

Abstract
We used a behavioral learning paradigm followed by electrophysiological analysis to find sites in theLymnaea feeding network in which electrical changes could be recorded after appetitive conditioning. Specifically, we analyzed conditioning-induced changes in cellular responses in the mechanosensory conditioned stimulus (CS) pathway, in the central pattern generator (CPG) network, and in feeding motoneurons. During training, experimental animals received 15 pairings of lip touch (the CS) with sucrose (the unconditioned stimulus, US). Control animals received 15 random CS and US presentations. Electrophysiological tests on semi-intact preparations made from conditioned animals demonstrated a network correlate of the overall feeding conditioned response, a touch-evoked CPG-driven fictive feeding rhythm. At the motoneuronal level, we found significant conditioning-induced increases in the amplitude of an early touch-evoked EPSP and spike activity, recorded from the B3 feeding motoneuron. Increases in EPSP amplitude and motoneuronal spike activity could occur independently of conditioned fictive feeding. These changes in response recorded at the level of CPG interneurons, and motoneurons were preceded by changes recorded in the CS pathway. This was demonstrated by recording a conditioning-induced increase in the number of touch-evoked spikes in the cerebrobuccal connective, which forms part of the CS pathway. The finding that electrophysiological changes after conditioning can be recorded at multiple sites in this simple system provided an important intermediate level of analysis between whole animal behavior and cellular studies on the synaptic sites of plasticity.
- classical conditioning
- memory trace
- cellular plasticity
- feeding behavior
- invertebrates
- semi-intact preparations
- Lymnaea
One successful approach in the search to understand the cellular basis of learning in both vertebrates and invertebrates (for review, see Byrne, 1987) is based on subjecting intact animals to conditioning and then recording learning-induced changes in the nervous system. The aim is to establish direct links between behavioral and cellular/molecular events. Ideally, plastic changes should be studied at the level of specific synapses between identified neurons. We have used this approach to study appetitive conditioning in the pond snail, Lymnaea stagnalis.
In molluscs, the neuronal basis of appetitive conditioning has been relatively little studied compared with aversive paradigms (for review, see Carew and Sahley, 1986; Byrne, 1987; Glanzman, 1995). So far neuronal correlates of appetitive conditioning only have been demonstrated in Lymnaea (Whelan and McCrohan, 1996; Kemenes et al., 1997; Staras et al., 1998b), although recent studies show promise for successful cellular analyses of nonaversive learning in other molluscs as well (Peschel et al., 1996; Colwill et al., 1997). We have previously shown the survival of a neurophysiological trace (fictive feeding) of an appetitively conditioned feeding response to touch (Kemenes and Benjamin, 1989a) in a semi-intact Lymnaeapreparation (Staras et al., 1998b). This was an important finding because it established that behaviorally produced appetitive learning is amenable to cellular analysis in preparations made from the same animals. The same type of preparation was used in the present work to examine more detailed neural changes in the feeding circuit, after appetitive conditioning.
The feeding circuit of Lymnaea is more complex than those involved in simple defensive reflexes used in aversive paradigms. It consists of sensory neurons, motoneurons, central pattern generating interneurons (CPG), and modulatory neurons, whose synaptic connectivity is known in great detail (Benjamin and Elliott, 1989). Studies of appetitive conditioning in another complex invertebrate system, the honeybee (for review, see Hammer, 1997), revealed that changes associated with learning could be recorded in various structures within the CNS. This and other studies on vertebrates (Lisberger, 1998) emphasized the importance of studying the whole neural system involved in learning. This intermediate level of analysis was the one adopted here. We had no previous knowledge of the parts of theLymnaea circuit that were likely to express changes occurring after behavioral conditioning, so electrical responses were recorded at different sites within the network as a prelude to more detailed studies on the specific synaptic sites of plasticity. We will show that electrophysiological correlates of learning can be recorded at multiple sites within the feeding network. As well as increasing the CPG-driven fictive feeding response to the CS, training also increased the strength of a specific synaptic response in an identified motoneuron and induced an increase in early spike activity in the CS pathway.
MATERIALS AND METHODS
Experimental subjects
Wild-type specimens of adult Lymnaea stagnalis, obtained from animal suppliers (Blades Biological, Kent, UK), were kept in groups in large holding tanks containing Cu2+-free water at 18–20°C on a 12 hr light/dark cycle and fed lettuce three times a week. At least 1 week before behavioral experiments, the animals were kept in a subsatiated state in 2 l tanks in the laboratory. The water in the tanks was replaced daily in the pretraining period and throughout the training procedure. All lettuce was removed from the home tank at least 2 hr before testing or training. These maintenance procedures were the same as those described in a recent Lymnaea conditioning study (Staras et al., 1998b).
Behavioral experiments
Appetitive classical conditioning. Behavioral experiments were performed to establish conditioned feeding responses to a tactile CS in intact animals before testing cellular responses in semi-intact preparations made from the same animals. In these experiments, we conditioned three groups of a total of 36 experimental animals, and these were compared both in vivo and in vitro with three corresponding groups of a total of 35 random control animals. This is the most important control in this type of experiment, because it includes the use of both CS and US. The lack of a conditioned response in other types of controls, such as CS alone, US alone, and handling, also has been demonstrated in earlier experiments using touch as a CS and sucrose as a US in Lymnaea (Kemenes et al., 1989a; Staras et al., 1998b).
The classical conditioning procedure in all three training experiments was performed as follows: subjects from the experimental groups were removed from their home tank and placed in individual training dishes. Two minutes after transfer, each snail was given a touch stimulus to the lips (the CS) delivered by a hand-held probe. The duration of this stimulus was ∼1 sec. This was followed immediately (interstimulus interval, <1 sec) by the presentation of sucrose (the US) from a 10 ml syringe into the water surrounding the animals (0.01m final concentration). Previous work demonstrated that sucrose at this concentration evokes strong unconditioned rhythmic feeding movements (rasps) in Lymnaea (Kemenes et al., 1986;Staras et al., 1998b). After 2 min in sucrose, the snails were moved to a clean water rinsing tank. After 5 min rinsing, all snails were transferred back to their home tanks. In random CS–US controls, the procedures were identical except that the intervals between CS and US were varied between 0 and 30 min, and the order (CS–US or US–CS) was randomized. In each trial a single random order was used for all control animals.
Both experimental and control animals received five trials a day with 60 min intertrial intervals. Training continued for 3 d so that a total of 15 trials were performed in a spaced training regimen. In all control and experimental groups the animals received a small amount of lettuce in the evening of each day. After the 15th trial, animals were placed in clean water and left undisturbed for at least 2 hr, before testing them on the same day as the final training trials.
Testing the conditioned response. Testing was performed using a blind procedure, by an experimenter who had no knowledge of the training history of the animals being tested. During testing an animal was removed from the tank and placed in a test dish containing 100 ml of water, and all subsequent feeding events were recorded with custom written event-recording software. Two minutes after emergence, the tactile stimulus was presented to the lips, and the feeding activity was recorded for a further 2 min. The animal was removed, numbered using an indelible ink marked onto the shell, and replaced in the tank. The training and test protocols described here have been reported in a previous Lymnaea conditioning study (Staras et al., 1998b).
