

Abstract
The population of mesolimbic dopaminergic neurons is believed to be a primary site at which opiates produce their rewarding effects. Using an unbiased, counterbalanced place conditioning paradigm, we reexamined the contribution made by these cells to the rewarding properties of morphine. Rats were conditioned such that distinct environments were paired with an intra-ventral tegmental area (VTA) microinfusion of either 500 ng per 0.5 μl per side morphine or 0.5 μl per side sterile saline. Furthermore, rats were conditioned either previously drug-naive or while in a motivational state of opiate dependence and withdrawal. We report that pretreatment with the broad-spectrum dopamine antagonist α-flupentixol blocked the acquisition of conditioned place preferences for environments paired with morphine microinjections directly into the VTA in opiate-dependent and withdrawn, but not in previously drug-naive, rats. Lesions of the tegmental pedunculopontine nucleus (TPP) produced exactly the opposite pattern of results. TPP lesions blocked the acquisition of conditioned place preferences for environments paired with VTA morphine microinjections in previously drug-naive, but not in opiate-dependent and withdrawn, rats. These data double-dissociate two independent reward substrates within the VTA itself and suggest that deprivation state selects which of these two substrates will be active. Furthermore, these findings are the first to demonstrate a nondopaminergic substrate for reward within the VTA itself.
The mesolimbic dopaminergic pathway originating from the cell bodies in the ventral tegmental area (VTA) and projecting to the nucleus accumbens is considered a fundamental component in the neural system mediating the motivational properties of opiates (Wise and Rompre, 1989; Wise, 1996). Opiate receptors in the VTA are thought to be a primary site at which systemically administered opiates act to produce reward (Britt and Wise, 1983). Indeed, even critics of this model who posit that the mesolimbic pathway is sufficient, but not necessary, for mediating the motivational properties of systemic opiates concur that the rewarding properties of intra-VTA morphine are mediated by this ascending dopaminergic pathway (Koob and Bloom, 1988;Koob, 1992). In contradiction to the hypothesis that morphine’s rewarding properties are dopamine-dependent are findings that double-dissociate two separate motivational systems mediating the motivational properties of systemic morphine, systemic heroin, and food (Bechara and van der Kooy, 1992a,b; Bechara et al., 1992; Nader et al., 1994; Nader and van der Kooy, 1996). With respect to opiates, this model posits that dopamine transmission is only important in mediating the motivational effects of opiates when animals are in a deprived state (i.e., opiate-dependent and in withdrawal). The TPP, on the other hand, mediates morphine’s rewarding properties only when animals are in a nondeprived state (cases when animals are not in a state of withdrawal: previously drug-naive rats, rats that have recovered from a state of dependence, and rats that are opiate-dependent but not in a state of withdrawal) (Bechara and van der Kooy, 1992b; Bechara et al., 1992; Nader et al., 1994; Nader and van der Kooy, 1996). Specifically, the ventromedial portion of the TPP, which contains only a sparse population of cholinergic cells, has been identified as the critical region mediating opiate reward in previously drug-naive rats (Bechara and van der Kooy, 1989). Furthermore, once in a state of deprivation, the TPP-mediated nondeprived system is inhibited (Bechara and van der Kooy, 1992b; Nader and van der Kooy, 1994).
We undertook a rigorous test of whether the mechanisms mediating morphine’s motivational properties within the VTA can be best described by actions either through a single dopaminergic mechanism or through multiple dissociable mechanisms that are contingent on an animal’s motivational state. Animals were conditioned using an unbiased, counterbalanced place conditioning procedure such that one environment was paired with a just suprathreshold dose (500 ng per 0.5 μl per side) of morphine bilaterally microinjected directly into the VTA, and the other environment was paired with sterile saline (0.5 μl per side) microinjected into the VTA. We predicted that if the dopaminergic mesolimbic pathway is necessary for mediating morphine’s motivational properties, then dopamine antagonists should block the acquisition of conditioned place preferences for environments paired with intra-VTA morphine, regardless of whether animals are trained previously drug-naive or in a state of opiate dependence and withdrawal. On the other hand, if separate systems mediate morphine’s rewarding properties, then we predicted that dopamine antagonist pretreatment would block the acquisition of conditioned place preferences for environments paired with intra-VTA morphine in opiate-dependent and withdrawn, but not in previously drug-naive, animals. Furthermore, TPP lesions should block the acquisition of place preferences for environments paired with intra-VTA morphine in previously drug-naive, but not in opiate-dependent and withdrawn, animals.
MATERIALS AND METHODS
Subjects. All animals used in these experiments were adult (350–400 gm) male Wistar rats (Charles River). Subjects were individually housed in suspended gray wire cages in a room kept at a temperature of 22°C with lights on from 9:00 P.M. to 9:00 A.M. Throughout the duration of the experiments, animals had ad libitum access to food and water.
