

Abstract
Cutaneous spinal sensory transmission appears to lack inhibitory control in the newborn spinal cord, but the properties of GABAergic and glycinergic synapses in the neonatal dorsal horn have not been characterized. Whole-cell patch-clamp recordings from rat superficial dorsal horn neurons in spinal cord slices at postnatal day 0 (P0) to P2, P6–P7, and P13–P14 revealed an age-dependent increase in the frequency of spontaneous IPSCs, which were abolished by the GABAA receptor (GABAAR) antagonist bicuculline between P0 and P7 but not at P14. GABAAR-mediated miniature IPSCs (mIPSCs), but not glycinergic mIPSCs, were present at birth, and GABA mIPSCs remained more frequent than glycine mIPSCs at all ages. Sciatic nerve stimulation resulted in IPSCs with both GABAergic and glycinergic components, although a larger contribution arose from GABAA receptors at all ages. In gramicidin perforated patch-clamp recordings, exogenous GABA applications produced depolarization in 40% of neurons at P0–P2, but the reversal potential of GABA-evoked currents (EGABA) was consistently more negative than action potential threshold at this age. By P6–P7, GABA evoked only membrane hyperpolarization. The GABABR agonist baclofen elicited an outward current in all neurons with peak amplitudes observed by P6–P7 and abolished sciatic nerve-evoked monosynaptic glutamatergic EPSCs in all groups. The results show considerable postnatal development of inhibitory processing in the dorsal horn with GABAergic mechanisms initially dominant over glycinergic events. GABAAR-mediated depolarizations during the first postnatal week are likely to be important for the maturation of spinal networks but do not provide a major excitatory drive to the newborn dorsal horn.
Introduction
Cutaneous sensory processing in the postnatal period is characterized by an apparent lack of inhibition. Cutaneous flexion reflexes evoked by mechanical stimulation of the skin are exaggerated in neonatal humans, kittens, and rats (Andrews and Fitzgerald, 1994; Fitzgerald and Jennings, 1999). The cutaneous receptive fields of neonatal rat dorsal horn neurons are large at birth and decrease rapidly over the first 2 postnatal weeks (Fitzgerald, 1985; Torsney and Fitzgerald, 2002). In addition, excitation of dorsal horn neurons at postnatal day 0 (P0) to P3 often results in a prolonged afterdischarge of action potentials (APs) that decreases with age (Fitzgerald, 1985). Because primary afferent inputs to the dorsal horn strengthen with postnatal age (Fitzgerald and Jennings, 1999; Baccei et al., 2003), these properties suggest that inhibitory systems in the spinal cord may be immature in the neonate. However, little is known about the properties of inhibitory synaptic transmission in the developing dorsal horn.
GABA and glycine serve as the main inhibitory neurotransmitters in the superficial dorsal horn of the adult spinal cord (Todd and Spike, 1993) through the activation of ligand-gated anion channels [Cl–-permeable GABAA receptors (GABAARs), GABACRs, or glycinergic receptors (GlyRs)] or G-protein-coupled GABAB receptors (Bormann, 1988). A combination of GABAAR and GlyR antagonists are required to block primary afferent-evoked IPSCs in adult substantia gelatinosa (lamina II) neurons (Moore et al., 2002). At present, it is unknown whether GABAergic and glycinergic systems are both functional during early postnatal life. An absence of one or several components of synaptic inhibition could contribute to the enlarged receptive fields and afterdischarges seen in neonatal dorsal horn neurons.
These properties could also be potentially explained by excitation of immature dorsal horn neurons by GABA or glycine. In a variety of CNS regions (including the hippocampus, neocortex, and hypothalamus), high intracellular chloride concentrations ([Cl–]i) found in neonatal neurons result in membrane depolarization after GABAAR or GlyR activation (Ben Ari, 2002; Owens and Kriegstein, 2002), sometimes resulting in action potential discharge (Ben Ari et al., 1989; Chen et al., 1996; Gao and van den Pol, 2001). In cultured embryonic dorsal horn neurons, GABA or glycine application causes depolarization and a rise in intracellular [Ca2+], which progressively decline over time in culture (Reichling et al., 1994; Wang et al., 1994). However, it is still unclear whether GABA-mediated depolarizations persist in the intact superficial dorsal horn throughout the first 2 postnatal weeks and are sufficient to elicit action potentials.
The present study characterizes the properties of GABAergic and glycinergic signaling in the immature superficial dorsal horn, an area known to be crucial for the processing of painful stimuli, to determine whether the mechanisms of synaptic inhibition that dominate in the adult spinal cord are also functional at early postnatal ages.
Materials and Methods
All experimental procedures were performed according to the United Kingdom Animals (Scientific Procedures) Act of 1986.
Preparation of spinal cord slices. Neonatal Sprague Dawley pups (P0–P14) were humanely killed by an overdose of halothane (5% in medical oxygen) and decapitated. The spinal column was quickly removed and placed in an ice-cold dissection solution containing the following (in mm): 125 NaCl, 2.5 KCl, 25 NaHCO3, 1.0 NaH2PO4, 6 MgCl2, 0.5 CaCl2, and 25 glucose (continuously bubbled with 95% O2–5% CO2). After a ventral laminectomy was performed, the spinal cord was removed and immersed in low-melting-point agarose (3% in above dissection solution; Invitrogen, Paisley, UK) at 37°C. After cooling, the block of agarose was glued to the chamber of a Vibroslice tissue slicer (HA-752; Campden Instruments, Leicester, UK), and 400 μm sagittal slices were cut from the lumbar region. For experiments involving the recording of IPSCs evoked by primary afferent stimulation, the sciatic nerve and the L4 and L5 DRGs were exposed after the ventral laminectomy and removed with the spinal cord. Subsequently, using a modified Vibroslice, a sagittal slice (400–600 μm) was cut that preserves the entire L4–L5 dorsal root entry zone with the L4 and L5 DRGs and sciatic nerve attached. The slices were transferred to a chamber filled with oxygenated dissection solution and allowed to recover for 1.5–2 hr at room temperature.
