

Abstract
At the Drosophila glutamatergic neuromuscular junction, the postsynaptic cell can regulate synaptic strength by both changing its sensitivity to neurotransmitter and generating a retrograde signal that regulates presynaptic transmitter release. To investigate the molecular mechanisms underlying these forms of plasticity, we have undertaken a genetic analysis of two postsynaptic glutamate receptors that are expressed at this synapse. Deletion of both genes results in embryonic lethality that can be rescued by transgenic expression of either receptor. Although these receptors are redundant for viability, they have important differences. By transgenically rescuing the double mutant, we have investigated the relationship of receptor gene dosage and composition to synaptic function. We find that the receptor subunit composition regulates quantal size, Argiotoxin sensitivity, and receptor desensitization kinetics. Finally, we show that the activity of the receptor can regulate the retrograde signal functioning at this synapse. Thus, the diversity of receptors expressed at this synapse provides the cell with mechanisms for generating synaptic plasticity.
- glutamate receptor
- Drosophila
- neuromuscular junction
- retrograde signal
- genetics
- synaptic plasticity
- quantal size
- quantal content
Synaptic strengths change as neuronal circuits develop and are modified by experience. The postsynaptic cell can contribute to this plasticity by changes in its sensitivity to transmitter and, at some synapses, by the generation of a retrograde signal that regulates presynaptic transmitter release. Both postsynaptic mechanisms function at theDrosophila neuromuscular junction (NMJ).
One mechanism for modulating the postsynaptic response to transmitter is by regulating the subunit composition of neurotransmitter receptors. If receptor subunits have different physiological properties, then the postsynaptic response could be regulated by the differential expression, targeting, membrane insertion, degradation, or posttranslational modification of these subunits. Such a regulatory mechanism functions at the developing vertebrate NMJ; the switch from the fetal acetylcholine receptor γ subunit to the adult ε subunit changes the open time and conductance of the receptor (for review, see Mishina et al., 1986; Schuetze and Role, 1987; Gu and Hall, 1988). A similar mechanism may regulate GABAA (Brooks-Kayal and Pritchett, 1993; Tia et al., 1996), glycine (Takahashi et al., 1992), and NMDA receptors (Sheng et al., 1994). Vertebrate AMPA-type glutamate receptor subunits differ in physiological parameters, such as ion permeability, desensitization kinetics, and toxin binding (for review, see Hollmann and Heinemann, 1994; Westbrook, 1994). It has been shown that the differential expression of these subunits gives rise to postsynaptic receptors with different properties (Jonas et al., 1994;Geiger et al., 1995; Washburn et al., 1997).
The Drosophila NMJ, like most central vertebrate excitatory synapses, is glutamatergic, expresses homologous ionotropic receptors, is organized into boutons, and exhibits dynamic functional plasticity. Two muscle-specific glutamate receptors, DGluRIIA andDGluRIIB, function at this synapse (Schuster et al., 1991;Petersen et al., 1997). In previous work, we showed that deletion ofDGluRIIA leads to a decrease in quantal size and a compensatory upregulation of quantal content, indicating the presence of a muscle-to-motoneuron signal-regulating presynaptic transmitter release (Petersen et al., 1997; Davis et al., 1998; Landmesser, 1998). A similar homeostatic compensation has been observed at the NMJ of both crayfish (Lnenicka and Mellon, 1983) and mammals (Cull-Candy et al., 1980; Plomp et al., 1992; Sandrock et al., 1997) and may also function at vertebrate excitatory (Turrigiano et al., 1998) and inhibitory (Nusser et al., 1998) central synapses.
Here, we present a genetic analysis of these Drosophilaglutamate receptors and investigate their role in the postsynaptic regulation of synaptic strength at the NMJ. We demonstrate that deletion of both receptors leads to embryonic lethality and that either receptor is sufficient for viability. We exploit the ability to rescue the double mutant via the transgenic expression of either receptor to test the effect of receptor gene dosage and composition on synaptic function. We show that receptor subunit composition (i.e., the receptor subtype expressed by muscle) regulates quantal size, Argiotoxin sensitivity, and receptor desensitization kinetics. We next examine the relationship of receptor expression to the retrograde regulation of quantal content. Finally, by overexpressing a dominant negative receptor with a mutation in the putative ion conduction pore, we show that the muscle-to-motoneuron signal is regulated by the activity, and not density, of the channel.
MATERIALS AND METHODS
Rescue constructs and dominant negatives. DGluRIIA rescue constructs were made as previously described (Petersen et al., 1997). The genomic DGluRIIB rescue construct consists of a genomic fragment extending from theHindIII site in the 3′ end of DGluRIIA toSalI site ∼500 bp downstream of DGluRIIB, subcloned into pUAST (Brand and Perrimon, 1993). TheDGluRIIB cDNA was cloned into a transformation vector containing the myosin heavy chain (MHC) promoter (Wassenberg et al., 1987), as well as into pUAST.
To create a dominant negative receptor, M614 in DGluRIIA was changed to R, using a PCR-based strategy (Hollmann et al., 1994). Briefly, two complementary primers were created containing the desired mutation. Each of these primers was used in a PCR reaction in combination with a primer at the 3′ or 5′ end of the cDNA as appropriate. The two resulting PCR products were annealed and amplified in a secondary PCR reaction. An Sfi/Asc fragment from this mutated cDNA was subcloned into a transformation vector containing the myc-tagged DGluRIIAcDNA in pUAST (Petersen et al., 1997). The XL-PCR kit (Perkin-Elmer, Emeryville, CA) was used for all PCR reactions.
Mutations in DGluRIIA and DGluRIIB. Mutations inDGluRIIA were made using a P element-hopping strategy as described previously (Petersen et al., 1997). Two deletions that removed both DGluRIIA and DGluRIIB,DGluRIIA&BSP22 andDGluRIIA&BAD1, were recovered using the same strategy. DGluRIIA&BSP22 results from the imprecise excision ofP[w+60] (located immediately upstream of DGluRIIA). It extends ∼8 kb upstream ofDGluRIIA and downstream beyond the coding region ofDGluRIIB. DGluRIIA&BAD1derives from an imprecise excision ofP[w+228] (immediately upstream of DGluRIIA) andP[w+72] (located between the two genes). It extends ∼200 bp upstream of DGluRIIA and 1 kb into the coding region of DGluRIIB (Petersen et al., 1997) (Fig. 1A).