Statistical analysis of behavioral data. The feeding responses to touch were quantified by awarding a behavioral score to each animal in a blind procedure. The score was calculated by subtracting the rasp rate for the minute preceding the touch CS from the rasp rate for the minute after the first rasp after the same stimulus. If no rasps occurred within 1 min after the stimulus, the post-touch count was taken as 0. This scoring procedure is based on a method previously used by Kemenes and Benjamin (1994) and Staras et al. (1998b) in appetitive learning experiments with Lymnaea.
As in previous work on Lymnaea learning (Audesirk et al., 1982; Whelan and McCrohan, 1996; Staras et al., 1998b), nonparametric statistical tests were used. This allowed us to avoid making assumptions about the normal distribution of the data. Also, conclusions based on nonparametric statistics are more robust given that they are less sensitive than parametric tests. For one type of nonparametric analysis (χ2 test), we compared the proportions of experimental and control animals giving a positive response to the CS (increase in feeding rate) versus the proportion of animals in the same groups not giving a positive response (decrease in feeding rate or no change in feeding rate after CS). Further nonparametric comparisons (Mann–Whitney U tests) were made using the CS-evoked feeding rasp scores in all experimental and control animals.
Electrophysiological experiments
The aim of these experiments was to record cellular changes in semi-intact preparations resulting from behavioral conditioning in intact snails.
General procedures. Random control and conditioned animals were dissected in HEPES-buffered snail saline (Benjamin and Winlow, 1981). All the subsequent electrophysiological tests were performed by a second experimenter who had no knowledge of the behavioral history of the individual preparations. The semi-intact preparation (Fig. 1A) was transferred to a Sylgard-coated electrophysiology chamber (volume 2, 3 ml) which contained saline and usually pinned dorsal-side up. The outer ganglionic sheath of the cerebral and buccal ganglia was removed surgically and the second, inner sheath was softened using a nonspecific solid protease (Sigma XIV; Sigma, Poole, UK).

The semi-intact preparations used to record neurophysiological feeding responses to a conditioned tactile stimulus. The lips are connected to the CNS by peripheral lip nerves. Rhythmic bursting activity in the feeding CPG, which underlies rhythmic feeding movements in whole animals, can be monitored by making intracellular recordings from identified buccal motoneurons, such as B3. A tactile stimulus (CS) could be presented to the lips using a switch-operated probe. A, Arrangement of the preparation for intracellular recording of CPG-driven fictive feeding activity and synaptic inputs to an identified motoneuron, B3. Note that the cerebrobuccal connective (B, CBC) is twisted to allow exposure of the dorsal side of the buccal ganglia for impalement of B3 with a microelectrode. B, Arrangement of the preparation for extracellular recording of signals travelling on the CBC after tactile stimulation of the lips.
Whole-lip CNS preparation. For these experiments we used preparations in which the lip sensory structures, the median and superior lip nerves, and the CNS were left completely intact (for more details of this type of preparation, see Staras et al., 1998b). The preparations were arranged in the electrophysiology chamber so that the lip structures were accessible for stimulation, and the buccal ganglia and cerebrobuccal connectives (CBCs) were accessible for electrophysiological recording (Fig. 1).
Intracellular recording techniques. Glass microelectrodes (2 mm; Clark Electromedical, Redding, UK) were pulled to a tip resistance of 10–50 MΩ when filled with 4 m potassium acetate. Micromanipulators with attached headstage preamplifiers (Neurolog; Digitimer, Welwyn Garden City, UK) were arranged around the perfusable electrophysiology chamber, permitting up to four simultaneous intracellular recordings. Signals were fed into amplifiers (model NL102G; Digitimer) incorporating a bridge-balance circuit for current injection and then sent to a storage oscilloscope (Gould model 1604; Gould Instrument Systems, Ltd., Hainault, UK) and a chart recorder (Gould model TA240S). All signals were recorded digitally using a DAT recorder (Biologic model DTR-1801; Biological Science Instruments, Claix, France).
Identification and selection of cell types. The main objective of the first type of electrophysiological experiment reported in this work was to intracellularly monitor neuronal activity in semi-intact preparations known to underlie feeding responses in whole animals. This neuronal activity, which is called fictive feeding, is generated by a set of premotor CPG interneurons (Rose and Benjamin, 1981b; Elliott and Benjamin, 1985). These neurons belong to three main types, N1, N2 and N3, each active in one of the three behavioral phases of feeding, protraction (N1), rasp (N2), and swallow (N3). However, recording activity directly from these small interneurons in semi-intact preparations is technically difficult, and impalement with microelectrodes often triggers nonspecific excitation in them, making quantitative assessments of fictive feeding responses more difficult (Staras et al., 1998b). An alternative and less invasive approach is to monitor CPG activity indirectly using identified buccal motoneurons that receive well characterized monosynaptic inputs from the CPG during each phase of fictive feeding (Benjamin and Elliott, 1989; Staras et al., 1998a). These large motoneurons could be located visually by size, position, and color so that they could be impaled with minimal disturbance to the rest of the system. In these experiments the identified buccal motoneuron B3 was used as the main cell type to monitor fictive feeding because it is both large and has a very characteristic firing pattern. Previous work has established that the B3 cell is inhibited during N1 but is excited during both N2 and N3 because of inputs from the appropriate CPG interneurons (Benjamin and Rose, 1979; Rose and Benjamin, 1981a,b; Elliott and Benjamin, 1985). By recording the B3 we could obtain information on activity in all three classes of feeding CPG interneurons in response to tactile stimuli as well as nonpatterned synaptic input arising directly from the tactile CS. It was important to be able to record changes both at the level of the CPG and the motoneurons, and recording B3 enabled both these objectives to be fulfilled. In addition to B3, in all experiments in which touch-evoked fictive feeding was assessed, either the B4 or B1 motoneuron, or both, were also recorded. The B4 cells are inhibited during N1 and the first phase of N2 but excited during the second phase of N2 and during N3, whereas the B1 cells are excited during N1 and silent in the N2 and N3 phases (Benjamin and Rose, 1979; Rose and Benjamin, 1981a,b; Elliott and Benjamin, 1985). Recording from these cells aided the unequivocal establishment of the occurrence of fictive feeding cycles that were primarily monitored on the B3 motoneuron (Fig.1A). The procedures described here also have been reported in a previous Lymnaea conditioning study (Staras et al., 1998b).
Extracellular recording. In a second type of electrophysiological experiment it was necessary to extracellularly record spike activity on the CBCs, which link the buccal ganglia with the rest of the CNS and the lips. A special electrophysiology chamber was used, in which two large Sylgard-lined compartments were separated by an empty trough with a slot in each partition wall. A whole-lip preparation (Fig. 1B) was placed in the chamber containing normal HEPES, and the head structures were pinned firmly on one of the large Sylgard compartments. The buccal ganglia were pulled through the slots and pinned down in the other compartment, so that CBCs spanned the gap. The saline was removed from all compartments and the slots, through which the CBCs ran, were rapidly sealed with a silicon grease. Two platinum hook electrodes mounted on a small manipulator were placed on one of the CBCs. The trough was filled with paraffin oil to prevent the nerve drying out, and the two large compartments were refilled with a modified High Mg2+, High Ca2+ (HiDi) saline (see below). The extracellular signals were fed into an AC extracellular amplifier (Neurolog) and displayed and stored using the same methods used for intracellular recording.
