

Abstract
The rhombic lip (RL) is the neuroepithelium immediately adjacent to the roof plate of the fourth ventricle, and it gives rise to various brainstem and cerebellar cell types. Our study shows that the bHLH (basic helix-loop-helix) transcription factor Olig3 is expressed in the progenitors of RL, and ablation of Olig3 significantly affects the development of RL. In Olig3−/− caudal RL, the expression level of Math1 in the dorsal interneuron 1 (dI1) domain is reduced, and the formation of four mossy-fiber nuclei is compromised; dI2–dI3 neurons are misspecified to dI4 interneurons, and the climbing-fiber neurons (inferior olive nucleus) are completely lost. In addition, the formation of brainstem (nor)adrenergic centers and first-order relay visceral sensory neurons is also dependent on Olig3. Therefore, Olig3 plays an important role in the fate specification and differentiation of caudal RL-derived neurons.
Introduction
The rhombic lip (RL) is a specific germinal matrix that is located in the dorsal hindbrain neuroepithelium between the roof plate of fourth ventricle and neural tube. Along the anterior–posterior axis of the rhombomeres, the RL is divided into rostral and caudal parts (Lumsden and Krumlauf, 1996). The rostral RL (rRL) is related to the neuroepithelium of rhombomere 1 (r1) and specifically expresses basic helix-loop-helix (bHLH) transcription factor Math1. The neurons derived from rRL migrate along the rostral rhombic-lip migratory stream and sequentially populate a series of nuclei within the rostral hindbrain and cerebellum, of which most are Math1 dependent (Machold and Fishell, 2005; Wang et al., 2005). In cerebellum, rRL gives rise to glutamatergic external granule layer (EGL) cells, large deep cerebellar nuclei (DCN) neurons, and unipolar brush cells.
The caudal RL (cRL) (r2–r5) comprises the auditory lip and gives rise to the cochlear extramural migratory stream (EMS) to form the cochlear nuclei (CN) (Farago et al., 2006). The generation of ventral CN and cochlear granule neurons requires Math1 expression (Wang et al., 2005). The more caudal cRL (r6–r8) produce the precerebellar mossy-fiber (MF) nuclei and climbing-fiber (CF) nuclei (Ambrosiani et al., 1996; Cambronero and Puelles, 2000). There are four major MF nuclei that project mossy fibers to the EGL and DCN, including the pontine gray nuclei (PGN), reticulotegmental nuclei (RTN), lateral reticular nuclei (LRN), and external cuneate nuclei (ECN). The PGN and RTN migrate in the anterior precerebellar EMS (AES) and settle in the rostral-ventral region of the pons, whereas the LRN and ECN migrate in the posterior precerebellar EMS (PES) and settle in the medial-dorsal and lateral-ventral region of medulla, respectively (Altman and Bayer, 1987a,b,c,d). Loss of Math1 completely inhibits the formation of AES and PES and the generation of four MF nuclei (Wang et al., 2005). In contrast, the inferior olive nucleus (ION) projects climbing fibers to the Purkinje cells, forming the olivocerebellar circuit (Altman and Bayer, 1987a,b,c,d; Sotelo, 2004). The ION migrates from the cRL via the precerebellar intramural migratory stream (IMS) and settles in the medial-ventral region of the medulla (Altman and Bayer, 1987a,b,c,d). The generation of ION is Math1 independent, but the migration of CF neurons requires Ptf1a, which is expressed ventral to the Math1 domain (Yamada et al., 2007).
Previous studies showed that Olig3 bHLH transcription factor was expressed in the RL (Takebayashi et al., 2002). In this study, we showed that Olig3 was specifically expressed in dorsal progenitor 1 (dp1)–dp3 domains of neural progenitor cells in the caudal hindbrain (r6–r8) and regulated the fates of these neural progenitor cells. In Olig3 mutant hindbrain, the cell fate of dp2 and dp3 progenitors was changed to that of the more ventral interneuron progenitors. The generation of four MF nuclei was markedly reduced. More strikingly, ION CF neurons were completely lost, providing evidence that these neurons may arise from the ventral dorsal interneuron 3 (dI3) domain, which is dependent on both Olig3 and Ptf1a. In addition, development of dorsal Phox2b+ neurons, brainstem (nor)adrenergic centers, and first-order relay visceral sensory neurons was also disrupted.
Materials and Methods
Generation of Olig3 knock-out mice.
A BAC clone containing Olig3 genomic DNA was purchased from Invitrogen. The Olig3 targeting vector contained a 4 kb 5′ arm and a 3.5 kb 3′ arm at the two ends of neo cassette. Targeting vector was linearized and electroporated into mouse embryonic stem (ES) cells. G418-resistant cells were selected. Genomic DNA from drug-resistant cells was digested with enzyme and analyzed by Southern hybridization using 5′ or 3′ probe for Olig3. Chimeric mice and F1 heterozygous mice were generated from Olig3 mutant ES cell lines. Germline transmission of the target allele was confirmed by both Southern analysis and PCR.
In situ RNA hybridization and immunofluorescent staining.
Brain tissues were isolated from embryonic day 10.5 (E10.5)–E18.5 mouse embryos and then fixed in 4% paraformaldehyde at 4°C overnight. After fixation, tissues were transferred to 20% sucrose in PBS overnight, embedded in OCT media, and then sectioned (20 μm thickness) on a cryostat. Sections at similar positions from the wild-type and mutant embryos were subsequently subjected to in situ hybridization (ISH) or immunofluorescent staining. ISH was performed as described by Schaeren-Wiemers and Gerfin-Moser (1993) with minor modifications. cDNA templates for ISH were obtained either by reverse transcription-PCR of postnatal day 0 brain tissues or from a commercial source (Invitrogen), and confirmed by sequencing.
