

Abstract
α-Synuclein has been reported to be important in modulating brain plasticity and to be a key protein in neurodegenerative diseases, including Lewy body dementia (LBD). We investigated how α-synuclein levels modulate adult neurogenesis and the development of dendritic arborization and spines in the dentate gyrus, in which new neurons are constantly added. In the human hippocampus, levels of endogenous α-synuclein were increased in LBD, and the numbers of SOX2-positive cells were decreased. We investigated whether newly generated neurons were modulated by endogenous α-synuclein, and we found increased adult neurogenesis in α/β-synuclein knock-out mice. In contrast, overexpression of human wild-type α-synuclein (WTS) decreased the survival and dendritic development of newborn neurons. Endogenous α-synuclein expression levels increased the negative impact of WTS on dendrite development, suggesting a toxic effect of increasing amounts of α-synuclein. To attempt a rescue of the dendritic phenotype, we administered rolipram to activate the cAMP response element-binding protein pathway, which led to a partial rescue of neurite development. The current work provides novel insights into the role of α-synuclein in adult hippocampal neurogenesis.
Introduction
Cognitive decline is an important symptom in synucleinopathies, having a severe impact on the quality of life (Possin et al., 2008). Clinical and imaging studies have attributed cognitive decline in Parkinson's disease (PD) and Lewy body dementia (LBD) to impairments in hippocampal and cortical circuitries (Camicioli et al., 2003; McKeith et al., 2005; Apostolova et al., 2010; Molano et al., 2010; Pavese et al., 2010; Politis et al., 2010). Recently, a correlation between memory-encoding performance and hippocampal volume (using voxel-based morphometry) in nondemented PD patients was reported (Weintraub et al., 2011). In addition, using diffusion tensor imaging techniques, Carlesimo et al. (2012) showed that microstructural hippocampal alterations in nondemented PD patients were related to decreased declarative memory. The pathological hallmark of PD and LBD is the aggregation of α-synuclein as Lewy bodies and Lewy neurites (Spillantini et al., 1997; Baba et al., 1998). As proposed by Braak et al. (2003), Lewy body formation is observed in different brain regions as the disease progresses, including the limbic system in many cases. Although a physiological presynaptic role for α-synuclein is well established, dysfunction of neurites is presently less understood (Kramer and Schulz-Schaeffer, 2007).
The integration and connectivity of new neurons play important roles in adult neurogenesis in the hippocampus. Adult neurogenesis persists throughout life in the hippocampal dentate gyrus (DG) (Altman and Das, 1965) in all mammals, including humans (Eriksson et al., 1998; Knoth et al., 2010). In the DG, adult neurogenesis contributes to hippocampus-dependent learning and memory (i.e., in spatial pattern separation, as shown in low-dose x-irradiation models; Clelland et al., 2009). Studies in various animals and humans affected by neurodegenerative diseases have convincingly shown that hippocampal neurogenesis is modulated and, under many circumstances, decreased, indicating a limited cellular plasticity (for review, see Winner et al., 2011). The clinical significance of human hippocampal adult neurogenesis in neurodegenerative diseases is still under debate.
The role of α-synuclein in adult neurogenesis has been extensively studied in the subventricular zone/olfactory bulb system, but less is known about its function in the generation of new neurons in the DG. Impaired neurogenesis in the DG was reported in brains from PD patients with dementia and in murine models of synucleinopathies (Höglinger et al., 2004; Winner et al., 2004). We chose a selective and complementary approach to understand the role of α-synuclein in regulating adult hippocampal neurogenesis in PD and LBD. In the work presented here, we examined the endogenous expression of α-synuclein in mouse and human hippocampus with LBD. We observed an increase in new neurons in the DG in an α/β-synuclein knock-out background. In addition, we found that α-synuclein is associated with dendrite and spine development, depending on the expression level. We show here that α-synuclein regulates dendritic maturation of newly generated neurons.
Materials and Methods
Human specimen processing, neuropathological evaluation, and criteria for disease stage.
A total of 12 human cases (n = 6 nondemented controls; n = 6 LBD) was included in the present study (Table 1). Human brain tissue was obtained at autopsy from patients studied at the Alzheimer Disease Research Center/University of California, San Diego. The last neurobehavioral evaluation was performed within the 12 months before death and included Blessed score (range of 0–28; 0–4, normal cognition; ≥10, impairment consistent with dementia). Autopsies were performed within 24 h of death (Table 1, postmortem hours). Brains were processed and evaluated according to standard methods. At autopsy, brains were divided sagittally; the left hemibrain was fixed in Formalin or 4% paraformaldehyde for subsequent neuropathological and immunohistochemical analysis.
Summary of clinico-pathological characteristics of human cases
Animals.
All animal procedures were performed in accordance with the protocols approved by the animal care use committees of the Salk Institute for Biological Studies and University of California, San Diego following National Institutes of Health guidelines for the humane treatment of animals. All mice were kept in a normal 12 h light/dark cycle and had access to food and water ad libitum. Mice overexpressing human wild-type α-synuclein (WTS) under the PDGF promoter were compared with non-transgenic (Tg) littermate controls (NTG). The PDGF WTS group was the WTS high expresser line D; the genetic background was DBA (Masliah et al., 2000; Rockenstein et al., 2002).