Surgery. Rats were anesthetized with 0.8 ml/kg intraperitoneal doses of sodium pentobarbital (Somnitol) and placed in a stereotaxic apparatus with the incisor bar set at −3.3. All coordinates were taken from Paxinos and Watson (1982). Cannulae were positioned into the VTA using the same procedure as described previously (Jaeger and van der Kooy, 1993). Briefly, 22-gauge guide cannulae were angled 10° toward the midline and implanted 1.5–2 mm dorsal to the VTA (from bregma: AP −5.0, L ±2.3; from the dural surface: V −8.0). The coordinates used for cannulae aimed dorsal to the VTA were AP −5.0, L ±2.3 and from the dural surface V −7.0. The guide cannulae were anchored to the skull by dental acrylic cement and stainless steel screws.
Lesions of the ventromedial TPP were performed by injecting bilaterally 0.2 μl of either a 2% ibotenic acid solution (lesion group) or physiological saline (sham group) via a 1 μl Hamilton microsyringe over a 20 min period. The needle was left in place for 5 min after the end of the infusion. The injection coordinates for the ventromedial TPP were AP 7.8 mm posterior to bregma, L 1.6 mm lateral to the midline, and DV 6.8 mm below the dura. For animals that received both cannulae implantation and TPP lesions, the two surgical procedures were performed at the same time. At least 2 weeks were allowed for recovery from surgery before any conditioning.
Histology. At the end of the behavioral experiments, all rats were deeply anesthetized and intracardially perfused with isotonic saline, followed by 10% formalin. The brains were removed and post-fixed in a 20% sucrose solution. The brains were then cut in 32 μm sections, which were mounted and stained with cresyl violet to verify the placement of both cannulae and TPP lesions. Sections were examined and photographed under bright-field microscopy.
Microinfusion. For intracranial microinfusions, obturators were removed from the animal’s skull cap, and a 28-gauge injector cannula was lowered to the injection site. The injector cannula protruded 1.5–2.0 mm ventral to the guide cannulae. Polyethylene tubing (PE-50, Clay Adams) connected the injector to a 1 μl Hamilton microsyringe, which was loaded with the appropriate solution before microinfusion. The rats were held by the experimenter while infusions were made over a 1 min period. The accuracy of the volume infused was assessed by observing the progress of an air bubble within the tubing. After termination of the infusion, the injector was left in place for an additional 1 min before being removed.
Apparatus and place conditioning procedure. The place conditioning apparatus was identical to that described previously (Mucha et al., 1982; van der Kooy, 1987). Briefly, conditioning took place in one of two distinct environments that differed in color, smell, and texture. One environment was white with a wire mesh floor that was covered with wood chips. The other environment was black with a smooth Plexiglas floor that was wiped down with a 2% acetic acid solution before each conditioning session. At testing, a narrow, neutral gray zone (on which the rats were placed at the beginning of each test session) separated the two test compartments.
All animals were conditioned using a standard place conditioning procedure (Mucha et al., 1982; van der Kooy, 1987). With the exception of the preliminary investigations, the conditioning protocol was identical for all experiments. The only independent measures that were manipulated were the state animals were trained in (previously drug-naive vs in a motivational state of opiate dependence and withdrawal) and pretreatment (TPP lesions vs dopamine antagonist). During conditioning, animals were given an intracranial microinfusion of morphine directly into the VTA and exposed to one of the two conditioning environments for 45 min. On the alternate day, rats received microinfusions of sterile saline and were placed in the alternate conditioning environment for the same period of time. This procedure was repeated until each animal had received a total of four drug-environment and four saline-environment pairings. Both treatment compartment and presentation order of drug and vehicle were counterbalanced within groups. Once training was terminated, all rats received a 1 week recovery period during which they were left undisturbed in their home cages. On test day, rats were individually given free access for a 10 min period to both training environments, and the subsequent times spent in each were recorded. A comparison of the times spent in the drug versus saline paired environments was used as an index for whether animals preferred one compartment over the other.
Animals trained in a state of opiate dependence and withdrawal received daily 0.5 mg/kg subcutaneous injections of heroin commencing 4 d before the start of conditioning. During conditioning, this dose of heroin was administered daily 3.25 hr after the termination of training. Although four subcutaneous injections of 0.5 mg/kg over 4 d do not induce dramatic classical somatic withdrawal signs, they are sufficient to induce a state of motivational dependence, and 20 hr after heroin injection, the time at which conditioning was performed in the present study, rats are undergoing withdrawal as measured by the conditioned aversions to environments paired with the absence of drug (Nader et al., 1994). The motivational effects of withdrawal induced by this regimen are isomorphic with those observed after a 3 week regimen of morphine administration, which produces aversive motivational withdrawal effects (that are dopamine-mediated) as well as intense somatic withdrawal signs (Bechara et al., 1995).