Immunofluorescent staining was performed as follows. After rinsing with PBS, sections were permeabilized in 0.1% Triton X-100 in PBS for 10 min, rinsed with PBS to remove the excessive Triton, incubated in blocking solution (5% normal serum in PBS plus 1% BSA) at room temperature for 1 h, and incubated in diluted primary antibody in blocking solution at 4°C overnight. On the next day, sections were washed in PBS three times for 10 min each, and incubated with the secondary antibody in blocking solution at room temperature for 1 h. After incubation, sections were washed in PBS three times for 10 min each, before they were mounted in Mowiol mounting medium on glass slides. The staining was examined under a Nikon fluorescence microscope. Rat polyclonal antibody anti-Olig3 was generously provided by Dr. Hirohide Takebayashi. Rabbit polyclonal antibody anti-Ngn1 was kindly provided by Dr. Qiufu Ma. Guinea-pig polyclonal antibody anti-Ptf1a was kindly provided by Dr. Jane Johnson. Rabbit polyclonal antibody anti-Ptf1a was kindly provided by Dr. Helena Edlund. Anti-Brn3a (1:200; Millipore Bioscience Research Reagents), anti-Mash1 (1:400; BD Biosciences), and anti-Pax2 (1:200; Zymed) were obtained from commercial sources. Guinea-pig polyclonal antibody anti-Lbx1 and rabbit polyclonal antibody anti-Tlx3 were kindly provided by Dr. Carmen Birchmeier. The Alexa-488- or Alexa-594-conjugated secondary antibodies were obtained from Invitrogen. The nucleic acid dye DAPI (4′,6-diamidine-2′-phenylindole dihydrochloride) was obtained from Roche.
Olivocerebellar projection tracing with DiI.
Brain tissues were isolated from E18.5 mouse embryos and fixed in 4% paraformaldehyde. Single or multiple small crystals of DiI were placed in one side of the cerebellar hemisphere far from the midline in E18.5 wild-type or Olig3−/− animals. Tissues were stored in 4% paraformaldehyde in dark at 37°C. After 5–6 weeks for DiI diffusion, tissues were transferred to 20% sucrose in PBS overnight, embedded in OCT media, and then sectioned (30 μm thickness) on a cryostat. The DiI staining was examined under a Nikon fluorescence microscope.
Results
Olig3 expression pattern in the rhombic lip
It has been well documented that several nuclei in the hindbrain originate from the rhombic lip. A previous report and our present studies demonstrated that Olig3 is expressed in the rhombic lip in the entire hindbrain, isthmus area, and the subventricular zone of cerebellum and pons (Fig. 1A,B) (Takebayashi et al., 2002). To investigate the function of Olig3 in the development of hindbrain, we performed detailed studies on the expression pattern of Olig3 in cRL at early embryonic stages by comparing Olig3 expression with various specific interneuron markers. Similar to that in the dorsal spinal cord, the dorsal neuroepithelium in the caudal hindbrain could be divided into several domains along the dorsal–ventral axis based on the expression of multiple interneuron markers (Sieber et al., 2007). At early stages of neural development, Math1 and Ngn1 specifically label the progenitor pool of dp1 and dp2, respectively, whereas Tlx3 marks dI3 and dI5 interneurons (Gowan et al., 2001; Qian et al., 2002; Sieber et al., 2007). Because the dorsal Ngn1 expression was only detected in cRL at r7–r8 level, which lies between the otic vesicle and cervical spinal cord (Landsberg et al., 2005), we used Ngn1 as the rostral–caudal boundary marker. At E10.5, Olig3 marked the dorsalmost domains (dp1–dp3) of neuroepithelium in the caudal hindbrain and was colabeled with Math1, Ngn1, and Tlx3, respectively (Fig. 1C–E,G,H). There was no Olig3 expression detected in dp4–dp6 domains, from which Pax2-labeled dI4 and dI6 neurons arose (Fig. 1I) (Helms and Johnson, 2003; Mizuguchi et al., 2006). There was a partially overlapping region between Olig3 and Ptf1a in the ventral dp3 domain (Fig. 1C,F,K). The overlapping expression of Olig3 and Ptf1a in the dorsal neuroepithelium became more apparent at E11.5 (Fig. 2A). This overlapping domain appeared to be tightly associated with the generation of Brn3a+ glutamatergic neurons, instead of the Pax2+ GABAergic inhibitory neurons (Fig. 2B,C), in keeping with the recent observation that Brn3a+ CF neurons were labeled by the Ptf1a-Cre fate mapping experiments (Yamada et al., 2007). Olig3 remained to be expressed in the dorsal neural progenitor cells at E12.5, but its expression domain was dorsally retracted (Fig. 2D–F). Together, these studies indicated that Olig3 is selectively expressed in the progenitor pool of the cRL at early stages of neurogenesis, suggesting that Olig3 may be involved in the fate specification and/or migration of the neurons originated from the cRL.

Expression of Olig3 in the mouse hindbrain. A, Unstained bright-field photograph of E12.5 mouse brain sagittal section. B, Olig3 expression in E12.5 sagittal hindbrain corresponding to the box position shown in A. C–F, In situ hybridization of various progenitor TFs in the dorsal hindbrain at E10.5. All transverse sections are oriented with lateral to the left. Olig3 expression region included the expression domain of Math1 (D) and Ngn1 (E). There is partial overlap between Olig3 (C) and Ptf1a (F) expression in dp3 domain. G–K, Double-immunofluorescence analysis of Olig3 expression pattern in E10.5 transverse hindbrain sections at r6–r8 level. Olig3 expression overlapped with Ngn1 (G) and Tlx3 (H), but not with Pax2 (I) in the dorsal hindbrain. Ptf1a was expressed in dp4 and part of dp3 domains (J). Olig3 and Ptf1a were coexpressed in some ventral dp3 progenitors (K, arrows). L, Summary of distinct dp domains and dIs in which the transcription factors were expressed. Cb, Cerebellum; Isth, isthmus. Scale bars: A, 1 mm; B, C (for C–F), G (for G–K), 100 μm.