Genetic analysis of DGluRIIA andDGluRIIB. A, Excisions ofDGluRIIA and DGluRIIB were generated by P-element mutagenesis. Excisions deleting only DGluRIIA(SP16 and AD9) were described previously (Petersen et al., 1997). Excisions that disrupt both receptors (SP22 and AD1) were generated by the imprecise excision of nearby P elements. B, The lethality associated with the receptor double mutant is rescued by the transgenic expression of the genomic region encompassing eitherDGluRIIA or DGluRIIB or by the expression of either cDNA driven by the muscle-specific MHC promoter.
Genetic strains. Low levels of transgenicDGluRIIA or DGluRIIB were introduced with the insertion of a genomic fragment spanning the complete coding sequence (see above). Higher levels and targeted expression were accomplished using the Gal4-upstream activation sequence (UAS) system (Brand and Perrimon, 1993). In Drosophila, there is a close relationship between gene copy number and expression level. Independent insertions of each receptor transgene were tested and found to give similar results. The Gal4 driver lines used to cross toUAS-DGluRIIA or UAS-DGluRIIB were 24B-Gal4 (Brand and Perrimon, 1993), an enhancer trap line, which expresses strongly in embryonic and larval somatic muscle MHC Gal4 (myosin heavy chain promoter fused to Gal4) (M. Winberg, personal communication) and H94-Gal4, an enhancer trap line variably expressing in muscle (D. Lin, unpublished observations).
Physiology. Intracellular recordings were done on muscle 6, segment A3 of third instar larvae. Physiology and analysis were done as described previously (Petersen et al., 1997), except that determination of quantal size for the dominant negative experiments was done using the Mini Analysis program (Jaejin Software, Leonia, NJ) instead of pClamp6 software. The bath saline contains (in mm): 70 NaCl, 5 KCl, 20 MgCl2, 10 HCO3, 5 trehalose, 115 sucrose, and 5 HEPES, the concentration of calcium described in the text, pH adjusted to 7.2 (HL3) (Stewart et al., 1994). Spontaneous synaptic events with very slow kinetics are caused by electrical coupling with neighboring muscle (Ueda and Kidokoro, 1996) and were excluded from analysis. Argiotoxin 636 (Accurate Chemicals, Westbury, NY) was prepared in physiological saline and was bath applied.
For patch-clamp recording, dissected third instar larvae were bathed in Schneiders Drosophila medium (Life Technologies GmbH, Eggenstein, Germany) containing 30 μg/ml collagenase type 1A (Sigma-Aldrich Chemie GmbH, Deisenhofen, Germany) for 15 min before patching and then superfused with physiological saline HL3 (Stewart et al., 1994). Patch pipettes had resistances of ∼5 MΩ when filled with intracellular solution containing 150 mm K-propionate, 5 mm Na-propionate, 10 mmMgCl2, 1 mm CaCl2, 10 mm EGTA, and 10 mm Tris-maleate buffer, pH adjusted to 7.4 with 5 N NaOH. Outside-out patches were taken from extrajunctional regions of muscle 6 and 7 of the abdominal segments of the larvae. The patches were voltage clamped to −60 mV and moved to an application chamber that was perfused with a solution containing 135 mm NaCl, 5 mm KCl, 4 mmMgCl2, 2 mm CaCl2, and 5 mm HEPES, pH adjusted to 7.4 with 5 N NaOH. Glutamate was applied with a liquid filament switch every 1–2 sec, and the currents were recorded and evaluated as described previously (Heckmann and Dudel, 1997). Channel activity is seen in ∼50% of patches.
RESULTS
DGluRIIA and DGluRIIB are functionally redundant For viability
Two ionotropic glutamate receptors, DGluRIIA andDGluRIIB, are expressed at the Drosophilaneuromuscular junction. The genes for these receptors are more similar to each other than to any other glutamate receptor, share a similar intron–exon structure, are adjacent in the genome, and are coexpressed in all somatic muscles. However, they only share 44% amino acid identity and have amino acid differences in functionally important parts of the molecule, such as the putative ion conduction pore (Petersen et al., 1997). To assess the functional role of these two receptors in vivo, we wished to analyze synaptic function at neuromuscular junctions expressing either one or the other receptor. To do this, we generated double mutants in which both genes are deleted and then rescued these mutants with either gene alone.
We have generated previously mutants lacking DGluRIIA via excision of nearby P elements. Using these same P elements, we have now generated excisions that delete both DGluRIIA andDGluRIIB (Fig. 1).P[w+228], which is located 300 bp upstream of DGluRIIA, was imprecisely excised to generate a mutant, DGluRIIA&BSP22, which deletes the entire coding regions for both genes. A second mutant,DGluRIIA&BAD1, was derived from the simultaneous and imprecise excision of two P elements,P[w+228] andP[w+72], which are located between the two genes. This mutant removes the entire coding region ofDGluRIIA and approximately the first 1 kb ofDGluRIIB.
Both of these double mutants are embryonic lethal. The homozygous mutants or the mutants in combination with a large deficiency of the region (Df(2L)clh4) develop to be late embryos but are unable to hatch. When mechanically removed from the chorion and viteline membranes, the mutant embryos appear to be anatomically grossly normal, but they are unable to crawl. The head is capable of some coordinated movements, but the abdominal body wall muscles merely fibrillate and there are no coordinated peristaltic waves. Therefore, these two receptors are essential for synaptic transmission at the neuromuscular junctions of the abdominal musculature.
To demonstrate that this phenotype is caused by disruption of the glutamate receptor genes, we generated transgenic flies carrying genomic rescue fragments of either DGluRIIA orDGluRIIB (Fig. 1B). The transgenic expression of either gene is able to rescue the lethality associated with both DGluRIIA&BSP22 andDGluRIIA&BAD1. The rescued flies appear behaviorally normal. This demonstrates that either gene is sufficient and that neither is necessary for viability. The transgenic expression of either cDNA driven by the muscle-specific myosin heavy chain promoter is also able to rescue lethality. Therefore, the essential function of these genes is in the somatic musculature.