There appear to be a group of nerve fibers in the CBCs that carry sensory information from the lips to the buccal ganglia where the feeding motoneuronal and interneuronal targets for the touch-activated input are located. Tactile responses still occur in these fibers when the preparation is bathed in HiDi (see Fig. 5Bii). This type of saline is known to increase the threshold of intermediate neurons in polysynaptic pathways (for the composition of this saline, see Elliott and Benjamin, 1989). In the present experiment it reduced the level of both spontaneous and touch-evoked extracellular activity, making it likely that the majority of remaining early signals in the CBC were arising from the CS sensory pathway. The bathing of the whole preparation, including the cerebral ganglia, in HiDi made it more probable that these signals were travelling directly from the lips. Therefore, these early tactile responses on the CBCs were more likely to represent the activity of primary or secondary components of the CS pathway. This was further supported by data from pilot experiments with preparations from naive animals (n = 5) in which normal saline was used and which, therefore, allowed both CBC spikes and buccal synaptic activity to be recorded simultaneously. These experiments showed that the earliest component of the tactile response in the CBCs occurred within 50 msec of touch to the lips and always preceded any touch-evoked synaptic potentials seen on neurons of the buccal feeding network by at least 20 msec.
The preparations used for the extracellular recordings of the CBCs were sufficiently stable to allow repeated responses to the CS to be recorded from control and experimental animals that had been subjected to the same behavioral conditioning procedures described for the intracellularly recorded responses in the previous experiments. Measurements made from each control and experimental animal were based on the average of four repeated presentations. Previous behavioral work (Kemenes and Benjamin, 1994) has shown that four repeated presentations of the CS alone does not result in extinction of the conditioned response, and it seemed reasonable to assume that this was also likely to be the case in the reduced preparations.
Tactile stimulation of the lips in semi-intact preparations.In the semi-intact preparations, we tried to reproduce the tactile stimulus used in intact animals as closely as possible. The only difference was that instead of the hand-held probe that had to be used in the freely moving intact animals, we used an electromagnetic coil-driven mechanical probe (Fig. 1) to deliver 1 sec duration tactile stimuli of a consistent force to the lips in the semi-intact preparations. However, the end of the probe consisted of the same thin wedge of soft, flexible plastic that was used in the behavioral experiments, and this hit the same target area on the lips in the preparations as in the intact animals. Like the behavioral tests, the neurophysiological experiments on trained animals were performed blind so that the presentation of the tactile CS to a preparation was performed without the identity of the subject (control or conditioned) being revealed. All electrophysiological experiments with previously trained animals were completed within 16–36 hr after the last training trial with a dissection schedule designed to ensure that the range of intervals between training and electrophysiological tests were the same for experimental and control groups. We assumed that there was no loss of the memory trace in the time interval between the behavioral and electrophysiological tests because previous experiments have shown that appetitive conditioning with 15 trials produces long-term memory and that experimental snails tested up to 3 d after the last training trials show conditioned responses that are not significantly different from the responses seen several hours after the end of training (Kemenes and Benjamin, 1994).
Analysis of electrophysiological data. All the analysis was performed blind; the person analyzing records from individual preparations was unaware of their status as experimental or control animals.
After experiments involving intracellular recording, the number of fictive feeding cycles (full sequences of N1, N2, and N3 phase activity) occurring on the B3 (and when these were also recorded, on the B1 and B4) motoneurons of the semi-intact preparations were counted for 1 min before the application of a touch (CS) to the lips and for 1 min after the first cycle after the stimulus. As in the behavioral tests, if no fictive feeding cycle occurred within 1 min after the stimulus, the post-touch count was taken as 0. The difference between the numbers of poststimulus and prestimulus cycles yielded a score that was used as a measure for conditioned fictive feeding. The same nonparametric statistical methods were used to analyze this electrophysiological data as had previously been used for the whole animal behavioral data.
In each experiment, the peak voltage of the touch-evoked B3 EPSP was measured against the prestimulus membrane potential, and this was used to quantify the EPSP amplitude. The amplitude data were compared by unpaired nonparametric statistical tests in the experimental and control groups.
For statistical comparisons of B3 resting potential and the latency and duration of the touch-evoked EPSP response, all normally distributed variables, parametric tests were used.
After the extracellular recording experiments, the number of touch-evoked spikes on the CBC were counted and compared by parametric statistical tests between experimental and control preparations.
We were aware of the potential importance of checking the experimental against both a random and an untrained control group in experiments of this type but decided only to include the most important control group (random) to allow more time for detailed electrophysiological experiments. Comparing experimental preparations with random control ones only was justified because our previously published behavioral experiments (Kemenes and Benjamin, 1989a; Staras et al., 1998b) had shown no difference between the random and untrained control group, whereas both showed the same difference when compared with an experimental group in the same experiment. Also, pilot experiments recording B3 had shown no difference in the touch response in these two groups. This validated the design of the present experiments and allowed us to measure more parameters of the conditioned and control responses in a larger number of snails. A practical reason for this design was that in each experiment, the time available between the last trial and the electrophysiological tests was limited to <36 hr, after which time the conditioned response was known to decline sharply (Kemenes and Benjamin, 1994). This restricted the number of different groups we could test in any one experiment to two (given the statistical requirement to electrophysiologically test as many preparations in each group as possible). Because of potential batch differences (our unpublished observations) we did not think that it was justifiable to use an untrained control group from a different experiment for comparison with the experimental and random control groups we trained and tested together in the present experiments.
RESULTS
Appetitively conditioned animals show increased CPG-driven feeding responses to the CS both in vivo and in vitro
After 15 conditioning trials, 89% of the experimental animals used in the first experiment (n = 18) showed an increased behavioral feeding response to the CS (defined as the difference between the number of pre-CS and post-CS feeding rasps per time unit). Eleven percent of the experimental animals showed inhibition of ongoing weak spontaneous rasping (Fig.2Ai). By contrast, only 37% of the random control animals (n = 19) showed an increase in rasping rates after the application of CS after training. In 47% of the control animals, the response to CS was inhibitory, and 16% completely failed to respond to touch (Fig.2Ai). The proportion of animals that showed spontaneous rasping before touch versus the proportion of animals that did not was not significantly different between the control and the experimental group (χ2 = 1.79; df = 1;p = 0.1). By contrast, the proportion of animals responding with an increase in feeding rate to the touch CS versus the proportion of animals not showing an increase (i.e., showing either no response or a negative response) was significantly greater in the experimental than in the control group (χ2 = 10.7; df = 1; p < 0.01).