α/β-Synuclein knock-out mice lacking genes for α- and β-synuclein (SNCA, SNCB) were obtained from The Jackson Laboratory and have been characterized extensively (Chandra et al., 2004). As controls, we used B6129SF2/J (stock #101045), which are F2 hybrids of C57BL/6 and 129S1/SvImJ mice, because according to The Jackson Laboratory (personal communication), the background of these mice best approximates the mixed background of the B6;129-Snca〈tm1Sud〉 Sncb〈tm1.1Sud〉/J (stock #006390) mice. Acknowledging that the levels of adult neurogenesis can vary between C57BL/6 and 129S1/SvImJ, we added another group of pure C57BL/6 mice to our study.
Analysis of neurogenesis.
Analysis of neurogenesis, including BrdU injection, tissue preparation, and immunostaining, was performed as described previously (Winner et al., 2004). A daily intraperitoneal BrdU injection (50 mg/kg) was given for 5 d. The animals were killed after 1 month. The paradigms are indicated with the respective figures.
Retrovirus-mediated labeling and analysis of new neurons in the mouse hippocampus.
We used a Moloney murine leukemia retrovirus that has been described previously (Zhao et al., 2006) and was a gift from Drs. Gerald Pao and Inder Verma (Salk Institute, La Jolla, CA). In brief, CAG–GFP uses the backbone of pCLNCXv.2. The expression of enhanced green fluorescent protein (GFP) is driven by the compound promoter CAG. The concentrated viral solution (108 pfu/ml) was prepared with human embryonic kidney 293T packaging cells. To perform a detailed analysis of dendritic arborization and spines, adult female PDGF–WTS mice and NTG controls, all aged 4 months (n = 8 per group), were stereotactically injected with 1.5 μl of retrovirus into the right DG at a flow rate of 0.25 μl/min at the following coordinates: anteroposterior, 2 mm; mediolateral, +1.6 mm from bregma; dorsoventral, −2.3 mm from skull.
CAG–WTS–GFP retrovirus was used to selectively study the effect of WTS in newly generated neurons. WTS cDNA was amplified by PCR (nucleotides 53–475; GenBank accession number L08850). To generate the human WTS–GFP fusion construct, WTS cDNA was cloned without a stop codon and ligated to the 5′ end of GFP (nucleotides 826–1545; GenBank accession number U43284), cloned into a Bluescript cloning vector (Invitrogen) and sequenced for accuracy. In this construct, GFP was thus fused to the C-terminal site of WTS. The 5′ WTS–GFP 3′ cDNA fragment was cloned into the CAG vector, concentrated, and stereotactically injected as described above. To compare CAG–GFP and CAG–WTS–GFP, naive female C57BL/6 mice were injected (n = 7, 7 weeks old). For the analysis of CAG–GFP versus CAG–WTS–GFP in α/β-synuclein knock-out mice, α/β-synuclein knock-out mice and controls were injected (n = 7, 7 weeks old). One or 3 months after the injection, the animals were killed and further processed for immunostainings as described previously (Winner et al., 2004).
Rolipram treatment.
Animals in the rolipram experiment received rolipram (1.25 mg/kg; Sigma; dissolved in sterile saline and freshly prepared daily) or sterile saline by intraperitoneal injection once daily at the age of 7 weeks until perfusion 35 d later. Five days after the first injection, CAG–GFP or CAG–WTS–GFP retrovirus was infused into the right hippocampus as described above. In addition, BrdU was administered once daily for 5 d starting on the day of stereotactical infusion as described above, and animals were perfused 1 month later (n = 4 per group).
Antibodies and immunochemicals for mouse tissue.
Primary antibodies and final dilutions for mouse brains included the following: rat α-BrdU at 1:500 (Oxford Biotechnology), mouse α-NeuN at 1:500, mouse α-Sox2 (sex-determining region Y-box 2; marker of neural stem cells), rabbit α-VAMP2 (Millipore Bioscience Research Reagents), goat α-doublecortin (DCX) at 1:500 (Santa Cruz Biotechnology), rat α-human α-synuclein 15G7 at 1:500 (Zymed), chicken α-GFP at 1:250 (Invitrogen), rabbit α-proliferating cell nuclear antigen (PCNA) at 1:500 (Abcam), and rabbit α-cleaved Caspase-3 at 1:250 (Cell Signaling Technology). Secondary antibodies for immunofluorescence included the following: donkey α-goat, α-mouse, α-rabbit, α-chicken, or α-rat conjugated with fluorescein (FITC), rhodamine X, or Cy5 at 1:500 (Jackson ImmunoResearch). To control the specificity of antibody staining, we performed the following negative controls: (1) leaving primary antibody incubation out, (2) preincubation with blocking peptide, (3) isotype controls, and (4) comparison with knock-out animals.
Immunohistochemical analysis of human tissue.
For analysis of levels of α-synuclein and SOX2 in the hippocampus of LBD patients and controls, systematically sampled brain sections were immunolabeled with the antibodies against α-synuclein at 1:1000 (rabbit α-synuclein, 5038P; Millipore). SOX2 antibodies at 1:500 (mouse monoclonal; Sigma-Aldrich) were detected with the Tyramide Signal Amplification-Direct (Red) system (PerkinElmer Life and Analytical Sciences). A subset of sections immunolabeled with the SOX2 antibody was colabeled with the antibody against α-synuclein detected with FITC-conjugated secondary antibodies (1:75; Vector Laboratories).
