

Abstract
The trafficking of GABAA receptors is an important component of the pathway that regulates plasticity of inhibitory synapses. The 17 kDa GABAA receptor-associated protein (GABARAP) has been implicated in the trafficking of GABAA receptors because of its ability to interact not only with the γ2 subunit of the receptor but also with microtubules and the N-ethylmaleimide-sensitive factor (NSF). To elucidate the role of GABARAP in the trafficking of GABAA receptors, we have constructed a yellow fluorescent protein (YFP) fusion protein of GABARAP and expressed it in neurons using adenovirus, so that its function may be examined. YFP-GABARAP colocalized with γ2 subunit-containing GABAA receptors and NSF to the perinuclear cytoplasm in cultured hippocampal neurons and to the proximal regions of dendrites that are making synaptic contact. Expression of YFP-GABARAP in Cos7 cells and cultured hippocampal neurons was able to increase the level of GABAA receptors detected at the plasma membrane, even at low levels of YFP-GABARAP expression. This effect is specific to the function of GABARAP on GABAA receptor trafficking, because point mutations in the γ2-binding domain of YFP-GABARAP interfered with the ability of YFP-GABARAP to increase GABAA receptor surface levels. These mutations also disrupted the colocalization of YFP-GABARAP with the γ2 subunit and with NSF in hippocampal neurons. The results of this study show for the first time that GABARAP has a functional effect on the trafficking of GABAA receptors and provide decisive evidence for the role of GABARAP in transporting GABAA receptors to the plasma membrane in neurons.
Introduction
The GABAA receptor is the predominant inhibitory neurotransmitter receptor in the brain (Macdonald and Olsen, 1994). The pentameric channel can be composed of many possible subunits, but the most common combination in the brain is composed of α1, β2, and γ2 subunits (McKernan and Whiting, 1996). The plasticity of inhibitory synapses in neurons probably involves the regulation of GABAA receptor trafficking between subcellular compartments and the plasma membrane, which requires the major components of the cytoskeleton, because disruption of microtubules and actin microfilaments has been shown to reduce cell surface expression (Meyer et al., 2000) and alter channel properties of GABAA receptors (Petrini et al., 2003). In addition, GABAA receptors can immunoprecipitate actin and tubulin from rat brain homogenates (Kannenberg et al., 1997). GABAA receptor-associated protein (GABARAP) may provide a link between GABAA receptors and the cytoskeleton during trafficking, because it binds to the intracellular loop of the γ2 subunit of the GABAA receptor in vitro and immunoprecipitates with GABAA receptors from rat brain homogenates (Wang et al., 1999; Nymann-Andersen et al., 2002b), while it also interacts with microtubules in cells and immunoprecipitates with brain tubulin (Wang et al., 1999; Wang and Olsen, 2000).
The crystal structure of GABARAP suggests that GABARAP forms a compact globular structure made of a flexible N terminus (1-27) composed of two helices and a C terminus (27-117) that forms a compact globular structure that is structurally homologous to ubiquitin (Coyle et al., 2002). The binding domain for the γ2 subunit is from amino acids 36-52 of GABARAP (Nymann-Andersen et al., 2002b), which lies between the β1 and β2 strands of the ubiquitin-like core domain (Coyle et al., 2002).
GABARAP also interacts with the N-ethylmaleimide-sensitive factor (NSF) and colocalizes with NSF to intracellular membrane compartments and subsynaptic cisternae in neurons (Kittler et al., 2001). NSF is involved in a variety of intracellular vesicle transport and fusion events (May et al., 2001), including intra-Golgi trafficking through its interaction with the GABARAP homolog GATE-16 (Sagiv et al., 2000) and AMPA receptor trafficking through its interaction with the glutamate receptor 2 (GLUR2) subunit (Hanley et al., 2002). GABARAP also interacts with gephyrin (Kneussel et al., 2000), which binds microtubules and is known to be an important component for anchoring both glycine and GABAA receptors at the plasma membrane (Kneussel and Betz, 2000). However, a recent study shows that gephyrin assists in the trafficking of glycine receptors to synapses (Hanus et al., 2004), implying that gephyrin may be involved in the trafficking of GABAA receptors, perhaps through its interaction with GABARAP.
Although the literature suggests that GABARAP is involved in the trafficking of GABAA receptors in neurons, the precise role of GABARAP in this process is unclear. By using adenovirus to express yellow fluorescent protein (YFP)-GABARAP in cultured neurons, we have shown that its overexpression increases the surface levels of γ2 subunit-containing GABAA receptors. In addition, we have shown that point mutations in the γ2-binding domain of GABARAP can disrupt this phenotype, suggesting that the GABARAP binding to GABAA receptors is essential for its ability to traffic receptors to the plasma membrane.
Materials and Methods
Plasmid construction and mutagenesis of GABARAP. The wild-type rat GABARAP cDNA was PCR amplified from pGEX-226(Full) (Wang et al., 1999) using primers that introduced a 5′ NotI (5′-GCGGCCGCTATGAAGTTCGTGTACAAAGAGG-3′) and a 3′ KpnI (5′-ACGGTACCTCACAGACCATAGACCCTTTC-3′) restriction site and was then cloned into the NotI/KpnI sites of pShuttle (BD Biosciences-Clontech) to generate pSh-RAP. To generate the YFP-GABARAP fusion construct, the enhanced YFP (EYFP) cDNA was PCR amplified from pEYFP (BD Bioscience-Clontech) using primers that introduced a 5′ NheI (5′-ATGCTAGCCACCATGGTGAGCAAGGG-3′) and a 3′ NotI (5′-GCGGCCGCCTTGTACAGCTCGTCCAT-3′) restriction site and was then subsequently cloned upstream of the GABARAP cDNA into the NheI/NotI sites of pSh-GABARAP to generate pSh-YFPRAP. The mouse cDNA for the γ2L subunit of the GABAA receptor was PCR amplified from the pRK5-γ2L (Chen et al., 2000) using primers that introduced a 5′ NotI (5′-GCGGCCGCTAATACATGGAGCACTGGAAGC-3′) and a 3′ KpnI (5′-ACGGTACCTTACAGATAAAGATAGGAGACCC-3′) restriction site and was then cloned into the NotI/KpnI sites of pShuttle to generate pSh-γ2. The cDNA for enhanced cyan fluorescent protein (CFP) was PCR amplified using primers that introduced a 5′ NheI (5′-ATATCCTAGCCACCATGAGTTCGCCAATGGTGAGCAAGGGCGAGG-3′) and a 3′ NotI (5′-GCGGCCGCCTTGTACAGCTCGTCCAT-3′) restriction site and was then cloned upstream of the γ2L cDNA in pSh-γ2 using the NheI/NotI sites to produce pSh-CFPγ2. The rat cDNA for the α1 subunit was PCR amplified from pBS-α1 (a donation from Allan Tobin) using primers that introduced a 5′ NotI (5′-GCGGCCGCATGAAGAAAAGTCGGGGTCTC-3′) and a 3′ XhoI (5′-AACTCGAGCTATTGATGGGGTGTGGGGGC-3′) and was then cloned into the NotI/XhoI sites of pShuttle to produce pSh-α1.