Receptor subunit composition regulates quantal size
Although the two receptors are redundant at the level of viability, the many differences in amino acid sequence suggested that they might have physiological differences. One measure of receptor function is the quantal size, or response of the muscle to the spontaneous release of a single synaptic vesicle. Quantal size reflects the postsynaptic sensitivity to transmitter, which is determined in large part by the properties of the transmitter receptor. With the genetic tools at hand, we were able to vary both the receptor subunit composition and gene dosage and assess the effect on quantal sizein vivo.
Before comparing the properties of the two genes, we wished to verify that the transgenic genomic rescue fragments functioned in the same manner as the endogenous genes. We have demonstrated previously that expression of the DGluRIIA genomic rescue fragment quantitatively mimics the endogenous DGluRIIA gene in the presence of DGluRIIB (Petersen et al., 1997, their Fig. 4B). To assess the DGluRIIB genomic rescue fragment, we compared the quantal size of the double mutant rescued with the transgenic genomic DGluRIIB to aDGluRIIA mutant expressing the endogenousDGluRIIB and found no difference in quantal size (transgenicDGluRIIB, 0.25 ± 0.01 mV; n = 11; endogenous DGluRIIB, 0.24 ± 0.01 mV; n= 10). Therefore, these rescue transgenes can be used to assess differences between the two receptors.
Comparison of quantal size at synapses expressing one or the other receptor revealed that DGluRIIA-expressing synapses exhibit a significantly larger response to transmitter thanDGluRIIB-expressing synapses (Fig.2). To ensure that this was not an artifact of level of expression, we expressed either one genomic copy or two genomic copies or we grossly overexpressed the receptor cDNAs using the Gal4-UAS expression system (Brand and Perrimon, 1993). At all gene dosages, DGluRIIA-expressing synapses had a significantly larger quantal size. In fact, the lowest level ofDGluRIIA expression still gave a threefold larger quantal size than did the highest level of DGluRIIB expression. In addition to the difference in amplitude, there was also a difference in the kinetics of the synaptic potentials. The time constant of the miniature extrajunctional potential (mEJP) decay is significantly shorter in DGluRIIB expressing larvae than inDGluRIIA-expressing larvae (21.6 ± 0.6 msec;n = 9; and 32.9 ± 1.2 msec; n = 12, respectively; p < 0.001).

Receptor subunit composition regulates quantal size. A, Representative traces of spontaneous transmitter release recorded from muscle 6, segment A3 ofDGluRIIA&BSP22 double-mutant larvae rescued by DGluRIIA or DGluRIIB expressed by a genomic transgene (1×) or a UAScDNA driven by 24BGal4 (>2×). Calibration: 200 msec, 2 mV. B, Quantal size is significantly larger at synapses rescued with DGluRIIA. The mean ± SEM for the mEJP amplitude is shown for rescue of the glutamate receptor mutant by various doses of DGluRIIAand DGluRIIB: 1×, a single copy of a genomic rescue transgene (DGluRIIA, n = 12;DGluRIIB, n = 11;p < 0.001); 2×, two genomic copies (DGluRIIA, n = 12;DGluRIIB, n = 11;p < 0.001); and >2×, UAScDNA driven by 24BGal4 (DGluRIIA, n = 11;DGluRIIB, n = 14;p < 0.001). The mean ± SEM of the resting potentials: 1×, DGluRIIA, 70.8 ± 1.5 mV;DGluRIIB, 69.7 ± 1.6 mV; 2×,DGluRIIA, 69.7 ± 1.2 mV; DGluRIIB, 70.9 ± 1.2 mV; >2×, DGluRIIA, 71.3 ± 1.0 mV; DGluRIIB, 70.4 ± 0.7 mV.
The data above suggest that the ratio of receptor subunits at the wild-type synapse could regulate quantal size. A larger proportion of DGluRIIA would increase quantal size, whereas more DGluRIIB would decrease quantal size. We have demonstrated previously that when DGluRIIA is overexpressed in a wild-type background, there is a significant increase in quantal size (Petersen et al., their Fig. 7). However, this result is equally consistent with quantal size being regulated by receptor subunit composition or receptor density. To distinguish between these two possibilities, we have overexpressed DGluRIIB in a wild-type background. We used a late driver, MHC Gal4, which initiates expression in the first larval instar after endogenous receptor expression has begun, and an early driver, 24B Gal4, which initiates expression in myoblasts. In both cases, there is a significant decrease in quantal size (Fig. 3). Late expression of DGluRIIB leads to a 44% reduction in mEJP amplitude, whereas earlier expression produces a 68% decrease. A similar change is mEJP amplitude is seen when DGluRIIB is directly overexpressed from the myosin heavy chain promoter (wild type, 1.08 ± 0.05 mV; n = 9; MHC-DGluRIIB, 0.56 ± 0.06 mV; n = 9; p < 0.001). Despite the likely increase in receptor density caused by overexpression, quantal size fell because of a change in the relative abundance of receptor subtype.

Overexpression of DGluRIIBdecreases quantal size at a wild-type synapse. A, Representative traces of spontaneous transmitter release recorded from muscle 6, segment A3 of third instar larvae expressing only endogenous glutamate receptors (WT, wild type), overexpressingDGluRIIB at an otherwise wild-type synapse (WT + P[B], UASDGluRIIB × MHCGal4), or strongly overexpressingDGluRIIB at an otherwise wild-type synapse (WT ++ P[B], UASDGluRIIB × 24BGal4). Calibration: 200 msec, 2 mV. B, The mean ± SEM of the mEJP amplitude is shown for WT (n = 13), WT + P[B] (n = 10), and WT ++ P[B] (n = 15). Overexpression of DGluRIIB in a wild-type background leads to a significant decrease in quantal size [WT + P[B] (p < 0.001) and WT ++ P[B] (p < 0.001)]. All three lines had similar resting potentials (WT, −72.4 ± 1.4 mV; WT + P[B], −68.5 ± 1.7 mV; WT ++ P[B], −69.5 ± 1.3 mV).