Human sections were mounted under glass coverslips with ProLong Gold antifade reagent with DAPI (Invitrogen) and imaged with a Carl Zeiss 63× [numerical aperture (NA) 1.4] objective on an Axiovert 35 microscope (Carl Zeiss) with an attached MRC1024 laser scanning confocal microscope system (Bio-Rad; Masliah et al., 2000). All sections were processed simultaneously under the same conditions and the experiments were performed twice to assess reproducibility. To confirm the specificity of primary antibodies, control experiments were performed in which sections were incubated overnight in the absence of primary antibody or of primary antibody preincubated with blocking peptide. For all human immunolabeling studies, assessment of levels of immunoreactivity was performed using the Image-Pro Plus program (Media Cybernetics). For each case, three systematically sampled sections (10 images per section) were analyzed to estimate the average number of immunolabeled cells per unit area (square millimeters).
Immunofluorescence.
Free-floating sections were washed in TBS and then incubated in TBS/0.25% Triton X-100/3% donkey serum for 30 min, followed by incubation with primary antibodies in TBS/donkey serum overnight at 4°C. Afterwards, primary antibodies were applied in TBS/donkey serum for 48 h at 4°C, followed by incubation with secondary antibodies as described above. After several washes in TBS, sections were mounted on gelatin-coated glass slides and coverslipped using Prolong (Invitrogen). For detection of BrdU-labeled nuclei, the following DNA denaturation steps preceded the incubation with anti-BrdU antibody: 2 h incubation in 50% formamide/2× SSC (2× SSC: 0.3 m NaCl and 0.03 m sodium citrate) at 65°C, 5 min rinse in 2× SSC, 30 min incubation in 2 m HCl at 37°C, and 10 min rinse in 0.1 m boric acid, pH 8.5.
Counting procedures.
For quantification, a systematic, random counting procedure, similar to the optical dissector (Gundersen et al., 1988), was used as described by Williams and Rakic (1988).
Every sixth section (200 μm interval) was selected from each animal and processed for immunohistochemistry. Sections were analyzed for BrdU-positive (BrdU+) cells in the hippocampal DG, which were exhaustively counted on each section under exclusion of the uppermost focal plane. The reference volume was determined by tracing the areas using a semiautomatic stereology system (Stereoinvestigator; MicroBrightField).
To determine the frequency of neuronal differentiation of newborn cells, a series of every sixth section (150 μm interval) was examined using a confocal laser microscope (Leica TCS-NT) equipped with a 40× Plan Apochromat oil objective (NA 1.25) and a pinhole setting that corresponded to a focal plane of 2 μm or less. On average, 50 BrdU+ cells were analyzed in the DG of each animal. Cells were randomly selected and analyzed by moving through the z-axis of each cell to exclude false-positive double labeling attributable to the overlay of signals from different cells (Kuhn et al., 1996). Colabeling was analyzed sequentially at each wavelength and in the z-axis to ensure that the fluorophores were present in the identical focal plane.
Dendritic growth and spine density of GFP+ neurons.
These analyses were conducted as described previously (Zhao et al., 2006). Briefly, for dendrite analysis, z-series at 1.5 μm intervals were acquired with a Bio-Rad Radiance 2100 confocal system from sections stained with the GFP antibody. Two-dimensional maximum intensity projections were created with Confocal Assistance Program (Bio-Rad), and files were imported into IGL Trace to measure total length of the dendritic tree within the section. The number of branching points was counted manually from the same images. At least 15 cells from three mice were analyzed for each data point. For spine density analysis and mushroom spine classification, close-up images of GFP+ dendritic processes were acquired in the outer molecular layer at 0.5 μm intervals with a 60× oil lens (NA 1.4; Nikon) and a digital zoom of 5. For spine analyses, sections were not stained to preserve the GFP signal. The Bio-Rad image files were subjected to five iterations of deconvolution with AutoDeblur (AutoQuant). Maximum intensity projections of z-series were created with the Confocal Assistance Program, and files were imported into IGL Trace. The length of each dendritic segment was determined by tracing the center of the dendritic shaft, and the number of spines was counted manually from the two-dimensional projections. Spine density was calculated by dividing the total number of spines by the length of the dendritic segment. For classification of mushroom spines, major and minor axes of each spine head were identified, and the size of the spine head was represented by the estimated surface area with the function 0.25 × π × Dmajor × Dminor. A spine was considered to be of mushroom type if the average estimated area from three measurements was >0.4 μm2 (Zhao et al., 2006). At least 40 dendritic segments from three to four mice were analyzed for each data point. Confocal imaging and data quantification were performed blinded to the experimental conditions.
Statistical analyses.
The data were expressed as mean ± SD values and were tested by an unpaired, two-sided t test for significant differences. Dendrite and spine analyses were compared by one-way ANOVA, and analyses in the rolipram experiment were compared by two-way ANOVA with the variables treatment and virus, both followed by Bonferroni's multiple comparison post hoc tests. Data were analyzed in Prism (GraphPad Software), and the significance level was p < 0.05.