Patch-clamp recordings. After recovery, slices were transferred to a submersion-type recording chamber (RC-22; Warner Instruments, Hamden, CT) and mounted on the stage of an upright microscope (Axioskop 2; Zeiss, Welwyn Garden City, UK). The slices were continually perfused at room temperature with oxygenated artificial CSF (aCSF) solution containing the following (in mm): 125 NaCl, 2.5 KCl, 25 NaHCO3, 1.0 NaH2PO4, 1.0 MgCl2, 2.0 CaCl2, and 25 glucose (at a rate of 1–3 ml/min).
Patch electrodes were constructed from thin-walled single-filamented borosilicate glass (1.5 mm outer diameter; World Precision Instruments, Hertfordshire, UK) using a two-stage vertical microelectrode puller (PP-830; Narishige, London, UK). Pipette resistances ranged from 5 to 7 MΩ, and seal resistances were >1 GΩ. For voltage-clamp experiments in the whole-cell recording configuration, electrodes were filled with a solution containing the following (in mm): 130 Cs-gluconate, 10 CsCl, 10 HEPES, 11 EGTA, 1.0 CaCl2, and 2.0 MgATP, pH 7.2 (305 mOsm). Current-clamp experiments used an electrode solution containing the following (in mm): 130 K-gluconate, 10 KCl, 10 HEPES, 1.0 EGTA, 0.1 CaCl2, and 2.0 MgATP, pH 7.2 (305 mOsm).
For perforated patch-clamp experiments, gramicidin (Sigma, Dorset, UK) was dissolved at 2.5 mg/ml in DMSO and stored at –20°C for up to 1 week. Electrode tips were filled with the above K-gluconate solution and subsequently backfilled with gramicidin diluted in the same solution to a final concentration of 25 μg/ml. Access resistance (Racc) was carefully monitored during perforation and throughout the experiment, and any neurons in which a sudden drop in Racc occurred were discarded. In voltage-clamp experiments to measure the reversal potential for GABA-evoked currents (EGABA), holding potentials were corrected for voltage errors across uncompensated series resistance according to Vcorrected = Vcommand–IRacc. Membrane voltages were also adjusted for liquid junction potentials (approximately–14 mV) calculated using JPCalc software (P. Barry, University of New South Wales, Sydney, Australia; modified for Axon Instruments, Union City, CA).
Dorsal horn neurons were visualized with infrared-differential interference contrast, and patch-clamp recordings were obtained using a Multiclamp 700A amplifier (Axon Instruments). Spontaneous IPSCs (sIPSCs) were recorded at a holding potential of 0 mV, thus minimizing the contribution of NMDA and AMPA/kainate receptor-mediated events (Yoshimura and Nishi, 1993). Miniature IPSCs (mIPSCs) were isolated via the application of 500 nm tetrodotoxin (TTX) with 20 μm d(–)2-amino-5-phosphonovaleric acid [d(–) AP-5] and 10 μm 1,2,3,4-tetrahydro-6-nitro-2,3-dioxo-benzo(f)quinoxaline-7-sulfonamide (NBQX) added to block currents through ionotropic glutamate receptors. Bicuculline methiodide (10 μm) or strychnine (0.5 μm) was added to the above solution to isolate GlyR-mediated or GABAAR-mediated mIPSCs, respectively. IPSCs were evoked via electrical stimulation of the sciatic nerve with a suction electrode connected to a constant-current stimulator (NeuroLog system; Digitimer, Hertfordshire, UK).
Postsynaptic GABABR effects were investigated using the above K+-gluconate solution with 0.3 mm Na3GTP added to avoid depletion of intracellular GTP. Baclofen was bath applied in the presence of 500 nm TTX, 20 μm d(–) AP-5, 10 μm NBQX, 10 μm bicuculline, and 0.5 μm strychnine.
Currents were filtered at 4–6 kHz through a –3 dB, four-pole low-pass Bessell filter, digitally sampled at 20 kHz, and stored on a personal computer (Viglen, Middlesex, UK) using a commercially available data acquisition system (Digidata 1322A with pClamp 8.0 software; Axon Instruments).
Drugs. GABA, (–)-bicuculline methiodide, strychnine, d(–) AP-5, gramicidin D, R(+)-baclofen hydrochloride, GTP (Na3GTP), and guanosine 5′-O-(2-thiodiphosphate) (GDPβS) were purchased from Sigma (Dorset, UK). TTX, NBQX, and CGP 55845 [(2S)-3-{[(1S)-1-(3,4-dichlorophenyl)ethyl]amino-2-hydroxypropyl)(phenylmethyl) phosphinic acid] were purchased from Tocris Cookson (Bristol, UK). All drugs except GABA were bath applied at 1–3 ml/min. GABA (1 mm) was pressure ejected (50–200 msec, 5–6 psi) through a patch electrode near the recorded neuron using the Picospritzer application system (General Valve Operation; Parker Hannifin, Fairfield, NJ).
Data analysis and statistics. Data were analyzed via visual inspection using Mini Analysis (version 5.4.1; Synaptosoft, Decatur, GA) and Clampfit (Axon Instruments) software. The threshold for mIPSC and sIPSC detection was set at twice the mean amplitude of the background noise. Multiphasic IPSCs evoked by primary afferent stimulation were quantified by measuring the area under the curve (AUC) for a 1 sec period from the IPSC onset. Because it has previously been observed that IPSC (or EPSC) frequencies often fail to exhibit a normal distribution in newborn dorsal horn neurons, we used nonparametric statistical tests to determine whether significant differences in IPSC frequency existed between various groups (Wilcoxon matched-pairs or Mann–Whitney tests for two groups, Kruskal–Wallis test for more than two groups; Prism 3.0 software; GraphPad Software, San Diego, CA). Nonparametric tests were also used in cases in which the number of observations was insufficient (n < 24) to definitively conclude that data were distributed in a Gaussian manner. Data are expressed as means ± SEM.