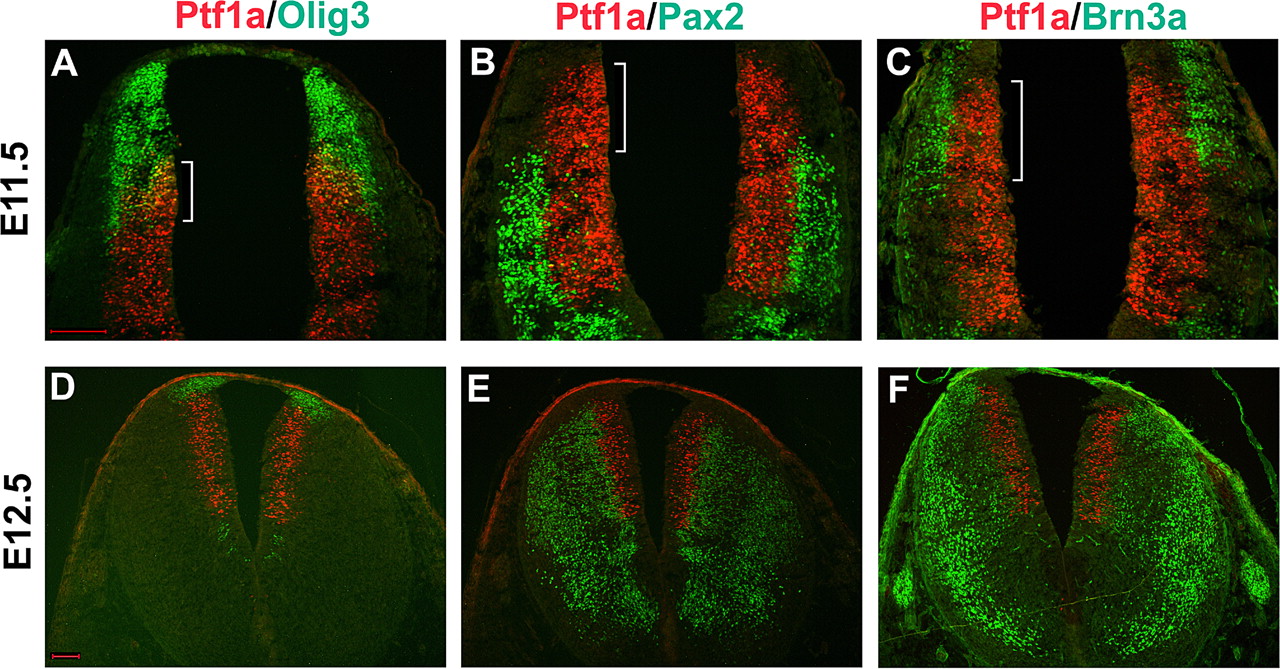
Expression of neural progenitor genes and neuron-specific markers in the dorsal caudal hindbrain. Transverse sections from E11.5 (A–C) and E12.5 (D–F) rhombomere 7 were subject to double immunostaining with various progenitor and subtype-specific markers. At E11.5, there was a considerable overlapping between Olig3 and Ptf1a (indicated by bracket; A), and this overlapping region was associated with the generation of Brn3a+ excitatory neurons (indicated by bracket; C), but not Pax2+ inhibitory neurons (indicated by bracket; B). Scale bars: A (for A–C), D (for D–F), 100 μm.
Generation of Olig3 mutant mice
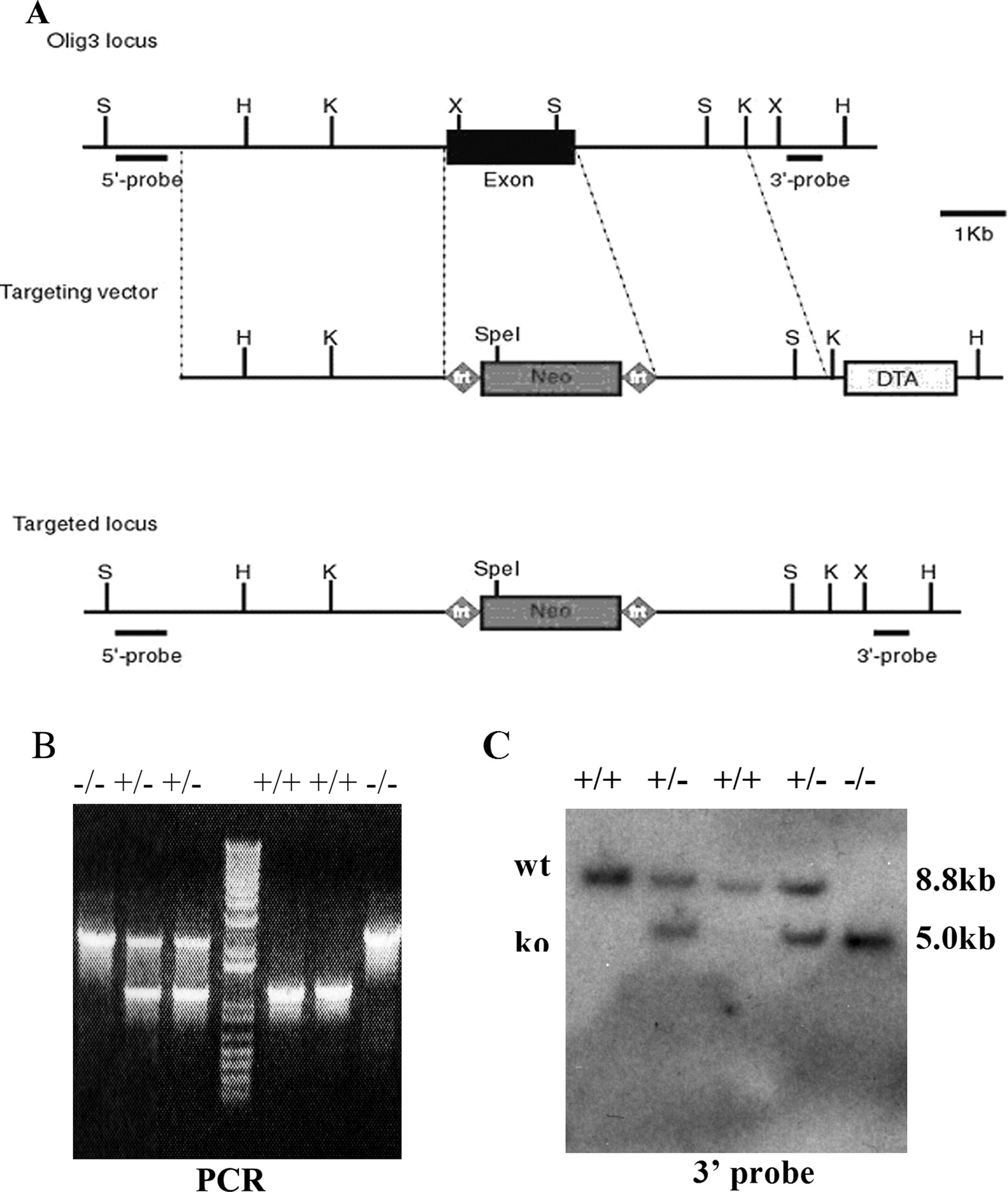
To elucidate the function of Olig3 in hindbrain development, we performed gene targeting of Olig3 by homologous recombination in ES cells. The targeting vector was designed to replace the entire coding region with the neomycin (neo) gene (Fig. 3A). After electroporation and drug selection, the ES clones harboring homologous recombination were identified by genomic Southern blot with the flanking probes (Fig. 3C). These ES cells were injected into blastocysts, and the targeted alleles were transmitted through germ line. Heterozygous mice were born and mated to obtain homozygous mutant mice. As expected, Olig3 expression was not detected in the Olig3−/− mutants (data not shown). Most Olig3 homozygous mutants died soon after birth, and few could stay alive until 12 h. Olig3−/− mice displayed cyanosis caused by respiratory failure, resembling the congenital hypoventilation syndrome in humans, suggesting that there is a defect in the developing respiratory system, which resides in the brainstem (Shirasawa et al., 2000).