Results
Endogenous expression of α-synuclein in human and mouse
We compared α-synuclein expression in the hippocampi of LBD patients and controls. α-Synuclein+ cells were rarely found in controls (Fig. 1A,B). In LBD patients, α-synuclein was expressed in the deep granule cell layer (DGC), subgranular zone (SGZ), and hilus (Fig. 1A) of the hippocampus, and the number of α-synuclein+ cells was increased in the SGZ of LBD patients (Fig. 1B, Table 1). We found a decrease in SOX2+ cells in the SGZ of LBD patients (Fig. 1C). SOX2 and α-synuclein colabeled in human LBD brains (Fig. 1D). In the DG of murine control mice, endogenous α-synuclein was expressed mainly in the hilus and at low levels in the SGZ and DGC (Fig. 1E). α-Synuclein was absent in the hippocampus of α/β-synuclein knock-out mice (Fig. 1F). Endogenous α-synuclein colabeled with SOX2 in the SGZ and DCX in the DG in mouse brain (Fig. 1G,H).

Endogenous expression of α-synuclein in human and mouse DG. α-Synuclein (α-syn) staining in the brains of LBD patients and controls shows expression in the CA regions and DGC (A) and, less extensively, in the molecular layer (ML; black boxes further magnified in D). The number of α-synuclein+ cells in the DG is significantly increased in the brains of LBD patients (B). SOX2+ cells are decreased in the brains of LBD patients (C). SOX2 and α-synuclein colabel in human LBD brains (D; arrows indicate α-synuclein and SOX2 double-labeled cells). In murine brains, α-synuclein is strongly expressed in the hilus and, to a lesser extent, the DG in control mice (E) but is absent in α/β-knock-out (ko) mice (F). Endogenous α-synuclein colabels with SOX2 and DCX in mouse brain (G, H indicate examples of a double-labeled cell). H, Arrows indicate coexpression with α-synuclein: arrow shows a DCX+ cell with a neurite reaching into the DG, and arrowhead shows a tangential DCX+ cell. Scale bars: A, 500 μm; D, 30 μm; E–H, 50 μm. *p < 0.05 in B and D.
Increased adult neurogenesis in α/β-synuclein knock-out mice
Because α-synuclein was endogenously expressed in the murine and human DG, we asked whether the number of newly generated neurons was also modulated by endogenous α-synuclein. We used α/β-synuclein knock-out mice (Chandra et al., 2004) that were killed 1 month after BrdU injection (Fig. 2A). There was no difference in survival of BrdU+ newly generated cells (Fig. 2B), but there was a significant increase in neuronal differentiation (percentage of BrdU/NeuN double-positive cells) in α/β-synuclein knock-outs (Fig. 2C,E), which led to a significant increase in new neurons (as calculated by the ratio of BrdU × %BrdU/NeuN+ cells; Fig. 2D). A similar increase in neuronal differentiation in α/β-synuclein knock-outs was observed when compared with C57BL/6 mice (Fig. 2B–D). The numbers of PCNA+ cells (Fig. 2F) and activated Caspase-3+ cells (Fig. 2G) were not altered in α/β-synuclein knock-outs.

Neurogenesis in α/β-synuclein knock-out mice. A, Adult neurogenesis was quantified in α/β-synuclein knock-out mice (α/β-syn ko) and two control groups using a 1 month BrdU survival paradigm (n = 7 animals per group). B, Survival of newly generated cells (number of BrdU+ cells) was unchanged between groups. However, there was a significant increase in relative (C) and absolute (D) numbers of newborn neurons in α/β-synuclein knock-out mice. Representative micrographs of the DG of littermate control and in α/β-synuclein knock-out mice (E). Proliferation (number of PCNA+ cells; F) and cell death (number of activated Caspase-3+ cells) in G were not significantly different. Scale bars: E, 50 μm; F, 200 μm; G, 20 μm. *p < 0.05, **p < 0.01.
Temporal expression pattern of WTS Tg mice resembles the endogenous expression
To further investigate the role of α-synuclein on newly generated neurons, we investigated colocalization of WTS with markers of specific stages of neuroblast development. In PDGF WTS mice, WTS was already expressed at the stem cell stage, as depicted by nuclear colabeling with SOX2 (Fig. 3A). WTS was present throughout the development of the newly generated neurons, as indicated by colabeling with DCX (Fig. 3B), and it was also expressed in NeuN+ mature neurons (Fig. 3C). In this PDGF WTS mouse model, α-synuclein was expressed from birth through maturation of adult-born neurons. Equally important was that, in the DG, α-synuclein accumulated along neurites in close proximity to the newly generated neurons (Fig. 3B).

Colocalization of α-synuclein (α-syn) in PDGF WTS mice. Colocalization analysis of human WTS with markers at different stages of neuroblast development in the DG reveals temporal expression patterns of WTS during the generation of new neurons in PDGF WTS mice. WTS is expressed at all adult neural progenitor stages, colabeling with SOX2 (A), DCX (B), and NeuN (C). Arrows indicate examples of double-labeled cells in A–C. Scale bars, 25 μm.
Compromised dendritic morphology in α-synuclein Tg animals
To further investigate the impact of α-synuclein on neurites and to determine the impact of WTS on dendrite and spine development in newly generated neurons, we injected a retrovirus expressing green fluorescent protein (CAG–GFP) into PDGF WTS mice (paradigm; Fig. 4A). Four weeks after viral injection, the dendrites of newborn granule cells in NTG mice showed a highly polarized morphology and apical dendritic processes with abundant spines extending into the molecular layer (Fig. 4D). Compared with NTG mice, the dendritic length in PDGF WTS mice was significantly decreased (Fig. 4B,E); however, no differences in branching points were observed between the groups (Fig. 4C). Given that dendritic growth was perturbed in PDGF WTS mice, we next asked whether spine formation or spine maturation of granule neurons was affected. Surprisingly, we found a significant increase in spine density in the PDGF WTS group (Fig. 4G,H). Tg overexpression of WTS also affected spine maturation in PDGF mice; the formation of mushroom spines was decreased in PDGF WTS mice (Fig. 4I). These results led us to further determine the impact of α-synuclein within the newly generated neurons compared with the effect induced by WTS of the surrounding neurogenic niche.