Generation of point mutations in GABARAP was performed as described previously (Ho et al., 1989). pSh-RAP was used as a template for the flanking primers pSh-MCSUpst (5′-GCAAATGGGCGGTAGGCGTG-3′) and pSh-MCSDst (5′-GGAGGGGCAAACAACAGATGGC-3′) with the internal mutagenic primers as follows: for mutation of Pro37 to Ala (GRapFM-P37A, 5′-GGAAAAAGCCGCCAAAGCTCGG-3; GrapRM-P37A, 5′-CCGAGCTTTGGCGGCTTTTTCC-3′); for mutation of Lys38 to Ala (GRapFM-K38A, 5′-CCCCCGCAGCTCGGATAGG-3′; GRapRM-K38A, 5′-CCTATCCGAGCTGCGGGGG-3′); for mutation of Lys38 and Arg40 to Ala (GRapFM-K38A/R40A, 5′-CCGCAGCTGCGATAGGAGACCTG-3′; GRapRM-K38A/R40A, 5′-CAGGTCTCCTATCGCAGCTGCGG-3′). After mutagenesis PCR, the PCR products were cloned into the TA cloning vector pCR2.1 (Invitrogen, San Diego, CA) and subsequently cloned back into pSh-YFPRAP using the NotI/KpnI sites to replace the existing wild-type GABARAP sequence.
Generation of adenoviral targeting vectors. Generation of recombinant adenovirus was performed essentially as described in the BD AdenoX user manual (BD Biosciences-Clontech). The expression cassettes containing the GABAA receptor subunits, YFP-GABARAP, and the mutated forms of YFP-GABARAP from pShuttle were cloned into pAdenoX using the PI-SceI/I-CeuI restriction sites. Identification of genomically complete recombinant viral constructs was performed as described previously (Mizuguchi and Kay, 1998). Infectious adenoviral particles were packaged in human embryonic kidney-293 (HEK-293) cells (American Type Culture Collection, Manassas, VA) as described in the AdenoX user manual (BD Biosciences-Clontech).
Primary culture of hippocampal neurons. To study the effects of YFP-GABARAP on neurons, primary culture of rat hippocampal neurons was prepared in the following manner. Adult female rats, pregnant with embryonic day 18 embryos, were killed with halothane, the embryos were removed, and the hippocampi were dissected in HBSS (Invitrogen). The dissected hippocampi were cut into small pieces and processed using the Papain Dissociation System (Worthington Biochemical, Freehold, NJ). After dissociation, the cells were counted and plated in serum-free Neurobasal medium supplemented with B27 (Invitrogen), l-glutamine (0.5 mm), and glutamate (25 μm) and grown at 37°C in a humidified incubator with 5% CO2. After 4 d, the medium was replaced with Neurobasal/B27 medium containing only l-glutamine. For biochemical assays, neurons were plated at a density of 300,000 cells per well in 6-well plates coated with poly-d-lysine (50 μg/ml); for microscopy, neurons were plated at a density of 40,000 cells per well in 24-well plates containing coverslips that had been coated with poly-d-lysine (100 μg/ml). Primary neurons were cultured for 7-10 d before infection with adenovirus at titers of 50-100 virus particles per neuron.
Immunoprecipitation and immunoblotting. Homogenates were prepared from cultured neurons that had been infected with YFP-GABARAP adenovirus on day 8. On day 11, hippocampal neurons were scraped and homogenized in (in mm) 20 3-[(3-cholamidopropyl) dimethylammonio]-1-propanesulfonate, 50 Tris, pH 8.0, 50 KCl, 5 MgCl2, 1 EDTA, and 2 phenylmethylsulphonyl fluoride at 4°C. Immunoprecipitation was performed at 4°C overnight, using polyclonal goat anti-green fluorescent protein (GFP) antibody (Rockland Immunochemicals, Gilbertsville, PA) cross-linked to protein G-agarose (Sigma, St. Louis, MO). The GFP antibody was cross-linked to protein G-agarose using the disuccinyl suberate reagent (Pierce, Rockford, IL) according to the manufacturer's specifications. After washing the immunoprecipitate three times with immunoprecipitation buffer, the protein bound to the column was eluted with the Immunopure IgG Elution Buffer (Pierce) supplemented with 1% Triton X-100. The eluted protein was separated on a gradient (4-15%), Tris-HCl, and SDS-PAGE (Bio-Rad, Hercules, CA), then transferred and blocked in PBS, 0.1% Tween, and 5% dry milk for 1 hr and probed using rabbit anti-GABARAP 2627 (1:5000) (Wang et al., 1999), rabbit anti-γ2 (1:2500; Chemicon, Temecula, CA), or mouse anti-NSF (1:5000; Transduction Laboratories, Lexington, KY) antibody in blocking buffer for 1 hr. Secondary detection was performed using either HRP-goat anti-rabbit (1:5000; Santa Cruz Biotechnology, Santa Cruz, CA) or HRP-goat anti-mouse (1:5000; Santa Cruz Biotechnology) in blocking buffer for 1 hr, followed by chemiluminescent detection using the ECL kit (Amersham Biosciences, Piscataway, NJ). All Western blots were exposed to Biomax film (Eastman Kodak, Rochester, NY), the films were scanned, and immunoreactive bands were quantified using Scion Image software (Scion, Frederick, MD).
Immunofluorescence microscopy and colocalization. On the third day after infection, neurons were immunofluorescence labeled at room temperature in the following manner. Coverslips were washed once with PBS, then fixed in 4% paraformaldehyde in PBS for 15 min, followed by permeabilization in PBS and 0.1% Triton X-100 for 10 min (for surface labeling, the permeabilization step was performed after surface labeling with primary antibody and fixation). Neurons were then incubated in blocking buffer (PBS, 10% goat serum, 0.2% BSA, and 0.01% sodium azide) for 1 hr, followed by incubation with primary antibodies for 2 hr in antibody buffer (PBS, 2% goat serum, 0.2% BSA, and 0.01% sodium azide), washed three times (5 min each) in PBS, then incubated with fluorescently labeled secondary antibodies in antibody buffer for 1 hr and washed again three times (5 min each) in PBS. The primary antibodies used were as follows: rabbit anti-γ2 (1:200; GAG2; Alpha Diagnostics); mouse anti-NSF (1:400; Transduction Laboratories); mouse anti-GAD65 (1:400; Santa Cruz Biotechnology); and mouse anti-synaptophysin (1:150; Sigma). Secondary antibodies conjugated to fluorophores were as follows: goat anti-rabbit conjugated to Alexa Fluor 568 (1:500; Molecular Probes, Eugene, OR) and goat anti-mouse conjugated to Alexa Fluor 647 (1:500; Molecular Probes). For visualization of cell nuclei, 4′,6-diamidino-2-phenylindole, dihydrochloride (DAPI; Molecular Probes) was incubated with cells at a concentration of 1 μm in PBS for 5 min subsequent to antibody immunostaining.