Results
Patch-clamp recordings were obtained from 306 neurons in the neonatal rat dorsal horn from the following three age groups: P0–P2 (n = 131), P6–P7 (n = 91), and P13–P14 (n = 84). All neurons were located in the superficial dorsal horn (laminas I–II) as judged by visual inspection. Because the sampled neurons were located at an average distance (measured from the dorsal white matter) of 60.1 ± 1.4 μm at P0–P2, 76.1 ± 2.3 μm at P6–P7, and 93.9 ± 2.7 μm at P13–P14, it seems likely that the majority of these neurons were located in lamina II.
Spontaneous and miniature IPSCs in the superficial dorsal horn are mediated by GABAA receptors during the first postnatal week
In the whole-cell patch-clamp experiments of the present study, sIPSCs are observed as transient outward currents from a holding potential of 0 mV (Fig. 1A). The frequency of sIPSCs recorded in aCSF increased significantly over the first 2 postnatal weeks (Fig. 1B) from 0.22 ± 0.08 Hz (n = 35) at P0–P1 to 0.95 ± 0.28 Hz (n = 31) by P6–P7 (p < 0.001; Kruskal–Wallis test) and 2.09 ± 0.58 Hz (n = 29) by P13–P14 (p < 0.001). In contrast, the size of the sIPSCs did not change with advancing postnatal age, with mean amplitudes of 15.30 ± 1.79 pA (n = 26) at P0–P1, 18.52 ± 1.42 pA (n = 29) at P6–P7, and 18.63 ± 1.63 pA (n = 29) at P13–P14 (p > 0.20; one-way ANOVA) (Fig. 1C).

sIPSCs in the superficial dorsal horn as a function of postnatal age. A, IPSCs are evident as transient outward currents from a holding potential of 0 mV. B, sIPSC frequency increases significantly with postnatal age (*p < 0.001 compared with P0–P1; Kruskal–Wallis test). C, The mean sIPSC amplitude was not significantly different between groups.
To determine the relative contribution of GABAARs and GlyRs to the observed sIPSC frequency, the selective GABAAR antagonist (–)bicuculline methiodide (10 μm) was added to the bath solution. During the first postnatal week, spontaneous activity appeared to be mediated predominantly by GABAARs because bicuculline significantly reduced the overall sIPSC frequency by 89.3 ± 6.8% at P0–P1 (from 0.31 ± 0.11 to 0.05 ± 0.04 Hz; n = 11; p < 0.005; Wilcoxon matched-pairs test), with a total abolition of sIPSCs (Fig. 2A) observed in 6 of the 11 neurons sampled at this age. A recovery in sIPSC frequency was observed during washout of bicuculline (data not shown), suggesting that a time-dependent rundown of synaptic currents cannot explain the decrease in sIPSC frequency during bicuculline application. At P6–P7, the vast majority of IPSCs were still mediated by GABAARs because bicuculline decreased the mean sIPSC frequency by 88.5 ± 5.5% (from 0.83 ± 0.18 to 0.07 ± 0.03 Hz; n = 15; p < 0.0001). By this age, most neurons did exhibit both GABAAR- and GlyR-mediated sIPSCs (3 of 15 cells showed a complete absence of sIPSCs in bicuculline). In contrast, bicuculline application did not significantly affect sIPSC frequency in the overall population of P13–P14 neurons sampled (aCSF, 1.82 ± 0.78 Hz; bicuculline, 1.44 ± 0.55 Hz; n = 18; p > 0.20; Wilcoxon test). This seems to reflect the variability in bicuculline effects on individual dorsal horn neurons at this age rather than a disappearance of GABAAR activity. Bicuculline treatment significantly decreased the sIPSC frequency in 6 of 18 cells at P13–P14 (p < 0.05; Kolmogorov-Smirnov two-sample test), with complete abolition of sIPSCs observed in only two of these neurons, suggesting that most P13–P14 neurons possessed both GABAAR- and GlyR-mediated sIPSCs. Meanwhile, bicuculline significantly elevated the overall sIPSC frequency in 4 of 18 neurons by an average of 165.7 ± 102.5% (from 1.79 ± 0.99 to 3.09 ± 1.20 Hz; n = 4), which may result from disinhibition of spinal interneurons. The mean frequency of sIPSCs observed in the presence of bicuculline at different postnatal ages is illustrated in Figure 2B and demonstrates that spontaneous glycine-mediated currents are rare in the first postnatal week. This is supported by the finding that sIPSC frequency in the presence of the selective GlyR antagonist strychnine (0.5 μm) was 93.1 ± 15.1% of control in P0–P7 neurons (aCSF, 0.25 ± 0.07 Hz; strychnine, 0.23 ± 0.06 Hz; n = 13; p > 0.30; Wilcoxon test) (Fig. 2C). The presence of functional GlyRs in the membrane was confirmed by pressure ejections of exogenous glycine (1 mm, 200 msec) near the recorded cell in the presence of bicuculline (10 μm). Glycine application resulted in an outward current in all cells tested (n = 18) (Fig. 2D), with a mean amplitude of 417.0 ± 42.4 pA at a holding potential of 0 mV. Pretreatment with strychnine (5 μm) abolished the glycine-evoked currents (n = 6; data not shown).

sIPSCs are predominantly mediated by GABAARs during the first postnatal week. A, Abolition of sIPSCs by the GABAAR antagonist bicuculline (10 μm; Bicuc) in a P7 neuron. B, Frequency of glycinergic sIPSCs (isolated with 10 μm bicuculline) increases during the second postnatal week (†p < 0.001; *p < 0.05; Kruskal–Wallis test). C, Application of the GlyR antagonist strychnine (0.5 μm; Strych) does not significantly affect sIPSC frequency in P0–P7 dorsal horn neurons (n = 13), consistent with a dominant role for GABAARs during the first week. D, Exogenous applications of glycine (1 mm) elicited outward currents in all cells examined at P0–P7 (n = 18).