Although receptor subunit composition is a primary determinant of quantal size, the data suggest that receptor density may also regulate postsynaptic sensitivity to single quantum. When the double mutant is rescued with increasing gene dosages of DGluRIIA, there is a significant increase in quantal size (Fig. 2). There is an 18% increase from one to two genomic copies of A (p< 0.05) and a further 20% increase from two genomic copies to gross overexpression of the cDNA (p < 0.05). Because no DGluRIIB is expressed in any of these genotypes, these results cannot be explained by a change in subunit composition between these two receptors, although we cannot rule out the existence of a third receptor that may function at this synapse. Similarly, there is a 24% increase in quantal size when the gene dosage ofDGluRIIB is doubled (p < 0.01) while rescuing the null mutant. However, there is no further increase in mEJP amplitude when the DGluRIIB cDNA is overexpressed. These data are consistent with a model in which receptor density is a determinant of quantal size.
Receptor subunit composition regulates sensitivity to Argiotoxin 636
The orb web spider toxin Argiotoxin 636 is a specific open-channel blocker of invertebrate glutamate receptors (Jackson and Usherwood, 1988). Previous studies have demonstrated that it is capable of blocking synaptic transmission at the Drosophilaneuromuscular junction (Broadie and Bate, 1993; Jarecki and Keshishian, 1995; Zhong and Peña, 1995). By genetically manipulating receptor composition, we have begun a molecular characterization of the site of action of the toxin.
Baseline synaptic transmission was assessed, 0.5 μm toxin was perfused in the saline, >1000 stimuli were given to allow the use-dependent blocker access to the receptors, and then residual synaptic transmission was measured (Fig.4A). This concentration of Argiotoxin 636 reduced the EJP size by 80% at wild-type synapses. The inhibition was significantly greater at synapses that only expressDGluRIIA (94%; p < 0.05) and significantly less at synapses expressing only DGluRIIB (40%;p < 0.01) (Fig. 4B).DGluRIIB-expressing synapses are sensitive to the toxin, because 1 μm Argiotoxin 636 blocked synaptic transmission by 75%. Therefore, this toxin blocks channels containing either subunit but is a much more potent blocker in the absence ofDGluRIIB.

Differential block by Argiotoxin 636.A, Baseline evoked synaptic transmission in third instar larvae was assessed, 0.5 μm Argiotoxin 636 was added to the bath, 1000 stimuli were given to allow the blocker access to the receptor pore, and then residual synaptic transmission was assessed. EJP amplitude was normalized to the mean baseline response for each cell. To reduce quantal variation, each point represents the average of three consecutive EJPs. A representative experiment is shown for a wild-type larvae (open circles), aDGluRIIA&BSP22 double mutant rescued by the expression of DGluRIIA (filled circles), and aDGluRIIA&BSP22 double mutant rescued by the expression of DGluRIIB (filled triangles). B, The mean ± SEM of the normalized EJP amplitude after block with 0.5 μmArgiotoxin 636 for wild-type larvae (n = 5),DGluRIIA&BSP22 double-mutant larvae rescued by the expression of DGluRIIA(n = 5), andDGluRIIA&BSP22 double-mutant larvae rescued by the expression of DGluRIIB(n = 5).
Receptor subunit composition regulates desensitization
To investigate the underlying biophysical basis for the observed differences in quantal properties, we have begun a single-channel analysis of the two receptor subunits. Outside-out patches were isolated from extrajunctional regions of muscle 6 of wild-type third instar, as well as DGluRIIA&BSP22 mutant larvae rescued with either DGluRIIA or DGluRIIB. The patches were held at −60 mV, and 10 mm glutamate was applied with a rapid application system. In response to glutamate, the channels opened rapidly, flickered between open and closed states, and desensitized in the continued presence of glutamate. In Figure5A, the top two traces show single responses, and the third trace shows an average current response to glutamate application. There is no significant difference in single-channel current amplitudes in the three genotypes (Fig. 5B). Their single-channel conductance is very similar to what has been observed previously for wild-type channels from larvae (Heckmann and Dudel, 1995) and embryos (Broadie and Bate, 1993; Nishikawa and Kidokoro, 1995). There is, however, a marked difference in the time course of desensitization. When fit with an exponential function, the time constant of decay is 18 msec for channels from wild-type larvae, 19 msec from larvae expressing DGluRIIA, and 2.0 msec for channels from larvae expressing DGluRIIB (Fig.5C). Because we have not observed channels from wild-type patches that desensitize as quickly as the DGluRIIB channels (present study; Heckmann and Dudel, 1997), DGluRIIBhomomultimers must be quite rare in a wild-type cell. The channels analyzed here are extrajunctional; however, we have found no evidence for a difference in the time course of patch and quantal currents (Heckmann and Dudel, 1998). Therefore, this difference in the time course of desensitization seen with single channels may explain some of the differences in quantal amplitude and time course seen at synapses in larvae rescued with either DGluRIIA orDGluRIIB.

Receptor subunit composition regulates desensitization kinetics. A, Outside-out patches were isolated from the muscle membrane of wild-type andDGluRIIA&BSP22 double-mutant larvae rescued by the expression of DGluRIIA andDGluRIIB. The top two traces show single responses, and the third trace shows an average response to the rapid application of 10 mm glutamate (solid black line). The average responses are shown normalized with respect to the peak current amplitude and are not to scale. The single-channel current amplitude (B) is not significantly different in the three genotypes (wild type, 8.8 ± 0.4 pA; n = 6; DGluRIIA, 9.2 ± 0.3 pA; n = 6; DGluRIIB, 8.1 ± 0.5 pA; n = 5), but the time constant of desensitization (C) is much more rapid inDGluRIIB-expressing larvae (wild type, 17.5 ± 1.0 msec; n = 6; DGluRIIA, 18.8 ± 3.1 msec; n = 6; DGluRIIB, 2.0 ± 0.7 msec; n = 4; p < 0.005). The mean ± SEM is shown for B andC. Calibration: 10 msec, 5 pA.
An inverse relationship between quantal size and quantal content inDGluRIIB mutants
We have demonstrated previously that, in DGluRIIAmutants, the amplitude of evoked synaptic events remain normal despite a large decrease in quantal size because of a compensatory increase in quantal content, the number of vesicles released by the nerve (Petersen et al., 1997). These data were taken as evidence for a retrograde signal linking postsynaptic activity with presynaptic transmitter release properties. We wished to assess whether a similar form of retrograde signaling is active at synapses mutant forDGluRIIB.