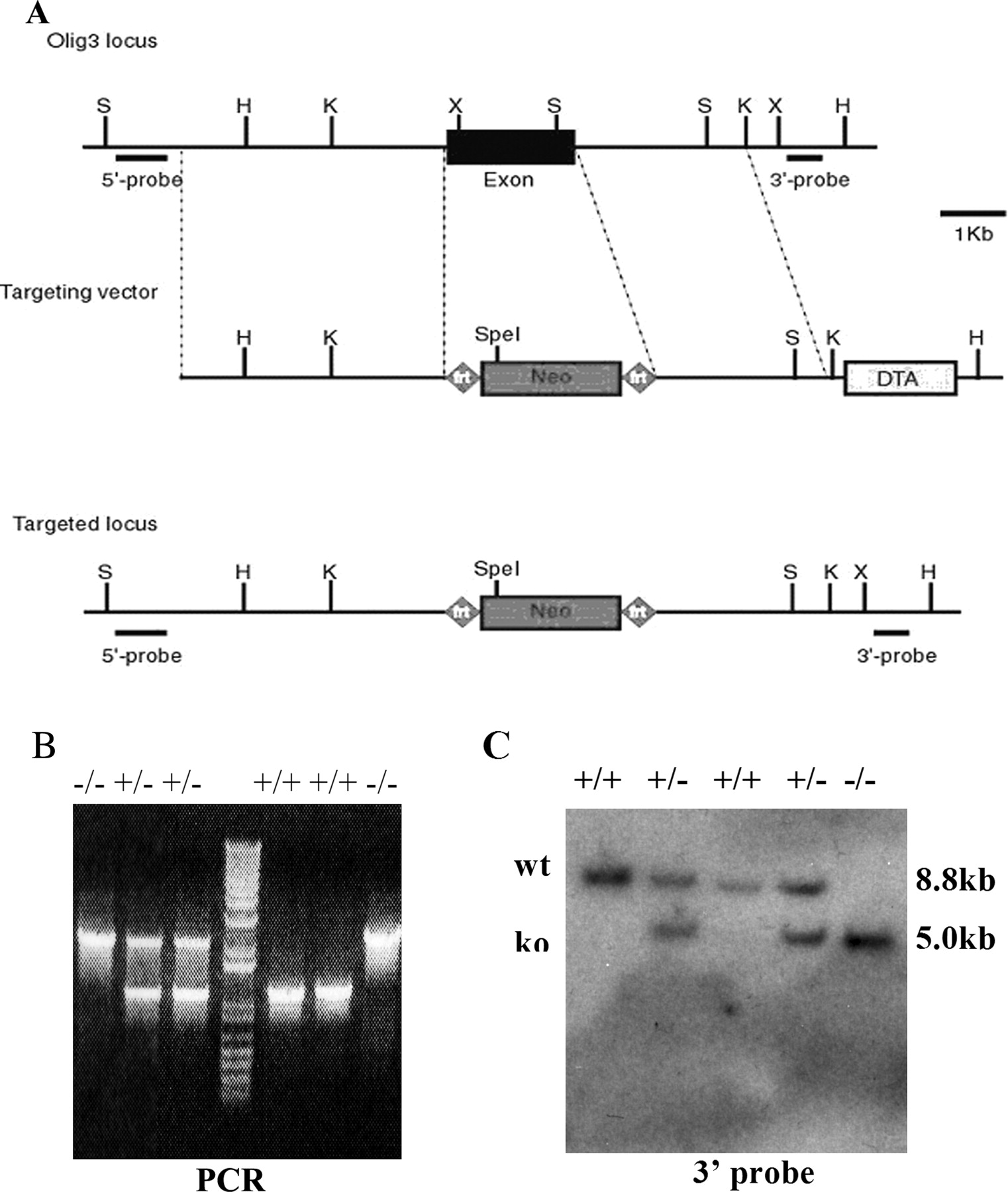
Inactivation of Olig3 by homologous recombination. A, Schematic of the strategy for Olig3 gene targeting. The wild-type Olig3 locus, targeting vector, and the targeted allele after homologous recombination are shown. 5′ and 3′ probes are used for Southern blot analysis. Neo, Neomycin positive selection marker cassette; DTA, diphtheria toxin gene negative selection marker. B, PCR analysis of genomic DNA extracted from wild-type, Olig3+/−, or Olig3−/− tails. C, Genotyping by Southern blot analyses. Genomic DNA was digested with HindIII and SpeI and hybridized with the 3′ probe. The wild-type allele (wt) appeared as a 8.8 kb band and the mutant allele (ko) as a 5.0 kb band.
Olig3 regulates the formation of precerebellar mossy-fiber nuclei
To test the possibility that loss of Olig3 function may affect the development of RL-derived neurons, we first examined the effects of Olig3 mutation on the correct establishment of brainstem precerebellar nuclei (PCN). There are five major PCN that arise from rhombomere 6–8 cRL. The ION provides climbing fibers to the cerebellum. The remaining four are the MF precerebellar nuclei (PGN, RTN, LRN, and ECN), tangentially migrating from the cRL in the precerebellar EMS. Transcription factor Barhl1 has been used as a generic marker for migrating and mature MF neurons (Bulfone et al., 2000). Expression analyses on E14.5 hindbrain coronal sections revealed that the expression level of Barh1 in EMS was significantly downregulated in Olig3−/− mutants compared with wild-type tissues (Fig. 4A,B). As a result, the formation of four MF nuclei was markedly inhibited in Olig3−/− hindbrains, as revealed by Barhl1 expression in four discrete MF nuclei in E16.5 and E18.5 tissues (Fig. 4E,F,I,J,M,N). Our results showed that transcription factor Zic1 is also expressed in MF neurons. Compared with wild-type littermates, Olig3−/− embryos showed apparently reduced Zic1 mRNA expression within the area of the EMS and all four MF nuclei, similar to Barhl1 expression (Fig. 4A–P). Together, these findings demonstrated that Olig3 expression was required for the proper formation of both the EMS and the precerebellar MF nuclei.

