic damage. In two of these cases, there was some additional minor damage to the dorsal edge of the AD nucleus in the hemisphere containing the large hippocampal lesion (Fig. 3).

Diagrammatic reconstructions showing the cases with the largest (gray) and smallest (black) lesions in the HPC+ group. Thenumbers refer to the plate numbers from Swanson (1998).
Behavior
Comparison of the HPC+ and the AT-HPC Ipsi groups
Statistical analysis revealed that the performance of the HPC+ group did not differ significantly from those animals in the AT-HPC Ipsi group in the T-maze task [F(1,7)≤ 1.0 (mean correct trials: HPC+, 16.3; AT-HPC Ipsi, 15.9); water maze acquisition, distance swum, F(1,7) ≤ 1.0 (mean distance: HPC+, 15.1 m; AT-HPC Ipsi, 13.3 m); radial arm maze, trials correct,F(1,7) = 1.34 (means: HPC+, 20.54; AT-HPC Ipsi, 19.40); radial arm maze, total trials to complete task,F(1,7) = 1.22 (means: HPC+, 12.46; AT-HPC Ipsi, 11.93)]. Because these two groups did not differ on any of the performance measures, they were combined and called the COMB Ipsi group for the subsequent analyses. However, additional analyses were also performed comparing the SHAM, AT-HPC Contra, and AT-HPC Ipsi groups without the inclusion of the HPC+ group to ensure that this group was not influencing the overall pattern of results.
Spatial forced alternation in the T-maze
The performance of the three groups of rats (SHAM, AT-HPC Contra, and COMB Ipsi) is depicted in Figure 4. ANOVA revealed a significant effect of lesion group [F(2,26) = 10.77 (p < 0.01)], a significant main effect of session [F(4,104) = 12.27 (p < 0.01)], and a significant lesion group by session interaction [F(8,104) = 2.32 (p < 0.03)]. Post hoc analyses showed that the performance of the AT-HPC Contra group was significantly worse (p < 0.01) than the SHAM and the COMB Ipsi groups, but the SHAM group did not differ significantly from the COMB Ipsi group (means: SHAM, 17.05; AT-HPC Contra, 14.00; COMB Ipsi, 16.07). Analysis of the simple main effects showed that the performance of the SHAM and COMB Ipsi groups improved significantly across the training sessions (p < 0.01), whereas the performance of the AT-HPC Contra group did not. A very similar pattern of results was obtained if the analyses were conducted on the data from the SHAM, AT-HPC Contra, and AT-HPC Ipsi groups (i.e., the Ipsi group without the inclusion of the HPC+ group). Once again, there was a significant effect of lesion [F(2,22) = 7.07 (p < 0.01)], and post hoc analyses revealed that the performance of the AT-HPC Contra group was significantly worse than that of the AT-HPC Ipsi group (p < 0.05). The SHAM group did not differ significantly from the AT-HPC Ipsi group (mean AT-HPC Ipsi, 15.92).

T-maze alternation. The graph shows the mean ± SEM number of correct responses made by each of the three groups (SHAM, COMB Ipsi, and AT-HPC Contra) in each block of three sessions (maximum of 18, chance of 9). SEM values not visible are smaller than the symbol.
Water maze
Acquisition. ANOVA of the length of the swim path to reach the escape platform revealed a significant difference between the lesion groups [F(2,26) = 6.22 (p < 0.01)] (Fig.5). Post hoc analyses showed that the AT-HPC Contra group swam significantly farther to reach the escape platform compared with the SHAM (p < 0.01) and COMB Ipsi (p < 0.05) groups (means: SHAM, 12.2 m; COMB Ipsi, 14.2 m; AT-HPC Contra, 19.1 m). There was a highly significant main effect of session [F(9,234) = 64.70 (p < 0.01)] as distance swum to reach the escape platform decreased across training sessions for all lesion groups. The lesion by session interaction did not reach significance [F(18,234) = 1.55 (p = 0.074)]. When the performance of the AT-HPC Ipsi group was compared with the SHAM and AT-HPC Contra groups, the analyses again revealed a significant main effect of lesion [F(2,22) = 4.57 (p < 0.05)]. Post hoc analyses showed that the distance swum by the AT-HPC Ipsi group (mean of 13.52) was significantly shorter than that of the AT-HPC Contra group (p < 0.05), but the AT-HPC Ipsi group did not differ from the SHAM group.