Preliminary investigations. These studies differed from the above protocol with respect to dose of intra-VTA morphine used to condition place preferences, placement of the injector cannulae, or the training drug. To determine a dose of morphine that produces reliable conditioned place preferences, we performed a dose–response curve for intra-VTA morphine-conditioned place preferences. Separate groups of previously drug-naive animals were conditioned with bilateral intra-VTA morphine 250 ng per 0.5 μl per side (n = 8), 500 ng per 0.5 μl per side (n = 7), or 2500 ng per 0.5 μl per side (n = 11) injections using the above procedure. On saline days, animals received bilateral 0.5 μl per side saline microinfusions into the VTA. Based on the findings from this study, a dose of 500 ng per 0.5 μl per side morphine was chosen for all subsequent experiments.
We tested whether morphine’s rewarding properties were the result of actions within, and not dorsal to, the VTA. One group of previously drug-naive (n = 7) and one group of opiate-dependent and withdrawn (n = 6) rats were conditioned with 500 ng per 0.5 μl per side morphine microinfused 1 mm dorsal to the VTA. Furthermore, to verify that the mechanisms in the VTA that supported morphine reward were receptor-mediated, we conditioned another group of previously drug-naive rats (n = 8) with 500 ng per 0.5 μl per side of the inactive (+)-morphine isomer (Jacquet et al., 1977) directly into the VTA.
Effects of α-flupentixol on intra-VTA morphine-mediated place preferences. A total of six groups of rats were used for this experiment. Two groups were trained previously drug-naive using a dose of 500 ng per 0.5 μl per side of the active (−)-morphine isomer. Rats were pretreated 2.5 hr before every conditioning session with either 0.8 mg/kg α-flupentixol (i.p.) (n = 7) or saline (n = 7) injections. At this dose, α-flupentixol blocks both D1 and D2 receptors (Creese et al., 1976). Furthermore, at this pretreatment time and dose, α-flupentixol does not have any motivational properties itself (Harrington and van der Kooy, 1992). Two other groups of rats were made opiate-dependent and trained in a state of opiate withdrawal using the same 500 ng per 0.5 μl per side intra VTA dose of morphine. Again, rats were pretreated 2.5 hr before each conditioning session with either 0.8 mg/kg α-flupentixol (i.p.) (n = 8) or saline (n = 9) injections.
The last two groups of rats were conditioned previously drug-naive with 500 ng per 0.5 μl per side intra VTA morphine. Again, rats received either saline (n = 6) or α-flupentixol (n = 8) injections (i.p.) 2.5 hr before each conditioning session. After testing, these last two groups of animals were counterbalanced between groups with respect to pretreatment (α-flupentixol or saline) and within groups for morphine-paired environments (black or white). That is, half the animals that were in the α-flupentixol pretreatment group were now placed in the saline pretreatment group and vice versa. Similarly, half the animals in each group were slotted to receive morphine pairings to the same side as the previous experiment, whereas the other half of the animals in each group were slotted to receive morphine pairings to the side that animals had received saline in the previous experiment. One week after testing for the conditioned place preferences in previously drug-naive animals, these same rats were made dependent (dependence was induced with systemic heroin as described above) and then trained again using the identical protocol as above, but this time while in a state of opiate withdrawal. Rats were pretreated 2.5 hr before conditioning with either 0.8 mg/kg (i.p.) α-flupentixol (n = 7) or saline (n = 7). After a 1 week recovery period, all animals were tested as described above.
Effects of TPP lesions on intra-VTA morphine-mediated place preferences. A total of four groups, two sham and two TPP-lesioned, were used for this experiment. One group of sham (n = 9) and one group of TPP-lesioned (n = 8) animals were conditioned while previously drug-naive. The remaining two groups, one sham (n = 8) and one TPP-lesioned (n = 7), were made opiate-dependent and trained while in a state of opiate withdrawal. In all cases, animals were conditioned with 500 ng per 0.5 μl per side intra VTA morphine dose. Once testing was completed, sham and TPP-lesioned animals that were conditioned opiate-dependent and in a state of withdrawal were given an additional week to recover from dependence. To verify behaviorally the effectiveness of the TPP lesions, the previously opiate-dependent animals were counterbalanced with respect to the drug-paired environment within groups and subsequently conditioned using a systemic 10 mg/kg (i.p.) dose of morphine (the animals conditioned with intra-VTA morphine while drug-naive were not retrained). That is, half the animals in each of the TPP and sham previously dependent groups were slotted to receive morphine pairings to the same side as the first time they were trained, whereas the other half of the animals in each group were slotted to receive morphine pairings to the side that animals had previously received intra-VTA saline. Previous evidence shows that once animals have recovered from dependence, the rewarding properties of 10 mg/kg (i.p.) morphine are TPP-dependent (Nader et al., 1994). Thus, effective lesions of the TPP should block the acquisition of place preferences for this dose of morphine.