Compromised dendritic morphology in α-synuclein Tg animals. A, Paradigm: CAG–GFP retrovirus was stereotactically injected into the DG of PDGF WTS and NTG (n = 8 animals per group). B, Total dendritic length of newly generated neurons was significantly decreased in PDGF WTS mice. C, No significant differences were detected in the number of dendritic branching points. Representative images of dendrites (D, E) and spines (F, G; arrows in F and G and insets indicate mushroom spine example) analyzed from the respective groups. The analysis of dendritic spines (H) revealed an increase in spine density of newly generated neurons in PDGF WTS animals. I, Density of mushroom spines was significantly decreased in PDGF WTS mice. Scale bars: D, E, 25 μm; F, G, 10 μm. *p < 0.05.
Overexpression of WTS in newly generated neurons leads to decreased dendritic outgrowth and fewer spines
To investigate the cell-autonomous effects of WTS on newly generated neurons, we used a WTS–GFP fusion protein retrovirus (CAG–WTS–GFP) and a GFP control retrovirus (CAG–GFP). CAG–WTS–GFP and CAG–GFP were injected into the DG of C57BL/6 mice, and the animals were killed 1 month (Fig. 5) or 3 months (data not shown) after the stereotactic injection. We first explored the expression of the CAG–WTS–GFP retrovirus and found it to be similarly distributed throughout the neurites when compared with a coinjected CAG–RFP retrovirus (Fig. 5B). Moreover, CAG–WTS–GFP colabeled with α-synuclein (Fig. 5C) and the mossy fiber synaptic marker VAMP2 in the CA3 region, indicating that the retrovirus was distributed throughout the entire newly generated neuron (data not shown). The amount of overexpression, as analyzed by quantitative PCR of the microdissected CAG–WTS–GFP-injected DG (data not shown), was similar to what has been reported in WTS Tg mice (Rockenstein et al., 2002). One month after retroviral injection, dendritic length, dendritic branching points (Fig. 5D,E), and spine density (Fig. 5F,G) were significantly decreased by overexpression of WTS selectively in newly generated neurons compared with the control CAG–GFP retrovirus injection alone. These findings indicate that overexpression of WTS within the newly generated neuron significantly impaired dendritic and spine outgrowth. No significant differences were detected in the numbers of mushroom spines (Fig. 5F,G). The effect of decreased dendritic length and decreased spine density was present even after 3 months (data not shown). Because of this effect of overexpression of WTS in newly generated neurons on dendrites and spines, we next asked whether endogenous mouse α-synuclein expression within the DG had an impact on dendrite and spine development.

Effects of WTS on dendrite and spine morphology. A, Paradigm: retrovirus overexpressing a human wild-type α-synuclein–GFP fusion protein (CAG–WTS–GFP) or GFP control (CAG–GFP) was injected into the DG of naive mice, and the mice were analyzed 1 month later (n = 7 animals per group). B, Retrovirus CAG–RFP that overexpresses RFP in infected cells was coinjected with CAG–WTS–GFP. Overlay of GFP (green) and RFP (red) shows that both fluorophores were similarly distributed throughout dendrites and the perikaryon. C, Colabeling for GFP and α-synuclein (15G7; α-syn) shows colabeling of dendritic trees and perikarya of infected newborn neurons. Representative images of dendrites (D) and spines (F; inset shows examples of mushroom spines, arrows). Dendritic length, dendritic branching points (E), and spine density (G) were decreased in the CAG–WTS–GFP group. No significant differences were detected in the number of mushroom spines (G). Scale bars: B–D, 50 μm; F, 10 μm. *p < 0.05, **p < 0.01, ***p < 0.001.
Effects of endogenous α-synuclein on dendrite and spine development
To investigate the impact of hippocampal α-synuclein on the development of new neurons, we analyzed the effects on neurite outgrowth of newly generated neurons in the presence or absence of α-synuclein. We first asked whether the DG neurite and spine development of newly generated neurons was altered in these α/β-synuclein knock-out mice, and we labeled the newly generated neurons by injection of retrovirus (CAG–GFP). No differences in dendrite (Fig. 6A,B,E,G) or spine (Fig. 6C–E,G) development in newly generated neurons for CAG–GFP were observed in the α/β-synuclein knock-out mice compared with controls (control and C57BL/6).

Effects of WTS on dendrite and spine development in α/β-synuclein knock-out mice (α/β-syn ko). We asked how endogenous α-synuclein levels influenced the effects of cell-autonomous WTS on dendrite and spine morphology. We performed the same injection paradigm as in Figure 5A in α/β-synuclein knock-out mice and two wild-type groups (controls and C57BL/6, n = 7 animals per group). In the absence of endogenous α-synuclein, overexpressed WTS does not impair dendrite outgrowth (A) or dendritic branching (B). The effects on spines, however, are present despite a deficiency in endogenous α-synuclein (C). The mushroom spine subpopulation is unchanged regardless of cell-autonomous α-synuclein content (D). Representative micrographs of controls injected with CAG–GFP (E) or CAG–WTS–GFP (F) and α/β-synuclein knock-out mice injected with CAG–GFP (G) or CAG–WTS–GFP (H). Scale bars (in E–H): top row, 100 μm; bottom row, 10 μm. *p < 0.05, **p < 0.01, ***p < 0.001.