At synapses lacking DGluRIIB, the quantal size is near wild-type levels. To assess the relationship between quantal size and quantal content over a wide range of values, we rescued the double mutant with a transgenic UAS DGluRIIA cDNA driven by a Gal4 line (H94) that gives quite variable levels of expression. Recordings of spontaneous miniature junctional potentials and evoked excitatory junctional potentials were made from muscle 6, segment A3 of third instar larvae (Fig.6A), and quantal content was estimated by dividing the mean EJP amplitude by the mean mEJP amplitude. There was a significant difference in quantal content when cells were grouped by quantal size; cells with the smallest quantal size tend to have the largest quantal content (Fig.6B). This suggests that, at synapses lackingDGluRIIB, changes in postsynaptic activity are compensated for by regulating presynaptic transmitter release. In this genotype, the amplitude of the evoked events is significantly larger than in wild type (25.1 ± 1.4 mV; n = 16; and 15.4 ± 2.0 mV; n = 10, respectively; p < 0.001) (see below).

An inverse relationship between quantal size and quantal content. A, Representative traces of spontaneous and trace averages of evoked transmitter release recorded in 0.42 mm calcium from muscle 6, segment A3 ofDGluRIIA&BSP22 double-mutant larvae rescued by the expression of UASDGluRIIA driven by H94Gal4. H94Gal4 gives variable levels of expression from segment to segment, and Cell 1 and Cell 2 are examples of cells with very different sensitivity to transmitter. Note the similarity in evoked release despite the large difference in quantal size. Calibration: spontaneous, 200 msec, 2 mV; evoked, 10 msec, 5 mV. B, The mean ± SEM of the quantal content is shown for cells grouped by quantal size (<0.9 mV,n = 4; 0.9–1.2 mV, n = 8; >1.2 mV, n = 4). The calculated quantal content for the 0.9–1.2 mV bin is significantly smaller than for the <0.9 mV bin (p < 0.05) and significantly larger than for the >1.2 mV bin (p < 0.01). The linear regression for quantal size versus quantal content has a coefficient of r2 = 0.66. The mean quantal content was calculated for each cell by dividing the average suprathreshold EJP amplitude (n > 75) by the average amplitude of the spontaneous miniature events (n > 60). After correcting for nonlinear summation (assumed reversal potential of 10 mV; Martin, 1955), the same trend is apparent (quantal content: <0.9 mV, 51 ± 6; 0.9–1.2 mV, 41 ± 4; >1.2 mV, 20 ± 8).
Quantal content overcompensates at synapses expressing low levels of DGluRIIA
To assess in a more quantitative manner the relationship between gene dosage of DGluRIIA and quantal content, we compared the synaptic response in 0.3 mm external calcium at the wild-type synapse and in the double mutant rescued with one genomicDGluRIIA transgene, two genomic DGluRIIAtransgenes, or by overexpression of the DGluRIIA cDNA (Fig.7A). As would be expected from the results above, the single genomic DGluRIIA, with the smallest mean quantal size, gave the largest quantal content. The single DGluRIIA showed a significant increase in quantal content compared with wild type (237%; p < 0.01), two copies of genomic DGluRIIA had a smaller increase (180%;p < 0.05), and overexpression of DGluRIIAhad no change in quantal content (114%; p = 0.58).
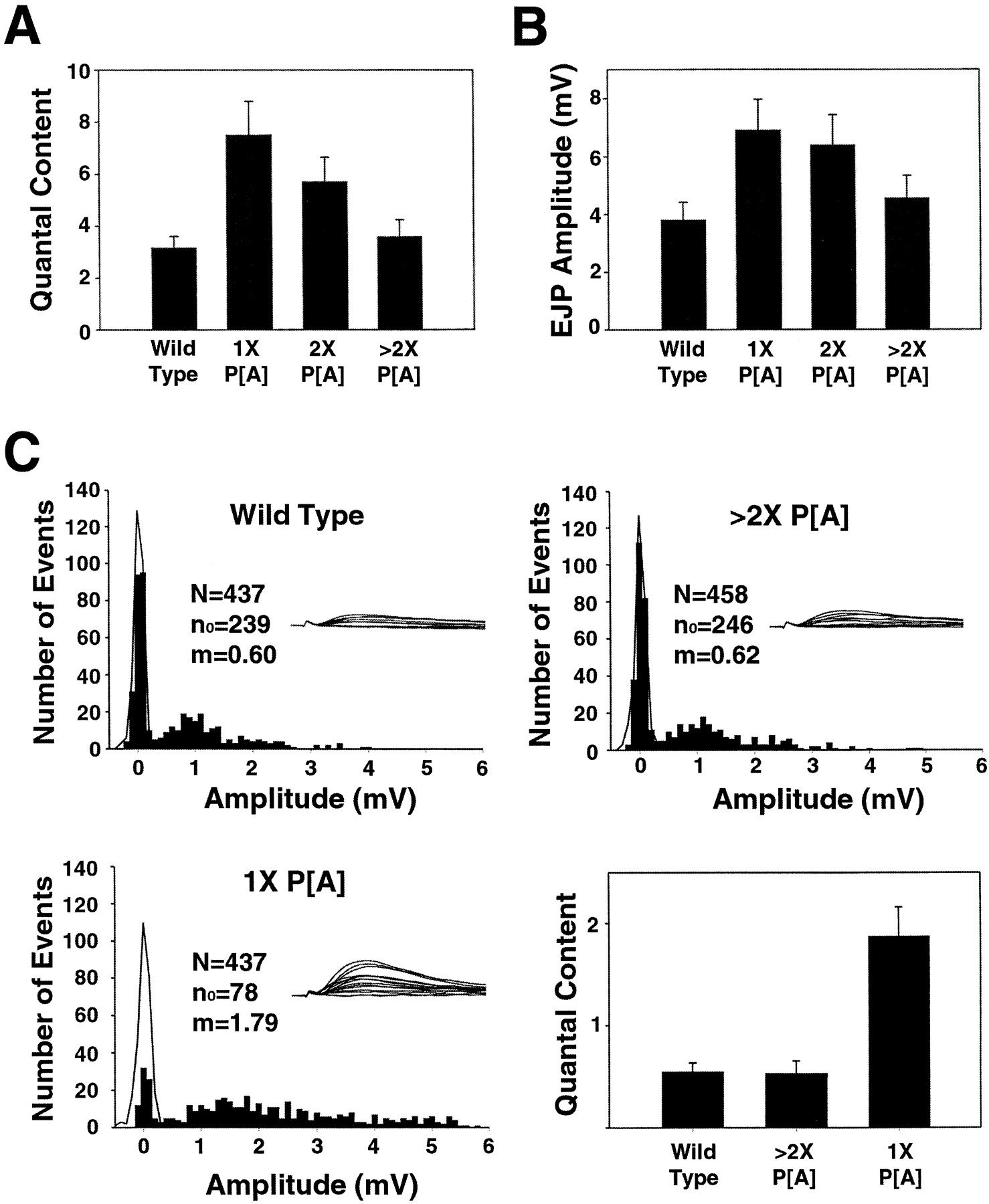
Quantal content overcompensates at synapses expressing low levels of DGluRIIA. The mean ± SEM of the calculated quantal content (A) and EJP amplitude (B) is shown for wild-type larvae (Canton S, n = 12) and for rescue of the glutamate receptor double-mutant DGluRIIA&BSP22by various doses of DGluRIIA: 1× P[A], a single copy of the DGluRIIA genomic rescue transgene (n = 12); 2× P[A], two copies of theDGluRIIA genomic rescue transgene (n= 12); and >2× P[A], UASDGluRIIA driven by 24BGal4 (n = 11). The mean quantal content was calculated for each cell by dividing the average suprathreshold EJP amplitude (n > 75) by the average amplitude of the spontaneous miniature events (n > 60). Recordings were made from muscle 6, segment A3 in saline containing 0.3 mm calcium. The mean ± SEM of the resting potentials: wild type, 70.0 ± 0.9 mV; 1×, 70.8 ± 1.5 mV; 2×, 69.7 ± 1.2 mV; >2×, 71.3 ± 1.0 mV. In C, frequency histograms of evoked release recorded in 0.18 mmcalcium from muscle 6, segment A3 from representative wild-type, >2× P[A], and 1× P[A] larvae. Fifteen consecutive traces from each genotype are shown above the histogram and demonstrate that, after the stimulus artifact, release events are separated from failures. Amplitudes of evoked events are plotted as filled bars, and the amplitudes of noise measurements are shown as theblack line. Evoked events within the distribution of the noise measurement are considered failures. N, Number of trials; n0 , number of failures;m, quantal content calculated by the method of failures (ln[N/n0]). The bar graph shows the mean ± SEM of the quantal content calculated by the method of failures for wild type (n = 10), >2× P[A] (n = 10), and 1× P[A] (n = 11).
