

Abstract
The extracellular signal-regulated kinases (ERKs) 1/2 pathway is stimulated by drugs of abuse in striatal neurons through coincident activation of dopamine D1 and glutamate NMDA receptors and is critical for long-lasting behavioral effects of these drugs. Although regulation of transcription is a major target of ERK, the precise mechanisms by which it contributes to behavioral alterations is not known. We examined the role of Zif268, an immediate-early gene induced by drugs of abuse under the control of ERK, in behavioral responses to cocaine using knock-in mutant mice in which Zif268 was replaced by LacZ. No biochemical or behavioral differences between mutant and wild-type mice were observed in basal conditions or in acute responses to cocaine injection. In contrast, locomotor sensitization to single or repeated cocaine injections was dramatically diminished in both heterozygous and homozygous Zif268 mutant mice. Conditioned place preference in response to cocaine was prevented in Zif268-deficient mice. This effect was not attributable to a general learning deficit because the mutant mice displayed normal conditioned place preference when food was used as reward. Our results provide direct genetic evidence for the requirement of Zif268 for long-lasting association of environmental context with specific behavioral responses after short exposures to cocaine. They also underline the common molecular machinery involved in long-lasting drug-induced behavioral alterations and the formation of other types of memory.
Introduction
Integration of dopamine (DA)- and glutamate-coded signals is thought to underlie long-term plasticity and reward-related learning in corticostriatal networks (Berke and Hyman, 2000; Hyman and Malenka, 2001; Reynolds and Wickens, 2002; Kelley, 2004). This integration plays a central role in the long-lasting effects of drugs of abuse, which have the common property to stimulate DA transmission in the nucleus accumbens (NAcc) (Di Chiara, 1999). Drugs of abuse are thought to divert the normal role of DA neurons in coding reward prediction errors and regulating synaptic plasticity at corticostriatal synapses (Di Chiara, 1999; Everitt and Wolf, 2002; Schultz, 2002; Nestler, 2004).
An important signaling pathway by which drugs of abuse exert their long-lasting effects involves extracellular signal-regulated kinase (ERK) (Valjent et al., 2000, 2005). [The ERK family characterized by a Thr–Glu–Tyr motif in the activation loop includes several isoforms (ERK1–ERK8). Only ERK1 and ERK2 have been studied in the action of cocaine, and ERK2 appears to be the major isoform involved.] Activation of ERK in the NAcc is a common and specific effect of drugs of abuse (Valjent et al., 2004), which requires the coincident activation of DA D1 and glutamate NMDA receptors, providing a basis for the integration of the signals generated by mesostriatal and corticostriatal pathways (Valjent et al., 2000, 2005). The events downstream from ERK activation are poorly characterized. Substrates of ERK include transcription factors, which control the expression of immediate-early genes (IEGs), many of which are themselves transcription factors (Hope et al., 1992; Moratalla et al., 1992, 1996; Kano et al., 1995). This induction depends on activation of both D1 and NMDA receptors (Young et al., 1991; Wang et al., 1994; Konradi et al., 1996). However, the precise contribution of the various IEGs to the behavioral effects of cocaine remains unknown.
ERK activates mitogen- and stress-activated protein kinase-1 (MSK1) in the striatum. In the absence of MSK1, phosphorylation of cAMP-regulated element binding protein (CREB) and induction of c-fos and preprodynorphin are virtually absent (Brami-Cherrier et al., 2005). In contrast, the induction of Zif268 (also termed NGFI-A, Egr-1, or Krox24), an IEG induced by cocaine or cocaine-associated clues (Bhat et al., 1992; Hope et al., 1992; Moratalla et al., 1992; Thomas et al., 2003), is not altered (Brami-Cherrier et al., 2005). Interestingly, locomotor sensitization in response to repeated injections of cocaine was significantly reduced but not completely absent in MSK1 mutant mice, whereas the rewarding effects of cocaine were increased. The maintenance of these cocaine-induced behavioral effects suggested that Zif268 could be implicated in their development. To test this hypothesis, we explored cocaine-induced locomotor sensitization and cocaine-rewarding effects in Zif268 mutant mice (Topilko et al., 1998). Our results show that Zif268 is a target of ERK, essential for long-lasting behavioral effects of cocaine.