After immunofluorescent labeling, the coverslips were mounted onto slides using FluorSave reagent (Calbiochem, La Jolla, CA) and stored overnight in the dark. All multicolor colocalization studies were performed on the Leica TCS SP MP inverted or fixed stage upright confocal microscope (Carol Moss Spivak Cell Imaging Facility, Gonda Center for Neuroscience and Genetics, University of California Los Angeles, Los Angeles, CA) using the 100× oil immersion objective. Z-stacks (optical sections) of the images were collected with an optical thickness of 0.2 μm and processed using the Leica TCS-NT/SP software Metavue (Universal Imaging, West Chester, PA) and Adobe Photoshop (Adobe Systems, Mountain View, CA). Fluorochromes were excited using the argon laser at 488 nm for YFP, the krypton laser at 568 nm for Alexa Fluor 568, or the neon-helium laser at 633 nm for Alexa Fluor 647.
Quantification of colocalization was performed using Image Pro Plus (Media Cybernetics, Silver Spring, MD). Pearson's r values were determined according to the Image Pro Plus software application note 1, “Colocalization of Fluorescent Probes” (Media Cybernetics). The significance of the Pearson's r values for single data sets was determined using a two-tailed Student's t test, whereas statistical comparison between data sets was performed using the JMP Statistical Package (SAS Institute, Cary, NC).
Measurement of GABAA receptor surface expression. Cos7 cells (American Type Culture Collection) were grown in DMEM and 10% FBS (Invitrogen) for 3 d in a T-25 until 50% confluent, then transfected with pCDNA3-β2 (Chen et al., 2000) using Lipofectamine reagent (Invitrogen) to express the rat β2 subunit. After 24 hr, the growth medium was replaced with medium containing the α1 and CFPγ2 adenovirus at a multiplicity of infection (MOI) of ∼20 virus particles per cell and one of either the wild-type or mutant YFP-GABARAP adenovirus at an MOI of ∼10 virus particles per cell. Forty-eight hours after viral infection, the cells were dislodged from the T-25 flask using PBS and 50 mm EDTA and placed on ice. Cells were spun down at 4°C, resuspended in antibody blocking buffer (PBS, 10% goat serum, 0.2% BSA, and 0.01% sodium azide), and blocked for 30 min on ice; cells were spun down again, resuspended in antibody incubation buffer (PBS, 2% goat serum, 0.2% BSA, and 0.01% sodium azide) containing mouse anti-GFP (1:500; Santa Cruz Biotechnology) antibody, and incubated on ice for 30 min. Cells were then washed three times with cold PBS, resuspended in antibody incubation buffer containing cyanine 5 (Cy5)-conjugated goat anti-mouse secondary antibody (1:500; Chemicon), and incubated for 30 min, followed by three PBS washes. Cells were then fixed in PBS and 4% paraformaldehyde for 10 min, washed three times in ice-cold PBS, and placed at 4°C to await flow cytometry. Flow cytometry was performed using the FACSVantage SE sorting flow cytometer (Becton Dickinson, Mountain View, CA). Fluorochromes were excited using the argon laser at 355 nm for CFP, another argon laser at 488 nm for YFP, and the krypton-argon mixed gas spectrum laser at 647 nm for Cy5. A pool of 2000-5000 cells that displayed both CFP fluorescence (expressing CFP-GABAA receptors) and Cy5 fluorescence (surface labeling with GFP antibody) was selected for selected for additional analysis. The flow cytometry data were analyzed using the JMP Statistical Package (SAS Institute).
Measurement of GABAA receptor surface expression in cultured hippocampal neurons was performed as follows. Nonpermeabilized infected neurons were immunofluorescence labeled for surface γ2 as described above, then permeabilized and labeled using mouse anti-MAP2 (1:150; Sigma) primary antibody and goat anti-mouse secondary antibody conjugated to Pacific blue (1:300; Molecular Probes), followed by mounting of the coverslips in FluorSave reagent (Calbiochem). Fluorescence microscopy was performed on the Nikon Eclipse TE200 inverted epifluorescence microscope, and three-color digital images were collected using the Photometrics CoolSNAP cf cooled CCD camera. The CFP filter was used for detection of MAP2 immunofluorescence, the YFP filter was used for detection of YFP fluorescence, and the tetramethylrhodamine isothiocyanate filter was used for detection of γ2 fluorescence. For each experimental condition, three-color images from 30-40 neurons were collected from four separate wells and analyzed quantitatively using Metavue (Universal Imaging). For each neuron, a region of interest (ROI) was drawn around the cell body of the neuron based on the somatic labeling by MAP2, and the average pixel intensity for the three-color channels in that ROI was measured. The average pixel intensity from the γ2 immunofluorescence from 30-40 neurons for each sample was averaged for each sample and analyzed statistically using the JMP Statistical Package (SAS Institute).
Results
Localization of YFP-GABARAP in hippocampal neurons
To examine the expression of GABARAP in cultured neurons, an N-terminal fusion protein between YFP and wild-type GABARAP was generated. Infection of cultured hippocampal neurons with YFP-GABARAP adenovirus resulted in high expression levels as well as a relatively high efficiency of gene transduction. In most cases, infection of cultures with an MOI of 50 adenovirus particles per cell resulted in ∼30% of the neurons displaying YFP-GABARAP fluorescence, while minimizing the extent of toxicity to the neurons. Glial cells appeared to be more easily transduced by adenovirus but were rarely observed in cultures because the neurons were grown in serum-free Neurobasal medium, which inhibits proliferation of glial cells (Xie et al., 2000). Neurons infected with YFP-GABARAP appeared morphologically normal and expressed the neuronal marker MAP2 (Fig. 1A), which is known to be a specific marker of neuronal cell bodies and dendrites (Fischer et al., 1986). Examination of the expression pattern for wild-type YFP-GABARAP demonstrated primarily cytoplasmic fluorescence (Fig. 1A), with some punctate staining extending out into some of the MAP2-labeled dendrites of the neuron. Nuclear staining of cultured hippocampal neurons expressing YFP-GABARAP revealed that the somatic fluorescence of YFP-GABARAP is primarily perinuclear (Fig. 1B). This perinuclear region also contained significant amounts of γ2 subunit-containing GABAA receptors (Fig. 1B).

