

Abstract
Temperature transduction in mammals is possible because of the presence of a set of temperature-dependent transient receptor potential (TRP) channels in dorsal root ganglia neurons and skin cells. Six thermo-TRP channels, all characterized by their unusually high temperature sensitivity (Q10 > 10), have been cloned: TRPV1–4 are heat activated, whereas TRPM8 and TRPA1 are activated by cold. Because of the lack of structural information, the molecular basis for regulation by temperature remains unknown. In this study, we assessed the role of the C-terminal domain of thermo-TRPs and its involvement in thermal activation by using chimeras between the heat receptor TRPV1 and the cold receptor TRPM8, in which the entire C-terminal domain was switched. Here, we demonstrate that the C-terminal domain is modular and confers the channel phenotype regarding temperature sensitivity, channel gating kinetics, and PIP2 (phosphatidylinositol-4,5-bisphophate) modulation. Thus, thermo-TRP channels contain an interchangeable specific region, different from the voltage sensor, which allows them to sense temperature stimuli.
Introduction
Transient receptor potential (TRP) ion channels are one of the largest groups of ion channels comprising six protein families (Montell et al., 2002; Clapham, 2003; Moran et al., 2004). A subset of these channels, the thermo-TRPs, is activated by distinct physiological temperatures. Six thermo-TRP channels, all characterized by their unusually high temperature sensitivity (Q10 > 10), have been cloned: TRPV1–4 are heat activated (Caterina et al., 1997, 1999; Peier et al., 2002b; Xu et al., 2002), whereas TRPM8 (McKemy et al., 2002; Peier et al., 2002a) and TRPA1 (Story et al., 2003) are activated by cold. With a Q10 of ∼26 for TRPV1 (Liu et al., 2003) and ∼24 for TRPM8 (Brauchi et al., 2004), they far surpass the temperature dependence of the gating processes characterized by other ion channels (Q10, ∼3) (Hille, 2001). Because of the lack of structural information, the molecular basis for regulation by temperature remains unknown.
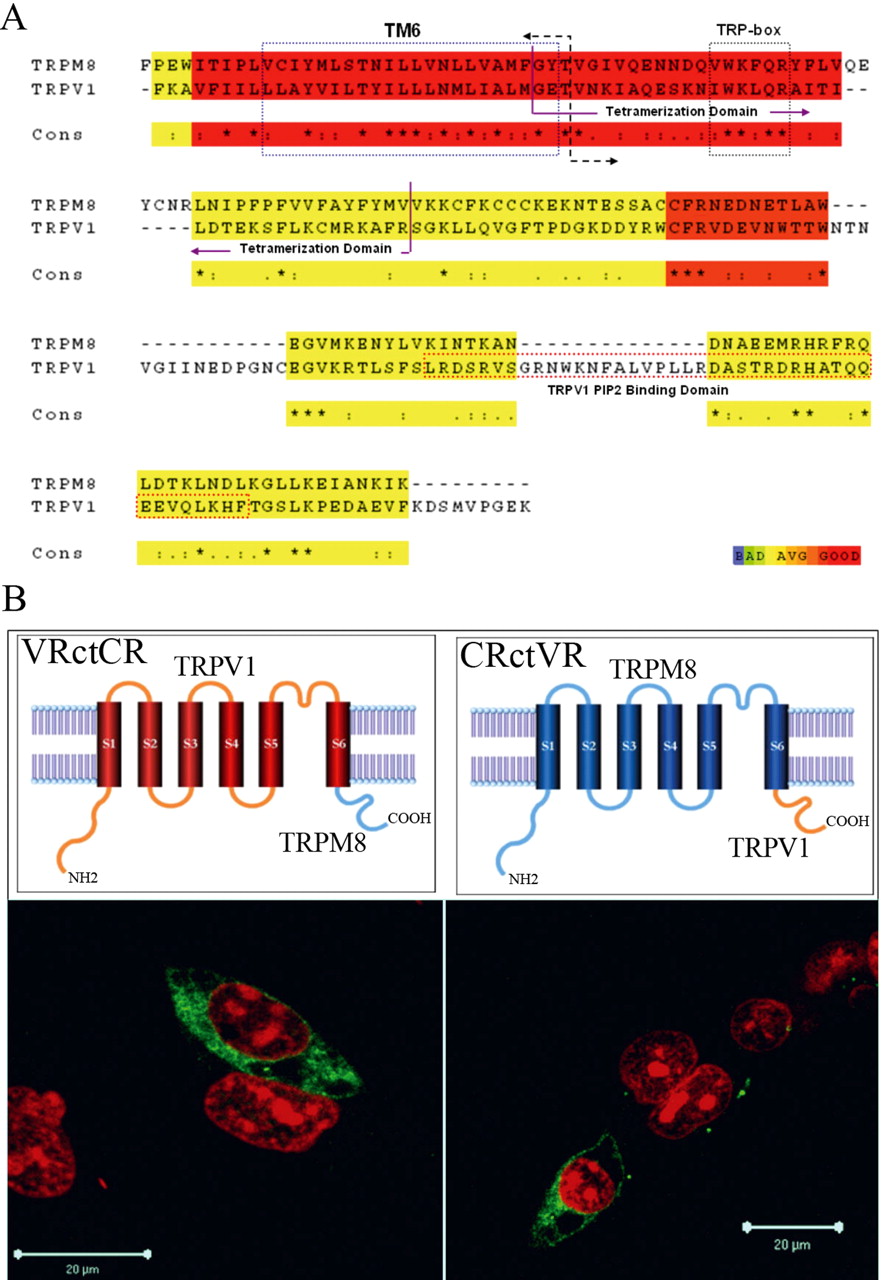