Materials and Methods
Animals and treatments
Mice were generated as described previously (Topilko et al., 1998) and backcrossed onto a C57BL/6N background (Supplementary Material, available at www.jneurosci.org as supplemental material). Experiments were performed in accordance with the guidelines of the French Agriculture and Forestry Ministry for handling animals (Decree 87849, License 01499) in 2- to 10-month-old male and female mice of the three genotypes. Cocaine-HCl (Sigma, St. Quentin Fallavier, France) was dissolved in 0.9% (wt/vol) NaCl solution (saline) and administrated by intraperitoneal injection.
Protein analysis and immunohistochemistry
Procedures were as described previously (Valjent et al., 2000, 2005). For details, see Supplementary Material (available at www.jneurosci.org as supplemental material).
Behavioral analysis
Locomotor activity and sensitization.
Locomotor activity was measured in a circular corridor with four infrared beams placed at every 90° (Imetronic, Pessac, France) in a low luminosity environment. Counts were incremented by consecutive interruption of two adjacent beams (i.e., mice moving through one-quarter of the circular corridor). Locomotor sensitization induced by a single or repeated cocaine injection was studied as described previously (Brami-Cherrier et al., 2005; Valjent et al., 2005).
Conditioned place preference.
The place preference apparatus (Imetronic, Pessac, France) comprised two different compartments distinguished by different patterns on floors and walls separated by a central neutral area. The conditioned place preference (CPP) protocol was as follows. (1) In the preconditioning test (day 1), mice were placed in the central neutral area, and the time spent in each compartment was measured during 18 min. (2) For conditioning (days 2–7), after injection of cocaine (10 mg/kg, days 2, 4, and 6) or saline (days 3, 5, and 7), mice were alternatively confined in each compartment during 20 min. Control mice received saline every day. (3) In the postconditioning test (day 8), mice had free access to both compartments, and the time spent in each compartment was measured as in the preconditioning test. Place preference was evaluated for each mouse as the difference between the times spent in the drug- and saline-paired compartments.
CPP induced by food reward was tested in mice that had limited access to food during the time of test and the 5 d before (90% of their normal weight). The protocol was similar to that described above for cocaine, except that, for conditioning, mice were alternatively confined in one compartment with food (4 g of normal mouse chow plus sucrose; days 1, 3, and 5) and in the other compartment without food (days 2, 4, and 6) (Maldonado et al., 1997). Control mice had no access to food in either compartment on days 1–6. Place preference was calculated as above.
Results
Induction of Zif268 by cocaine is mediated by the ERK pathway
Although ERK activation mediates Zif268 induction in many models (Davis et al., 2000; Valjent et al., 2001; Derkinderen et al., 2003; Radwanska et al., 2005), its implication in cocaine-induced transcription of Zif268 in the striatum has not been demonstrated. In agreement with previous reports (Bhat et al., 1992; Hope et al., 1992; Moratalla et al., 1992), injection of cocaine (20 mg/kg) increased the number of Zif268-positive cells in the dorsal striatum (DStr) (272 ± 33% of control) and the NAcc (194 ± 12% of control) (supplemental Fig. 1, available at www.jneurosci.org as supplemental material). When cocaine was preceded by an injection of SL327 (α-[amino[(4-aminophenyl)thio]methylene]-2-(trifluoromethyl)benzeneacetonitrile), a drug that prevents ERK activation in vivo by blocking the upstream kinase MEK (mitogen-activated protein kinase/ERK kinase) (Valjent et al., 2000), the induction of Zif368 was completely abolished (DStr, 75 ± 11% of control; NAcc, 69 ± 8% of control) (supplemental Fig. 1, available at www.jneurosci.org as supplemental material), supporting the role of ERK in the control of Zif268 by cocaine in the striatum.
Signaling cascades triggered by cocaine are normal in Zif268-deficient mice
To test the role of Zif268, we used knock-in mutant mice in which the Zif268 gene was inactivated by the insertion of a LacZ-Neo cassette (Topilko et al., 1998). Morphological analyses and measurement of multiple markers (tyrosine hydroxylase, synapsin I/II, glutamate receptor subtype GluR1/2, NMDA receptor NR2B, postsynaptic density-95, Gαs, Gαolf, protein-phosphatase-1, dopamine- and cAMP-regulated phosphoprotein-32, regulator of calmodulin signaling, striatal-enriched tyrosine phosphatase-46/61, ERK1/2, and CREB) in the DStr and prefrontal cortex did not reveal any alteration in Zif268−/− mice (supplemental Table 1, available at www.jneurosci.org as supplemental material). To check whether the absence of Zif268 altered the signaling pathways upstream from its induction, we took advantage of the LacZ reporter gene inserted in place of Zif268. Cocaine increased β-galactosidase immunostaining in the DStr and NAcc shell in Zif268−/− mice (Fig. 1a). Moreover, the induction by cocaine of c-fos, another ERK-controlled IEG, was comparable in Zif268+/+ and Zif268−/− mice (Fig. 1b). These results show the preservation of signaling cascades triggered by cocaine in Zif268−/− mice.