The predominance of GABAAR-mediated events during the first postnatal week is also illustrated by the recording of mIPSCs in 500 nm TTX with 20 μm d(–) AP-5 and 10 μm NBQX added to block currents through ionotropic glutamate receptors. The frequency of TTX-resistant mIPSCs was significantly lower than the rate of sIPSCs at all ages tested (compare Figs. 1B, 3A), suggesting a role for presynaptic action potentials in sIPSC generation. Importantly, no glycinergic mIPSCs (isolated via the bath application of 10 μm bicuculline) were observed in neurons sampled at P0–P1 (n = 14) (Fig. 3A). In addition, bath application of ruthenium red (100 μm), known to enhance mIPSC frequency via calcium-independent mechanisms (Trudeau et al., 1996; Keller et al., 2001), failed to reveal Gly mIPSCs in 7 of 10 neurons examined at P0–P1 (data not shown). Meanwhile, GABA mIPSCs (recorded in the presence of 0.5 μm strychnine) were observed at a rate of 0.005 ± 0.001 Hz at this age (Fig. 3A). Gly mIPSCs were sporadically observed across the population of P6–P7 neurons (0.004 ± 0.003 Hz; n = 11) but were absent in 8 of the 11 neurons examined. The frequency of GABA mIPSCs increased significantly over the first 2 postnatal weeks to 0.14 ± 0.02 Hz (n = 13) at P6–P7 and 0.17 ± 0.12 Hz (n = 14) at P13–P14 (p < 0.001; Kruskal–Wallis test). As depicted in Figure 3A, the frequency of Gly mIPSCs remained low at P13–P14, with a mean rate of 0.011 ± 0.008 Hz (n = 10), and was significantly lower than the GABA mIPSC frequency at all ages tested (p < 0.001; Mann–Whitney test).

GlyR- and GABAA R-mediated mIPSCs in the P0–P14 dorsal horn. A, Frequency of GABAAR mIPSCs increases with postnatal age (*p < 0.001 compared with P0–P1; Kruskal–Wallis test). GlyR mIPSCs were not observed in any of the P0–P1 neurons tested (n = 14) but were observed by the end of the first postnatal week at a significantly lower rate than GABAAR events (†p < 0.001; Mann–Whitney test). B, Mean mIPSC amplitudes at different postnatal ages (*p < 0.01, **p < 0.001 compared with P0–P1 with Kruskal–Wallis test; †p < 0.05, ‡p < 0.005 compared with GABA mIPSC amplitude at given age with Mann–Whitney test).
The size of GABA mIPSCs was significantly lower at birth than later postnatal ages, with mean amplitudes of 5.41 ± 0.62 pA (n = 7) at P0–P1, 12.51 ± 2.07 pA (n = 13) at P6–P7 (p < 0.001; Kruskal–Wallis test), and 9.03 ± 0.49 pA (n = 14) at P13–P14 (p < 0.01). In the few neurons that displayed Gly mIPSCs in the P6–P7 and P13–P14 groups, the size of the Gly mIPSCs were significantly smaller than GABA mIPSCs, with average amplitudes of 5.76 ± 0.46 pA at P6–P7 (n = 3; p < 0.05; Mann–Whitney test) and 5.11 ± 0.33 pA at P13–P14 (n = 5; p < 0.005) (Fig. 3B).
Pharmacological properties of primary afferent-evoked IPSCs in the neonatal dorsal horn
Electrical stimulation of the sciatic nerve at C-fiber intensity (1 mA, 1 msec at 0.033 Hz) in aCSF evoked multiphasic IPSCs in neonatal dorsal horn neurons of all ages from a holding potential of 0 mV. Bath application of bicuculline (10 μm) led to a substantial reduction in the composite IPSC, and the subsequent addition of strychnine (0.5 μm) abolished the outward current (Fig. 4A), confirming the selective involvement of GABAARs and GlyRs in the observed response. To quantify the respective contributions of the different receptors to the evoked IPSC, electronic subtraction was used to isolate the bicuculline-sensitive and strychnine-sensitive components of the IPSC. For each component, the AUC was calculated for a 1 sec period after the onset of the IPSC as a measurement of the net movement of charge through the receptors.

Primary afferent-evoked IPSCs in the neonatal superficial dorsal horn. A, Multiphasic IPSCs were evoked from a holding potential of 0 mV via electrical stimulation of the sciatic nerve (1 mA, 1 msec, 0.033 Hz) and dissected into its composite subtypes with selective GABAAR and GlyR antagonists. For each cell, the areas under the bicuculline- and strychnine-sensitive waveforms were calculated for a 1 sec period from the onset of the IPSC. B, Average values of the AUC for GABAAR- and GlyR-mediated components at different postnatal ages (†p < 0.01, ‡p < 0.05 compared with P2 with Kruskal–Wallis test; *p < 0.05 compared with GABAAR with Mann–Whitney test).
As documented in Figure 4B, the GABAAR-mediated component of the evoked IPSC significantly increased during the first postnatal week, because the mean AUC for the bicuculline-sensitive currents was 17.26 ± 4.14 nA/msec (n = 15) at P2, 97.78 ± 21.62 nA/msec by P6–P7 (n = 17; p < 0.001; Kruskal–Wallis test), and 73.50 ± 15.11 nA/msec at P13–P14 (n = 16; p < 0.01). The GlyR-mediated component also increased with post-natal age, with mean AUCs of 2.96 ± 0.81 nA/msec at P2 (n = 15), 14.92 ± 4.30 nA/msec at P6–P7 (n = 17), and 24.22 ± 5.71 nA/msec (n = 16) at P13–P14 (p < 0.05 compared with P2 group). In addition, the GlyR component of the evoked IPSC was significantly smaller than the GABAAR component at all ages tested (p < 0.005; Mann–Whitney test) (Fig. 4B), further supporting the dominant role of GABAARs in spontaneous and evoked neurotransmission in the neonatal dorsal horn.