RESULTS
Histology and preliminary investigations
All animals that were included in the behavioral analysis had appropriate injector cannulae placements either within the VTA (Fig.1) or dorsal to the VTA for the dorsal control groups. As with all central cannulae experiments, some cell death was noted around the tip of the injector. Animals conditioned with the 500 ng per 0.5 μl per side dose (mean time spent in the saline paired environment at testing was 206.9 ± 11 sec and in the morphine-paired environment was 270.8 ± 19 sec; t= 2.6, p < 0.05) or 2500 ng per 0.5 μl per side dose (mean time spent in the saline-paired environment at testing was 173.7 ± 30 sec and in the morphine-paired environment was 340.9 ± 38 sec; t = 2.5, p < 0.05), but not the 250 ng per 0.5 μl per side dose (mean time spent in the saline-paired environment at testing was 253.2 ± 54 sec and in the morphine-paired environment was 276.1 ± 52 sec;t = 0.2, p > 0.05), showed significant place preferences for the morphine versus the saline paired environment. Therefore, the dose of 500 ng per 0.5 μl per side intra-VTA morphine was chosen as the training dose of morphine for the remaining studies.

A photomicrograph of a Nissl-stained coronal section showing representative cannulae placements into the VTA.
Animals trained previously drug-naive or while in a state of opiate dependence and withdrawal did not demonstrate significant preferences for environments paired with 500 ng per 0.5 μl per side morphine-injected 1 mm dorsal to the VTA (for the naive group, mean time spent in the saline paired environment at testing was 215.1 ± 29 sec and in the morphine-paired environment was 256.7 ± 23 sec; t = 0.86, p > 0.05; for animals trained in withdrawal, the mean time spent in the saline-paired environment at testing was 232 ± 16 sec and in the morphine-paired environment was 213.5 ± 26 sec; t= 0.5, p > 0.05). These results demonstrate that the place preferences acquired with morphine microinjections into the VTA are not the product of the drug diffusion up the cannulae tract with action at a distant dorsal site. Furthermore, the mechanisms supporting morphine’s rewarding properties within the VTA are opiate receptor-mediated. Previously drug-naive animals did not spend significantly more time in environments paired with microinjections of 500 ng per 0.5 μl per side of the inactive (+)-morphine isomer (Jacquet et al., 1977) into the VTA than in environments paired with intra-VTA saline injections (the mean time spent in the saline-paired environment at testing was 214.4 ± 39 sec and in the morphine-paired environment was 216.3 ± 34 sec; t= 0.03, p > 0.05). Together, these results demonstrate that our microinjected, just suprathreshold dose of morphine must be acting via a stereospecific opiate receptor-mediated mechanism within the VTA itself to produce its rewarding properties.
Effects of α-flupentixol on intra-VTA morphine-mediated place preferences
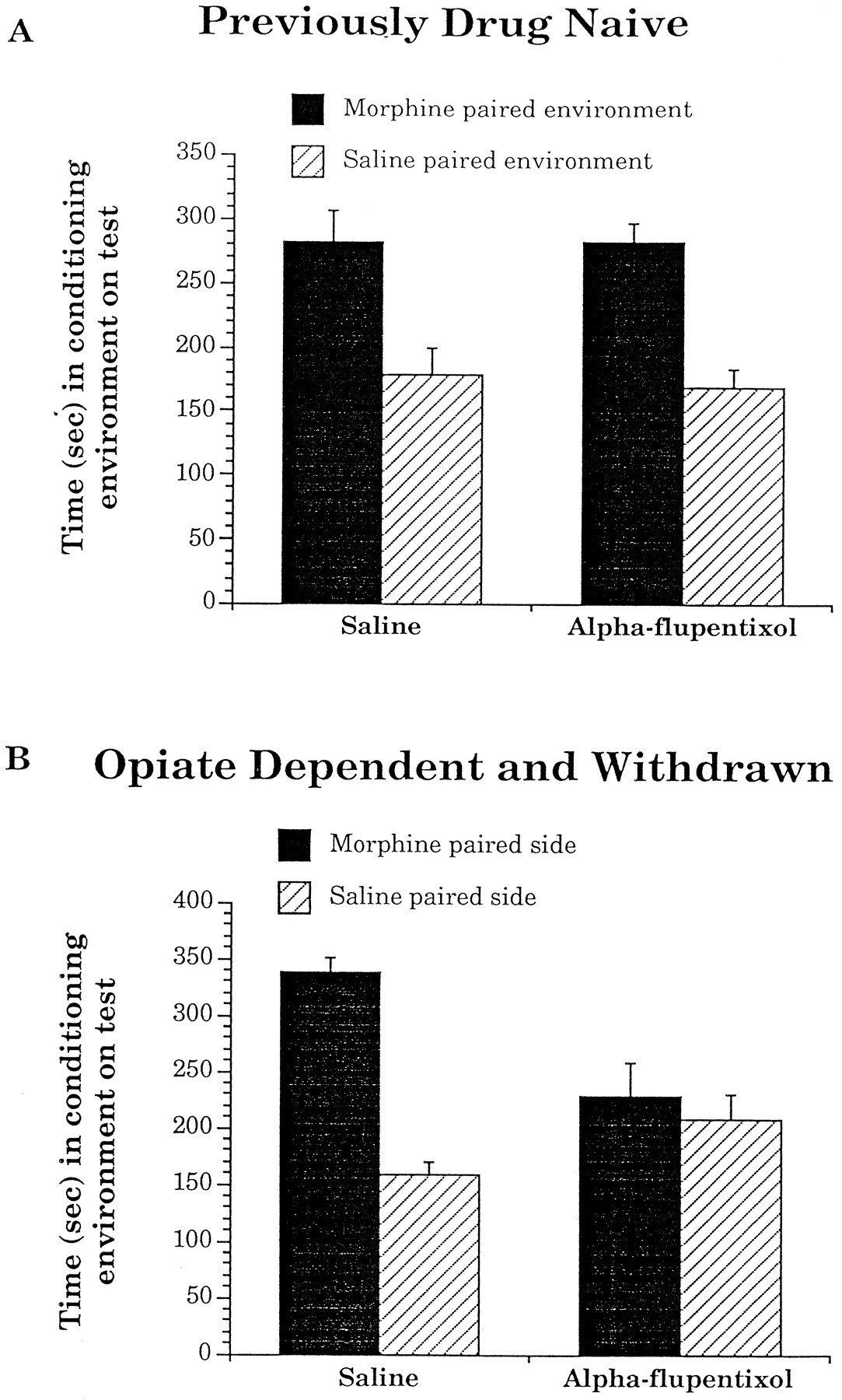