Normal gene induction by cocaine in Zif268−/− mice. a, β-Galactosidase immunostaining in knock-in mutant mice (Zif268−/−) in which LacZ replaces Zif268 90 min after acute administration of saline or cocaine (20 mg/kg). No immunostaining was detected in wild-type littermates (Zif268+/+). Scale bars, 40 μm. b, c-fos induction was quantified in the DStr and in the shell of the NAcc in the same conditions in Zif268−/− and Zif268+/+ mice. Data (means ± SEM; n = 3 per group) were analyzed using two-way ANOVA: effect of treatment, DStr, F(1,8) = 38.55, p < 0.01; shell, F(1,8) = 82.38, p < 0.01; effect of genotype, DStr, F(1,8) = 0.171, not significant (NS); shell, F(1,8) = 2.054, NS. Post hoc comparison (Bonferroni's test), ∗∗p < 0.01 saline versus cocaine.
Single and repeated injections of cocaine are known to induce different molecular responses. For example, induction of Zif268 in the NAcc is known to desensitize after repeated injections of cocaine (Bhat et al., 1992; Hope et al., 1992). We used the LacZ reporter gene to examine this point in our model. In agreement with previous studies, β-galactosidase induction diminished in the NAcc of Zif268+/− mice after repeated cocaine administration and a 7 d withdrawal period (Fig. 2). In contrast, no persistent desensitization was found in the DStr, indicating different mechanisms of regulation in these two striatal regions.

Effects of chronic cocaine treatment on the induction of β-galactosidase in the dorsal striatum and nucleus accumbens shell of Zif268+/− mice. The induction of β-galactosidase was tested in Zif268+/− mice in which an allele of Zif268 was replaced by LacZ. Naive mice (Naive) received an acute injection of saline (Sal) or cocaine (20 mg/kg; Coc), as indicated. Pretreated mice (Chronic Coc) were injected daily for five consecutive days with cocaine (20 mg/kg) and, after a 7 d withdrawal, received an additional injection of saline or cocaine (20 mg/kg). β-Galactosidase-immunoreactive cells (as in Fig. 1a) were quantified in DStr and nucleus accumbens shell using an image analyzer (Image Pro Plus). Data are means ± SEM (n = 3 mice per group) and were analyzed using one-way ANOVA: effect of treatment in the DStr, F(3,11) = 11.20, p < 0.001; NAcc, F(3,11) = 10.76, p < 0.001. Post hoc comparison (Newman–Keuls test), ∗p < 0.01 saline versus cocaine; °p < 0.05 cocaine versus chronic cocaine.