Whereas the inverse relationship between the gene dosage ofDGluRIIA and quantal content was expected, the magnitude of the change in quantal content was a surprise. Although the null mutant rescued with a single genomic DGluRIIA transgene does have a slightly smaller quantal size than wild type (0.97 ± 0.06 vs 1.19 ± 0.07 mV), the increase in quantal content more than compensates for this postsynaptic deficit. As a result, the postsynaptic response to nerve stimulation is significantly increased (182%; p < 0.05) (Fig. 7B). As the gene dosage of DGluRIIA (and hence the response to a single vesicle) is increased, the response to nerve stimulation decreases because quantal content is no longer upregulated. This result, in addition to the increase seen in the EJP amplitude in theH94-DGluRIIA rescue larvae described above, suggests that the mechanism monitoring postsynaptic activity and regulating presynaptic transmitter release is not directly sensitive to depolarization of the muscle.
A second, independent estimate of quantal content can be derived from the method of failures. In low-calcium saline (0.18 mmcalcium), both wild type and the double mutant rescued by overexpression of the DGluRIIA cDNA show a large proportion of failures. In the double mutant rescued with a single genomic copy ofDGluRIIA, there are many fewer failures (Fig.7C). Quantal content was estimated as the natural log of the ratio of trials of nerve stimulation to the number of failures of the nerve to release transmitter. No difference in quantal content was observed between wild-type and overexpressing DGluRIIAsynapses. However, there was a large and significant increase in quantal content in the mutants expressing just a single genomic copy ofDGluRIIA (290%; p < 0.01) (Fig.7C, histogram). Therefore, two methods confirm that when low levels of DGluRIIA are expressed postsynaptically, there is a large increase in presynaptic transmitter release that overcompensates for the decrease in postsynaptic sensitivity to transmitter.
Quantal content compensation is sensitive to the activity of the postsynaptic receptor
Activation of ionotropic glutamate receptors leads to the generation of two types of signals. The postsynaptic cell is depolarized by the influx of cations through the open channel, and second messenger systems can be activated through either the influx of calcium or the interaction of the receptor with other signaling molecules (Dong et al., 1997; Wang et al., 1997; Sprengel et al., 1998). The overcompensation of quantal content seen in the single genomic DGluRIIA rescue suggests that depolarization is not the determinant being sensed in the postsynaptic cell. In fact, this result could suggest that the retrograde signal is not even sensitive to the activity of the channel but instead is measuring the amount of channel present. To distinguish between the activity and amount of postsynaptic receptor, we generated a dominant negative mutant ofDGluRIIA. Using site-directed mutagenesis, we changed a single residue in the channel pore M614 to an R. The analogous mutation in homologous vertebrate channels is thought to coassemble with wild-type receptors and produce nonfunctional channels (Dingledine et al., 1992).
Transgenic flies were generated carrying the M/R mutant cloned downstream of the UAS promoter. Expression of two copies of this transgene in a wild-type background driven by the strong mesodermal promoter 24B Gal4 is lethal. Driving expression of a single copy of the mutant with 24B Gal4 produces viable adults with no obvious behavioral abnormalities. Staining of the larval neuromuscular junction shows that this mutant receptor does localize to the synapse. As with overexpression of the wild-type receptor, however, much of the transgenic receptor is present extrasynaptically (data not shown). Recordings of spontaneous excitatory junctional potentials reveal that expression of this mutant receptor leads to a dramatic decrease in quantal size (1.01 ± 0.05 vs 0.33 ± 0.02 mV;p < 0.001) (Fig. 8). Hence, this pore mutant acts as a dominant negative receptor in vivo. Analysis of evoked synaptic potentials revealed no significant change, indicative of a large increase in quantal content in the mutant (3.5 ± 0.6 vs 12 ± 2.0; p < 0.01) (Fig. 8B). These data do not support the model that a low channel density is the signal controlling the retrograde regulation of presynaptic transmitter release. Normal levels of the endogenous DGluRIIA and DGluRIIB receptors are expressed in addition to the transgenic expression of aDGluRIIA pore mutant, and yet quantal content is upregulated. Although we cannot rule out the possibility that the mutant channel could disrupt localization of the endogenous receptors to the synapse, we favor the model that it acts as a dominant negative by disrupting the pore of the channel. Therefore, these data imply that the activity of the channel and ion flux through the pore are the initiating events for the measurement of postsynaptic activity and the regulation of presynaptic function.