Behaviorally (A) and electrophysiologically (B) recorded feeding responses to a conditioned touch stimulus (CS). Ai, Percentage distribution of intact animals in the experimental and control group showing an increase (+ response), a decrease (− response), or no change (no response) in feeding rate after the application of the CS. The experimental group has a much higher proportion of animals showing an increased feeding rate after touch than the control group (for detailed statistical data, see Results). Aii, The change in feeding rate after tactile presentation in the experimental group is significantly larger than the change in the control group (for detailed statistical data, see Results). Data are shown as medians and interquartile ranges of data from all experimental and control animals.Aiii, Example of a feeding response to lip tactile stimulation in an appetitively conditioned intact animal. Thevertical lines represent individual rasps on a continuous time base (horizontal line).Aiv, Example of the lack of a feeding response to lip tactile stimulation in a control subject. Bi, Percentage distribution of preparations from animals in the experimental and control group showing an increase (+ response), a decrease (− response), or no change (no response) in fictive feeding rate after the application of the CS. These preparations were made from the same groups of animals for which behavioral data are shown in A. The experimental group has a much higher proportion of preparations showing an increased fictive feeding rate after touch than the control group (for detailed statistical data, see Results). Bii, The change in fictive feeding rate after tactile presentation in the experimental group is significantly larger than the change in the control group (for detailed statistical data, see Results). Data in Bii are shown as medians and interquartile ranges of data from all experimental and control preparations (including nonresponding ones).Biii, Example of a fictive feeding response to tactile stimulation of the lip in a semi-intact preparation made from an appetitively conditioned whole animal. The touch-evoked feeding activity is monitored by intracellular recording from the identified rasp-phase feeding motoneuron B3. The touch stimulus in this preparation evoked three long CPG-driven fictive feeding cycles (indicated by dots), two of which occurred within a minute after the onset of the first cycle. Biv, Lack of a fictive feeding response to a lip tactile stimulus in a semi-intact preparation derived from a control animal. The lack of cyclical synaptic input after the lip touch shows that in this preparation the stimulus is not effective in triggering CPG-driven fictive feeding activity.
The median rate of the feeding response to the touch CS in the experimental group was 7.0 rasps/min [interquartile range (IQR), 3.5–11.3], significantly greater than the median rate of the response in the control animals (0.0 rasps/min; IQR, −2.0–4.2; Mann–WhitneyU test; U = 61.5; p < 0.001) (Fig. 2Aii). Examples of typical feeding responses (or the lack of them) to the touch stimulus in experimental and control animals are shown in Figure 2, Aiii andAiv.
After an overnight period after training in which the animals had no access to food, they were dissected and prepared for electrophysiological experiments. The person dissecting the animals was not aware of their individual behavioral scores, and the animals to be dissected were drawn randomly from the experimental and control groups. Successful electrophysiological tests were performed on 13 experimental and 12 control semi-intact lip-CNS preparations made from the same groups of animals that were used in the behavioral experiments. Technical problems, mainly caused by damage occurring during dissection, prevented all the behaviorally tested animals being used for electrophysiology.
To measure changes in CPG activity after training, fictive feeding responses to two touch stimuli (separated by a 2 min interval) were recorded in each preparation, and these were averaged. Again, the response was defined as the difference between the number of pre-CS and post-CS fictive feeding cycles per time unit. Fictive feeding consists of bursts of N1, N2, and N3 (CPG) driven action potentials recorded in the appropriate feeding motoneurons that were illustrated in detail in a previous paper describing both unconditioned fictive feeding responses to sucrose and conditioned fictive feeding responses to touch (Staras et al., 1998b). In these electrophysiological experiments, statistically significant differences in fictive feeding responses to touch were found between the experimental and control groups. In 54% of the experimental preparations (n = 13), a positive fictive feeding response to touch was seen, 38% failed to respond, and 8% showed a small negative difference (−0.5 cycles/min) between the post-touch and pre-touch rate (Fig. 2Bi). In contrast, 84% of the control preparations (n = 12) failed to respond, and 16% of the preparations showed a decrease between their pre-CS and post-CS fictive feeding rates (Fig.2Bi). As in the behavioral tests, the experimental and control group were found to have similar proportions of preparations showing versus not showing spontaneous fictive feeding activity before the touch CS (χ2 = 0.25; df = 1;p = 0.5). The median rate of spontaneous fictive feeding before touch was 0.0 cycles/min in both groups (control interquartile range, 0.0–1.6; experimental interquartile range, 0.0–1.0). However, the nonparametric difference between experimental and control preparations, giving positive versus not positive responses, was highly significant (χ2 = 7.3; df = 1;p < 0.01). The median rate of fictive feeding response (Fig. 2Bii) in all 13 experimental preparations was 1.5 cycles/min (IQR, 0.0–2.0), significantly greater than the median control rate (0.0 cycles/min; IQR, 0.0–0.0; Mann–Whitney Utest; U = 36.0; p < 0.01). Typical examples of fictive feeding responses (or the lack of them) to lip tactile stimulation in semi-intact preparations made from trained experimental and control animals are shown in Figure 2, Biiiand Biv.
A further comparison between the behavioral and electrophysiological data revealed a strong positive correlation between CS-evoked behavioral and fictive feeding rates measured in the same animals [n = 25; Spearman correlation coefficient (rS), 0.57; significance test forrS, p < 0.03].
Appetitively conditioned animals show an increased CS-evoked synaptic input (B3 EPSP) and spike activity at the level of motoneurons
The expression of a significant positive fictive N1, N2, N3-driven feeding response to CS, monitored at the level of motoneurons, is good evidence that a conditioned response was occurring at the level of the CPG interneurons. To investigate further the cellular changes that accompany and potentially also contribute to this conditioned response it was necessary to examine changes in early synaptic events evoked by the CS. Of most interest were the earliest nonpatterned synaptic responses after lip touch, known to be present on many neurons in the feeding system, including motoneurons (Staras, 1997). The B3 cell was chosen to monitor these early responses because the synaptic inputs it receives are consistent and easy to identify. The most striking component of the B3 response to touch was a short-latency (∼75 msec) synaptic input that is a compound EPSP (Fig.3B). The B3 EPSP was the earliest component of a series of touch-evoked synaptic responses on the same cell. It was large, and its amplitude could be reliably measured.