C-terminal domains of TRPV1 and TRPM8 are structures that are critical for proper channel function. For example, the TRPV1 C terminal contains an inhibitory phosphatidylinositol-4,5-biphosphate (PIP2) binding site (Prescott and Julius, 2003) and a tetramerization domain in the proximal C-terminal region (Garcia-Sanz et al., 2004) (Fig. 1 A). Partial deletions performed in the C-terminal domain of TRPV1 result in functional channels with attenuated heat sensitivity, and truncation of the whole TRPV1 C-terminal domain completely hindered channel expression (Vlachova et al., 2003; Liu et al., 2004). In TRPM8 channels, binding of PIP2 leads to channel activation (Liu and Qin, 2005; Rohacs et al., 2005). The proximal C-terminal TRP domain is conserved in TRPM8 and appears to serve as a PIP2 site (Rohacs et al., 2005). These results underscore the importance of the C-terminal domain of TRPV1 and TRPM8 as a structural domain able to modulate thermo-TRP channel activity.

Design and expression of thermo-TRP C-terminal chimeras. A, Sequence alignments between rTRPV1 and rTRPM8. The cut–paste limit for chimera construction is marked by a double arrow. Several important features are highlighted in the alignment: the TM6 segment, TRPbox, and TRPV1 PIP2 binding domain. B, Chimeras and their expression in HEK-293 cells. Each of the top schemes corresponds to the immunochemistry below (scale bar, 20 μm). It is clear that the chimeric channels are able to reach the membrane, but large amounts of them are retained on intracellular compartments.
We performed multiple alignments of the different C terminals of thermo-TRPs. These alignments showed that the tetramerization domain is conserved between thermo-TRPs. Secondary structure analysis revealed the existence of three α helices in the region near S6, which were also found to be conserved among thermo-TRPs (supplemental material, available at www.jneurosci.org). In contrast, almost all similarities among the C terminals of these channels are lost in the distal region. These previous observations, and the fact that the key question regarding what makes thermo-TRPs temperature sensitive remained unanswered, prompted us to build C-terminal chimeras between TRPV1 and TRPM8.