Overexpression of a dominant negative pore mutant of DGluRIIA leads to a decrease in quantal size and a compensatory increase in quantal content. A, Representative traces of spontaneous and trace averages of evoked transmitter release recorded from muscle 6, segment A3 of wild-type (Canton S) and dominant negative (UASDGluRIIA M614R driven by 24BGal4) third instar larvae. Calibration: spontaneous, 200 msec, 2 mV; evoked, 24 msec, 1 mV. B, The mean ± SEM for the mEJP amplitude, EJP amplitude, and quantal content is shown for wild-type (Canton S; n = 13) and dominant negative (UASDGluRIIA M614R driven by 24BGal4;n = 13) third instar larvae recorded in 0.3 mm calcium from muscle 6, segment A3. The mean quantal content was calculated for each cell by dividing the average suprathreshold EJP amplitude (n > 75) by the average amplitude of the spontaneous miniature events (n > 60). Expression of the dominant negative receptor does not change the kinetics of depolarization (EJP width at the half-maximal amplitude, 32.7 ± 2.5 vs 34.7 ± 3.0 sec;p = 0.61). Mean resting potentials ± SEM: wild type, −72.4 ± 1.4 mV; dominant negative, −71.2 ± 1.6 mV.
DISCUSSION
In this study, we have investigated postsynaptic mechanisms of synaptic plasticity that function at the Drosophilaglutamatergic neuromuscular junction. We present a genetic analysis of two ionotropic glutamate receptors, DGluRIIA andDGluRIIB, that are expressed at this synapse. We show that deletion of both receptors results in embryonic lethality, and that this lethality is rescued by expression of either receptor. Although these receptors are redundant for viability, they have important physiological differences. The receptor subunit composition regulates quantal size, Argiotoxin 636 sensitivity, and receptor desensitization kinetics. We also examine the relationship of receptor expression to the generation of the muscle-to-motoneuron retrograde signal that regulates presynaptic function at this synapse.
Two glutamate receptors at the neuromuscular junction
The genes for DGluRIIA and DGluRIIB are adjacent in the genome, share a similar intron–exon structure, and encode receptors that are more similar to each other than to any other known glutamate receptor. We have demonstrated previously that animals lacking expression of DGluRIIA are viable, suggesting that these genes may be redundant (Petersen et al., 1997). Here, we present double-mutant analysis that demonstrates that the receptor complex is essential for viability. These mutants are unable to hatch from the egg case and, when mechanically removed from the chorion, exhibit no coordinated peristaltic waves, suggesting that these receptors are required for normal synaptic transmission (Broadie and Bate, 1993). However, these mutants do have some coordinated movement of the head, so residual synaptic transmission occurs in at least a subset of muscles. This may be attributable to a second transmitter system or to a third, unidentified glutamate receptor.
In a series of rescue experiments, we have demonstrated that these receptors are functionally redundant for viability and that their essential function is in the somatic musculature. Because neither receptor is required, each must be capable of assembling as a homomultimer and localizing to the NMJ. Alternatively, each may coassemble with a third, unidentified subunit.
Receptor subunit composition and quantal size
The presence of two receptors with different physiological properties provides the cell a simple mechanism for regulating the postsynaptic response to transmitter. To investigate whether this type of plasticity could function at the Drosophila NMJ, we have used our mutants and rescue transgenes to systematically investigate the relationship of receptor gene dosage and composition to synaptic function.
We have demonstrated previously that quantal size is reduced in the absence of DGluRIIA and that quantal size is increased whenDGluRIIA is overexpressed (Petersen et al., 1997). These findings suggest that receptor density is a primary determinant of postsynaptic responsiveness. However, they are equally consistent with a model in which the relative level of DGluRIIA regulates quantal size, with a higher proportion of DGluRIIA favoring a larger postsynaptic response. The data presented here favor this second model. Regardless of the level of expression, synapses lackingDGluRIIB have a large quantal size, and synapses lackingDGluRIIA have a small quantal size. In fact, overexpression of the DGluRIIB subunit at a wild-type synapse leads to a dose-dependent decrease in quantal size. In this case, the receptor density should be increasing, but the quantal size is decreasing. This is most easily explained if the primary determinant of quantal size at this synapse is the relative abundance of each receptor subtype.
Although subunit composition is the primary factor controlling postsynaptic responsiveness, our data does suggest that receptor density may also regulate quantal size. In the absence of theDGluRIIB subunit, increasing the gene dosage ofDGluRIIA increases the quantal size. However, we cannot exclude the alternate explanation that the subunit composition is changing between DGluRIIA and an unidentified third receptor.
How might the cell exploit the differences in receptor function to regulate synaptic strength? First, the two receptors could be differentially expressed. During embryonic development,DGluRIIB is initially expressed at a high level and then declines, whereas DGluRIIA expression slowly rises throughout embryogenesis (Petersen et al., 1997). Such a mechanism is used at the vertebrate NMJ in the switch from a fetal to adult acetylcholine receptor subunit. Second, the two receptors could be differentially regulated by second messengers. Davis et al. (1998) have demonstrated that activation of PKA decreases the quantal size at theDrosophila NMJ and that this modulation requires the presence of DGluRIIA. Similar subunit-specific modulation has been seen for numerous vertebrate transmitter receptors (for review, see Smart, 1997). Finally, the localization of receptor subunits could be regulated. In Drosophila, there is no evidence for differential localization of these two receptors, although the regulated membrane insertion of homologous vertebrate AMPA receptors has been proposed as a mechanism for long-term potentiation in the hippocampus (Isaac et al., 1995; Liao et al., 1995).