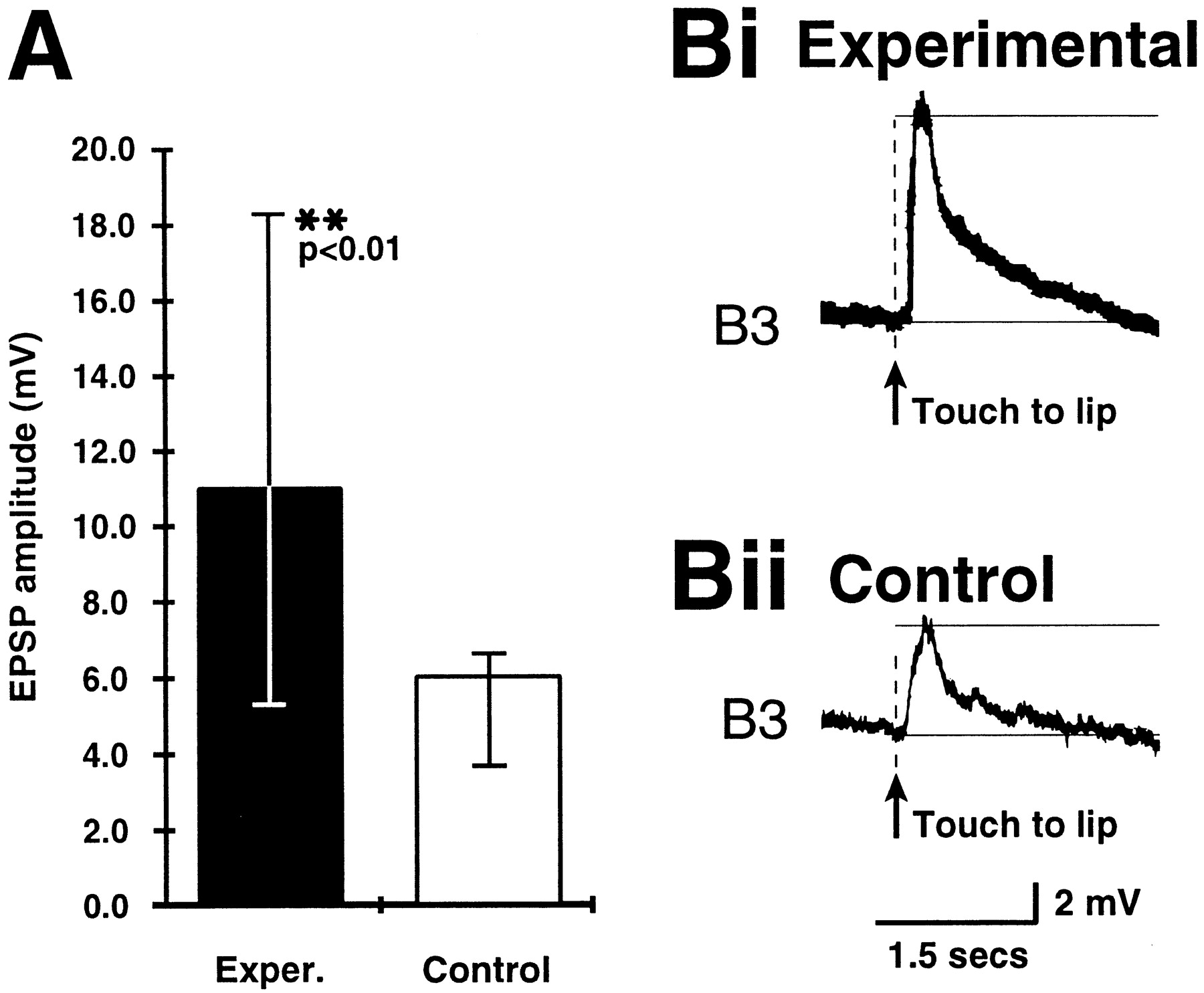
A comparison of the amplitude of the EPSP evoked by the touch CS on the B3 motoneuron in preparations from experimental and control animals. A, The EPSP amplitude is significantly larger in the experimental group than in the control group. Medians and interquartile ranges are shown. For detailed statistical data, see Results. Bi, Bii, Examples of the touch-evoked EPSP on the B3 motoneuron in an experimental preparation and in a control preparation, respectively.
The B3 EPSP amplitude was measured in the same 13 experimental and 12 control preparations in which the rate of CS-evoked fictive feeding was also measured. In each experiment two touch responses were recorded, and from these an average EPSP amplitude was calculated for each preparation. The median and IQR of the averaged amplitude data were calculated for both the experimental and the control group, and these are shown in Figure 3A. The experimental group had a median B3 EPSP amplitude of 11 mV (IQR, 5.3–18.3), which was >80% larger than the median of the control group (6 mV; IQR, 3.6–6.5). A comparison between the experimental and the control amplitude data revealed that the difference between the two groups was highly significant (Mann–Whitney U test; U = 32.5;p < 0.01). Figure 3, Bi and Bii, respectively, show examples of EPSP responses in experimental and control subjects.
We also compared the number of cases in both the experimental and control groups where the touch-evoked EPSP reached firing threshold and triggered spikes in B3. In the control group, only 4 of the 24 touch-evoked EPSPs reached threshold, whereas in the experimental group 14 of the 26 EPSPs triggered spikes in B3. The difference in the ratio of EPSPs reaching versus not reaching firing threshold between experimentals and controls was highly significant (χ2 = 7.49; df = 1; p < 0.001). This shows that in conditioned animals the B3 motoneuron is significantly more likely to fire in response to inputs arising from the CS pathway than in control animals.
The B3 EPSP amplitude is correlated with the rate of fictive feeding response to the conditioned stimulus
The B3 motoneuron is known to play no role in the generation or modulation of the feeding rhythm (Rose and Benjamin, 1979; Staras et al., 1998b), so the function of this conditioned increase in EPSP amplitude is unknown. However, it does show clearly that changes as a result of learning can occur at the level of the motoneurons. Although the increase in strength of the touch-induced response in the motoneurons themselves is not thought to be directly involved in the activation of the CPG that underlies conditioned fictive feeding, the conditioned increase in EPSP amplitude is thought to reflect similar changes occurring in other parts of the network, more directly involved with rhythm generation. For instance, the important CPG interneurons known as the N1Ms also receive a similar synaptic input (Staras, 1997). If this hypothesis is correct, then the amplitude of the B3 EPSP should be significantly positively correlated to the rate of fictive feeding response to touch. This was examined by correlation analysis of B3 EPSP amplitudes and the post-CS fictive feeding rates obtained in the same preparations. The procedure revealed that the CS-evoked EPSP amplitude was significantly positively correlated to the number of cycles of fictive feeding activity that followed it in the same B3 motoneurons (n = 25; rS, 0.43; significance test for rS,p < 0.05). The significantly larger B3 EPSP amplitudes in the experimental versus the control group is most likely to be indicative of increased activation of the same and/or additional components of the CS pathway that play a more direct role in the conditioned activation of the feeding CPG by the CS. Thus, changes in the size of the B3 EPSP represent a useful monitor of cellular changes that more directly contribute to appetitive learning in theLymnaea feeding system.
The conditioned B3 touch response can be separated from CPG-driven synaptic activity
It was important to test if, after conditioning, an increased motoneuronal EPSP could be evoked without subsequent activation of fictive feeding. This would show that plastic changes at the motoneuronal level could be recorded independently of conditioned fictive feeding. It would also rule out the possibility that a larger EPSP is simply correlated with a stronger expression of fictive feeding because of some mechanism(s) common to both in the same cell. This was achieved by first training unsatiated animals as before but then satiating the snails before dissecting them to produce the semi-intact preparations for electrophysiology. Successful pilot experiments showed that preparations made from sated animals show very low fictive feeding rates, and, therefore, satiety could be expected to have a suppressing effect on touch-evoked fictive feeding as well.
The initial behavioral procedures for conditioning animals were the same as those described previously. The experimental group (n = 9) again showed a significant positive feeding response to touch (median, 6.0 rasps/min; IQR, 5.0–7.5) compared with the control group (n = 8; median, −0.5 rasps/min; IQR, −8.5–1.0; Mann–Whitney U test; U = 5.0;p < 0.003). After an overnight period in which the animals were allowed to feed ad libitum they were dissected and prepared for electrophysiological experiments. B3 motoneurons were recorded successfully in all nine experimental and six of the eight control preparations with the emphasis on measuring the B3 EPSP latency, duration, and amplitude and the B3 resting potential.
As was the case in the preparations made from hungry snails in the previous experiment, spontaneous fictive feeding activity before the application of the CS was low in the preparations made from sated snails (Kruskall–Wallis one-way ANOVA; χ2 = 2.1; df = 3; p = 0.55). However, unlike the previous experiment with hungry snails, fictive feeding responses to touch were weak in both experimental and control preparations dissected from sated animals [medians, 0.0 cycles/min for both groups; IQRs, −1.12–0.88 (controls); 0.0–3.0 (experimentals); Mann–Whitney U test;U = 24.0; p = 0.22). In contrast, a conditioning-induced increase in the amplitude of the synaptic response in the B3 motoneuron to touch could still be recorded (Fig.4A), despite the lack of a conditioned fictive feeding response to touch. Thus, the effect of satiety was to remove the pattern of burst activity associated with CS-evoked fictive feeding in conditioned animals, without affecting the relative differences between CS-evoked motoneuronal EPSP size in experimental and control preparations (see below). The use of these more quiescent experimental preparations made it easier to make comparisons of the EPSP amplitude as well as other parameters, such as resting membrane potential, latency, and duration of the response with controls.