Materials and Methods
Molecular biology.
cDNAs coding for rat TRPV1 (GenBank accession number NM_031982) and rat TRPM8 (kindly provided by Dr. David Julius; GenBank accession number NM_134371) were used. The boundaries of the transmembrane domains of both channels were defined by consensus using multiple transmembrane prediction tools (supplemental material, available at www.jneurosci.org). Chimeric thermo-TRP channels were made by the overlapping extension method and confirmed by DNA sequencing. DNAs were subcloned either into pCDNA3 or pTracer-CMV2 vectors using suitable enzymes.
Cell culture and transfection.
Human embryonic kidney 293 (HEK-293) cells were transfected either with pCDNA3 or pTracerCMV2 vectors containing wild-type or chimeric coding DNA sequence. Transfection was performed using cationic liposomes (TransIT-HEK293; Mirus, Madison, WI). Transfection efficiency, as determined using the pTracer vector, was 30%.
HEK-293 electrophysiology.
Whole-cell currents were measured ∼30–40 h after transfection of HEK-293 cells. In general, we found that chimeric proteins express currents ∼5–10 times smaller than those obtained with the wild-type channels. Gigaseals were formed using 2–4 MΩ borosilicate pipettes (outer diameter, 1.5 mm; inner diameter, 0.86 mm; Warner Instruments, Hamden, CT). Whole-cell voltage clamp was performed at different temperatures (10−40°C). Macroscopic currents were acquired at 100 kHz and filtered at 10 kHz. Temperature ramps were performed, and the whole-cell current was acquired during the ramp; cells were maintained at a holding potential of +80 mV. Normalized conductance (G/Gmax) was obtained from the steady-state current (Isteady state/applied voltage) and from the tail current when possible. Activation and deactivation rates were calculated from the slow component of a double exponential fit. Activation rates in both cases were calculated from activation time constants of current traces elicited at +150 mV. Deactivation rates were calculated from deactivation time constants of current traces elicited at −40 mV for the case of VRctCR and −70 mV for the case of CRctVR. EPC7 patch-clamp amplifier (HEKA Elektronik, Lambrecht/Pfalz, Germany) and 6052E acquisition board (National Instruments, Austin, TX) were used.
Recording solutions, Calcium imaging, Immunocytochemistry, Temperature control and ramps, and Data analysis can be found in the supplemental material (available at www.jneurosci.org).
Results
We created chimeric channels between TRPV1 and TRPM8 in which the complete C-terminal domain was exchanged (Fig. 1A,B). The chimeras were dubbed CRctVR [C terminal of vanilloid receptor (TRPV1) attached to cold receptor core] and VRctCR [C terminal of cold receptor (TRPM8) attached to vanilloid receptor core].
C-terminal region of TMPM8 and TRPV1 constitute exchangeable regulatory modules
To check the presence of the chimeric channels in a subpopulation of transfected HEK cells, we performed immunocytochemistry experiments (Fig. 1B), and functional expression was determined using calcium imaging. Calcium changes induced by temperature modifications were detected in transfected cells loaded with Fluo-4. Approximately 30% of the cells exhibited fluorescent responses to temperature changes, a value in agreement with the transfection efficiency as determined with the pTracer vector. Steady-state measurements of internal Ca2+ indicate an increase in fluorescence (approximately threefold) when temperature was decreased (40→15°C) in the case of the VRctCR or when the temperature was raised (16→40°C) in the case of the CRctVR chimera. Nontransfected cells do not present significant changes in fluorescence at different temperatures. The baseline calcium levels were essentially the same when comparing cells expressing the chimeras or cells expressing wild-type channels. These values were, however, somewhat higher than nontransfected cells. This set of experiments demonstrates that both chimeric channels are permeable to calcium. After checking the functional expression of both chimeras, we measured whole-cell currents during temperature ramps. These experiments show that VRctCR and CRctVR are activated by cold and heat, respectively (Fig. 2A,B). The analysis of the current versus temperature plots gives a Q10 value of 7 ± 1 for VRctCR and 11 ± 1 for CRctVR. Although highly temperature sensitive, the Q10 values of the chimeric channels are about three times smaller than observed in wild-type channels. This suggests that the functional coupling between temperature and channel opening is maintained but with a lower efficiency in our chimeric channels. The overall behavior in terms of reliability of the chimeras was analyzed through a statistical analysis that comprises 51 experiments (supplemental material, available at www.jneurosci.org). CRctVR behaves inversely in 95% of trials, whereas VRctCR responds inversely in 74% of trials. Moreover, both chimeras have the tendency to lock themselves in one state, either activated or deactivated after stimulation, ∼20% of the time (supplemental material, available at www.jneurosci.org). These observations support the idea that the lower efficiencies (lower Q10 values) are attributable to the loss of some specific interactions present in wild-type channels.