Olig3 mutation affected the generation of precerebellar mossy-fiber nuclei. A–H, Coronal sections from E14.5 and E16.5 hindbrain tissues were examined by in situ hybridization for expression of Barhl1 (A, B, E, F) and Zic1 (C, D, G, H). Both of these genes were expressed in the migrating and mature mossy-fiber neurons. Loss of Olig3 function significantly downregulated the expression level of these two genes in the EMS and reduced the size of the mature mossy-fiber nuclei, LRN and ECN. I–P, E18.5 sagittal sections with rostral to the left. I, J, M, N, Sections were labeled with Barhl1 probe. K, L, O, P, Sections were stained with Zic1 probe. Olig3-null embryos displayed the reduced size of two other mossy-fiber nuclei, PGN and RTN. Ectopic Zic1 expression was detected in the nTS area (G, H, K, L). Scale bars: A (for A, B), C (for C, D), E (for E, F), G (for G, H), I (for I, J), K (for K, L), M (for M, N), O (for O, P), 100 μm.
Olig3 regulates the formation of the inferior olive nucleus
ION is the precerebellar CF nucleus, emigrating from the cRL via the IMS and settling in the most ventral region of caudal medulla (Altman and Bayer, 1987a,b,c,d). The migrating and mature CF neurons can be detected by the expression of Brn3a (Fedtsova and Turner, 1995; Xiang et al., 1996). At E14.5, the Brn3a+ CF neurons in ION were detected at the medulla in the wild-type tissues (Fig. 5A), but not in Olig3−/− mutants (Fig. 5B). The loss of CF neurons in Olig3-deficient mutants was further confirmed by the lack of expression of Rph3a (Fig. 5C,D) and Fgfr2 (data not shown), two other markers for ION neurons (Belluardo et al., 1997; Foletti and Scheller, 2001). The absence of Brn3a and Rph3a expression in Olig3−/− medulla persisted at E16.5 (Fig. 5E–H) and E18.5 (data not shown), indicating that the ION defect was not simply a developmental delay. The loss of ION structure was further confirmed by the lack of Nissl staining in the ION region of the Olig3 mutants (supplemental Fig. S1, available at www.jneurosci.org as supplemental material).

The Olig3−/− mutant lost the precerebellar climbing-fiber neurons. A–H, E14.5 (A–D) and E16.5 (E–H) coronal medulla sections were subjected to immunostaining with anti-Brn3a antibody (A, B, E, F) or in situ hybridization with Rph3a probe (C, D, G, H). Both genes were expressed in the climbing-fiber neurons. ION comprised of climbing-fiber neurons was lost in Olig3-null medulla. I–L, DiI labeling was used to examine the formation of the olivocerebellar projection in E18.5 tissues. DiI crystal was placed into one side of wild-type cerebellum, and projections (arrow) to the contralateral ION (circled area) were detected (I, K). However, the formation of the DiI labeling olivocerebellar projection was not detected in the Olig3−/− mutant (J, L). In addition, DiI labeling of LRN neurons was drastically reduced as well in the mutant (small circled area; I, J). The dashed boxes in A–J outline the ION area. K, L, High-magnification images of I and J. Scale bars: A (for A, B, E, F), C (for C, D, G, H), 100 μm; I (for I, J), 200 μm; K (for K, L), 50 μm.
Because no ectopic Brn3a+ CF neurons were found in Olig3−/− medulla, we proposed that ION neurons fail to form possibly as a result of the loss of CF progenitor pool, instead of their incorrect migration route. To examine whether an ectopic olivocerebellar projection existed in Olig3 homozygote mutants, we performed retrograde labeling of axon fibers in the medulla by placing DiI crystals into one side of E18.5 cerebellum and culturing them for 5 weeks in the dark at 37°C. In the wild-type brains, Purkinje cells incubated with DiI always received the climbing fibers from the contralateral ION (Fig. 5I,K). However, no DiI labeling of climbing fibers and cell bodies was observed in Olig3 mutant brain (Fig. 5J,L), consistent with the idea of the loss of the olivocerebellar circuit. The DiI labeling of the mossy fiber nucleus LRN was detected in the wild-type embryos, but was almost completely lost in Olig3−/− medulla (Fig. 5I–L), in keeping with the dramatic reduction of expression of LRN marker Zic1 (Fig. 4G,H). Based on these results, we concluded that Olig3 expression was indispensable for the generation of CF neurons and LRN mossy fiber neurons.
Olig3 regulates cell fate specification of dp1–dp3
Previous studies showed that the formation of MF nuclei was dependent on Math1, but the migration of CF neurons was regulated by Ptf1a in cRL (Wang et al., 2005; Yamada et al., 2007). Because Olig3 is expressed in the Math1+, Ngn1+, and dorsal part of Ptf1a+ domains in cRL, it is possible that Olig3 may control the formation of MF nuclei and CF nucleus by regulating the expression of Math1, Ngn1, and Ptf1a in the dorsal neural progenitor cells. Consistent with this idea, our expression studies in E10.5 hindbrains revealed a modest reduction of Math1 (we counted 42 ± 4 and 28 ± 6 Math1+ progenitors in control and Olig3−/−, respectively) and Barhl1 expression in dp1 domain (Fig. 6A–D), and a nearly complete loss of dorsal Ngn1 expression in the dp2 domain, although the ventral Ngn1 expression was not affected (Figs. 6E,F, 7E,F). The expression of Lmx1a resides immediately dorsal to Math1 and marks the roof plate. Lmx1a expression was similar in control and Olig3−/− embryos (Fig. 6K,L). Interestingly, the expression pattern of Neo in Olig3−/− mutants was nearly identical to that of the endogenous Olig3 gene in the wild-type embryos (Fig. 6G,H), indicating that Olig3 mutation did not affect the initial allocation and survival of neural progenitors in RL.