Pretreatment with a high dose of α-flupentixol (0.8 mg/kg, i.p.) 2.5 hr before each conditioning session had no effect on the acquisition of conditioned place preferences for environments paired with a low dose of intra-VTA morphine (500 ng per 0.5 μl per side) in previously drug-naive animals (Fig.2A), but completely blocked the acquisition of these same conditioned place preferences in rats trained while opiate-dependent and in withdrawal (Fig. 2B). An ANOVA that compared the effects of pretreatment (α-flupentixol vs saline) with times spent in the morphine versus saline paired environments revealed no significant interaction between these two variables for animals trained previously drug-naive (F(1,12) = 0.9, p > 0.05). There was, however, a significant main effect of time (F(1,12) = 11.9, p < 0.05), revealing the significant place preferences shown by both previously drug-naive groups. Conversely, a similar analysis on the scores of animals trained while in a state of opiate withdrawal revealed a significant interaction between pretreatment and times spent in the morphine- versus saline-paired environments (F(1,15) = 9.5, p < 0.05). These results suggest that α-flupentixol only blocked morphine’s rewarding properties in animals that were trained while in a state of opiate dependence and withdrawal.

The effects of saline or α-flupentixol pretreatment on the absolute times spent in environments paired with intra-VTA injections of either 500 ng per 0.5 μl per side morphine or 0.5 μl per side saline in separate groups of animals that were trained (A) previously drug-naive or (B) in a state of opiate dependence and withdrawal. Data are represented as mean ± SEM.
Animals conditioned initially while previously drug-naive and then again later while opiate-dependent and deprived showed the same pattern of results (Fig. 3). α-Flupentixol pretreatment had no effect on the initial acquisition of conditioned place preferences for environments paired with 500 ng per 0.5 μl per side morphine into the VTA in previously drug-naive animals (Fig.3A). An ANOVA comparing the effects of pretreatment (α-flupentixol vs saline) with times spent in the separate test environments (morphine- vs saline-paired) showed that there was no significant interaction between these two variables (F(1,12) = 0, p > 0.05). However, there was a significant main effect of time, (F(1,12) = 13.6, p < 0.05). When these same animals were subsequently retrained while in a state of dependence and withdrawal, α-flupentixol blocked the normal preferences for environments paired with 500 ng per 0.5 μl per side morphine into the VTA (Fig. 3B). An ANOVA revealed a significant interaction between the times spent in the test environments (morphine- vs saline-paired) with pretreatment (α-flupentixol vs saline) (F(1,12) = 6.1,p < 0.05). Furthermore, there were no interactions between either the environment to which morphine was paired or the pretreatment animals received the first time they were conditioned and the times spent in morphine- versus saline-paired environments on the second conditioning test (F(1,12) = 2.5,p > 0.05 and F(1,12) = 0.0,p > 0.05, respectively). Thus, even within the same animals, pretreatment with a broad-spectrum dopamine antagonist only blocked morphine’s rewarding properties in animals that were in an opiate-dependent and withdrawn state, but not when they were drug-naive.