Temperature sensitivity of chimeric channels. A, VRctCR chimera normalized current as a function of temperature. B, CRctVR chimera normalized current as a function of temperature. Data obtained using slow temperature ramps (0.25°C/s) and a holding potential of 80 mV. The solid lines are the best fits to a Boltzmann function. Each point represents an average of at least four different experiments. Error bars indicate SD.
To better characterize the effects of the C-terminal exchange between TRPV1 and TRPM8, we studied the voltage activation at fixed temperatures (Fig. 3). Figure 3, C and D, shows macroscopic current traces obtained from the same whole-cell patch at different temperatures for each chimera, showing the inverse gating behavior and a change in kinetic properties when compared with the wild-type channels (Brauchi et al., 2004; Voets et al., 2004). For both chimeras, at all of the temperatures tested, there was a strong outward rectification of the steady-state current. Figure 3, E and F, shows the normalized conductance–voltage curves for the VRctCR (n = 9) and CRctVR (n = 14) chimeric channels. These plots are well fitted by a Boltzmann function of the form G = Gmax/[1 + exp−(zF(V − V0.5)/RT], where z is the voltage dependency, V0.5 is the half-activation voltage, and Gmax is the maximum conductance (F, R, and T have their usual meanings). The voltage dependence of the parental ion channels are maintained in the chimeras, and the observed z values are in the range of 0.55–0.80, very similar to the ones reported previously for TRPV1 and TRPM8 (Brauchi et al., 2004; Voets et al., 2004) (Fig. 3A,B). Cooling induced a leftward shift of the voltage activation curve of the VRctCR chimera; the V0.5 decreases by ∼100 mV on cooling from 35 to 15°C (Fig. 3E). This shift in the voltage dependence of activation agrees reasonably well with the left shift of the voltage activation curve of the TRPM8 channel induced by cooling (Brauchi et al., 2004; Voets et al., 2004) (Fig. 3A). A leftward shift is also observed when heating in the case of the CRctVR chimera, and V0.5 decreases ∼75 mV when the temperature is increased from 15 to 35°C (Fig. 3F). Thus, the behavior of the CRctVR is similar to that of the TRPV1 channel (Fig. 3B).

Electrophysiological analysis of the chimeras. A, B, Plots showing the normalized conductance (for details, see Materials and Methods) in function of voltage at the indicated temperatures for TRPM8 and TRPV1, respectively. The solid lines correspond to the best fit to Boltzmann functions. C, Whole-cell recordings of a cell expressing the VRctCR chimera and exposed to the indicated temperatures. The voltage protocol is shown in the top of the figure. D, Whole-cell recordings of a cell expressing the CRctVR chimera and exposed to the indicated temperatures. E, F, Plots showing the normalized conductance in function of voltage at the indicated temperatures for VRctCR and CRctVR, respectively. The solid lines correspond to the best fit to Boltzmann functions. G, H, Arrhenius plots for activation and deactivation process of VRctCR and CRctVR chimeras, respectively (each data point represent at least three experiments). Activation (α) and deactivation (β) rates were calculated from double exponential function fits (see Materials and Methods). Activation energy was calculated directly from the slope of the Arrhenius plots (slope = −Ea/R). Error bars indicate SD.