Next we investigated the effect of expression of WTS within the newly generated neurons by injecting the CAG–WTS–GFP retrovirus into the DG in an environment not expressing α/β-synuclein. Control mice (control and C57BL/6) injected with CAG–WTS–GFP showed a significant decrease in dendrite length and branching points (Fig. 6A,B,F). Surprisingly, overexpression of WTS in newly generated DG neurons in the α/β-synuclein knock-outs did not show a reduction in dendrite length and branching points (Fig. 6A,B,H), but the values of the WTS–CAG–GFP group for dendrite length and branching points were similar to those of the CAG–GFP-injected controls (Fig. 6A,B,H). After WTS–CAG–GFP injection, spine density was equally impaired in α/β-synuclein knock-outs and littermate controls (Fig. 6C), but mushroom spine numbers were not altered under any of the conditions analyzed (Fig. 6D).
Rolipram reverses WTS-induced spine alterations
Because these results revealed that modulation of α-synuclein had a strong impact on adult neurogenesis and dendrite and spine development, we sought to determine whether we could reverse the deficit. We tested a pharmacological restoration of the WTS-induced impairment in dendrite and spine development by administration of rolipram. This selective cAMP-specific phosphodiesterase 4 inhibitor increases cAMP and, consecutively, phosphorylation of CREB (MacKenzie and Houslay, 2000). We treated C57BL/6 mice daily with rolipram for a total of 36 d, starting 5 d before retroviral labeling (CAG–GFP and CAG–WTS–GFP) of newly generated neurons, and we continued this treatment until perfusion (Fig. 7A). In the control CAG–GFP group, rolipram increased the number of mushroom spines (Fig. 7E). Dendritic length, dendritic branching, and spine density were not statistically different between treated and control groups (Fig. 7B–D). In addition, we confirmed an increase in adult neurogenesis in the rolipram-injected animals, as described previously (data not shown).
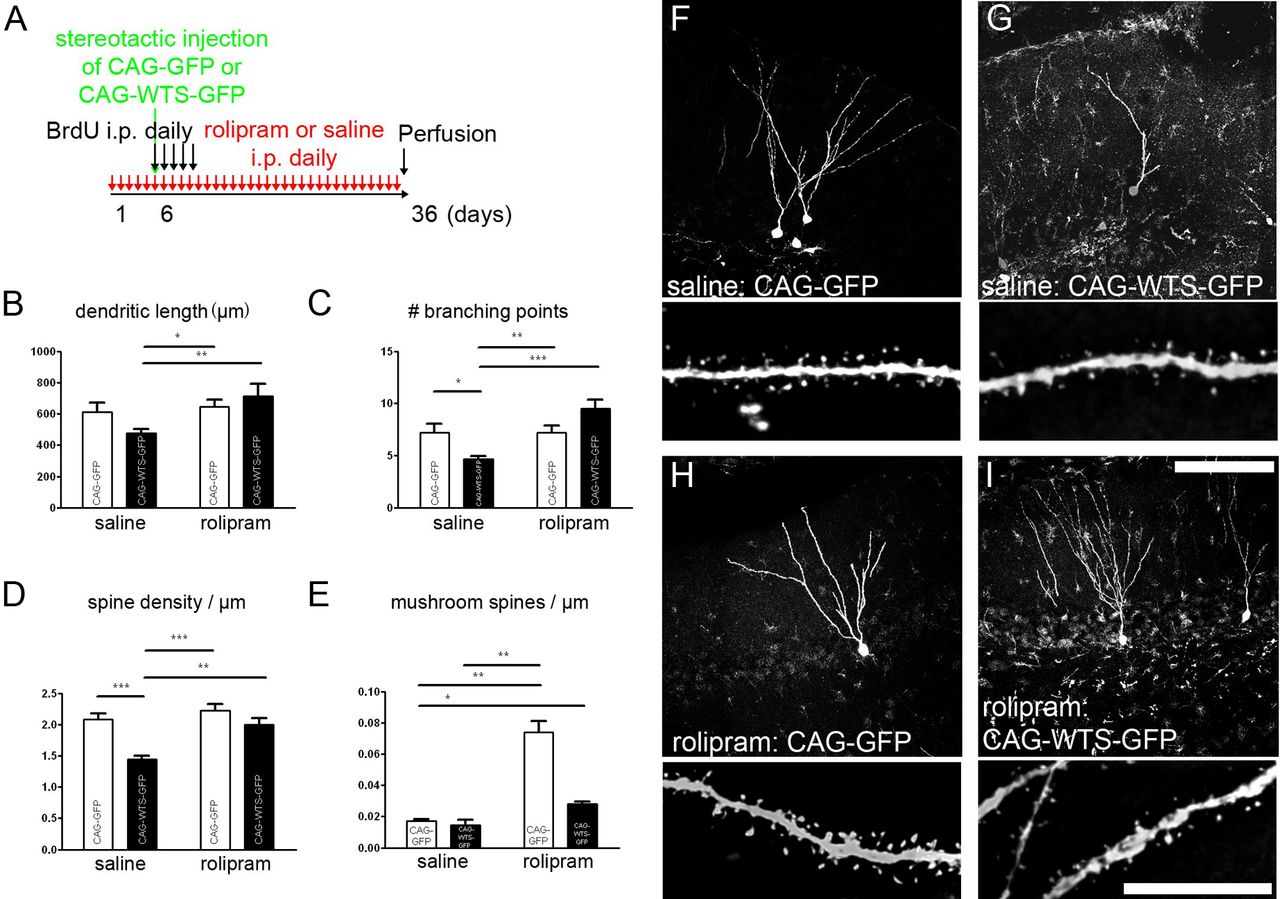
Partial reversal of WTS-induced spine alterations by CREB-activator rolipram. Daily treatment with rolipram (1.25 mg/kg) was started 5 d before retroviral labeling of newborn neurons and continued until perfusion (A). Dendritic length was significantly different between newborn neurons overexpressing CAG–WTS–GFP in saline versus rolipram-treated animals (B). Likewise, impairments in dendritic branching and spine density were not present in rolipram-treated animals injected with CAG–WTS–GFP (C, D). No significant effect of rolipram treatment on mushroom spines was observed in the CAG–WTS–GFP group (E). F–I, Representative images from saline-treated animals injected with CAG–GFP (F) or CAG–WTS–GFP (G) and from rolipram-treated animals injected with CAG–GFP (H) or CAG–WTS–GFP (I). Scale bars (in F–I): top row (dendrite overviews), 100 μm; bottom row (spine magnifications), 10 μm. *p < 0.05, **p < 0.01, ***p < 0.001.
The dendritic length of newly generated neurons overexpressing CAG–WTS–GFP was increased by rolipram treatment compared with the saline-treated CAG–WTS–GFP-injected animals (Fig. 7B). Rolipram treatment increased dendritic branching and spine density in CAG–WTS–GFP-injected animals (Fig. 7C,D). It has to be noted that, in saline-treated animals, WTS led to a trend of dendrite length reduction that, in contrast to the results above, was not significant.
Discussion