Expression of YFP-GABARAP in cultured hippocampal neurons. A, YFP-GABARAP adenovirus was used to infect cultured hippocampal neurons, then cells were fixed and immunostained with antibodies to MAP2 to visualize soma and dendrites of neurons. YFP-GABARAP fluorescence and MAP2 immunostaining were visualized using fluorescence microscopy. YFP-GABARAP was mostly localized to the MAP2-labeled cell body, but YFP-GABARAP puncta were occasionally seen in MAP2-labeled dendrites. B, Fluorescence images of another cultured hippocampal neuron infected with YFP-GABARAP adenovirus and subsequently fixed, permeabilized, immunostained with antibodies to the γ2 (red) subunit to visualize intracellular GABAA receptors, and stained with DAPI. The nuclear DAPI (blue) staining shows the cytoplasmic perinuclear expression pattern of YFP-GABARAP (green) and the colocalization of intracellular γ2 subunit immunostaining to this perinuclear region, which appears as yellow in the overlay image. Scale bars, 20 μm.
YFP-GABARAP did not appear to colocalize extensively with surface GABAA receptors (Fig. 2A), which is consistent with previously published reports (Kittler et al., 2001). Cultured hippocampal neurons express significant quantities of γ2 subunit-containing GABAA receptors in intracellular pools (Kneussel et al., 1999). Confocal microscopy using anti-γ2 subunit antibody in permeabilized neurons revealed that YFP-GABARAP fluorescence colocalizes extensively with γ2 immunofluorescence staining in an intracellular region of the cell body (Fig. 2B). Calculation of the Pearson correlation coefficient (Table 1) revealed that the colocalization between YFP-GABARAP fluorescence and γ2 immunofluorescence from confocal slices was highly significant (t test; p = 0.001). Pearson correlation coefficients (Table 1) also revealed a highly significant (t test; p < 0.0001) colocalization between YFP-GABARAP and NSF immunofluorescence. YFP-GABARAP can be seen to colocalize to a cytoplasmic, perinuclear region of the cell body with NSF in hippocampal neurons (Fig. 2C). Occasionally, YFP-GABARAP was found to colocalize with NSF in the dendrites of neurons (Fig. 2C, arrow).

YFP-GABARAP colocalizes with GABAA receptors intracellularly but not at the cell surface. A, YFP-GABARAP adenovirus was used to infect cultured hippocampal neurons, and surface GABAA receptors were detected before fixation and permeabilization using antibodies to the γ2 subunit (red); then the neurons were permeabilized and immunostained using antibodies to GAD65 (blue). A single three-color confocal z-plane is shown. YFP-GABARAP fluorescence (green) does not appear to colocalize with surface γ2 subunit-containing GABAA receptors. Synapses are labeled by colocalization of GAD65 and γ2 (arrows in merged image). B, A cultured hippocampal neuron expressing YFP-GABARAP (green), which was fixed and permeabilized to stain for intracellular γ2 (red) subunit-containing GABAA receptors. Three dual-color confocal z-planes from the same neuron are shown to demonstrate the three-dimensional intracellular colocalization (appears as yellow in the merged image) between YFP-GABARAP and the γ2 subunit. Colocalization of YFP-GABARAP and γ2 is apparent mainly in the perinuclear cytoplasm of the cell body but is also found in the proximal portions of dendrites. C, A cultured hippocampal neuron expressing YFP-GABARAP (green), which was fixed and permeabilized, then immunostained using antibodies to NSF (red). There is extensive perinuclear colocalization between YFP-GABARAP and NSF (appears as yellow in the merged image), but there is also some dendritic colocalization between the two proteins (arrow). Scale bars, 20 μm.
Quantification of colocalization with Pearson correlation coefficients
Synaptophysin is an established marker of synaptic vesicles (Floor and Feist, 1989). Immunofluorescence staining with synaptophysin revealed that YFP-GABARAP was expressed in dendrites that are making synaptic contact (Fig. 3A). Extensive punctate YFP-GABARAP fluorescence could be found in the proximal region of one of the dendrites that is making extensive synaptic contacts labeled by colocalization of γ2 and synaptophysin immunofluorescence, perhaps indicating a need for YFP-GABARAP to transport γ2 subunit-containing GABAA receptors to the distal portions of the dendrite. GAD65 is also a well established marker of GABAergic presynaptic nerve terminals (Benson et al., 1994). Figure 3B shows a permeabilized pyramidal neuron receiving GABAergic input as delineated by GAD65 staining. Magnification of the cell body revealed that γ2 colocalizes only with a subset of GAD65-labeled presynaptic nerve terminals in the cell body of this neuron (Fig. 3B). This is consistent with previously published reports suggesting that only a subset of GAD-positive presynaptic terminals contain γ2 immunoreactivity (Scotti and Reuter, 2001). Examination of YFP-GABARAP fluorescence in this neuron showed that YFP-GABARAP was primarily expressed in an intracellular perinuclear ring, where it colocalized with γ2 immunofluorescence. However, some YFP-GABARAP puncta were found with γ2 near the surface of the soma at points of synaptic contact labeled by GAD65 (Fig. 3B, arrows). Gephyrin is another marker of both synaptic and extrasynaptic γ2 subunit-containing GABAA receptors (Christie et al., 2002) but has recently been found to be involved in the intracellular trafficking of glycine receptors (Hanus et al., 2004). Colocalization between γ2 subunit and gephyrin immunofluorescence was evident in dendrites, but it was also apparent in a perinuclear region in the cell body of neurons (Fig. 3C). Interestingly, this perinuclear region also contained extensive YFP-GABARAP fluorescence (Fig. 3C).

Colocalization of YFP-GABARAP with synaptic markers and GABAA receptors. A, YFP-GABARAP adenovirus was used to infect cultured hippocampal neurons, then cells were fixed, permeabilized, and coimmunostained using antibodies to the γ2 subunit (red) and synaptophysin (blue). The top images show a single three-color confocal z-plane of two hippocampal neurons, one of which is expressing YFP-GABARAP (green), and the bottom images show a 2.5× zoom of one of the dendrites. Arrows indicate the expression of YFP-GABARAP in the proximal portion of a dendrite making extensive synaptic contacts as labeled by synaptophysin. B, A cultured hippocampal neuron expressing YFP-GABARAP (green) and permeabilized to stain for γ2 subunit-containing GABAA receptors (red) and GAD65 (blue). The top images show a single three-color confocal z-plane of a neuron displaying extensive GABAergic synapses as labeled by colocalization of γ2 and GAD65 (appears as magenta in the merged image), and the bottom images are a 2.5× zoom of the cell body. The arrows show points of colocalization between γ2, GAD65, and YFP-GABARAP. C, A cultured hippocampal neuron expressing YFP-GABARAP (green) and permeabilized to stain for γ2 subunit-containing GABAA receptors (red) and gephyrin (blue). The extensive intracellular colocalization between YFP-GABARAP, gephyrin, and γ2 can be seen in the merged image on the right. Scale bars, 20 μm.