Closer inspection of the records shows that the activation rate (α) of the VRctCR chimera has a shallow temperature dependence and that the closing rate (β) is steeply temperature dependent (Fig. 3G). This is a characteristic of the TRPM8 channel as reported by Voets et al. (2004) and Brauchi et al. (2004). On the other hand, analysis of the macroscopic current records of CRctVR chimera indicates that the temperature dependence resides mainly in the activation rate (Fig. 3H), which is a characteristic of the TRPV1 channel (Voets et al., 2004). This is an important result, because it allows us to conclude that the major determinants of TRPV1 and TRPM8 kinetic behavior are associated with the C-terminal domain. Thus, the functional coupling is maintained and the overall phenotype is switched.
PIP2 is able to inversely regulate the chimeras
PIP2 has been shown to be an important modulator molecule for thermo-TRP channels, and there are solid data supporting the idea that there is a PIP2 binding site and/or specific interaction between PIP2 and amino acids present in the C-terminal domain. The PIP2 site in the case of TRPM8 is located on the very proximal C terminus (Rohacs et al., 2005), whereas in TRPV1 it is located on the distal C-terminal domain (Prescott and Julius, 2003; Liu and Qin, 2005). Individual analysis of our chimeras revealed that VRctCR, which has the C terminal of TRPM8, loses its current almost completely after 10 or 15 min in a way that was independent of the applied voltage protocol, suggesting an inherited PIP2 control mechanism. Another characteristic of VRctCR is that the steady-state currents reach a maximum and saturate (macroscopic current showed in Fig. 3C). This saturation takes place at potentials >120 mV. This behavior is not present when channel activity is measured in cell attached patches or when PIP2 (10 μm) is added to the patch pipette (supplemental Fig. S5, available at www.jneurosci.org as supplemental material). Furthermore, we observed that the C-terminal domain carries the phenotype for PIP2-related responses. Responses from VRctCR chimera are potentiated by the presence of PIP2 (10 μm) in the patch pipette (Fig. 4A,C). As predicted from previous reports (Prescott and Julius, 2003), we observed an inhibitory response against PIP2 (10 μm) in the CRctVR chimera (Fig. 4B,C).

Pharmacological properties of the chimeras. Current records obtained at 100 mV. Temperature, 25°C. A, PIP2 (10 μm) activates the VRctCR channel. B, PIP2 (10 μm) inhibits the CRctVR channel. C, Bar plot of several experiments exposing the CRctVR and VRctCR channels to PIP2 (10 μm). Unpaired t test showed that both chimeras were affected by PIP2 addition. D, Current traces showing menthol-induced activation and La3+ blocking of CRctVR chimera. F, Current traces for capsaicin-induced activation and La3+ blocking of VRctCR chimera. E, G, A summary of the experiments performed for the indicated chimera. La3+ was added to check whether the chimeras were able to be blocked (the addition was always performed after a maximal activation of the respective channel). Capsaicin (Cap) (200 μm) and menthol (Men) (500 μm) were added to CRctVR and VRctCR, respectively, as control experiments. One-way ANOVA and Bonferroni’s statistical analysis were performed for the two data sets in E and G, and in both cases the populations were found to be statistically different (p < 0.05). H, Capsaicin dose–response curves for TRPV1 (IC50, 0.6 μm) and VRctCR (IC50, 7.8 μm). I, Menthol dose–response curves for TRPM8 (IC50, 22 μm) and CRctVR (IC50, 28 μm). Current values were normalized and expressed as percentage of maximal response to the agonist. Each point represents mean values ± SD from at least three different patches. The Hill equation was used to fit the data.