Cocaine-induced locomotor sensitization is decreased in Zif268 mutant mice
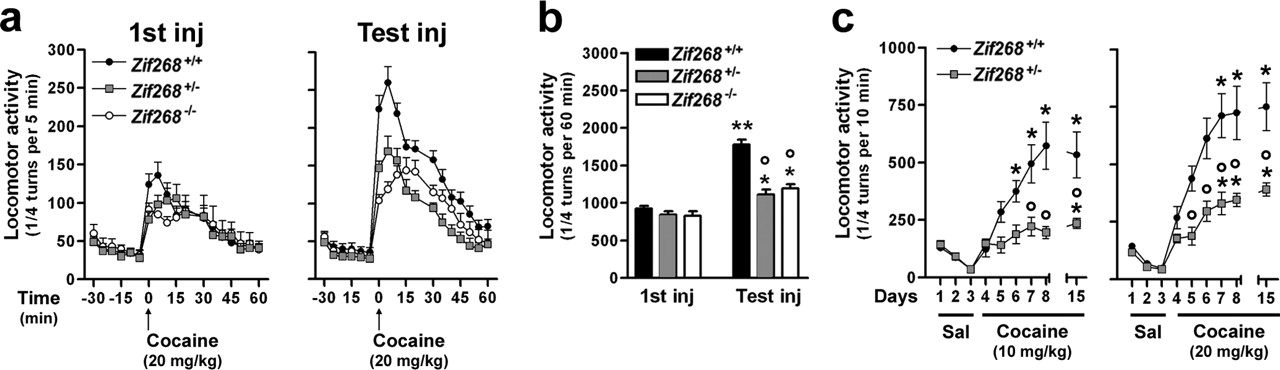
In agreement with previous studies (Jones et al., 2001), Zif268+/−, Zif268−/−, and Zif268+/+ mice displayed similar spontaneous locomotor activities and habituation to novel environment (data not shown). In response to a single injection of cocaine, the total locomotor activity over a 1 h period was not significantly altered (Fig. 3a,b). We then tested the sensitization of locomotor response to cocaine induced by a single dose of this drug administered 1 week before in the same environment. As reported previously (Valjent et al., 2005), a clear sensitization of cocaine locomotor effects was found in wild-type mice (Fig. 3a,b). In contrast, the increased responsiveness to the second cocaine injection was dramatically reduced in both Zif268−/− and Zif268+/− mutant mice (Fig. 3a,b). However, the locomotor response to cocaine was still significantly higher after the second injection than after the first injection in mutant mice (Fig. 3b). We also tested the locomotor sensitization induced by repeated exposure to cocaine (10 or 20 mg/kg) in Zif268+/− mutant mice. Sensitization was markedly reduced compared with wild-type mice, although a slight but significant sensitization was still observed (Fig. 3c). Altogether, these data provide strong evidence that locomotor sensitization involves both Zif268-dependent and independent mechanisms.

Alteration of cocaine-induced locomotor sensitization in Zif268 mutant mice. Locomotor sensitization after a single injection of cocaine (a, b). a, Time course of locomotor activity in Zif268+/+, Zif268+/−, and Zif268−/− mice after the first injection of cocaine (20 mg/kg; 1st inj) and a second injection 7 d later (Test inj). b, Total activity during 60 min. Data (means ± SEM, n = 6–8 per group) were analyzed using two-way ANOVA: effect of treatment, F(1,34) = 118.4, p < 0.01; effect of genotype, F(2,34) = 27.26, p < 0.001. Post hoc comparison (Bonferroni's test), ∗p < 0.05, ∗∗p < 0.001 test versus first injection; °p < 0.01 mutant versus wild type. c, Locomotor sensitization to repeated cocaine injections. After 3 d of habituation, Zif268+/+ and Zif268+/− mice were injected daily for 5 consecutive days with cocaine (10 and 20 mg/kg; n = 10–11 per group) and, after a 7 d withdrawal, received an additional injection of cocaine on day 15. Sal, Saline. Data were analyzed with a mixed-factor ANOVA (repeated measure over time): 10 mg/kg cocaine: effect of time, F(8,171) = 17.23, p < 0.01; effect of genotype, F(1,171) = 42.35, p < 0.01; 20 mg/kg cocaine: effect of time, F(8,177) = 25.80, p < 0.01; effect of genotype, F(1,177) = 66.15, p < 0.01. Post hoc comparison (Bonferroni's test), ∗p < 0.01 later days versus day4; °p < 0.01 Zif268+/+ versus Zif268+/−.
Cocaine-induced conditioned place preference is prevented in mice lacking Zif268
We tested the association between the rewarding effects of cocaine and specific environmental clues in the CPP paradigm. Before conditioning, Zif268+/+, Zif268+/−, and Zif268−/− mice spent the same amount of time in each compartment (Fig. 4a, Preconditioning). After conditioning, both Zif268+/+ and Zif268+/− mice developed a clear preference for the cocaine-paired compartment, whereas no CPP was observed in Zif268−/− mice (Fig. 4a, Postconditioning). The absence of CPP was not attributable to a locomotion impairment of Zif268−/− mice, because their number of visits to each compartment during the preconditioning phase was comparable with that of Zif268+/+ mice (data not shown). Interestingly, although Zif268 heterozygous mice displayed an altered sensitization (see above), their CPP was normal. In addition, when CPP was extinguished by repeated saline pairing in both compartments, its reinstatement by a priming injection of cocaine was similar in Zif268+/+ and Zif268+/− mice (data not shown). Altogether, these results demonstrate that Zif268 is required for the rewarding effects of cocaine.