GABAAR activation causes subthreshold depolarizations in dorsal horn neurons with intact [Cl–]i during the first postnatal week
The effect of GABAAR activation on the excitability of superficial dorsal horn neurons at different post-natal ages (P0–P1, P6–P7, and P13–P14) was evaluated using perforated patch-clamp recordings with gramicidin, which forms pores that are impermeable to Cl– ions (Kyrozis and Reichling, 1995; Akaike, 1996). In the presence of 20 μm d(–)AP-5, 10 μm NBQX, 0.5 μm strychnine, and 2 μm CGP 55845 (a selective GABABR antagonist), GABA (1 mm, 200 msec) was applied via pressure ejection near the recorded neuron at its resting membrane potential (Vrest), and the membrane potential (Vm) was subsequently monitored in the current-clamp configuration. The average Vrest was –58.1 ± 1.1 mV at P0–P1 (n = 15), –60.1 ± 1.1 mV at P6–P7 (n = 15), and –69.1 ± 1.5 mV at P13–P14 (n = 16). It should be noted that these voltages were not corrected for any errors that can result from the higher ratio of input resistance to seal resistance characteristic of immature neurons (Tyzio et al., 2003).
During the first days of postnatal life (P0–P1), GABA evoked depolarizations in 40% (6 of 15) of superficial dorsal horn neurons at Vrest (Fig. 5B,C). In these neurons, Vm at the peak of the depolarization averaged –50.5 ± 3.7 mV (n = 6). However, the depolarizations were not sufficient to elicit APs from Vrest in any of the neurons, although APs were evoked by intracellular current injection at an average threshold of –42.2 ± 2.5 mV (n = 13). The remaining cells in the P0–P1 group hyperpolarized in response to GABA (Fig. 5A,C) to an average Vm of –68.6 ± 2.2 mV (n = 9). In contrast, at P6–P7, GABA produced hyperpolarization in all cells examined (p < 0.01; χ2 test) (Fig. 5D) to a mean Vm of –72.6 ± 1.2 mV, whereas the mean AP threshold was –41.2 ± 1.9 mV at this age (n = 15). Similarly, 15 of 16 cells at P13–P14 hyperpolarized in response to GABA application (Fig. 5E) to a peak Vm of –78.3 ± 1.2 mV, which was significantly more negative than the peak hyperpolarizations seen at P0–P1 (p < 0.01; Kruskal–Wallis test). In a single P13–P14 neuron with a Vrest of –80 mV, we observed a depolarization to –73 mV in response to GABA application. The average AP threshold in this age group was –47.4 ± 1.7 mV (n = 15).

Effect of GABA on Vm of neonatal dorsal horn neurons with intact [Cl–]i. A, Perforated patch-clamp recording showing example of hyperpolarization in response to GABA application (1 mm, 200 msec) in a P0 neuron at its resting membrane potential. B, GABA application results in membrane depolarization but no action potential discharge in a different P0 neuron. C–E, Plots depicting the response to GABA in individual neurons at different ages. Vrest, Resting membrane potential of the cell at the time of GABA application; Vpeak in GABA, membrane voltage at the peak of the GABA response in the same neuron. There was a significant difference in the distribution of depolarizing versus hyperpolarizing responses across the different age groups (p < 0.01; χ2 test).
At the end of each experiment, brief pulses of negative pressure were applied to the electrode to rupture the membrane and establish a whole-cell recording configuration. Under these conditions, voltage-clamp recordings demonstrated that the reversal potential for GABA-evoked currents was close to the ECl predicted by the Nernst equation, suggesting that a Cl– conductance underlies the effects of GABA on Vm (data not shown). Also, the effects of GABA were abolished by pretreatment with 100 μm bicuculline, confirming the role of GABAARs in the observed voltage responses. It should be noted that, when the neurons were dialyzed with a high [Cl–]i solution (150 mm KCl and 10 mm HEPES) in the whole-cell recording configuration, identical GABA applications (1 mm, 200 msec) from Vrest led to depolarization and AP discharge in all cells examined (data not shown), demonstrating that the ejection parameters produced sufficient concentrations of GABA at the recorded neuron to evoke APs if ECl was significantly more positive than the AP threshold (i.e., ECl of ∼0 mV).
In the six neurons that depolarized in response to GABA at P0–P1 (Fig. 5B,C), the activation of GABAARs will also produce significant shunting of the membrane that may prevent the firing of APs during the slow depolarizations, even if ECl values were located above AP threshold. To address this possibility, we measured EGABA under perforated patch-clamp conditions (Fig. 6A,B) in an additional group of P0–P1 neurons. Meanwhile, APs were elicited by intracellular current injections from Vm of –60 to –65 mV in the same cells, and the AP threshold was measured to allow for a direct comparison of EGABA and AP threshold in individual neurons (Fig. 6B). EGABA was located at more negative membrane potentials than the AP threshold in all cells examined at this age (n = 11). As illustrated in Figure 6C, the average value of EGABA was –65.3 ± 2.1 mV at P0–P1, which was significantly lower than the mean AP threshold in the same neurons (–44.6 ± 1.2 mV; n = 11; p = 0.001; Wilcoxon matched-pairs test). This strongly suggests that the absence of GABA-evoked APs in the current-clamp experiments results from the relationship between EGABA and AP threshold rather than the shunting action of the GABAAR conductance.