Does the cell use these postsynaptic mechanisms to regulate synaptic strength? When a Drosophila muscle is hypoinnervated, it compensates with an increase in quantal size (Davis and Goodman, 1998a). We suggest that this increase in postsynaptic sensitivity may reflect an increase in the proportion of DGluRIIA at the synapse or a decrease in the PKA-dependent modulation ofDGluRIIA.
Single-channel properties of DGluRIIA and DGluRIIB
As a probe for molecular differences between the two receptors, we studied block by the use-dependent spider venom Argiotoxin 636. This toxin is likely to block the pore of the channel (Jackson and Usherwood, 1988). We found that channels containing theDGluRIIA subunit are much more sensitive to toxin than channels composed of DGluRIIB. Similar results have been found with vertebrate glutamate receptors in which block requires the absence of the GluRB subunit (Brackley et al., 1993; Herlitze et al., 1993).
To investigate the biophysical basis of the differences between the receptors, we have initiated a single-channel analysis. We find no evidence for differences in the single-channel conductance of the two receptors. However, we do find a major difference in the time course of desensitization between the two subunits. Similar differences have been observed for vertebrate glutamate receptors in which subunit composition, flip–flop splice variants, and the R/G edited site all regulate desensitization kinetics (Lomeli et al., 1994; Mosbacher et al., 1994).
How might this difference in channel properties affect synaptic function at the NMJ? Channel desensitization mediates short-term depression during rapid firing at this synapse (M. Heckmann, unpublished observation). Therefore, differences in receptor subunit composition could regulate this form of postsynaptic plasticity. The faster decay kinetics of mEJPs at synapses lacking DGluRIIAlikely reflects the faster desensitization kinetics ofDGluRIIB channels. To demonstrate this point, however, will require an analysis of the time course of synaptic currents at this synapse. In vertebrates, desensitization of AMPA receptors may affect the kinetics of synaptic events at some, but not all, glutamatergic synapses (for review, see Jonas and Spruston, 1994; Westbrook, 1994). Finally, what leads to the decrease in quantal size seen in the absence of DGluRIIA? Previous work has demonstrated thatDrosophila glutamate receptors can desensitize before opening (Heckmann and Dudel, 1997), and modeling of their kinetics suggests that an increase in the rate of desensitization would lead to a decrease in the synaptic current. In addition, because synaptic currents are much shorter than the membrane time constant of the postsynaptic muscle (Jan and Jan, 1976), the more rapid currents mediated by DGluRIIB will lead to a smaller synaptic depolarization.
Receptor expression regulates the retrograde signal that modulates presynaptic function
We have demonstrated previously that, in the absence ofDGluRIIA, an increase in quantal content compensates for the decrease in quantal size so that postsynaptic excitation remains normal after nerve stimulation (Petersen et al., 1997). These data lead to the simple model of a homeostatic mechanism in which a muscle-to-motoneuron signal regulates presynaptic release to ensure appropriate depolarization of the muscle. Similar compensation may occur at the vertebrate and crayfish NMJ and at central excitatory and inhibitory synapses (Cull-Candy et al., 1980; Lnenicka and Mellon, 1983; Plomp et al., 1992; Sandrock et al., 1997; Nusser et al., 1998; Turrigiano et al., 1998). Such a mechanism could function during development to match the release capacity of the nerve to the ever growing requirements of the muscle (Davis and Goodman, 1998b; Landmesser, 1998). However, increases in quantal size do not lead to a decrease in quantal content, so this compensatory mechanism must not be strictly regulated by depolarization (Petersen et al., 1997; Davis et al., 1998).
In this study, we find that, at synapses lacking DGluRIIB, this compensatory mechanism is also functional. Over a wide range ofDGluRIIA expression, we see an inverse correlation between the quantal size and quantal content. To our surprise, however, we find that, at synapses expressing low levels of DGluRIIA, the increase in quantal content overcompensates, leading to a significant increase in postsynaptic excitation. We demonstrate that this overcompensation reflects a presynaptic change, because the proportion of failures at these synapses is much reduced.
This increase in quantal content, despite the increase in postsynaptic excitation, supports the previous finding that postsynaptic depolarization is not the primary determinant being sensed by the muscle. However, this leaves open the question of why quantal content overcompensates at synapses expressing low levels ofDGluRIIA. At synapses expressing no DGluRIIB and either low or high levels of DGluRIIA, the postsynaptic sensitivity to transmitter is similar. However, there could be a difference in the ion permeability of these channels if there is a third receptor subunit. For example, channels composed primarily of the third subunit (in the case of low DGluRIIA expression) could have lower calcium permeability than channels containing primarilyDGluRIIA. In this case, synaptic calcium influx could be sensed by the muscle. Alternatively, the receptor may synaptically localize signaling molecules that are required for the compensation. Evidence from other systems suggests that glutamate receptors can interact with such molecules and that this may be important for some forms of plasticity (Dong et al., 1997; Wang et al., 1997; Sprengel et al., 1998). With low levels of receptor, too few signaling molecules may be present at the synapse, resulting in the inappropriate activation of the retrograde signal.
We have tested this second model by overexpressing a dominant negative pore mutant at an otherwise wild-type synapse and find that the decrease in quantal size is compensated for by an increase in quantal content. This argues that the amount of receptor is not the primary determinant sensed by the muscle, because receptor is abundantly expressed. Instead, these data suggest that something related to ion flux through the channel is sensed by the muscle. Of course, these two models are not mutually exclusive; the receptor may localize molecules to the synapse that are then activated by channel activity.
Footnotes
A.D. was supported by a Helen Hay Whiney Postdoctoral Fellowship and a Burroughs Wellcome Career Award. S.A.P. is a predoctoral fellow, and C.S.G. is an Investigator with the Howard Hughes Medical Institute. M.H. was supported by Deutsche Forschungsgemeinschaft Grant SFB 391/A4.
Drs. DiAntonio and Petersen contributed equally to this work.
Correspondence should be addressed to Corey S. Goodman, Howard Hughes Medical Institute, Department of Molecular and Cell Biology, Life Sciences Addition Room 519, University of California at Berkeley, Berkeley, CA 94720.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}