C terminus is not essential for capsaicin or menthol responsiveness
A comparison between mammalian and avian (heat-sensitive but capsaicin-insensitive) TRPV1 channels resulted in a map of the structural determinants for capsaicin interaction. Results revealed that residues located at the transition between the second intracellular loop and the third transmembrane domain (Jordt and Julius, 2002) are required for capsaicin binding. The same approach revealed that the amino acid residues essential for the binding of icilin, a super-cooling agent, are also located in the intracellular loop connecting transmembrane domains 2 and 3 (Chuang et al., 2004). The menthol binding site has not yet been identified. Taking into account these observations, we tested the VRctCR and CRctVR chimeras for their ability to be activated by capsaicin and menthol, respectively (Fig. 4D,F). We found that 1 mm menthol and 50 μm capsaicin were able to strongly activate CRctVR and VRctCR, respectively (Fig. 4E,G). Full dose–response curves for these two compounds show that the VRctCR chimera is ∼10-fold less sensitive to capsaicin than the TRPV1 channel (Fig. 4H) and that the CRctVR chimera has the same sensitivity to menthol as the TRPM8 channel (Fig. 4I). For the TRPV1 and the TRPM8 channels, the IC50 values for capsaicin and menthol were 0.6 and 22 μm, respectively (Fig. 4H,I). These values are in reasonable agreement with those reported previously by Caterina et al. (1997) (capsaicin IC50, 1 μm) and by Chuang et al. (2004) (menthol IC50, 50 μm). These results indicate, as previously proposed, that the transmembrane core of TRPV1 carries the response element to capsaicin and suggest that the region for menthol interaction in TRPM8 is located outside of the C-terminal domain. Moreover, we found that the VRctCR chimera is insensitive to menthol and that the CRctVR chimeric channel is not activated by capsaicin (Fig. 4E,G).
Discussion
A regulatory role for the C terminus of thermo-TRPs has been proposed before, in terms of its participation in the effect of PIP2 and temperature on channel behavior (Prescott and Julius, 2003; Vlachova et al., 2003; Liu et al., 2004; Liu and Qin, 2005; Rohacs et al., 2005). Deletions in the distal portion of the C-terminal domain of TRPV1 have severe repercussions on channel activity (Vlachova et al., 2003). TRPV1 channel expression is affected by deletions in the TRP domain, located in the proximal C-terminal sequence (near TM6) (Fig. 1A), likely because of the removal of a putative tetramerization domain (Garcia-Sanz et al., 2004).
The relevant contribution of this study is the finding that, for both TRPV1 and TRPM8, modulation of channel gating behavior by temperature arises from the C-terminal structure that follows the S6 inner helix. A notable aspect of these results is the exchangeable modularity not only of the temperature-sensitive elements but also of the PIP2 regulatory elements. Thus, most of the key structural elements of the ligand regulatory process, including binding and coupling machinery that converts the free energy of ligand binding into mechanical work (pore opening) are intrinsic to the C terminal.
We have proposed a mechanism to describe the process of opening of thermo-TRP channels (Brauchi et al., 2004). In this model, both voltage-dependent and temperature-dependent activation of the channel involve an allosteric mechanism that demands the existence of separate structures able to sense specific stimuli and, in doing so, transfer the electrical or thermal energy to the gate. The modularity of the C-terminal structure suggests that most of key elements necessary for the main function of thermo-TRP channels (to sense temperature) are self-contained in this cytoplasmic region.
TRP channels form tetramers of identical subunits (Kadei et al., 2001); just as voltage-gated K+ channels (Clapham, 2003). The voltage dependence in these channels appears to be intrinsic to the channel-forming protein. The voltage sensor of these channels is, however, unknown, and a closer inspection of the predicted S4 segment of these channels reveals the presence of only one basic residue in TRPV1 and three in TRPM8. It is possible that the weak voltage dependence showed by thermo-TRPs is attributable to the scarcity of positive charges in the S4 domain.
C-terminal role in thermo-TRP channels
The observations presented here led us to the conclusion that the C terminal has an essential role not only as a regulatory domain involved in determining the so-called “activation thresholds” but also as a thermal modulator for the channel aperture. In support of this, we have clearly shown that the C terminal is able to define the phenotype of activation. The fact that temperature dependency is transposable constitutes a remarkable aspect of the results presented here, because they clearly show that the coupling between temperature stimuli and channel gating, although weakened, is maintained in the chimeras.