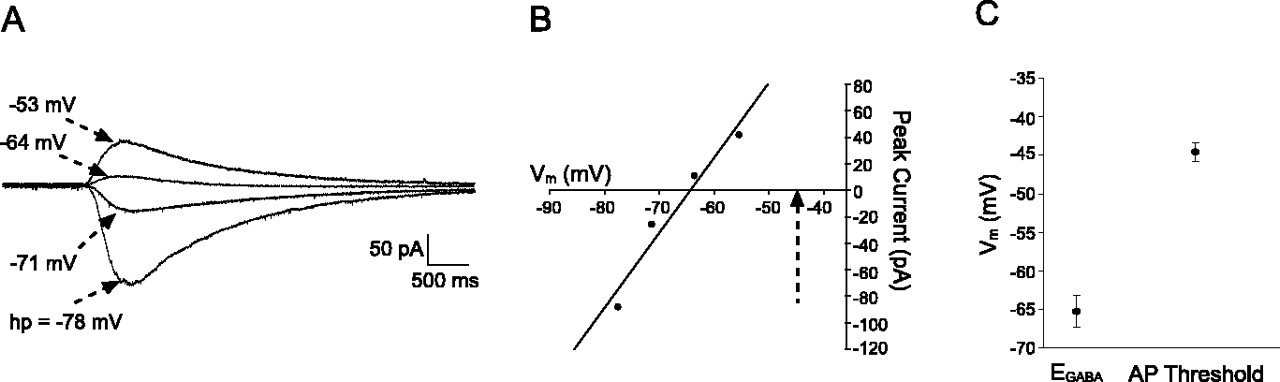
Comparison of EGABA and action potential threshold in newborn dorsal horn neurons. A, Perforated patch-clamp recording showing currents evoked by GABA application (1 mm, 200 msec) from different holding potentials (hp). Reported holding potentials have been corrected for errors attributable to uncompensated series resistance. B, Current versus voltage plot for the same neuron illustrated in A showing an EGABA of –64.4 mV as measured by linear regression analysis (r2 = 0.95). The dotted arrow identifies the AP threshold in the same neuron. C, Average values for EGABA and AP threshold in the sampled population of P0–P1 dorsal horn neurons (n = 11). EGABA was more negative than AP threshold in all cells examined.
Presynaptic and postsynaptic GABABR effects in the neonatal dorsal horn
To examine the development of presynaptic inhibition by the GABAB receptor in the superficial dorsal horn, monosynaptic glutamatergic EPSCs were elicited in neurons (at a holding potential of –60 mV in the whole-cell configuration) via electrical stimulation of the sciatic nerve (1.5× threshold at 0.033 Hz). EPSCs were classified as monosynaptic on the basis of stable latencies and an absence of failures in response to stimulation at 10 Hz. After observing a stable baseline of peak current amplitude, the selective GABABR agonist R(+)-baclofen hydrochloride (10 μm) was bath applied, and its effects on EPSC amplitude were subsequently measured and expressed as a percentage inhibition of the mean amplitude seen in the predrug trials. In these experiments, postsynaptic effects of GABABR activation were excluded by the use of a cesium-based electrode solution, which prevents current efflux through G-protein-coupled inwardly rectifying K+ channels (GIRKs), and the inclusion of the nonhydrolyzable GDP analog GDPβS in the electrode solution, which blocks the activation of G-proteins by the GABAB receptor. As demonstrated in Figure 7C, activation of presynaptic GABABRs abolishes primary afferent neurotransmission in the dorsal horn from the first days of postnatal life, because the mean EPSC amplitude was reduced by 93.4 ± 2.7% (n = 8) at P2, 96.0 ± 1.3% (n = 5) at P6–P7, and 94.3 ± 1.84% (n = 7) by P13–P14. An example of the block of primary afferent-evoked EPSCs by baclofen is illustrated in Figure 7A. The effects of baclofen on the evoked EPSCs were abolished by previous application of the GABABR antagonist CGP 55845 (at 2 μm), confirming the role of GABABRs in the observed response (Fig. 7B). Bath application of CGP 55845 (2 μm) alone did not significantly affect the amplitude of the evoked EPSCs (n = 3; data not shown), suggesting an absence of tonic activation of GABABRs on primary afferent terminals under our experimental conditions.

Potent modulation of primary afferent-evoked inputs to the superficial dorsal horn by the GABABR throughout the first 2 postnatal weeks. A, Glutamatergic inputs to dorsal horn cells were elicited via electrical stimulation of the sciatic nerve and classified as monosynaptic on observing a constant latency and absence of synaptic failures with 10 Hz stimulation. Example illustrates an EPSC in a P14 neuron evoked at a threshold (300 μA, 100 μsec) and latency (14.1 msec; approximate conduction velocity of 1.7 m/sec) suggestive of an input mediated by Aδ-fibers. Bath application of baclofen (10 μm) resulted in a reversible block of the evoked EPSCs. B, The effect of baclofen on EPSC amplitude was prevented by previous treatment with the selective GABABR antagonist CGP 55845 (2 μm). C, Baclofen abolishes primary afferent-evoked EPSCs in all age groups (expressed as the percentage of baseline EPSC amplitude inhibited by baclofen). D,Baclofen(Bac) significantly reduces the frequency, but not the amplitude, of mEPSCs in dorsal horn neurons, consistent with a presynaptic site of action (*p < 0.005; Wilcoxon matched-pairs test).
The lack of postsynaptic GABABR activation in these experiments was verified by measuring the effect of baclofen on mEPSCs recorded under the above conditions. In the presence of 500 nm TTX, baclofen significantly reduced the mEPSC frequency in a population of P2–P14 neurons from 0.45 ± 0.09 to 0.26 ± 0.06 Hz (n = 18; p < 0.005; Wilcoxon matched-pairs test). In contrast, Figure 7D demonstrates that mEPSC amplitude was not significantly altered by baclofen (TTX, 16.29 ± 2.13 pA; baclofen, 15.61 ± 2.06 pA; n = 17; p > 0.40; paired t test). The observed decrease in the frequency of mEPSCs in the presence of baclofen without a corresponding change in mEPSC amplitude strongly suggests a presynaptic site of action.