The effects of saline or α-flupentixol pretreatment on the absolute times spent in environments paired with intra-VTA injections of either 500 ng per 0.5 μl per side morphine or 0.5 μl per side saline in animals that were initially trained previously drug-naive (A) and then subsequently retrained while in a state of opiate dependence and withdrawal (B). Data are represented as mean ± SEM.
Effects of TPP lesions on intra-VTA morphine-mediated place preferences
All animals included in the behavioral analyses of this experiment had the majority of the ventromedial TPP bilaterally lesioned (Figs.4, 5). Histological analyses revealed that ∼50% of the ibotenic acid-lesioned rats had lesions with necrotic, vacuolated centers. Such a phenotype may be the result of the long interval between surgery and histology (Brown and Fibiger, 1993), which in the present case was ∼3 months. Lesions of the TPP blocked the acquisition of intra-VTA morphine place preferences in previously drug-naive, but not opiate-dependent and withdrawn, animals (Fig.6). For animals trained previously drug-naive, there was a significant interaction of surgery (sham- vs TPP-lesioned) with times spent in the morphine versus saline paired environments (F(1,15) = 8.8, p < 0.05; Fig.5A). A similar analysis on the scores of animals trained while in a state of opiate withdrawal showed that there was no interaction between these two variables (F(1,13)= 0.1, p > 0.05; Fig. 5B). There was, however, a significant main effect of times spent in the morphine- versus saline-paired environments (F(1,13) = 6.4, p < 0.05), demonstrating that both withdrawal groups showed significant conditioned morphine place preferences. Given that some of the TPP-lesioned rats had necrotic lesions, we also compared the scores of TPP-lesioned rats with and without necrotic lesions. There were no significant interactions between lesion type (necrotic vs non-necrotic) with times spent in the morphine- versus saline-paired environments for animals trained either previously drug-naive (F(1,6) = 0.34, p > 0.05) or in a state of opiate dependence and withdrawal (F(1,5) = 0.05, p > 0.05). Thus, any damage to fibers of passage did not produce any additional behavioral deficit. These results suggest that lesions of the TPP blocked intra-VTA morphine’s rewarding properties in previously drug-naive, but not in opiate-dependent and withdrawn, rats.

A, Photomicrograph of a Nissl-stained coronal section showing the extent (arrows) of a representative bilateral ibotenic acid induced lesion of the TPP. B, Higher-magnification photomicrograph of the left side of the ibotenic acid-induced lesion shown in A. The extent of the lesion is indicated byarrows, whereas a healthy neuron is indicated by anarrowhead. C, High-magnification photomicrograph of the left side of a sham lesion.

A schematic representation of the size and extent of the ibotenic acid lesions of the ventromedial TPP. Thefilled-in areas represent the volume of the smallest lesion. The open outlined areas represent the volume of the largest lesion. The numbers to theright indicate the distance caudal from Bregma in millimeters.

The effects of bilateral sham or ibotenic acid lesions of the TPP on the absolute times spent in environments paired with intra-VTA injections of either 500 ng per 0.5 μl per side morphine or 0.5 μl per side saline in separate groups of animals that were trained (A) previously drug-naive or (B) in a state of opiate dependence and withdrawal. Data are represented as mean ± SEM.
The sham- and TPP-lesioned animals that were trained in a state of opiate withdrawal were allowed to recover from dependence and were retrained with a 10 mg/kg systemic dose of morphine. Two rats were removed from this final portion of the study because of sickness. TPP-lesioned rats that had acquired previously conditioned place preferences for environments paired with intra-VTA morphine while in opiate withdrawal did not acquire conditioned place preferences for environments paired with systemic morphine after recovering from dependence. An ANOVA on the scores of the remaining rats comparing the effects of lesions (TPP or sham) with times (morphine- vs saline-paired environment) revealed a significant interaction between these two variables (F(1,11) = 5.3, p < 0.05). For the sham-lesioned group, the mean time spent in the saline-paired environment at testing was 166 ± 19.6 sec and in the morphine-paired environment was 305 ± 27.3 sec. For the TPP-lesioned group, the mean time spent in the saline-paired environment at testing was 235 ± 24.4 sec and in the morphine-paired environment was 215 ± 24.2 sec. Thus, in the same animals that were trained initially in a state of opiate withdrawal and then subsequently retrained after having recovered from dependence, lesions of the TPP only blocked opiate reward when animals are in nonwithdrawal states.