Altered gene expression in cRL of Olig3-null embryos. A–L, Hindbrain sections from E10.5 embryos at the cRL level were subjected to in situ hybridization with riboprobes for Math1 (A, B), Barhl1 (C, D), Ngn1 (E, F), Olig3 (G), Neo (H), Ptf1a (I, J), and Lmx1a (K, L). Loss of Olig3 significantly downregulated the expression level of Math1, Ngn1, and Barhl1, whereas Ptf1a expression was ectopically expanded into dp2–dp3 domains. The expression pattern of Neo in Olig3 mutant was the same as that of Olig3 in wild-type controls (G, H). K, L, Arrows indicate Lmx1a expression in the roof plate. Scale bar, 100 μm.
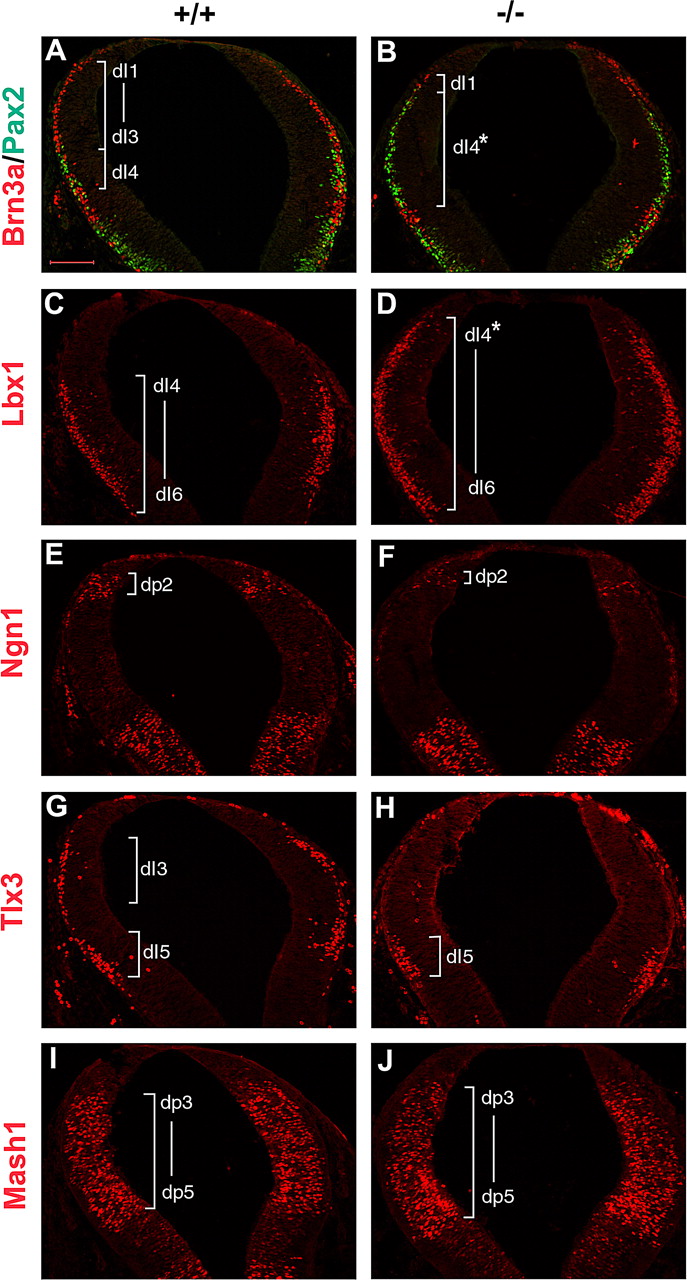
Olig3 is important for the specification of cRL. A–J, Coronal sections of E10.5 cRL were assessed by immunostaining with Brn3a and Pax2 (A, B), Lbx1 (C, D), Ngn1 (E, F), Tlx3 (G, H), and Mash1 (I, J). The Olig3−/− mutant lost the expression of Brn3a in dI2–dI3 neurons and Tlx3 in dI3 neurons, and had a reduced number of Ngn1+ progenitor cells in dp2 domain. Ectopic expression of Pax2 and Lbx1 was detected at the positions of dI2–dI3 neurons. B, D, dI4* indicates ectopic dI4 interneurons derived from dp2 and dp3 domains in Olig3−/− hindbrain. Expression of Mash1 in the dp3–dp5 domains was not altered. Scale bar, 100 μm.
Remarkably, Ptf1a was upregulated and expanded dorsally into the dorsal dp3 and dp2 domains in Olig3-null hindbrain (Fig. 6I,J). Ectopic expression of Pax2 (dI4 and dI6 marker) and Lbx1 (dI4–dI6 marker) was also detected at the positions of dI2 and dI3 interneurons (Fig. 7A–D). The dorsal expansion of Ptf1a and Pax2 expression was accompanied by the reduced expression of Brn3a in dI1–dI3 dorsal interneurons and the absence of Brn3a+ IMS in the Olig3−/− hindbrain (Fig. 7A,B; supplemental Fig. S2, available at www.jneurosci.org as supplemental material). In contrast, Brn3a+ dI5 interneurons persisted in the mutant embryos. In addition, Olig3 mutation resulted in the loss of Tlx3+ dI3 interneurons, but not Tlx3+ dI5 neurons (Fig. 7G,H). Expression of Mash1 (dp3–dp5 marker) was not affected by loss of Olig3 (Fig. 7I,J). Together, these observations indicated that in the absence of Olig3 expression, Brn3a+ dI2 and dI3 neurons were respecified to Pax2+ dI4 neurons. In other words, whereas the Ptf1a+ domain gives rise to both Brn3a+ and Pax2+ neurons in wild-type mice, the expanded Ptf1a+ domain in Olig3−/− mice only produces Pax2+ neurons.