Channels are most likely built as modular structures (Schreiber et al., 1999; Jiang et al., 2001; Choe, 2002; Zagotta et al., 2003; Long et al., 2005; Murata et al., 2005), and thus the interactions between different modules (e.g., pore, binding sites, sensors, etc.) are defined by mechanic and electrostatic interactions that could be represented or understood as coupling factors and/or allosteric constants. Our data indicate that the modular nature of the structures involved in activation processes allow different stimuli (voltage, temperature, and agonists) to promote thermo-TRP channel opening by different interrelated mechanisms as has been suggested in the form of allosteric interaction (Brauchi et al., 2004; Rohacs et al., 2005).
To explain our results, we suggest three possible scenarios: One possibility is that the temperature affects the interaction of the C terminus with PIP2. However, although TRPV1 has a PIP2 binding site, its presence is not necessary to maintain the temperature dependence (Prescott and Julius, 2003; Liu et al., 2004). A general mechanism of PIP2 activation on TRP channels has been proposed by Rohacs et al. (2005). They have hypothesized that the PIP2 activation site found in TRPM8 is present is most TRP channels, including TRPV1, because the TRP domain is highly conserved. If this PIP2 site were involved in temperature sensing, all TRPs channels would present some degree of temperature dependence, which is not the case. Another possibility could be that temperature affects the interaction between a particular portion of the proximal C-terminal and some other region of the channel, probably an intracellular loop. Finally, it may be that independent arrangements induced by temperature on C-terminal domains directly promote gate opening.
It is possible that channel opening is controlled by a mechanism similar to the one observed for HCN channels (Zagotta et al., 2003), considering then that a direct coupling between the S6 and the C-linker may exist in thermo-TRP channels. This is based on the structural similarities between the C terminals of HCN and TRPV1 channel. Garcia-Sanz et al. (2004) found that comparison between the predicted secondary structure of TRPV1 C-terminal domain and the C terminal of HCN channels reveals significant similarities. Moreover, the C-linker that connects the cAMP-binding site with the pore in the case of HCN channels and the TRP domain in TRPV1 functions as tetramerization domains. In HCN channels, it has been proposed that the structural rearrangement produced by cAMP leads to a change in tension on the helical linker connecting the C-terminal domains with S6 segment. This tension on the linker provides the energy necessary to move the S6 inner helix to the open conformation. Similar structural rearrangements may be induced by temperature in thermo-TRP channels.
In summary, our results show that the TRPV1 C terminal confers heat-dependent regulation on the TRPM8 pore module, whereas the TRPM8 cytosolic module confers cold-dependent regulation on the TRPV1 pore module. The main conclusion of our work is that thermo-TRP channels have a modular C-terminal domain that is able to determine channel phenotype regarding temperature sensitivity.
Footnotes
-
This work was supported by funds from Fondo Nacional de Investigacion Cientifica y Tecnologica 103-0830. Centro de Estudios Cientificos is funded in part by grants from Fundación Andes, the Tinker Foundation, and Howard Hughes Medical Institute, and hosts a Millennium Science Institute (Ministerio de Planificación, Government of Chile). We thank M. A. Castro for confocal microscopy technical assistance and discussion of results, C. Feinn for his contribution to this manuscript, O. Alvarez and R. A. Jorquera for discussion of results and figure preparation, and C. Zaelzer for help with chimeric constructions.
- Correspondence should be addressed to Dr. Ramon Latorre, Laboratory of Biophysics and Molecular Physiology, Centro de Estudios Científicos, Avenida Arturo Prat 514, Valdivia, Chile 905-9100. Email: rlatorre{at}cecs.cl





{kind=link}
{kind=link}
{kind=link}
{kind=link}