Water maze acquisition. The graph shows the mean ± SEM distance swum by the three groups (SHAM, COMB Ipsi, and AT-HPC Contra) to find the hidden platform within each session (made up of the mean of 4 trials).
Probe. Analysis of the percentage of time the animals spent in the correct quadrant during the 60 sec probe test showed a trend toward a difference between the performance of the three groups [F(2,26) = 2.95 (p = 0.07) (means: SHAM, 41.37%; COMB Ipsi, 41.01%; AT-HPC Contra, 27.63%)]. Additional analyses comparing the performance levels of the animals in the probe test revealed that the SHAM and COMB Ipsi groups spent a significantly greater proportion of time in the training quadrant (Fig. 6) than chance (25%) (t(11) = 4.03,p < 0.01; t(8) = 3.26, p < 0.03, respectively). In contrast, the AT-HPC Contra lesion group did not show a significant preference for the training quadrant (t(7) <1.0). Finally, an identical pattern of results was obtained when the comparisons were made between the AT-HPC Ipsi, AT-HPC Contra, and SHAM groups, [F(2,22) = 3.02 (p = 0.069) (mean AT-HPC Ipsi, 45.38%)].

Water maze probe. Mean percentage of time each group spent swimming in each of the four quadrants during the probe trial. The error bars represent the SEM.
Radial arm maze
ANOVA conducted on the number of correct arm choices made by the animals in the first eight trials (blocked across three sessions, maximum number of 24) revealed a significant difference between the lesion groups [F(2,26) = 13.81 (p < 0.01)], a significant main effect of session [F(5,130) = 7.27 (p < 0.01)], and a significant lesion group by session interaction [F(10,130) = 2.36 (p = 0.01)]. Post hoc analyses showed that the SHAM and COMB Ipsi lesion group made a significantly greater (p < 0.01) number of correct choices (means: SHAM, 6.87 trials; COMB Ipsi, 6.65 trials) than the AT-HPC Contra group (mean of 5.91). Analysis of the simple main effects showed that only the SHAM and COMB Ispi groups significantly improved their performance across the training sessions (p < 0.01). The data are presented in Figure7A.