Detailed analysis of the touch-evoked effects on B3 after separation of the EPSP response from the fictive feeding response to touch. Insets are diagrams showing the EPSP parameter being measured. A, The EPSP amplitude is significantly larger in the experimental group than in the control group. Amplitude data are shown as medians and interquartile ranges from all preparations. B, No significant difference between experimentals and controls was found in the B3 resting potential (RP). RP data are shown as means ± SE from all preparations. C, No significant difference between experimentals and controls was found in the latency of the onset of the B3 EPSP response to touch. Data are shown as means ± SE from all responding preparations. D, No significant difference between experimentals and controls was found in the duration of the touch-evoked EPSP on B3 neurons. Data are shown as means ± SE from all responding preparations. For a more detailed statistical analysis of data, see Results.
Amplitude of touch-evoked EPSP response in B3
Median EPSP amplitudes and IQRs were calculated as before for control and experimental animals, and these are shown in Figure4A). The amplitude of the B3 EPSP was significantly larger in the experimental group (5.8 mV; IQR, 4.2–6.5) versus controls (3.8 mV; IQR, 2.3–4.3; Mann–Whitney Utest; U = 9.0; p < 0.03). A statistical comparison between the results of the present experiment on sated snails and the previous experiment on subsatiated snails showed that in both sated and hungry animals classical conditioning led to a similar relative increase in the B3 EPSP amplitude over control levels (Mann–Whitney U test; U = 52.0;p = 0.66). This is an important result, because it clearly shows that an increase in EPSP amplitude from a control to a conditioned level is not sufficient for an enhanced fictive feeding response to touch to occur. It also suggests that satiety mechanisms include effects on the CPG, and these survive into reduced preparations. Interestingly, as in the previous experiment with subsatiated snails, we found a significant difference between the satiated experimental and control group in the number of cases in which the touch-evoked EPSP reached firing threshold and triggered spikes in B3. In the satiated control group, only 2 of the 12 touch-evoked EPSPs reached threshold, whereas in the satiated experimental group 11 of the 19 EPSPs triggered spikes in B3 (χ2 = 4.55; df = 1;p < 0.02). This showed that not only satiety did not abolish the difference in EPSP size between experimental and control preparations, but it also preserved the ability of motoneurons to produce spikes in response to the increased amplitude of the synaptic input.
B3 resting potential
The resting potential (RP), measured before the CS lip touch, was not significantly different in experimental [58.6 ± 2.3 (SE) mV] versus control animals (64.2 ± 3.2 mV; unpaired ttest; df = 13; t = 1.5; p = 0.17) (Fig. 4B). Moreover, no correlation was found between the RP of B3 and the amplitude of B3 EPSP in the ranges seen in the present experiments (RP, 48.8–72.5 mV; EPSP amplitude, 6.3–24.0 mV). Changes in B3 resting potential were, therefore, not responsible for the increase of the amplitude of the touch-evoked EPSP after conditioning.
Latency of touch-evoked EPSP response in B3
The latency of touch-evoked EPSP in B3 was defined as the time between the application of touch and the onset of the depolarizing input. Unlike the resting potential, this could only be measured when the touch-evoked EPSP amplitude size was >0 (nine experimental preparations and five control preparations). No statistically significant differences were found in this parameter between the experimental and control preparations (experimental group EPSP latency, 76.7 ± 8.3 msec; control group EPSP latency, 73.0 ± 12.8 msec; unpaired t test; df = 12; t = 0.3; p = 0.8) (Fig. 4C).
Duration of touch-evoked EPSP response in B3
The duration of touch-evoked EPSP in B3 was defined as the time period that the depolarization (if any) was elevated above the pre-CS membrane potential. There was a tendency for the EPSP to be longer in duration in experimentals (1.8 ± 0.3 sec; n = 9 EPSPs) than in controls (1.3 ± 0.2 sec; n = 5 EPSPs) (Fig. 4D), but this difference was not significant when analyzed statistically (unpaired t test; df = 12; t = 1.4; p = 0.2).
A comparison of the results obtained by measuring all the above EPSP parameters suggests that increases in the firing rates rather than earlier or more prolonged firing of a component or components of the CS pathway presynaptic to B3 or a preceding neuron were occurring as a result of conditioning. This would cause an increase in the amplitude of the EPSP without necessarily changing its latency or duration. This made it necessary to try to identify conditioning-evoked changes occurring earlier than the synaptic events recorded in buccal cells.
The mechanosensory CS pathway linking the lips to the feeding network is facilitated in appetitively conditioned animals
We have no direct knowledge of the neurons that form the CS pathway that might be responsible for the conditioned enhancement of CPG-mediated fictive feeding and/or the increases in EPSP amplitude on the B3 motoneuron. However, using en passant extracellular recordings of the cerebrobuccal connective in HiDi it has been possible to record touch responses from neuronal elements that have a shorter latency than the earliest EPSP responses on the B3 neurons (see Materials and Methods). It seemed reasonable to assume that these neurons formed part of the CS pathway from lips to buccal ganglia, and so experiments were performed to see if conditioning produced increases in the levels of their response to the lip touch CS.
In the behavioral tests (performed blind on animals whose identity was not known to the observer), forming the initial part of the experiment, the conditioned feeding response after 15 trials in the intact animals was again found to be significantly higher in the experimental group (n = 9) versus the control group (n = 8) (Mann–Whitney U test; U = 9.5;p < 0.01) (Fig.5A). All experimental and control animals were subsequently dissected. Again, the electrophysiological tests were performed by a second experimenter who had no knowledge of the behavioral history of the individual animals. Preparations were set up to make extracellular recordings from the CBC that allowed the tactile CS to be presented to the lips. It was possible to electrophysiologically analyze eight experimental and seven control animals from the behaviorally tested groups. Again, like the behavioral data, there were significant differences between controls and experimentals but only in the initial part of the response to touch. Touching the lips produced a complex and prolonged burst of action potentials in both control and experimental animals (Fig.5B). The frequency of firing appeared to be greater in the first part of the response in trained animals (Fig. 5Bi) compared with controls (Fig. 5Bii). To analyze this further, each trace was divided into a series of time bins (50 msec for each one) starting at the first spike in the touch-evoked burst (0 msec). The number of extracellular spikes was counted, and an average spike number was calculated for each bin from four touch-evoked bursts in each preparation. From these data, mean responses in the experimental and the control groups across time were calculated (Fig.5C). Only in the initial 50 msec bin did the experimental animals show a significantly larger number of spikes than the control subjects in the same bin (unpaired t test; df = 13;t = 2.2; p < 0.05). The pairwise analysis of total activity across the response period between 50 and 500 msec showed no significant difference in the number of spikes between experimentals and controls. These results are significant because they show that conditioning enhances the activity of early units, firing between ∼50 and 100 msec after lip touch. These are the earliest, probably pure sensory signals that can be measured after touch and they could contribute to the other, more delayed facilitated cellular responses seen after conditioning, such as B3 EPSP and fictive feeding.