Absence of cocaine-induced, but not food-induced, conditioned place preference in Zif268−/− mice. a, Cocaine CPP. Before conditioning (Preconditioning), mice showed no initial preference for either side in any of the experimental groups (effect of treatment, F(1,34) = 0.02, NS; effect of genotype, F(2,34) = 0.12, NS). After conditioning (Postconditioning), Zif268+/+ and Zif268+/−, but not Zif268−/−, mice developed a significant place preference for the cocaine-paired side (effect of treatment, F(1,34) = 10.02, p < 0.01; effect of genotype, F(2,34) = 2.46, NS). Data are means ± SEM (n = 6–8 per group). Bonferroni's test, ∗p < 0.05 cocaine versus saline; °p < 0.01 Zif268−/− versus Zif268+/+. b, Food-induced CPP. Before conditioning, no group of mice showed any initial preference for either side (Preconditioning) (effect of treatment, F(1,24) = 0.53, NS; effect of genotype, F(1,24) = 0.07, NS). After conditioning (Postconditioning), Zif268+/+ and Zif268−/− mice showed a significant place preference for the food-paired side (effect of treatment, F(1,24) = 25.39, p < 0.001; effect of genotype, F(1,24) = 0.62, NS). Data are means ± SEM (n = 6–8 per group). ∗p < 0.01 food versus no food.
Food-conditioned place preference in mice lacking Zif268
Because several types of long-term memory are impaired in mice lacking Zif268 (Davis et al., 2003), the alteration of CPP could result from deficits unrelated to the rewarding effects of cocaine but secondary to alterations of other functions, such as spatial memory. To test this hypothesis, we examined the ability of mildly food-deprived Zif268−/− mice to develop CPP to palatable food in a paradigm similar to that used for cocaine. Food reward produced comparable CPP in Zif268+/+ and Zif268−/− mice (Fig. 4b). This result is important because it shows that the mutant mice had the capacity to develop CPP and, thus, demonstrates the specificity of the blockade of cocaine-induced CPP in Zif268−/− mice.
Discussion
Regulation of gene expression is widely acknowledged to play a central role in the mechanisms by which drugs of abuse produce long-lasting changes in the brain (Kelley, 2004; Nestler, 2004). Although the control of gene regulation by the ERK signaling pathway is likely to play a role in behavioral alterations in response to drugs of abuse (Valjent et al., 2000; Valjent et al., 2005), direct evidence for this hypothesis is still missing. In the present study, we provide strong genetic evidence for the critical role of Zif268 in the long-lasting alterations of behavioral responses consecutive to short exposure to cocaine. It is important to underline that mouse genetically deficient for Zif268 had behavioral responses to acute injections of cocaine virtually identical to wild-type mice and displayed no significant alteration of striatal proteins important for signaling in response to cocaine. Moreover, the induction by cocaine of c-fos or the β-galactosidase reporter gene replacing Zif268 was normal, indicating that signaling pathways activated by cocaine upstream from Zif268 were not altered by the mutation. This lack of alteration of acute responses to cocaine gives all their importance to the alterations of delayed behavioral responses to cocaine in these mice.
Locomotor sensitization by single and repeated cocaine administration and cocaine-induced CPP were altered in the mutant mice, revealing the important role of Zif268 in these responses. Zif268 has been shown previously to be necessary for the long-term (i.e., >24 h) retention of memories in paradigms that primarily involve amygdala or hippocampus, such as object recognition and spatial localization (Jones et al., 2001; Bozon et al., 2003). Because the two paradigms used in the present study involve the recognition of environmental clues, it was important to determine whether the deficits in these tests were secondary to other alterations. To do so, we examined CPP to food, a natural reward that involves mechanisms in part different from those activated by cocaine (Kelley, 2004; Baunez et al., 2005) and found it was normal in Zif268 knock-in mice. Zif268 knock-in mice do not display complete loss of memory because they are able to learn after extensive training in several behavioral models (Jones et al., 2001). Normal food CPP demonstrates that mice had enough memory capacity to respond in a CPP paradigm and supports a specific requirement of Zif268 for the rewarding effects of cocaine.