Olig3 is required for proper formation of the brainstem (nor)adrenergic centers and visceral sensory neurons
Both relay visceral sensory and (nor)adrenergic neurons in the brainstem play crucial roles in regulating respiration pattern and cardiovascular activities, and these two classes of neurons originate from dp3 domain (Qian et al., 2001, 2002). Our studies show that Olig3−/− mice die from respiratory failure, suggesting that Olig3 may affect the generation of these two types of neurons. Because Phox2b is essential for formation of brainstem (nor)adrenergic centers and proper development of many visceral sensory neurons (Morin et al., 1997; Pattyn et al., 1999, 2000), we performed a detail examination of Phox2b expression in Olig3-deficient hindbrains. Dorsal Phox2b expression was completely inhibited in E11.5 and E12.5 Olig3−/− mutants, whereas its ventral expression in the dorsal motor nucleus of vagus (dmnX) was normal (Fig. 8A–D). At later stages, Phox2b expression was completely eliminated in the prospective nucleus of the solitary tract (nTS) area in the Olig3−/− hindbrain (Fig. 8E,F,I,J). The first-order relay visceral sensory neurons in the nTS receive signals generated from peripheral chemoreceptors and pulmonary stretch receptors and then influence respiration rates (Blessing, 1997). In addition, the expression level of Phox2b was reduced in the pons as well (Fig. 8I,J). In contrast, Phox2b expression in other brainstem structures was not affected, including the dmnX and nucleus ambiguus (Fig. 8E,F,I,J). Instead, ectopic Zic1 expression was detected at the nTS area in E16.5 and E18.5 Olig3−/− medulla (Figs. 4G,H, 8G,H). To determine the stages when ectopic migration of Zic1+ neurons occurs, we examined Zic1 expression in Olig3 mutant embryos at earlier stages. At E10.5, Zic1 expression was normal in the ventricular zone throughout the entire dorsal hindbrain (data not shown). At E12.5, Zic1+ neurons derived from dp1 and dp4 domains formed two ventrally migratory streams in wild-type embryos, whereas Zic1+ neurons from the entire dp1–dp4 region were able to migrate ventrally in Olig3−/− hindbrains (supplemental Fig. S3A–D, available at www.jneurosci.org as supplemental material), further supporting the notion that dI2–dI3 neurons are misspecified to dI4 interneurons and behave like dI4 neurons. However, misspecified dorsal interneurons still took their migration path to the nTS area, which is normally occupied by Phox2b+ neurons derived from dI3 domain (Fig. 8G,H) (Qian et al., 2001). Consistent with this idea, ectopic expression of Viaat, GABAergic neuron marker, was detected at the nTS area in E18.5 Olig3-deficient embryos (supplemental Fig. S3E–H). This finding is in keeping with the observations that Ptf1a promoted neural precursors to differentiate into GABAergic neurons (Hoshino et al., 2005; Mizuguchi et al., 2006) and was ectopically expressed in dp2 and dp3 domains in Olig3−/− hindbrain (Fig. 6I,J).

Formation of brainstem (nor)adrenergic centers and first-order relay visceral sensory neurons is dependent on Olig3. A–D, Phox2b expression was examined in the transverse sections of E11.5 (A, B) and E12.5 (C, D) caudal hindbrains. Dorsal expression of Phox2b was completely inhibited in Olig3 mutants. E, F, The nTS labeled with Phox2b was lost in E18.5 Olig3 mutants, whereas the dmnX, located ventral to the nTS structure in wild-type embryos (E), still existed in Olig3−/− medulla (F). G, H, Ectopic Zic1 expression was detected in the prospective nTS area in Olig3 mutants. I–L, All E18.5 sagittal sections were oriented with rostral to the left. I, J, Phox2b expression was checked in the sagittal sections. K, L, In Olig3 mutants, expression of TH was absent in the (nor)adrenergic centers of medulla, including A1/C1 and A2/C2. M, N, High-power images of A2/C2 areas. Scale bars: A (for A–D), E (for E–H), I (for I–L), 100 μm.
Our results also showed that loss of Olig3 function affected the formation of (nor)adrenergic centers. The generation of the A1/C1 and A2/C2 clusters in the medulla was completely absent in Olig3−/− brainstems, as indicated by the lack of expression of tyrosine hydroxylase (TH) (Fig. 8K,L). In summary, loss of Olig3 function completely inhibited the generation of Phox2b+ neurons derived from the cRL dp3 domain and disrupted the formation of brainstem (nor)adrenergic centers and first-order relay visceral sensory neurons.
Discussion
In this study, we present evidence that Olig3 is primarily expressed in RL cells in the developing hindbrain and that its expression is important for the fate specification of RL progenitor cells. In the absence of Olig3 function, development of precerebellar neurons derived from the caudal RL is either reduced or completely inhibited. Additionally, Olig3 plays a critical role in regulating the formation of the central respiratory system.
Olig3 is predominantly expressed in RL cells in the hindbrain
The RL was first reported as the germinal zone immediately adjacent to the roof plate of the fourth ventricle through morphological studies (His, 1891). The diversity of neurons in the mature brain is proposed to be generated from this neuroepithelium and to migrate to their destination. Although many attempts have been made, the exact distinction between the RL and the adjacent neuroepithelium still remains unclear. Our studies suggest that Olig3 can be used as a candidate molecular marker for labeling RL cells based on the following observations. (1) Olig3 is expressed in the RL throughout the anterior–posterior axis of the rhombomere (Fig. 1B) (Takebayashi et al., 2002). (2) No Olig3 expression is detected in the roof plate, and the mutation of Olig3 is not related to the development of the roof plate (Fig. 6G,K,L). (3) Olig3 is expressed in the Math1+ and dorsal part of Ptf1a+ domains in cRL, from which MF neurons and CF neurons arise (Fig. 1). (4) Olig3 mutation reduces the generation of MF nuclei and completely inhibits the formation of CF nucleus. Together, these observations strongly suggest that Olig3 predominantly labels RL progenitor cells in the entire hindbrain.
Origin and molecular specification of ION neurons in the caudal hindbrain
Our data showed that Olig3 is specifically expressed in dp1–dp3 domains of the caudal hindbrain at r6–r8 level (Fig. 1C–K). It is well known that Math1 is specifically expressed in the dp1 domain and determines the cell fate of dI1 neurons that substantially form the precerebellar MF neurons (Wang et al., 2005). Loss of Olig3 leads to a modest downregulation of Math1 expression (Fig. 6A,B) and partial reduction of four MF nuclei in the brainstem (Fig. 4). Although the origin of the MF neurons has been well defined, it remains unclear from which region of cRL the precerebellar CF neurons are derived. Classical ablation experiments indicated that removal of the dorsal third of the neuroepithelium in the caudal hindbrain could completely inhibit the formation of ION comprised of CF neurons (Harkmark, 1954). Our results suggested that CF neurons are derived from the Olig3-expressing domains. In Olig3 mutant embryos, dI2 and dI3 Brn3a+ neurons were not produced, in parallel to the complete loss of Brn3a+ CF neurons in the ION. Together, these findings suggested that cRL can be divided into two regions along dorsal–ventral axis: the dp1 progenitor domain that gives rise to the precerebellar MF neurons and the ventral Olig3+ progenitor domains that contribute substantially to the formation of precerebellar CF nucleus.