DISCUSSION
Lesions of the TPP and α-flupentixol pretreatment double-dissociate two independent motivational substrates within the VTA itself. TPP lesions, but not α-flupentixol pretreatment, blocked the rewarding effects both of intra-VTA morphine in animals that were conditioned previously drug-naive and of systemic morphine in animals that had recovered from dependence. Conversely, when animals were conditioned in a state of opiate dependence and withdrawal, the opposite pattern of results was found. α-Flupentixol pretreatment, but not TPP lesions, blocked the rewarding properties that resulted from morphine microinjections into the VTA. Furthermore, even in single animals that were initially conditioned previously drug-naive and then retrained while in a state of opiate withdrawal, α-flupentixol’s reward-blocking effects were constrained to when animals were in withdrawal. Thus, by manipulating only the state in which animals are conditioned but leaving constant the training dose, conditioning protocol, pretreatment, and variation in cannulae placement (by using the same animals, first trained in one state, then counterbalanced and subsequently retrained in the opposite state), we have demonstrated that two separate neural systems in the VTA can be engaged in predictable ways to mediate the motivational properties of morphine. Training animals while in a state of opiate dependence and withdrawal (deprivation) will cause morphine’s rewarding properties to be dopamine-mediated. On the other hand, training animals while in non-opiate-withdrawal states (nondeprivation) causes morphine’s rewarding properties to be TPP-dependent.
The behavioral deficits of α-flupentixol pretreatment in opiate-dependent and withdrawn rats and of TPP lesions in previously drug-naive animals are not attributable to nonspecific sensory, motor, or learning deficits. α-Flupentixol pretreatment in previously naive rats or TPP lesions in opiate-dependent and withdrawn rats had no effects on the normal acquisition of conditioned place preferences, demonstrating that these manipulations did not prevent animals from perceiving the environment and learning an association between the environment and morphine’s rewarding properties.
The most obvious candidate pathway mediating the dopamine-dependent system is the mesolimbic pathway. Indeed, recent unpublished results from our laboratory suggest that the acquisition of conditioned place preferences in opiate-dependent and withdrawn rats can be antagonized by α-flupentixol microinjections into the nucleus accumbens. Although the pathway mediating morphine reward in naive animals is not known, the most direct route is for morphine to act on opiate receptors on nondopaminergic VTA neurons (Johnson and North, 1992) that descend to the TPP (Semba and Fibiger, 1992; Steininger et al., 1992). This view is in contrast to the current focus on ascending catecholamine systems as the basis of reward (Koob, 1992; Wise, 1996).
An alternate interpretation of the present data is that the reported dissociation is quantitative, not qualitative, in nature. This interpretation posits a process of sensitization as being the critical variable underlying our dissociation. In this case, TPP lesions that are normally sufficient to block morphine’s rewarding properties in opiate-naive animals are ineffective in opiate-dependent and withdrawn rats because the pathway mediating morphine reward has been sensitized by chronic opiate administration. To antagonize this enhanced reward, interference of dopamine transmission is required. However, this interpretation is untenable for two reasons. First, interference with dopamine transmission that is sufficient to block the sensitized opiate reward present in dependent and withdrawn animals should have been sufficient to block the weaker opiate reward in previously drug-naive animals. The inability of the dopamine antagonist to block opiate reward in previously drug-naive rats suggests the dissociation is qualitative, not quantitative, in nature. Second, sensitization is a long-lasting process (Kalivas and Stewart, 1991; Robinson and Berridge, 1993). Indeed, studies have reported that the mesolimbic pathway remains hyper-responsive to morphine a month after the termination of initial opiate administration (Spanagel et al., 1993). This interpretation predicts that once the mesolimbic pathway has been sensitized, then morphine reward should continue to be dopamine-mediated regardless of whether rats are in a state of deprivation. Our results showing that TPP lesions blocked the acquisition of conditioned place preferences for environments paired with systemic morphine in rats that had recovered from dependence contradict this prediction. Furthermore, the behavioral effects of TPP lesions appear to be independent of reward magnitude because lesions of the TPP block the rewarding properties of systemic morphine over the entire dose range usable in drug-naive animals, but have no effect on the rewarding properties of a low, 2 mg/kg dose of morphine in opiate-dependent and withdrawn rats (Bechara and van der Kooy, 1989,1992a,b). Thus, we suggest that the critical qualitative difference in the processes being mediated by these two systems is the production of reward in the absence or presence of opiate dependence and withdrawal.