YFP-GABARAP increases surface expression of GABAA receptors
GABAA receptors composed of wild-type α1 and β2 subunits were expressed in Cos7 cells along with an engineered γ2 subunit in which the N terminus was fused with CFP. This resulted in the expression of GABAA receptors that contain a γ2 subunit that projects CFP to the extracellular side of the membrane. Expression of these receptors using adenovirus in both Cos7 and HEK-293 cells resulted in the formation of functional receptors as demonstrated by specific binding of the radioligands muscimol and flunitrazepam (data not shown). Muscimol is a GABAA agonist that binds at the interface between the α1 and β2 subunits, and flunitrazepam is a benzodiazepine that binds at the interface between the α1 and γ2 subunits (Smith and Olsen, 1995).
Flow cytometry was used to measure the surface labeling of GABAA receptors using an antibody to GFP on nonpermeabilized Cos7 cells, comparing between cells that were or were not expressing YFP-GABARAP. The levels of YFP-GABARAP expression were separated into four groups, based on the level of YFP fluorescence found in that group of cells (Fig. 4A). At the lowest level of YFP-GABARAP expression, the ratio of surface receptors to total receptors was increased by 58% compared with cells that had no YFP-GABARAP expression (Fig. 4A) (0 vs 1, p < 0.0001; one-way ANOVA). The increase in surface GABAA receptor labeling increased as the level of YFP-GABARAP expressed in the cells increased (Fig. 4A)(1 vs 2, p = 0.01; 2 vs 3, p = 0.006; one-way ANOVA). To determine whether YFP-GABARAP could have a similar effect in neurons, the surface levels of γ2 subunit-containing GABAA receptors were measured in infected cultures, and cells that expressed high levels of YFP-GABARAP were compared with cells that did not express YFP-GABARAP. Expression of high levels of YFP-GABARAP in cultured hippocampal neurons caused a 26% increase (t test; p < 0.0001) in the surface labeling of endogenous γ2 subunit-containing GABAA receptors (Fig. 4B).

Effect of YFP-GABARAP on surface expression of GABAA receptors. A, Expression of wild-type YFP-GABARAP caused a significant increase in the surface levels of heterologously expressed GABAA receptors (CFP-γ2:β2:α1) in Cos7 cells, as measured by flow cytometry. The levels of GABAA receptors detected at the surface increased in a dose-dependent manner as the level of YFP-GABARAP expression increased. The ratio of surface/total receptors is measured as the ratio of surface CFP-γ2 immunofluorescence (Cy5-conjugated GFP antibody fluorescence) to total CFP-γ2 fluorescence. The level of YFP-GABARAP expression is divided into four bins: 0, no expression; 1, low expression (bottom 40% of cells); 2, medium expression (middle 40% of cells); 3, high expression (top 20% of cells). B, Expression of YFP-GABARAP in culture hippocampal neurons caused a significant increase in the surface levels of endogenous GABAA receptors as detected by immunostaining of nonpermeabilized neurons using antibodies to the γ2 subunit of the receptor. Receptor levels were measured in units of florescence intensity.
Mutations in the γ2-binding domain of YFP-GABARAP reduce interaction with γ2 and NSF
The GABARAP homolog GATE-16/GABARAP L2 has 57% amino acid identity overall and has 50% identity within the γ2-binding domain from amino acids 36-52 (Fig. 5A). Because GATE-16 does not bind to γ2, it was hypothesized that the amino acids that differ between the two proteins may, if mutated, disrupt the binding of GABARAP to the γ2 subunit (Kneussel et al., 2000). Three amino acids likely to be involved in binding γ2 were selected in the region of GABARAP from amino acids 36-52 based on their lack of conservation within the amino acid sequence of GATE-16 (Fig. 5A). The solution structure of GABARAP has previously been resolved at high resolution (Stangler et al., 2002). Mapping of the three amino acids selected for alanine mutagenesis to the solution structure of GABARAP reveals that they are located between the first and second β-strand of GABARAP and that they are exposed to the surface of the protein (Fig. 5B).

Strategy for mutagenesis of GABARAP. A, GABARAP is highly homologous to GATE-16 but does not bind to the γ2 subunit of the GABAA receptor. The amino acids shown with the arrows were selected for mutagenesis based on the difference in homology between the two proteins. B, The crystal structure of GABARAP is shown. The globular structure with surface rendering is shown on the left, and the ribbon cartoon structure is shown on the right. The amino acids that were selected for mutagenesis are labeled in both structures (P37, red; K38, magenta; R40, gray). α Helices are blue, β-sheets are green, and random coils are yellow.
Previous studies have shown that antibody to GABARAP can immunoprecipitate with GABAA receptors from rat brain homogenate (Wang et al., 1999). To determine whether wild-type and mutated forms of YFP-GABARAP could immunoprecipitate γ2 subunit-containing GABAA receptors, cultured hippocampal neurons were infected with adenovirus to express wild-type, P37A, K38A, or K38A/R40A forms of YFP-GABARAP. As a positive control, adenovirus was used to express an N-terminal fusion of V5 with GABARAP as well. Wild-type YFP-GABARAP was expressed at similar levels to the mutated forms of YFP-GABARAP or to V5-GABARAP in homogenates of cultured hippocampal neurons (Fig. 6A). Wild-type and mutated forms of YFP-GABARAP were immunoprecipitated using GFP antibody, whereas V5-GABARAP was immunoprecipitated using antibody to the V5 epitope. Immunoreactivity to the γ2 subunit was found to coimmunoprecipitate with wild-type YFP-GABARAP and with V5-GABARAP (Fig. 6B). The quantity of γ2 that was pulled down by wild-type YFP-GABARAP and V5-GABARAP was very similar when quantified and normalized to the amount of GABARAP present in each lane (Fig. 6C). The mutated forms of YFP-GABARAP showed variable degrees of interaction with γ2 subunit-containing GABAA receptors (Fig. 6C). The YFP-GABARAP mutants P37A and the double mutant K38A/R40A displayed the weakest interaction with γ2 but were higher than the negative control, suggesting that they still possessed some interaction (Fig. 6B). The K38A mutant showed a stronger interaction with γ2 but was still reduced compared with wild type.