Radial arm maze. A, The mean ± SEM number of correct choices made in the first eight trials of each session. B, The mean ± SEM number of total arms visited to retrieve eight reward pellets. In A andB, the scores are blocked into groups of three sessions so that a perfect score would be 24 in both cases.
ANOVA conducted on the total number of visits (Fig. 7B) taken to obtain all of the reward pellets and complete the task (blocked across three sessions, minimum of 24) also revealed a significant effect of lesion group [F(2,26) = 22.4 (p < 0.01)], a significant main effect of session [F(5,130) = 14.61 (p < 0.01)], and a significant lesion by session interaction [F(10,130) = 2.15 (p < 0.03)]. Post hoc analyses showed that the SHAM and COMB Ipsi lesion groups took significantly fewer trials per session (both p < 0.01) to complete the task compared with the AT-HPC Contra group (means: SHAM, 30.6; COMB Ipsi, 33.6; AT-HPC Contra, 44.4). Analysis of the simple main effects showed that all lesion groups improved across the training sessions.
A similar pattern of results was obtained, for both trials correct and total trials, when the analyses were conducted on the SHAM, AT-HPC Contra, and AT-HPC Ipsi groups (i.e., without the inclusion of the HPC+ group). There was a significant main effect of lesion for trials correct [F(2,22) = 9.53 (p < 0.01)] and total trials [F(2,22) = 15.29 (p < 0.01)] and a significant main effect of session for trials correct [F(17,374)= 2.13 (p < 0.01)] and total trials [F(17,374) = 3.47 (p < 0.01)]. Post hoc analyses revealed that for both performance indices (trials correct and total trials), the AT-HPC Ipsi group was significantly better than the AT-HPC Contra group (p < 0.01) but that the AT-HPC Ipsi group did not differ significantly from the SHAM group.
DISCUSSION
The present study compared the behavioral consequences of a unilateral anterior thalamic lesion combined with a hippocampal lesion placed in either the contralateral hemisphere or the same hemisphere. In addition to a surgical control group, a fourth group of rats had just a unilateral hippocampal lesion. In the T-maze alternation and radial arm maze tasks, those animals with lesions placed in contralateral hemispheres were significantly impaired compared not only with the sham control group but also importantly with those animals with lesions placed in a single hemisphere. The anterior thalamic–hippocampal contralateral group was also impaired in both the acquisition of the water maze task and the water maze probe task. In the probe task, which most clearly indicates how well the animals are able to use allocentric spatial strategies, the sham control group and the combined ipsilateral group spent a significantly greater proportion of time searching for the escape platform in the correct quadrant. In contrast, the amount of time the anterior thalamic–hippocampal contralateral group spent in the training quadrant did not differ significantly from chance, indicating that they had failed to learn the location of the escape platform. The lack of a significant difference between the amount of time the ipsilateral and contralateral groups spent in the correct quadrant during the probe trial is probably a consequence of the fact that the probe data are derived from a single session and are therefore subject to individual variability. For this reason, it might be more appropriate to use the delayed matching-to-place task in the water maze (Steele and Morris, 1999) for any future studies.
Consistent with the present disconnection results, there was a lack of difference within the COMB Ipsi group, because the addition of a unilateral anterior thalamic lesion did not exacerbate the effect of a unilateral hippocampectomy. Together, these disconnection findings provide strong support for the view that the hippocampus and anterior thalamic nuclei function in a closely related way for the learning of spatial tasks taxing allocentric processing. Although the T-maze and radial arm maze tasks are not necessarily selective tests of allocentric spatial memory, previous studies with the same strain and protocol have shown that normal rats consistently do use allocentric cues when performing these tasks (Warburton et al., 1997; Bussey et al., 1999).
These results support and extend previous findings that showed that disconnection of the anterior thalamic and hippocampus, by a lesion of the fimbria fornix, also produced behavioral impairments in the same tasks as those used in the present study (Warburton et al., 2000). The present disconnection procedure (lesion of hippocampus rather than the fornix) did, however, result in more severe learning deficits in the behavioral tasks. This difference was most evident in the water maze task and presumably reflects the more complete disconnection between the two sites. This is because fornix lesions spare alternate direct routes between the presubiculum and parasubiculum and the anterior thalamic nuclei (Meibach and Siegel, 1977). Fornix lesions will also spare projections from the hippocampal formation to the posterior cingulated–retrosplenial cortex, another route by which the hippocampus may interact with the anterior thalamic nuclei. In addition, a unilateral fornix transection will spare crossed projections, including interhippocampal connections via the hippocampal commissure. Evidence that these commissural projections can help to support spatial memory performance (Olton et al., 1982; Warburton et al., 2000) reinforces the view that unilateral fornix lesions can only partially disconnect the hippocampus.
An additional explanation for why the effects of a crossed hippocampal–anterior thalamic lesion are more marked than the effects of a crossed fornix–anterior thalamic lesion is that projections from the anterior thalamus to the hippocampal formation pass via the cingulum bundle and not the fornix (Shibata, 1993). The hippocampal afferents would consequently be unaffected by the fornix lesion. Thus, the two disconnection procedures differ markedly in the extent to which information is truly disconnected between the hippocampus and the thalamus. This sparing may also explain why the effects of bilateral anterior thalamic lesions on the Morris water maze task can be more severe than those seen after fornix lesions (Warburton et al., 1999)
The data from the present study show that interactions between the hippocampus and anterior thalamus are required for the operation of several spatial memory tasks. Although it may seem intuitive to assume that the present disconnection primarily affects information passing from the hippocampus to the anterior thalamic nuclei (Parmeggiani et al., 1974), we cannot distinguish the contribution from the loss of inputs from the anterior thalamic nuclei to the hippocampus. Indeed, recent research on the importance of the projections from the anterior thalamic nuclei to the hippocampal formation primarily for the processing of head direction information (Taube, 1995; Taube and Muller, 1998) underlines the importance of these thalamic inputs. For example, lesions of the anterior thalamic nuclei result in a loss of head direction cells in the postsubiculum, but the opposite pattern of results is not observed (Goodridge and Taube, 1997), although hippocampal lesions have been shown to modify the thalamic head direction responses of the cell.
The current study is also consistent with previous studies in both human and nonhuman primates showing that the hippocampus, fornix, and anterior thalamus are important for spatial memory (Gaffan, 1992;Parker and Gaffan, 1997). Investigations of episodic-like memory in monkeys, using an “object-in-place” discrimination task, revealed that lesions of the fornix, mammillary bodies, and anterior thalamic nuclei all produce similar patterns of deficits (Gaffan, 1992;Parker and Gaffan, 1997). Furthermore, studies of human patients with neuronal loss or disconnection of the anterior thalamic nuclei, either as a result of Korsakoff's disease or thalamic infarction, have concluded that these patients all show patterns of episodic memory loss similar to those seen in patients with lesions in the medial temporal lobe (Harding et al., 2000; Van der Werf et al., 2000). Although it is only possible to infer this functional link from clinical studies, the present experiments test the link directly and provide clear evidence for the importance of hippocampal–anterior thalamic interactions.
The present results thus support the suggestion that the anterior thalamus and hippocampus form key components of a neural circuit important for the processing of mnemonic information, which extends beyond the medial temporal lobe (Aggleton and Brown, 1999). The present disconnection study only examined the processing of spatial information, and there are data suggesting that projections from the hippocampus to the anterior thalamic nuclei via the mamillothalamic tract are also important for discriminative avoidance learning processes (Gabriel et al., 1995). However, it remains to be determined whether these disconnection results extend to other forms of memory in the rat (Aggleton and Brown, 1999; Célérier et al., 2000). Finally, it remains to be established whether the critical flow of information for spatial memory processing is from the anterior thalamic nuclei to the hippocampus, from the hippocampus to the anterior thalamic nuclei, or both.
Footnotes
This research was supported by a project grant from the Wellcome Trust.
Correspondence should be addressed to Dr. E. C. Warburton at her present address: Medical Research Council Centre for Synaptic Plasticity, Department of Anatomy, University of Bristol, Medical School, University Walk, Bristol BS8 1TD, UK. E-mail:e.c.warburton{at}bristol.ac.uk.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}