This study provides evidence for the involvement of α-synuclein in regulating adult hippocampal neurogenesis. We showed that knock-out of α/β-synuclein increased neuronal differentiation, whereas overexpression of WTS within the newly generated neuron in PDGF WTS mice and retroviral CAG–WTS–GFP overexpression had a negative impact on adult neurogenesis and dendrite development. The overexpression of WTS within the hippocampal niche (e.g., PDGF WTS) also had an impact on spine development. Interestingly, the interaction between WTS expression within the newly generated neuron and expression of mouse endogenous α-synuclein around the newly generated neuron also played an important role in dendrite development in the newly generated neuron, as summarized in Figure 8.

Model of effects of human WTS and mouse endogenous α-synuclein. Model of the physiological development of a newborn neuron in the adult DG (A). A newly generated neuron integrating into the DG is shown on the left of all images; the right depicts a higher-magnified inset of a dendrite showing spines and the larger mushroom spines (A–D). B–D, Models showing the impact of WTS on dendrite and spine development. B, PDGF WTS mice; C, retroviral overexpression of WTS using a CAG–WTS–GFP construct; D, retroviral overexpression of WTS on a α/β-synuclein knock-out background (α/β-syn ko). Cell-specific overexpression of WTS results in decreased dendrite length and spine density (B, C), whereas nonspecific overexpression (B) leads to a decrease in mushroom spines, as indicated in the purple panels showing an example of magnification of the spines.
The increased number of α-synuclein+ cells in the human DG in LBD patients and the decreased progenitor cell population is suggestive of a decrease in adult neurogenesis in LBD. We provide evidence here that α-synuclein might contribute to decreased numbers of progenitors in synucleinopathies, which is in line with a recent human study in LBD brains (Johnson et al., 2011).
To further clarify the impact of α-synuclein on the DG, we investigated adult neurogenesis in the absence of α-synuclein. Because α-synuclein and β-synuclein are both widely colocalized in presynaptic nerve terminals and highly coexpressed, indicating redundancy (Li et al., 2002), we used double α/β-synuclein knock-out mice (Chandra et al., 2004). The downregulation of α/β-synuclein resulted in an increased percentage of surviving new neurons, which was unlikely to be background related, because the number of new neurons was even higher than pure C57BL/6 mice, which are known to have a high level of neurogenesis (Kempermann et al., 2006). Our group had shown previously that overexpression of α-synuclein decreased hippocampal neurogenesis (Winner et al., 2004; Nuber et al., 2008). The finding of increased neurogenesis in α/β-knock-outs further indicates a modulatory role for α- and/or β-synuclein in adult neurogenesis. Therefore, we pursued further the study of cell-autonomous versus non-autonomous interactions of α-synuclein in newly generated neurons in the DG.
α-Synuclein was expressed in the hippocampus within new neurons endogenously in controls and in PDGF WTS mice. The expression of endogenous α-synuclein was similarly shown by in situ hybridizations in the Allen Brain Atlas (mouse brain: SNCA, riboprobe identification number RP_Baylor_103043, coronal; http://www.brain-map.org/). Expression of transgenic α-synuclein in PDGF WTS mice (line D; Rockenstein et al., 2002) and its distribution in the hippocampus were also comparable. By analyzing neurite outgrowth after injection of a GFP retrovirus into the PDGF WTS mice and controls, we showed that dendrite outgrowth of newly generated neurons was significantly impaired in the PDGF WTS mice. To further test the impact of α-synuclein in newly generated neurons, we investigated selective overexpression of WTS in newly generated neurons. Again, we observed a decrease in dendritic length, and branching was also impaired.