The effects of postsynaptic GABABR activation at different postnatal ages was investigated via bath application of baclofen (100 μm) to dorsal horn neurons (using a K+-based electrode solution with 0.3 mm Na3GTP substituted for GDPβS) and measuring the resulting change in holding current, which reflects the opening of GIRKs. An example of the slow outward current induced by baclofen (from Vh of –60 mV) is illustrated in Figure 8A. Abolition of the baclofen effect by CGP 55845 (2 μm) confirms that the outward current results from GABAB receptor activation (Fig. 8B). As illustrated in Figure 8C, the amplitude of the GABAB-mediated current depended on postnatal age, because the mean amplitude was significantly greater at P6–P7 (40.0 ± 8.7 pA; n = 8; p < 0.001; Kruskal–Wallis test) and P13–P14 (17.21 ± 1.55 pA; n = 11; p < 0.01) than at P0–P1 (4.39 ± 1.59 pA; n = 12). Although current amplitudes appeared to peak at P6–P7 and subsequently decrease during the second postnatal week, there was no statistically significant difference between the mean amplitudes observed at P6–P7 and P13–P14 (p > 0.05; Kruskal–Wallis test). Similar results were seen when current amplitudes were normalized to cell capacitance to obtain measurements of current density at different ages (data not shown).

Effects of postsynaptic GABABR activation in dorsal horn neurons vary according to postnatal age. A, Bath application of the selective GABABR agonist R(+)-baclofen hydrochloride elicits a slow outward current from a holding potential of –60 mV in neonatal dorsal horn cells. B, The effect of baclofen on the holding current was completely blocked by the selective GABABR antagonist CGP 55845 (2 μm). C, The amplitude of the baclofen-evoked outward current was significantly greater at P6–P7 and P13–P14 than at P0 (*p < 0.01; **p < 0.001; Kruskal–Wallis test).
Discussion
Sensory networks in the immature spinal cord are characterized by a dependence on GABAAR signaling, because GlyRs play an extremely limited role in spontaneous neurotransmission in the superficial dorsal horn during the first postnatal week (Figs. 2, 3). Functional dominance of GABAA receptors in developing spinal networks has been reported in motoneurons in which the frequency of GABAAR-mediated mIPSCs was fourfold higher than that of GlyR-mediated mIPSCs at embryonic day 17 (E17) to E18 (Gao et al., 2001). Dorsal horn neurons lacking GlyR-mediated sIPSCs responded to exogenous glycine applications, suggesting the possibility that functional GlyRs have been inserted into the membrane by birth but have not yet clustered at synaptic sites. Synaptic localization of GlyRs in spinal neurons depends on the expression of gephyrin, which binds with high affinity to both GlyRs and the tubulin comprising the subsynaptic cytoskeleton (Kirsch et al., 1993). Gephyrin is also involved in the synaptic targeting of GABAARs containing the γ2or α2 subunits (Essrich et al., 1998; Kneussel et al., 1999) but is not required for stabilization of other GABAAR subunits at inhibitory synapses (Kneussel et al., 2001). Interestingly, Dumoulin et al. (2000) report that GABAARs accumulate at postsynaptic sites before gephyrin at inhibitory synapses in cultures of embryonic spinal neurons and suggest that GABAARs may be involved in the recruitment of gephyrin to synaptic loci (Dumoulin et al., 2000). An absence or low level of gephyrin expression in the neonatal dorsal horn during the first postnatal week could explain the observed dependence of sIPSCs on GABAARs.
In the adult superficial dorsal horn, approximately half of GABAergic neurons also express glycine (Todd and Sullivan, 1990), and the glycine transporter GLYT2 colocalizes with both known isoforms of the GABA synthetic enzyme glutamic acid decarboxylase in synaptic boutons (Mackie et al., 2003). Recent evidence suggests that GABA and glycine can be copackaged and released from the same presynaptic vesicles, resulting in mixed GABAAR–GlyR mIPSCs (Jonas et al., 1998; Keller et al., 2001). The expression of glycine and GABA transporters in the dorsal horn during early postnatal development has yet to be characterized. Although a failure to transport glycine from the extracellular space or into synaptic vesicles could explain the paucity of spontaneous GlyR currents in the P0–P7 dorsal horn, it seems unlikely because glycine is expressed in the dorsal horn by E8 (Berki et al., 1995) and is carried by the same vesicular transporter (VIAAT, for vesicular inhibitory amino acid transporter) as GABA (Gasnier, 2000).
Developmental changes in mIPSC kinetics have profound implications for the net synaptic inhibition of immature spinal neurons. Keller et al. (2001) described the kinetics of GABAergic and glycinergic mIPSCs in laminas I–II neurons between the ages of P8 and P23. GABA mIPSCs exhibited slower decay kinetics than Gly mIPSCs at all ages, with smaller differences seen in older neurons attributable to a fourfold increase in the GABA mIPSC decay rate during this period (Keller et al., 2001). Unfortunately, the very low frequency of GABA mIPSCs and absence of Gly mIPSCs at P0–P1, even in the presence of ruthenium red, meant that we could not compare the kinetics of GABA and Gly mIPSCs at birth. Additional experiments under conditions of elevated neurotransmitter release would be necessary to acquire enough events to accurately analyze mIPSC kinetics during the first post-natal days.
In many regions of the immature CNS, GABAAR activation causes membrane depolarization that is sufficient to elicit APs, because elevated levels of intracellular [Cl–] yield an ECl that is more positive than both the resting potential and AP threshold (Leinekugel et al., 1997; Gao and van den Pol, 2001; Staley and Smith, 2001). In the developing hippocampus, GABAergic synapses form before glutamatergic inputs on CA1 pyramidal cells (Ben Ari et al., 1989) and interneurons (Hennou et al., 2002) and provide the main excitatory drive to the network as spontaneous activity was abolished by bicuculline (Ben Ari et al., 1989). A decrease in [Cl]i over the postnatal period resulting from the expression of the K+-Cl– cotransporter KCC2 can lead to a negative shift in ECl, thus causing the effects of GABA and glycine to become progressively inhibitory (Owens et al., 1996; Rohrbough and Spitzer, 1996; Ehrlich et al., 1999; Rivera et al., 1999). GABAergic activity may itself promote the switch from synaptic excitation to inhibition in hippocampal neurons, because chronic blockade of GABAA receptors reduced the levels of KCC2 mRNA and delayed the onset of inhibition (Ganguly et al., 2001).