According to our model, dopamine should not play any role in mediating the rewarding properties of opiates in drug-naive animals. However, there are a number of previously reported findings with both heroin (Bozarth and Wise, 1981; Spyraki et al., 1983; Hand et al., 1989) and morphine (Phillips et al., 1983; Leone and Di Chiara, 1987; Shippenberg and Herz, 1988; Acquas and Di Chiara, 1994) that contradict this hypothesis. Although the majority of these studies can be questioned on procedural grounds (see Nader et al., 1994), one study that used an unbiased place conditioning paradigm has reported that unilateral intra-accumbens injections of SCH 23390, at a dose that has no unconditioned motivational effects itself, blocked the acquisition of low-dose systemic morphine place preferences in drug-naive animals (Shippenberg et al., 1993). Based on this finding, it could be argued that morphine’s rewarding properties are D1-mediated and, therefore, that SCH 23390 (a specific D1 antagonist) is more effective than α-flupentixol (a broad-spectrum antagonist) in blocking morphine’s motivational properties. This interpretation is unlikely, however, given that α-flupentixol is more effective than SCH 23390 at inhibiting dopamine-stimulated adenylate cyclase activity (a measure of D1 receptor activity) (Hyttel, 1978, 1984). Furthermore, it is difficult to reconcile the unilateral intra-accumbens SCH 23390 blockade of conditioned place preferences for environments paired with systemic morphine with the recent findings that lesions of the entire ventral striatum do not block the acquisition of conditioned place preferences for morphine-paired environments (Olmstead and Franklin, 1996).
A neurobiological distinction between nondeprived and deprived states may clarify some of the conflicting results obtained with heroin self-administration. The nondeprived/deprived model predicts that the acquisition of opiate self-administration (when animals have had minimal drug exposure) will be blocked by lesions of the TPP but will be insensitive to dopamine antagonist pretreatment. Consistent with these predictions are the findings that lesions of the TPP (Olmstead et al., 1993), but neither dopamine antagonist pretreatment nor 6-OHDA lesions of the nucleus accumbens (van Ree and Ramsey, 1987; Gerrits et al., 1994; Gerrits and van Ree, 1996), blocked the acquisition of heroin self-administration.
Previous findings have shown that the amount of heroin self-administered in order to attain a stable baseline of responding is sufficient to induce states of dependence (Nader et al., 1994). Thus, the nondeprived/deprived model predicts that once animals have acquired a stable baseline of operant responding, then dopamine antagonist pretreatment, but not TPP lesions, will block operant responding for heroin. Indeed, TPP lesions have no effect on the rate of heroin self-administration in animals that previously had acquired a stable baseline (Nader et al., 1994). The findings with dopaminergic manipulations on the maintenance of heroin self-administration are more controversial. Under conditions in which animals have acquired a stable baseline rate of responding for heroin, evidence from different studies suggests that the reinforcing effects of heroin are affected by dopaminergic manipulations (presumably because of opiate actions in the VTA) (Britt and Wise, 1983; Nakajima and Wise, 1987) or are not affected by dopaminergic manipulations (possibly because of actions in the nucleus accumbens) (Ettenberg et al., 1982; Pettit et al., 1984). We can imagine three reasons why 6-OHDA lesions of the nucleus accumbens may not block the maintenance of heroin self-administration (Pettit et al., 1984). First, it is possible that when the dopaminergic systems are challenged, opiates can sometimes still act on receptors located efferent to dopaminergic neurons in the deprived motivational system. Second, it is possible that the lesions did not decrease dopamine levels sufficiently. Third, 6-OHDA lesions may have blocked the primary motivational properties of heroin, but the amount of heroin self-administered due to conditioned responding alone was sufficient to take animals from the deprived state to the nondeprived state. Once in nondeprived states, 6-OHDA lesions of the nucleus accumbens should not affect self-administration behavior because heroin’s rewarding properties are no longer dopamine-dependent.
In conclusion, we have demonstrated that two independent motivational systems exist within the VTA itself, the differential activation of which is predicated on whether animals are in states of withdrawal (deprivation) or not (nondeprivation). A dopamine-independent mechanism mediates the rewarding properties of morphine only when animals are in nondeprived opiate states and is dependent on the TPP nucleus. In animals that are in opiate-deprived states, a dopaminergic system is engaged to mediate morphine reward. Furthermore, the psychological boundary between nondeprived and deprived states identified with intra-VTA morphine in the present study is consistent with a similar boundary between nondeprivation and deprivation states using neurobiologically manipulations of the reward produced by food, systemic morphine, and systemic heroin (Bechara and van der Kooy, 1992a; Bechara et al., 1992; Nader et al., 1994; Nader and van der Kooy, 1996).
Footnotes
This work was supported by a grant from the Medical Research Council of Canada. We thank Dr. Thomas Jaeger for his assistance in preliminary dose–response studies.
Correspondence should be addressed to Karim Nader, Neurobiology Research Group, Department of Anatomy and Cell Biology, University of Toronto, Toronto, Ontario, Canada M5S 1A8.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}