To distinguish the impact of endogenous and overexpression of α-synuclein in newly generated neurons, we used an α-/β-synuclein knock-out model. In the absence of endogenous mouse α-/β-synuclein, dendrite development of CAG–GFP-labeled newly generated neurons was not altered. However, the previously observed effect of dendrite length reduction after overexpression of α-synuclein in newly generated neurons (CAG–WTS–GFP) was no longer present in the α/β-synuclein knock-out mice. These experiments showed that cell-autonomous overexpression of WTS induced a reduction in dendrite length. This reduced dendrite length could be modulated by changes in the concentration of endogenous α-synuclein levels even if they were only detected at low levels.
Reductions in dendrite numbers, dendrite length and branching, and spine density have been reported in striatal medium spiny neurons in PD patients (McNeill et al., 1988; Zaja-Milatovic et al., 2005) and in the substantia nigra (Patt et al., 1991). In addition, in vitro studies reported a negative impact of α-synuclein on neurite outgrowth and synaptic integration (Takenouchi et al., 2001; Crews et al., 2008).
The postsynaptic effects of α-synuclein on dendrites and spines are less well understood. The increase in spine density observed in the PDGF WTS mice might be indicative of a compensatory effect in early presynaptic and postsynaptic pathology, in which pruning is reduced as a result of dendritic atrophy and reduced connectivity (for review, see Fiala et al., 2002). When WTS levels were selectively increased in the newly generated neurons but not increased in their surroundings, spine numbers were decreased. Similar observations were made in α/β-synuclein knock-out mice.
Reductions in mushroom spines, indicating a lack of maturation, have been observed in PDGF WTS mice. Because increased expression of α-synuclein reduces neurotransmitter release as well as synaptic vesicle reclustering (Nemani et al., 2010), we speculate that the overexpression of α-synuclein hinders normal levels of presynaptic activity. These findings further strengthen the impact of the presynaptic compartment of spine maturation and size in synucleinopathies.
Rolipram restored WTS-induced morphological dendrite alterations in dendritic length, dendritic branching points, and spine density. Rolipram itself is a specific phosphodiesterase-4 inhibitor leading to increased levels of cAMP, which activates PKA and CREB (Silva et al., 1998). Dominant-negative CREB overexpression prevents the proliferative effect on neural progenitor cells (Nakagawa et al., 2002). The impact of rolipram in the hippocampus may also involve the glucocorticoid receptor (GR), because the effect of rolipram on adult neurogenesis is prevented by a GR antagonist (Anacker et al., 2011).
In PD postmortem brains, the large amount of presynaptic α-synuclein aggregates has a major pathological impact on dendritic spine maintenance. Decreases in the postsynaptic scaffold protein postsynaptic density-95 and the f-actin-binding postsynaptic protein drebrin have been reported in LBD, indicating the impact of synaptic dysfunction in synucleinopathies (Kramer and Schulz-Schaeffer, 2007). These findings have parallels with other neurodegenerative diseases, e.g., Alzheimer's disease, in which a correlation between amyloid oligomer amount, dendritic spines, and memory deficit was shown (Terry et al., 1991; Walsh et al., 2002). It is intriguing that similar connections between protein aggregation, synaptic connectivity, and memory function might be present in synucleinopathies. Our results reveal a negative impact of α-synuclein on newly generated neurons, in particular on their dendrite outgrowth and spine development. These deleterious effects of α-synuclein may underline some components of the cognitive impairments reported in synucleinopathies.
Footnotes
This work was supported by grants from the Alexander von Humboldt Foundation, California Institute of Regenerative Medicine, the Michael J. Fox Foundation, the JPB Medical Foundation, the Lookout Fund, the Mather's Foundation, the Ellison Foundation, the Glenn Foundation, the Bavarian Ministry of Sciences, Research, and the Arts in the framework of the Bavarian Molecular Biosystems Reseach Network and ForNeuroCell, German Federal Ministry of Education and Research Grant 01GQ113, the Independent Research Program (School of Medicine, University of Regensburg), ELAN Program for supporting clinical research and teaching 11.08.19.1 (I.P.), and the Interdisciplinary Center for Clinical Research (University Hospital of Erlangen). We thank Robert Aigner, Daniela Graef, Eunice Meija, Barbara Miller, and Chris Tse for excellent technical support and Christian Alzheimer and Tobias Huth for the use of the confocal microscope. We are grateful to Stefan Aigner, Nada Ben Abdallah, Armin Blesch, Zacharias Kohl, and Chichung D. Lie for helpful comments on this manuscript and discussions and M. L. Gage for editing this manuscript.
- Correspondence should be addressed to Dr. Beate Winner, Interdisciplinary Center for Clinical Research, Nikolaus-Fiebiger Center for Molecular Medicine, Friedrich-Alexander-Universität Erlangen-Nürnberg, Glücksstrasse 6, 91054 Erlangen, Germany. beate.winner{at}med.uni-erlangen.de





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}