Recent Ptf1a-Cre lineage tracing experiments showed that Brn3a+ glutamatergic CF neurons are derived from Ptf1a+ progenitor cells, and Ptf1a-null mutation similarly resulted in the loss of ION (Hoshino et al., 2005; Yamada et al., 2007). In light of our finding that Ptf1a was coexpressed with Olig3 in the ventral dp3 domain and a dual requirement of Ptf1a and Olig3 for ION formation (Figs. 1K, 2A), we propose that CF neurons are specifically derived from the Olig3+/Ptf1a+ overlapping region of the ventral dp3 domain (Fig. 1K). Although Olig3 is required for the fate specification of Brn3a+ excitatory CF neurons, Ptf1a activity appears to be essential for the proper migration of CF neurons. In the absence of Ptf1a expression, the generation of Brn3a+ neurons was not compromised, but these neurons failed to migrate into the ION area and subsequently underwent apoptosis (Yamada et al., 2007).
Ptf1a is also expressed in dp4 progenitor cells that give rise to inhibitory neurons (Fig. 1C,F,J,K). In Olig3 mutant embryos, the Ptf1a expression was dorsally expanded in association with the dorsal expansion of Pax2+/Lbx1+ GABAergic neurons at the expense of Tlx3+/Brn3a+ glutamatergic neurons (Figs. 6I,J, 7A–D,G,H). As a result, the formation of Brn3a+ IMS of CF neurons and ION were completely abolished (Fig. 5; supplemental Figs. S1, S2, available at www.jneurosci.org as supplemental material). Collectively, these findings strongly suggested that in the absence of Olig3 expression, the dp2 and dp3 domains were respecified to the dp4 domain. In contrast, the generation of Brn3a+/Tlx3+ dI5 neurons was not affected (Fig. 7A,B,G,H), indicating that dI5 neurons do not contribute to ION population.
ION is largely composed of glutamatergic excitatory neurons (Brn3a+), but it also contains a small number of GABAergic inhibitory neurons (Fredette et al., 1992). The inhibitory ION neurons may originate from the more ventral Ptf1a domain of dorsal caudal hindbrain. The Ptf1a+ neural progenitor cells in dp4 domain are in close association with the Pax2+ neurons, consistent with the previous reports that Ptf1a promotes the fate of GABAergic inhibitory neurons in the dorsal spinal cord and cerebellum (Glasgow et al., 2005; Hoshino et al., 2005).
Olig3 deficiency inhibits the development of the central respiratory center
Olig3−/− mutants died at birth and displayed cyanosis, suggesting that Olig3 function is related to the development of the respiratory system. In support of this idea, Olig3 regulates the specification of dI1–dI3 interneurons, which contribute substantially to the diversity of cell types within the respiratory system. Olig3 mutation may affect several regulatory pathways that are important for the development of the respiratory system. First, loss of Olig3 function significantly affected the formation of four precerebellar MF nuclei through Math1 regulatory pathway (Fig. 4). In particular, the LRN is located in the ventrolateral region of the medulla, where the ventral medullary respiratory center resides. Development of LRN neurons might be particularly important for the functioning of respiratory system. Consistent with this idea, Math1-null mice died shortly after birth from respiratory failure (Ben-Arie et al., 1997). Second, absence of Olig3 completely inhibited the generation of Phox2b+ neurons from dI3 domain and subsequently disrupted the formation of brainstem (nor)adrenergic centers and first-order relay visceral sensory neurons that were critical for respiratory rhythms and cardiovascular control (Fig. 8). It is well known that Phox2b is an important transcription factor regulating respiratory activity; mutations in Phox2b are the main disease-defining mutation in human congenital central hypoventilation syndrome (CCHS), leading to life-threatening hypoxia immediately after birth (Amiel et al., 2003). Tlx3-null mice lost the expression of Phox2b at later embryonic stages and showed a rapid respiratory rate with short inspiratory duration and frequent apnea (Shirasawa et al., 2000; Qian et al., 2001). Third, Olig3 mutation resulted in the misspecification of dI2–dI3 neurons into dI4 interneurons, and Ptf1a was ectopically expressed in dp2–dp3 domains (Fig. 6I,J). Tlx3 expression in dp3 domain was lost in Olig3 mutant hindbrain (Fig. 7G,H). Previous reports demonstrated that Ptf1a promoted neural precursors to differentiate into GABAergic neurons and Tlx3 determined glutamatergic over GABAergic cell fates (Cheng et al., 2004, 2005; Glasgow et al., 2005; Hoshino et al., 2005; Mizuguchi et al., 2006). Consistent with these findings, ectopic expression of Viaat, a GABAergic neuron marker, was detected at the nTS area in E18.5 Olig3-deficient embryos (supplemental Fig. S3G,H, available at www.jneurosci.org as supplemental material). Thus, excess GABAergic neurons existed in Olig3 mutant respiratory circuit, and this could alter the respiratory pattern, offering an alternative explanation for the change of respiratory rhythm in Tlx3 mutant mice. Together, these observations strongly suggest that Olig3 is an important transcription factor regulating the embryonic development of the central respiratory system. Human sudden infant death syndrome (SIDS) is a major cause of infant mortality, but the exact cause of this disease is still unclear. Growing evidence suggests that the impairment of central respiratory control leads to the defect of SIDS patients in response to breath and blood pressure challenges during sleep (Gillan et al., 1989). In addition, one-third of CCHS victims do not carry the mutation of Phox2b gene (Amiel et al., 2003). Therefore, Olig3 should be a candidate gene for human SIDS and CCHS.
Footnotes
-
This work was supported by the National Institutes of Health Grants R21 NS 060033, R01 NS37717, and R01EY016724. We thank Drs. Hirohide Takebayashi, Carmen Birchmeirer, Helena Edlund, and Jane Johnson for generously providing the anti-Olig3, anti-Lbx1, anti-Tlx3, anti-Ngn1, and anti-Ptf1a antibodies.
- Correspondence should be addressed to Mengsheng Qiu at the above addresses. m0qiu001{at}louisville.edu





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}