Sensitization was similarly altered in Zif268+/− and Zif268−/− mice, whereas CPP to cocaine was normal in heterozygous animals but completely prevented in Zif268−/− mice. Different effects of Zif268 gene dosage have also been reported for other types of responses, including long-term stabilization of long-term potentiation in the dentate gyrus and several learning tests (Jones et al., 2001; Bozon et al., 2003). This variable impact of reduced levels of Zif268 for different responses may suggest the existence of different molecular mechanisms.
ERK can control gene expression either directly by phosphorylating transcription factors such as Elk1 or indirectly by activating kinases that regulate transcription factors and chromatin accessibility (Brami-Cherrier et al., 2005). We have shown recently that MSK1 is a major effector of ERK in response to cocaine in vivo, controlling the phosphorylation of CREB and histone H3 and the induction of c-fos and preprodynorphin (Brami-Cherrier et al., 2005). In contrast, the induction of Zif268 was normal in MSK1-deficient mice (Brami-Cherrier et al., 2005), indicating that it did not depend on MSK1. Comparison of the phenotypes of Zif268−/− and MSK1 knock-out mice provides interesting clues concerning the molecular mechanisms by which cocaine exerts its long-lasting effects. First, although sensitization was dramatically altered in Zif268−/− mice, they displayed a significantly higher locomotor activity in response to the second injection of cocaine than to the first injection, indicating the existence of a Zif268-independent component in the sensitization. A similar residual locomotor sensitization was observed in MSK1 mutant mice (Brami-Cherrier et al., 2005). Because Zif268 induction is independent of MSK1 (Brami-Cherrier et al., 2005), it appears likely that at least two different signaling mechanisms, respectively, MSK1 and Zif268 dependent, with additive effects, control locomotor sensitization downstream from ERK. In contrast, CPP was completely blocked in the absence of Zif268 (present study), whereas it was increased in MSK1 mutant mice (Brami-Cherrier et al., 2005). This contrast reveals the critical role of Zif268 compared with the MSK1-dependent events (including CREB and histone H3 phosphorylation) in CPP, i.e., in the association of environmental clues with the effects of cocaine. Because induction of Zif268 in NAcc is markedly decreased after repeated cocaine injections (Bhat et al., 1992; Hope et al., 1992; present study), it is likely that it plays a predominant role for initial plasticity induced by cocaine.
Recently, Zif268 expression was found to be increased by conditioned stimuli previously paired with cocaine self-administration in specific brain areas such as the NAcc, the ventral tegmental area, the prefrontal cortex, and the basolateral amygdala (Thomas et al., 2003). Zif268 also participates in the molecular mechanisms underlying memory reconsolidation (Bozon et al., 2003; Lee et al., 2004), and interesting results supporting the role of Zif268 in the reconsolidation of stimulus–drug associations have been obtained using stereotaxic injection of antisense oligonucleotides (Thomas et al., 2003; Lee et al., 2005). Our results directly demonstrate that Zif268 is required for the rewarding effects of cocaine. The precise relationships between the molecular mechanisms underlying the role of Zif268 in the response to cocaine injection (i.e., the unconditioned stimulus), as demonstrated here, and to environmental clues (i.e., conditioned stimuli) will be an interesting area of investigation. Moreover, the identification of genes specifically regulated by Zif268 will help to decipher how these transcriptional effects are implemented at the synaptic level.
Footnotes
-
This work was supported by Institut National de la Santé et de la Recherche Médicale (INSERM) and by grants from Agence Nationale de la Recherche, Fondation pour la Recherche Médicale, Fondation Liliane Bettencourt, and Mission Interministérielle pour la Lutte contre la Drogue et la Toxicomanie (J.-A.G.) and Action Concertée Incitative Physiologie et Développement (J.C., J.-A.G.). B.A. was supported by a fellowship from Fondation pour la Recherche Médicale. A.-G.C. was supported by a scholarship from Ecole de l'INSERM. We thank Dr. P. Greengard, P. Lombroso, and A. C. Nairn for providing antibodies.
- Correspondence should be addressed to Dr. Jean-Antoine Girault, Institut National de la Santé et de la Recherche Médicale Unité 536, Institut du Fer à Moulin, 17 rue du Fer à Moulin, 75005 Paris, France. Email: girault{at}fer-a-moulin.inserm.fr





{kind=link}
{kind=link}
{kind=link}
{kind=link}