Analysis of changes in extracellularly recorded afferent activity after conditioning. A, Behaviorally recorded feeding responses to a conditioned touch stimulus (CS) from the group of experimental and control animals from which semi-intact preparations were set up to record extracellular spike activity on the CBC. The change in feeding rate after tactile presentation in the experimental group is significantly larger than the change in the control group. Data in A are shown as medians and interquartile ranges of data from all experimental and control preparations. B, Representative extracellular recordings made in semi-intact preparations in HiDi saline from i, an experimental animal, and ii, a control animal. Thespaced vertical lines represent 10 50 msec bins. For the analysis, the number of spikes occurring in each bin was counted. The general appearance of the touch-evoked responses is the same, but the number of spikes occurring in the early phase of the response is larger in the experimental preparation. C, Graph showing the mean number of spikes (± SE) recorded in each of the 10 50 msec bins for control and experimental preparations. In both groups the overall decay in spike frequency after CS is very similar. However, in the initial 50 msec bin experimental preparations showed a significantly greater number of spikes. For detailed statistical analysis of data, see Results.
DISCUSSION
The experiments described in the present paper demonstrated that behavioral conditioning of the feeding response in Lymnaealeads to plastic changes (memory formation) that can be recorded electrophysiologically at several different locations in the neuronal network underlying rhythmic feeding. This represents an intermediate level of analysis between the behavior and a more detailed study of synaptic plasticity at each site where a change was recorded. This seems essential in a complex network like that involved in feeding. The expression of the memory or its “read-out” (Byrne, 1985) was mainly recorded as feeding motoneuronal responses to the CS, but enhanced activity was also recorded in nerve fibers that were considered to be elements of an “earlier” sensory stage of the CS-activated pathway. These results are consistent with a multistage or multisite model of appetitive learning but do not rule out a single common site of plasticity, say in the sensory periphery. The causal links between the various sites of electrical change have not been directly established, although certain inferences based on the temporal sequences of events recorded after the application of CS can be made. Motoneurons are driven monosynaptically by CPG interneurons in the Lymnaeafeeding network and so act as an indirect monitor of CPG activity. In post-training tests, the CS evoked a significant activation of the feeding CPG in preparations made from experimental but not from control animals, and the rate of CS-evoked fictive feeding was correlated with the rate of feeding response measured in the same animals. This indicates that the conditioned lip touch has a facilitated long-lasting excitatory effect on the feeding CPG. Conditioning enhanced the amplitude of a specific synaptic response evoked by the CS on an identified motoneuron, B3. This increase was not attributable to alterations in the B3 membrane potential, and, therefore, was more likely to involve presynaptic or possible nonvoltage-dependent postsynaptic mechanisms. The input resistance of the B3 neurons was not measured in these experiments, so an increased response of the B3 membrane to depolarizing inputs could also be partly responsible for the enhanced EPSP amplitudes.
The enhancement of the EPSP response (and resulting increase in B3 spike activity) could be made independent of the CPG-driven conditioned fictive feeding response by satiating the snails after conditioning. This rules out the possibility that an increase in EPSP amplitude and the expression of conditioned fictive feeding are linked by a common mechanism in the same cell. However, in hungry snails, conditioned fictive feeding responses were correlated with enhanced EPSP amplitudes in B3 motoneurons. This suggests some type of parallel processing (presumably in the CNS) of the inputs leading to an EPSP and spikes in B3, and to initiation of fictive feeding, respectively. Satiation affects both the acquisition (Audesirk et al., 1982; Kemenes and Benjamin, 1994) and expression (Audesirk et al., 1982) of conditioned feeding responses in Lymnaea. However, there is no direct evidence that a period of satiety will result in a loss of the memory trace, once this has been formed. On the other hand, satiety is known to reduce the number of spontaneously occurring rasps (Tuersley and McCrohan, 1987; Kemenes and Benjamin, 1994) as well as the intensity of the unconditioned feeding response itself (Kemenes and Benjamin, 1994). Therefore, it is more likely that the loss of fictive feeding response to touch in sated conditioned animals is caused by the strong effect of satiety on some aspect of the feeding motor program rather than to a direct effect of satiety on the memory trace. Moreover, the retention of the enhanced conditioned EPSP response in preparations made from satiated conditioned animals provided direct evidence that at least some aspect of the memory trace remained intact after satiation.
The conditioned feeding response to touch far outlasts the duration of the B3 EPSP, this again suggests that a separate pathway is involved in maintaining the former. However, parallel processing of the signals reaching B3 and the CPG does not rule out a common fundamental origin for all the changes recorded, upstream to both CPG interneurons and feeding motoneurons.
Some evidence for this was obtained by recording short-latency touch-induced activity in a central connective. Like CPG and B3 responses, these early responses to touch could be enhanced after behavioral conditioning. A variety of data suggested that the extracellularly recorded units are early in the sensory CS pathway and could be part of the pathway responsible for the conditioned fictive feeding response in the buccal feeding network. The conditioning-induced changes, seen at the cellular level, are summarized in Figure 6. In control preparations, touch to the lips gives rise to a brief burst of spikes on the CBCs, the pathway linking peripheral mechanosensory neurons to putative sensory integrating neurons in the buccal ganglia (Staras, 1997). It also evokes an EPSP in motoneuron B3, but triggers no fictive feeding (Fig. 6A). In contrast, in preparations from conditioned animals, touch to the lips evokes a facilitated spike response on the CBCs, a facilitated EPSP on B3, and CPG-driven fictive feeding (also recorded on B3) (Fig. 6B).