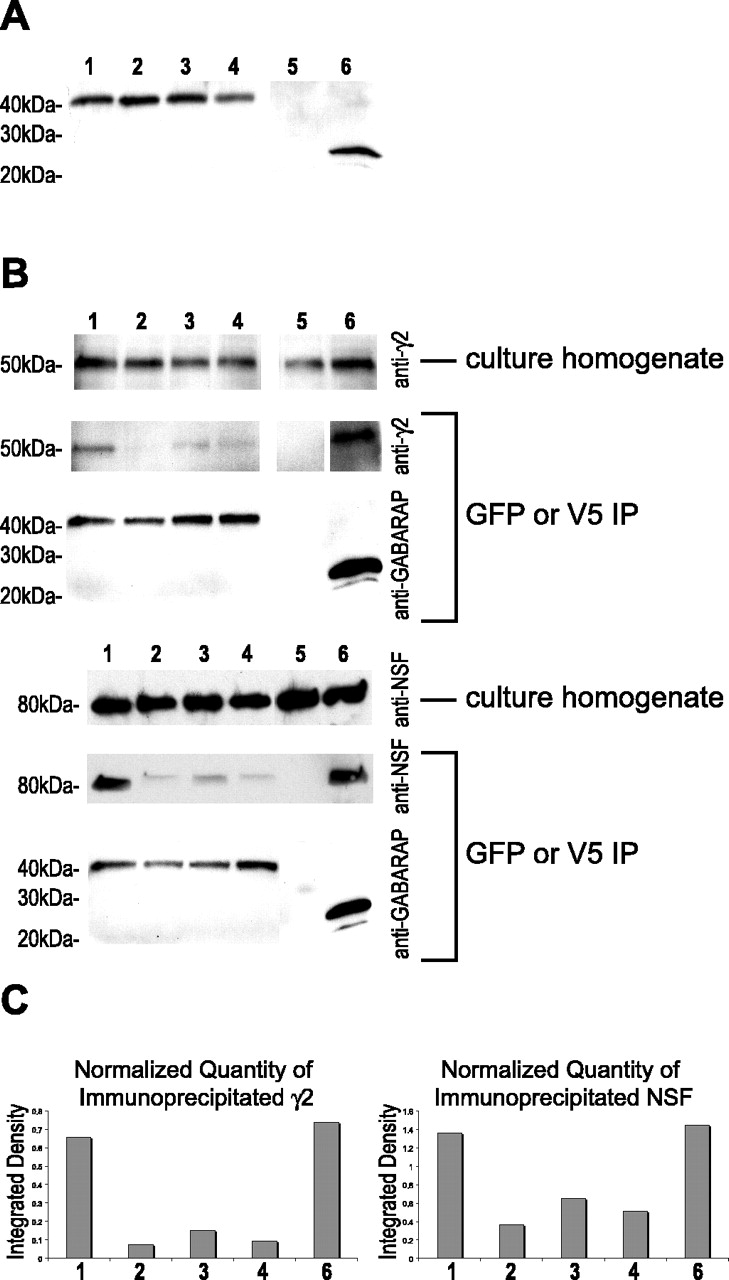
Mutations in the γ2-binding domain of YFP-GABARAP disrupt the interaction of YFP-GABARAP with γ2 subunit and NSF in cultured hippocampal neurons. A, Adenovirus was used to direct expression of either wild-type YFP-GABARAP or mutated forms of GABARAP or V5-GABARAP in cultured hippocampal neurons. Western blot of homogenates from cultured neurons using antibody to GABARAP revealed the presence of a 42 kDa band corresponding to wild-type YFP-GABARAP (lane 1), P37A mutated YFP-GABARAP (lane 2), K38A mutated YFP-GABARAP (lane 3), K38A/R40A mutated YFP-GABARAP (lane 4), or V5-GABARAP (lane 6). Cells that did not receive any adenovirus (lane 5) did not have this 42 kDa band. B, Immunoprecipitation of YFP-GABARAP using anti-GFP antibody (or V5-GABARAP using V5 antibody) from neurons infected with wild-type or mutated forms of YFP-GABARAP. Western blot of YFP-GABARAP immunoprecipitate using anti-γ2 antibody shows that YFP-GABARAP (or V5-GABARAP) is capable of pulling down γ2 immunoreactivity. Mutations in the γ2-binding domain reduced the ability of YFP-GABARAP to pull down γ2 immunoreactivity. YFP-GABARAP was also able to pull down NSF immunoreactivity. Mutations in the γ2-binding domain also reduced the ability of YFP-GABARAP to pull down NSF immunoreactivity. C, Quantitation of the amount of immunoprecipitated γ2 or NSF by integration of the optical density of the autoradiograph bands from Western blots revealed that the P37A and K38A/R40A mutants were the most disrupted in there ability to pull down either γ2 or NSF immunoreactivity. Quantities of γ2 or NSF were normalized to the amount of GABARAP detected in each lane.
GABARAP has previously been found to immunoprecipitate with NSF when coexpressed in Cos7 cells; however, it has yet to be shown that GABARAP interacts with NSF in neurons (Kittler et al., 2001). Immunoprecipitation with GFP antibody revealed that YFP-GABARAP does interact with endogenous NSF in hippocampal neuron cultures (Fig. 6B). V5-GABARAP also pulled down NSF immunoreactivity from these culture homogenates at quantitatively similar levels to YFP-GABARAP (Fig. 6C), suggesting that YFP did not interfere with the interaction between GABARAP and NSF under these conditions. Interestingly, similar to their interaction with γ2, all of the mutated forms of YFP-GABARAP were also altered in their interaction with NSF (Fig. 6C).
Mutations in the γ2-binding domain disrupt colocalization of YFP-GABARAP with γ2 and NSF
Confocal microscopy of neurons that had been labeled with γ2 and NSF antibodies revealed that colocalization of the P37A and K38A/R40A mutants of YFP-GABARAP were disrupted in their colocalization with both of these proteins (Fig. 7). The fluorescence of the P37A mutant appeared to be primarily in the perinuclear region, with a clumpy pattern of distribution, perhaps causing the uneven colocalization with γ2 and NSF (Fig. 7A). The fluorescence from the K38A/R40A double mutant appeared to be more random in its distribution, with less apparent perinuclear localization (Fig. 7B). Colocalization of the double mutant with γ2 and NSF immunoreactivity was much less apparent than with the wild-type YFP-GABARAP. This was most apparent in the neuron on the right in Figure 7B, where YFP-GABARAP fluorescence was found primarily at puncta that lack γ2 and NSF immunoreactivity. This neuron also showed an apparent reduction in NSF immunoreactivity, perhaps because of the high expression of this mutated form of YFP-GABARAP.

YFP-GABARAP mutants display altered colocalization. A, P37A mutated YFP-GABARAP adenovirus was used to infect cultured hippocampal neurons, then cells were fixed, permeabilized, and coimmunostained using antibodies to the γ2 subunit (red) and NSF (blue). The top images show a single three-color confocal z-plane of a hippocampal neuron expressing P37A mutated YFP-GABARAP (green), and the bottom images show a 2.5× zoom of one of the dendrites. The P37A mutant of YFP-GABARAP displays altered colocalization with NSF immunofluorescence and γ2 immunofluorescence. B, A cultured hippocampal neuron expressing the K38A/R40A double mutant of YFP-GABARAP (green) also shows altered colocalization with NSF (blue) and γ2 (red). The top images show a single three-color confocal z-plane of a hippocampal neuron, and the bottom images show a 2.5× zoom of one of the dendrites. Scale bars, 20 μm.