It is unlikely that the reported hyperexcitability of dorsal horn neurons during the first postnatal week (Fitzgerald and Jennings, 1999) results from a dominant excitatory role for GABA during early postnatal development. First, the distinct electrophysiological properties of immature dorsal horn neurons persist after the maturation of the GABAAR response at the end of the first post-natal week. For example, at P6 A-fiber-induced sensitization of dorsal horn neurons is still widespread (33% of cells compared with 0% at P21), and receptive fields remain large, corresponding to 36 ± 2.9% of the total hindpaw area (Fitzgerald and Jennings, 1999), despite the exclusively hyperpolarizing action of GABA at this age (Fig. 5). More importantly, the present data show that EGABA is consistently below AP threshold at birth (Fig. 6). This suggests that GABA-evoked depolarizations likely function to inhibit the firing of newborn dorsal horn neurons by clamping Vm to ECl, thereby shunting subsequent EPSPs via an increase in membrane conductance and enhanced inactivation of voltagegated Na+ channels (Bormann et al., 1987). Previous work has shown that GABA depolarizes embryonic and early postnatal motoneurons but still blocks AP discharge, suggesting that GABAergic synaptic inhibition is functional by E18.5 in the ventral horn (Hubner et al., 2001; Marchetti et al., 2002), consistent with the widespread expression of KCC2 in the spinal cord by this age (Hubner et al., 2001). The heterogeneity of GABA responses observed in the present study (Fig. 5C) suggests considerable diversity in KCC2 levels among individual superficial dorsal horn neurons at birth. Overall, the present data suggest that, in contrast to the hippocampus, GABAARs do not provide the main excitatory drive to the network in the newborn dorsal horn. It should be noted that these results do not exclude a transient excitatory role for GABA in the developing dorsal horn. Indeed, single-channel recordings of GABA-activated Cl– channels in embryonic (E15) spinal cord neurons have reported an approximate EGABA of –30 mV (Serafini et al., 1995). Our results instead suggest that the switch from excitation to inhibition begins during the embryonic period and appears complete by the end of the first postnatal week.
The presence of depolarizing, inhibitory GABAAR potentials in the neonatal superficial dorsal horn may promote Ca2+ entry into neurons without allowing unchecked excitation during a period in which primary afferent C-fiber synaptic inputs are rapidly strengthening (Baccei et al., 2003) and descending inhibitory systems are not yet functional (Fitzgerald and Koltzenburg, 1986). Previous reports have demonstrated that GABAAR-evoked depolarizations increase [Ca2+]i via activation of N-type and L-type voltage-gated calcium channels in embryonic dorsal horn neurons (Reichling et al., 1994). These depolarizations may also contribute to the relief of the Mg2+ block of NMDA receptors (Khazipov et al., 1997), further increasing calcium influx into newborn dorsal horn neurons. Such elevations of intracellular calcium levels may be important for various developmental processes, including synapse formation and remodeling, as well as neuronal differentiation (Wang et al., 1994; Ben Ari, 2002). Thus, at birth, GABAAR activation may facilitate the maturation of the dorsal horn network despite preventing AP discharge in dorsal horn neurons.
Presynaptic modulation of glutamatergic transmission by GABABRs is observed at birth in CA1 hippocampal neurons, but postsynaptic GABABR-mediated inhibition is absent (Gaiarsa et al., 1995). The present results suggest that both presynaptic and postsynaptic GABABRs in the superficial dorsal horn are functional at birth but are regulated differently over the early postnatal period (Figs. 7, 8). Presynaptic inhibition of primary afferent-evoked EPSCs by baclofen has been reported previously in the neonatal dorsal horn (Blake et al., 1993), and the present study demonstrates that it remains potent throughout the first 2 post-natal weeks. Meanwhile, postsynaptic GABABR effects depended on postnatal age with maximal outward currents seen by P6–P7. Recent work has identified three subunits (GABABR1a, GABABR1b, and GABABR2) that heterodimerize to form functional GABABRs (White et al., 1998; Kuner et al., 1999) and are expressed in the DRG and superficial dorsal horn, with GABABR1a preferentially localized on presynaptic terminals and GABABR1b on cell bodies in the region (Towers et al., 2000). Developmental changes in the expression of GABABR1b (or GABABR2) subunits, or postnatal regulation of downstream GIRKs (Chen et al., 1997), could account for the age-related differences in postsynaptic GABABR effects. Additional experiments are necessary to determine whether the synaptic activation of presynaptic and postsynaptic GABABRs varies as a function of age.
Additional investigation of the synaptic and intrinsic properties of neonatal dorsal horn neurons will help elucidate possible mechanisms underlying the lower reflex thresholds and enlarged receptive fields that characterize the immature spinal cord. Clearly, the patterns of inhibitory activity in the superficial dorsal horn, essential to the sensory processing of painful stimuli, are modified during the early postnatal period.
Footnotes
This work was supported by Medical Research Council Grant 52378. We thank Dr. D. Billups for advice during the project, Drs. R. Bardoni, M. Baker, and J. Wood for the kind loan of equipment, and Dr. D. Attwell for helpful comments on the preparation of this manuscript.
Correspondence should be addressed to Mark Baccei, Department of Anatomy and Developmental Biology, University College London, Gower Street, London WC1E 6BT, UK. E-mail: m.baccei{at}ucl.ac.uk.
DOI:10.1523/JNEUROSCI.5211-03.2004
Copyright © 2004 Society for Neuroscience 0270-6474/04/244749-09$15.00/0





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}