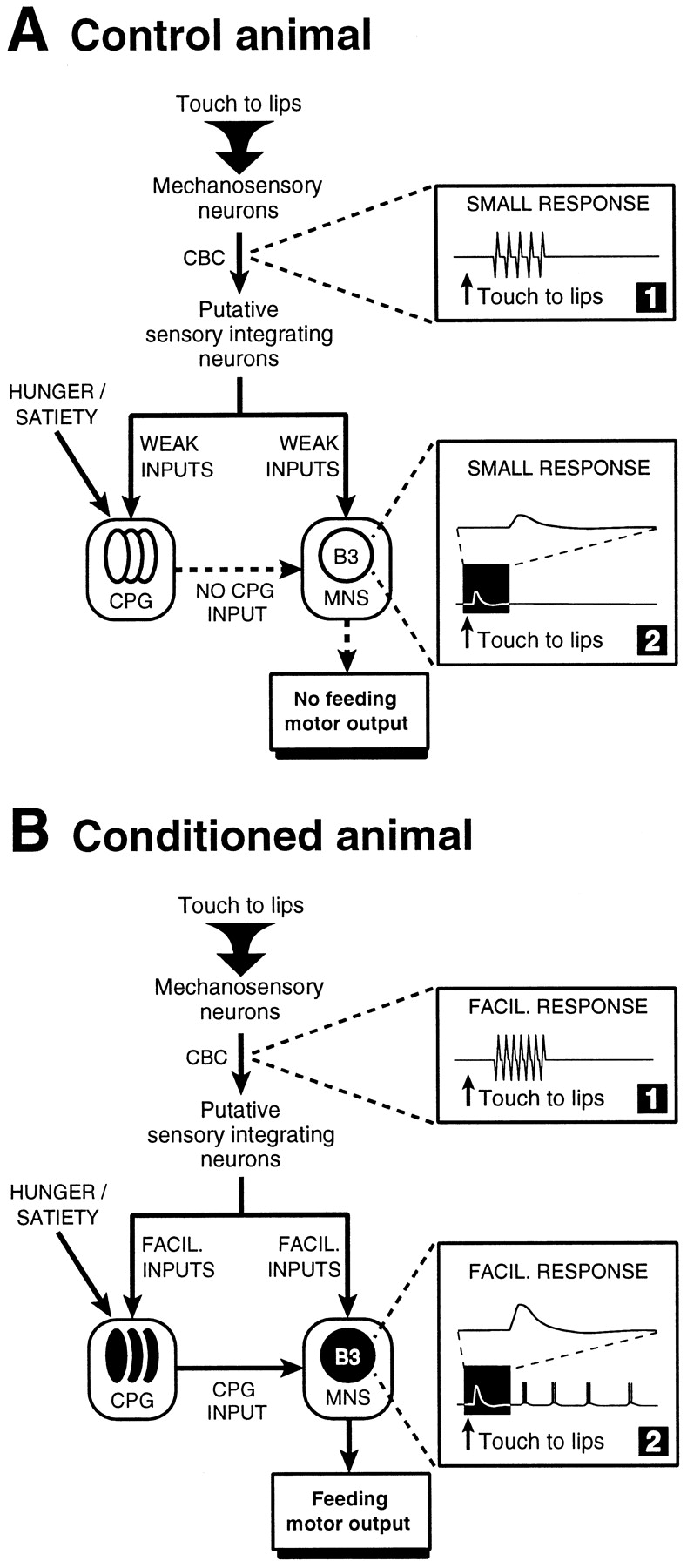
Summary of electrophysiological changes recorded after appetitive conditioning in Lymnaea. A, In control animals a lip touch activates early components of the mechanosensory pathway, which can be recorded as extracellular spikes on the CBCs (box 1). Putative sensory integrating neurons in the buccal ganglia (Staras, 1997) are thought to distribute weak synaptic inputs to many feeding neurons, including members of the interneuronal CPG network and identified motoneurons, such as B3. The synaptic response seen on the B3 after a lip touch consists of an EPSP (box 2). This, and the possible synaptic response on the CPG network are subthreshold. The lack of activation of the CPG by the touch input means that the pathways linking the CPG to the motoneuronal followers (indicated by dashed arrows) do not become activated, and, consequently, no rhythmic fictive feeding occurs. The CPG and the B3 in inactive state are shown as hollow circles. B, In conditioned animals, the lip touch evokes facilitated extracellularly recorded spike activity on the CBC (box 1). This may lead to an increased activation of putative sensory integrating neurons, which in turn, may contribute to the facilitated synaptic drive to both CPG and motoneurons, such as B3 (box 2, expanded time-base trace). Rhythmic activity in the CPG, triggered by the CS, provides synaptic drive to the motoneuronal network, and this leads to the patterned activity we observe after the touch-evoked EPSP on the B3 motoneuron in conditioned animals (box 2, bottom trace). The CPG and the B3 in activated state are shown as filled circles. Satiety will selectively suppress the expression of CPG response to touch without affecting the facilitated EPSP response in B3.
The CS-evoked sensory response was recorded on the cerebrobuccal pathway after a shorter latency than the other, more central, responses in the buccal ganglia. This suggests that the former might be responsible for the latter. However, it was not possible to prove a link between the facilitation of early activity and increases in buccal ganglion responses because isolation of the early components of the sensory responses using HiDi saline blocked most of the more central responses. Despite this, the facilitation of early afferent events in the CS pathways after conditioning may be an important common factor underlying all the other changes observed. It is possible, as was found for aversive conditioning in Aplysia (for review, seeHawkins, 1984; Carew and Sahley, 1986; Byrne, 1987; Glanzman, 1995), that critical changes occur in primary mechanosensory neurons during learning. However, postsynaptic changes, affecting secondary components of the CS pathway, such as putative sensory integrator neurons (Fig. 6) (Staras, 1997) or the responsiveness of CPG interneurons to touch-induced inputs, are also likely to play a role in the build-up of the conditioned feeding response in Lymnaea. Indeed, recent work (Kojima et al., 1997) showed that during taste aversion learning in Lymnaea, important changes take place in the responsiveness of the protraction phase CPG interneuron, N1M, to inputs arising from the serotonergic CGCs (Yeoman et al., 1996), modulatory cells that are activated by food (Kemenes et al., 1986). This indicates that the N1M interneuron itself may be an important site of plasticity. A similar suggestion was made in our previous work describing in vitro appetitive classical conditioning of the feeding response inLymnaea (Kemenes et al., 1997). This will make it worthwhile in future work to compare the relative contribution of changes upstream of the feeding CPG and changes taking place in the CPG itself to the shaping of the conditioned feeding response. More recent work inAplysia also suggests that in addition to presynaptic facilitation of the activity of primary mechanosensory cells, postsynaptic LTP-type plasticity in motoneurons also plays a role in classical conditioning of withdrawal responses (Glanzman, 1995; Bao et al., 1998).
Comparisons with previous experiments on appetitive learning in molluscs
Most previous experiments on appetitive learning in molluscs have been largely concerned with the behavioral expression of conditioning (Audesirk et al., 1982; Kemenes and Benjamin, 1989a,b, 1994; Sahley et al., 1990; Kojima et al., 1996; Colwill et al., 1997) and have offered little insight into the cellular mechanisms underlying this process. Although in a few cases neuronal correlates of conditioned feeding have been described in preparations dissected from trained animals, these never went beyond demonstrating the survival of a correlate of thein vivo rhythmic conditioned behavior. InPleurobranchaea, it was demonstrated that animals that were appetitively conditioned with touch CS and food US showed facilitated feeding CPG activity on a buccal nerve after electrical stimulation of the nerve carrying the CS pathway (Mpitsos and Davis, 1973). Recently, fictive feeding responses to both chemical and tactile conditioned stimuli have been reported in appetitively conditionedLymnaea (Whelan and McCrohan, 1996; Staras et al., 1998b). However, our present work provides the first detailed evidence in a CPG-driven system for changes recorded at specific locations, e.g., the level of the sensory CS pathway and synaptic inputs reaching identified cells through the CS pathway. Taken together with the results presented in two previous papers on the cellular analysis of tactile learning inLymnaea (Kemenes et al., 1997; Staras et al., 1998b), these findings suggest that Lymnaea is a suitable model for further analysis of the cellular basis of appetitive learning, particularly given that plasticity in the feeding network can readily occur with both chemical (Whelan and McCrohan, 1996) and tactile CS. This is important, because so far there has been only one type of invertebrate model in which cellular aspects of appetitive learning could be analyzed in detail, the conditioned proboscis extension reflex in the honey bee Apis mellifera (Hammer, 1993, 1997).
Footnotes
This work was supported by Biotechnology and Biological Sciences Research Council Grant GR/J33234 (United Kingdom).
Correspondence should be addressed to Dr. Kevin Staras (c/o Dr. G. Kemenes), Sussex Centre for Neuroscience, School of Biological Sciences, University of Sussex, Falmer, Brighton, United Kingdom, BN1 9QG.
Dr. Staras’s present address: Department of Physiology, Royal Free Hospital, School of Medicine, Rowland Hill Street, London, United Kingdom, NW3 2PF.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}