Pearson correlation coefficients were calculated for the colocalization of the mutated forms of YFP-GABARAP fluorescence with NSF or γ2 immunoreactivity in confocal images (Table 1). Among the mutated forms of YFP-GABARAP, only the K38A mutant showed a slightly significant correlation with γ2 (t test; p = 0.043), whereas the correlation of the other two mutants with γ2 was not significant, indicating that these mutations abolish the colocalization of YFP-GABARAP with γ2 (t test; P37A, p = 0.094; K38A/R40A, p = 0.235). The Pearson correlation for the colocalization of the P37A mutant with NSF was reduced compared with wild-type YFP-GABARAP, but still significant (t test; p = 0.030), whereas the colocalization of the K38A/R40A double mutants was not significant, indicating that this mutation also abolished the colocalization of YFP-GABARAP with NSF (t test; p = 0.096). In addition to the change in colocalization of YFP-GABARAP with γ2 and NSF, there may also be a change in the pattern of expression for the mutants. This is particularly true for the P37A mutation, which is less even in its distribution (Fig. 7A).
Mutations in the γ2-binding domain disrupt ability of YFP-GABARAP to increase surface expression of GABAA receptors
The mutated forms of YFP-GABARAP were compared with wild type as to their ability to increase surface expression of CFPγ2 subunit-containing GABAA receptors in Cos7 cells (Fig. 8A). At the lower level of expression, wild-type YFP-GABARAP showed a significantly greater increase in surface GABAA receptor expression when compared with all of the mutated forms of YFP-GABARAP (one-way ANOVA; WT vs P37A, p < 0.0001; WT vs K38A, p = 0.0004; WT vs K38A/R40A, p < 0.0001). At the higher levels of expression, the situation changed slightly: YFP-GABARAP displayed a significantly greater increase in surface GABAA receptor expression when compared with the P37A mutant (one-way ANOVA; WT vs P37A, p = 0.044) and the double mutants (one-way ANOVA; WT vs K38A/R40A, p = 0.009) but not when compared with the K38A mutant (one-way ANOVA; WT vs K38A, p = 0.59).

Mutations in the γ2-binding domain of YFP-GABARAP disrupt ability to increase surface receptor level. A, Flow cytometry was used to measure the level of GABAA receptors that reach the surface in Cos7 cells expressing the mutated forms of YFP-GABARAP. The γ2-binding mutants P37A, K38A, and K38A/R40A are significantly reduced in their ability to increase surface levels of GABAA receptors in Cos7 cells compared with wild-type YFP-GABARAP. At low expression levels (bottom 50% of cells with respect to YFP-GABARAP expression), the difference between wild type and mutants is more pronounced than at the high level of expression (top 50% of cells with respect to YFP-GABARAP expression). The y-axis shows the percentage of increase in γ2 subunit-containing GABAA receptors compared with cells that do not express YFP-GABARAP. B, Cultured hippocampal neurons infected with adenovirus to express wither wild-type or mutated YFP-GABARAP revealed that mutation of the γ2-binding domain drastically reduces the ability of YFP-GABARAP to increase surface levels of endogenous γ2 subunit-containing GABAA receptors. The y-axis shows γ2 surface fluorescence after subtraction of fluorescence from the noninfected neurons.
Similar results effects were seen on endogenous γ2 subunit-containing GABAA receptors when the mutated forms of YFP-GABARAP were expressed in cultured hippocampal neurons (Fig. 8B). All of the mutated forms of YFP-GABARAP were significantly reduced in their ability to increase surface expression of endogenous GABAA receptors when compared with wild type (one-way ANOVA; WT vs P37A, p < 0.0001; WT vs K38A/R40A, p < 0.0001; WT vs K38A, p = 0.002). The mutated forms of YFP-GABARAP did not show any significant difference from the noninfected control; however, the K38A mutant was nearly significant when compared with the double mutant K38A/R40A (one-way ANOVA; p = 0.083). Similar to the results with the Cos7 cells, wild-type YFP-GABARAP was able to significantly increase the surface levels of GABAA receptors in neurons, whereas the mutants P37A and K38A/R40A were dramatically reduced in this effect.
Discussion
In this study, YFP-GABARAP was expressed in cultured hippocampal neurons to compare its localization with γ2, NSF, and other markers of neuronal function. GABARAP has been shown to be expressed at high levels in the hippocampus (Okazaki et al., 2000), so cultured hippocampal neurons provide a good model system in which to study its function. Electron microscopy has revealed that GABARAP is found on Golgi membranes and in postsynaptic cisternae (Kittler et al., 2001), which is consistent with the cytoplasmic, perinuclear pattern of fluorescence seen with YFP-GABARAP.
Lack of colocalization between YFP-GABARAP and surface GABAA receptors confirms previous findings that GABARAP is not anchoring receptors at the surface (Kneussel et al., 2000). However, YFP-GABARAP fluorescence was found in the dendrites of neurons making synaptic contact (Fig. 3A) and was found beneath GAD65-labeled presynaptic nerve terminals (Fig. 3B). This is consistent with a role for GABARAP in the trafficking of γ2 subunit-containing GABAA receptors to the plasma membrane, and perhaps to synapses.
YFP-GABARAP colocalizes with γ2 subunit-containing GABAA receptors primarily in a cytoplasmic, perinuclear region in the cell body of most neurons (Figs. 1A, 2B), but YFP-GABARAP puncta can also be found in the proximal regions of some the dendrites in neurons. This excludes the possibility that GABARAP is restricted to the Golgi, like its homolog GATE-16, because the cis- and trans-Golgi compartments of hippocampal neurons are normally restricted to the cell body (Krijnse-Locker et al., 1995). This allows us to differentiate between the functions of GABARAP and GATE-16 (GABARAP L2), which also binds to NSF but does not bind to GABAA receptors or gephyrin (Kneussel et al., 2000; Sagiv et al., 2000). Although both GABARAP and GATE-16 are highly expressed in the brain (Xin et al., 2001), GABARAP appears to function in the transport of vesicles from the trans-Golgi to the plasma membrane, and GATE-16 functions in the transport of vesicles within the Golgi complex. GATE-16 appears to be Golgi restricted and participates with NSF by enhancing its ATPase activity, causing the disassembly of the vesicle-docking apparatus and allowing intra-Golgi vesicles to fuse with the Golgi membrane (Elazar et al., 2003). It is unclear whether GABARAP has any influence on the ATPase activity of NSF, but if it does, this may also have an effect on the ability of GABAA receptor-containing exocytotic vesicles to fuse with the plasma membrane.
Colocalization of YFP-GABARAP with NSF (Fig. 2C) implies that NSF plays an important role in the function of GABARAP. Perhaps NSF plays an analogous role with GABARAP, as it does with GATE-16, in an ATPase-dependent fusion of GABARAP-laden vesicles as they reach the plasma membrane from the trans-Golgi network (TGN). This process may also require vesicle (v)- and target (t)-soluble NSF attachment protein (SNAP) receptors (SNAREs) as with GATE-16, but occurring at the plasma membrane rather than in the Golgi. Because SNARE proteins are involved in most intracellular vesicle fusion events (Rothman, 1994), it is likely that they also play a role in docking and fusion of GABAA receptor-containing vesicles coated with GABARAP. In this model, the t-SNARE would be located on the plasma membrane, whereas the v-SNARE is located on the transport vesicle coated with GABARAP and GABAA receptors. GABARAP may recruit NSF to these vesicles to mediate the ATP-dependent fusion of the transport vesicle with the plasma membrane. The C-terminal lipidation of GABARAP may allow it to coat the transport vesicle, just as lipidation of the GABARAP homolog LC3 results in it coating the surface of autophagosomes during induction of starvation (Mizushima et al., 2004). This may allow GABARAP to transport not only GABAA receptors but other proteins that are required for GABAA receptor function, such as gephyrin. The importance of gephyrin for anchoring GABAA receptors at the plasma membrane is illustrated by the gephyrin knock-out, in which γ2 subunit-containing GABAA receptor clusters at the plasma membrane are reduced or lost (Kneussel et al., 1999). GABARAP is known to interact with gephyrin and causes its translocation to the plasma membrane in PC12 cells (Kneussel et al., 1999). Consistent with previously published reports (Kneussel et al., 2000), YFP-GABARAP was found to colocalize with gephyrin primarily in the intracellular region of the cell (Fig. 3C). This implies that GABARAP does not provide a link between gephyrin and GABAA receptors at the plasma membrane but may be involved in the trafficking of gephyrin. Because gephyrin is required for anchoring glycine receptors at the plasma membrane and appears to play a role in their trafficking to the plasma membrane as well (Hanus et al., 2004), it is possible that gephyrin is involved in the trafficking of GABAA receptors through its interaction with GABARAP.
Overexpression of YFP-GABARAP results in an increase in the level of GABAA receptors found at the surface of Cos7 cells and hippocampal neurons. It is possible that increasing the level of YFP-GABARAP expressed in cells induces an increase in the formation of vesicles that transport GABAA receptors from the Golgi to the cell surface. This may be the result of an increase in the amount of the lipidated form of GABARAP that can coat the surface of transport vesicles that are targeted to the plasma membrane. Although it is possible that the effect of YFP-GABARAP on surface GABAA receptor levels is attributable to a dominant-negative effect, it seems unlikely because the levels of surface receptors were increased even at the lowest levels of YFP-GABARAP expression. A dominant-negative effect of YFP-GABARAP overexpression would implicate GABARAP in the internalization and degradation of GABAA receptors, which could be ruled out by repeating the experiments on Cos7 cells in the presence of agents that block receptor internalization.
It is interesting that mutations in the γ2-binding domain of YFP-GABARAP did not completely abolish the effect of GABARAP on receptor surface expression. Different mutations were able to have differing effects on the transport of receptors to the cell surface. For instance, mutation of lysine 38 appeared to have the least effect and was not significantly different from wild type in its ability to transport receptors to the surface at the high levels of expression in Cos7 cells. In both Cos7 cells and hippocampal neurons, the mutations that had the greatest effect in abolishing the ability of YFP-GABARAP to traffic receptors to the cell surface were the P37A and the K38A/R40A double mutant. This is consistent with the results from the coimmunoprecipitation experiments that showed that the K38A mutation had the strongest interaction of the mutants, whereas the P37A and K38A/R40A mutants had the weakest interaction with the γ2 subunit. Even these mutants were able to have a significant effect on GABAA receptor surface levels at high expression levels, suggesting that these mutations result in a reduction of the affinity of GABARAP for the receptor rather than causing global disruption of protein structure.
The region of GABARAP selected for mutagenesis in this study has also been implicated in dimerization of GABARAP and in the formation of GABARAP oligomers (Coyle et al., 2002; Nymann-Andersen et al., 2002a). It is possible that this domain is responsible for the binding of GABARAP to many of its protein partners, including γ2 and NSF. Mutation of proline 37 within this domain appeared to have a major influence, not only on the interaction between GABARAP and γ2, but also on the interaction between GABARAP and NSF. Previous studies have demonstrated that GABARAP 36-117 is nearly four times higher in its interaction with the γ2 intracellular loop in a yeast two-hybrid assay compared with GABARAP 41-117, suggesting that the amino acids from 36 to 41 are the most important for the interaction (Nymann-Andersen et al., 2002b). Therefore, it is not surprising that the double mutant of YFP-GABARAP, K38A/R40A, would be disrupted in its ability to bind the γ2 subunit. It is slightly surprising that these amino acids would be disrupted in their ability to interact with NSF, because GATE-16 is capable of interacting with NSF, and it does not have these amino acids. It is possible that GABARAP uses these amino acids, in addition to those that are conserved with GATE-16, in its interaction with NSF.
In conclusion, this study has demonstrated an important functional role for GABARAP in the trafficking of GABAA receptors in neurons. A new model for GABARAP function in neurons can be proposed based on the results of this study. One can envision GABARAP associating with newly synthesized GABAA receptors in transport vesicles budding from the TGN and traveling along microtubules to the cell surface. At some point, GABARAP is converted to the lipid-conjugated form to increase membrane association, and gephyrin is recruited, possibly for anchoring of surface receptors. Finally, GABARAP recruits NSF to the vesicle to initiates fusion of GABAA receptor-containing vesicles with the plasma membrane with the help of the SNAP-SNARE complex. In the future, the role of NSF in this process should be clarified. For instance, the influence of GABARAP binding to NSF on activation of its ATPase activity, and whether this has an influence on the ability of GABARAP to transport GABAA receptors to the cell surface, can be examined.
Footnotes
This work was supported by National Institutes of Health Grant NS28772.
Correspondence should be addressed to Dr. Richard W. Olsen, Department of Molecular and Medical Pharmacology, Geffen School of Medicine, University of California Los Angeles, Room CHS 23-120, 650 Young Drive South, Los Angeles, CA 90095-1735. E-mail: rolsen{at}mednet.ucla.edu.
Copyright © 2004 Society for Neuroscience 0270-6474/04/2511429-10$15.00/0





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}